
Multi-Omics Analysis Reveals the Potential Effects of Maternal Dietary Restriction on Fetal Muscle Growth and Development


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
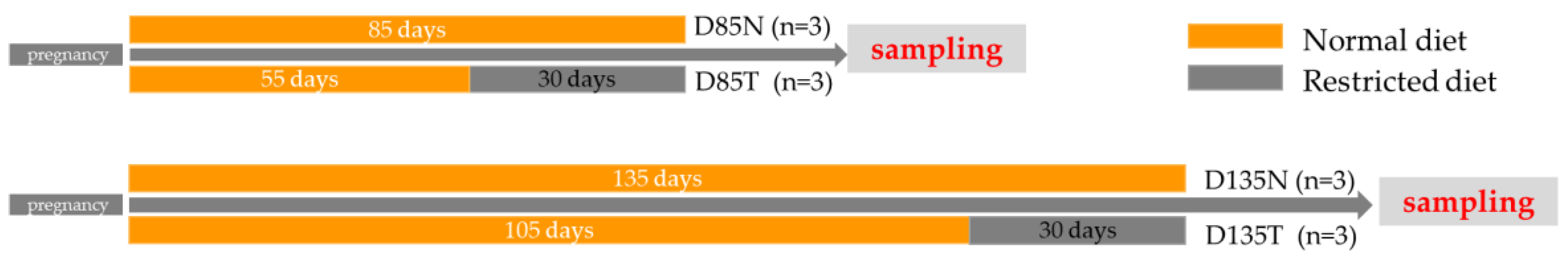
2.1. Animals, Diets, and Sample Collection
2.2. Phenotype Trait Analysis
2.3. mRNA Sequencing Analysis
2.4. Proteome Quantitative Analysis
2.5. Statistical Analysis
2.6. qRT-PCR and PRM of mRNAs and Proteins in Muscle Tissues
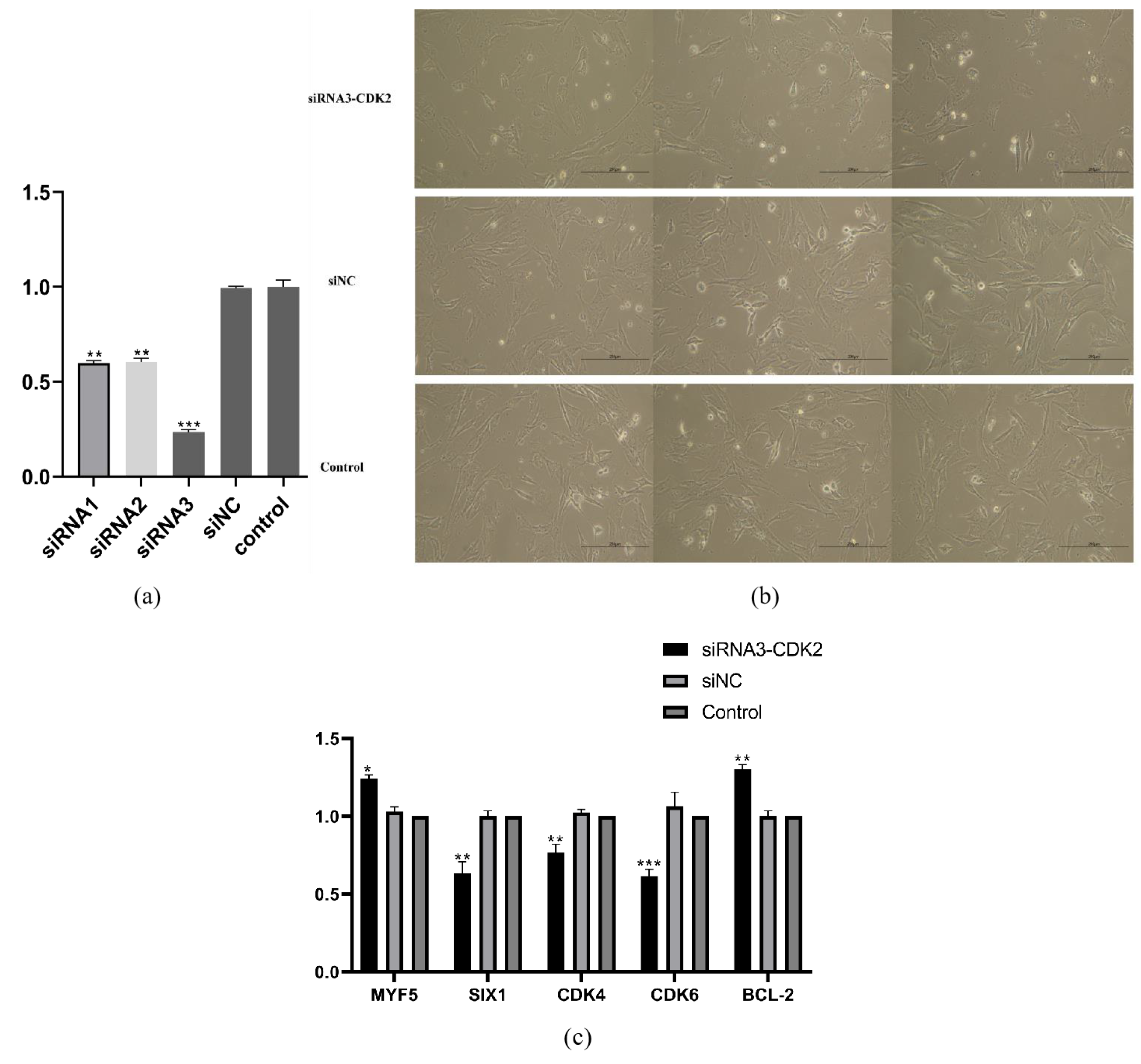
2.7. Knock-Down CDK2 Gene Expression in the Sheep Primary Embryonic Myoblasts
2.8. Gene and Phenotypic Analysis of Sheep Primary Embryonic Myoblasts Cultured under Different Nutrient Conditions
3. Results
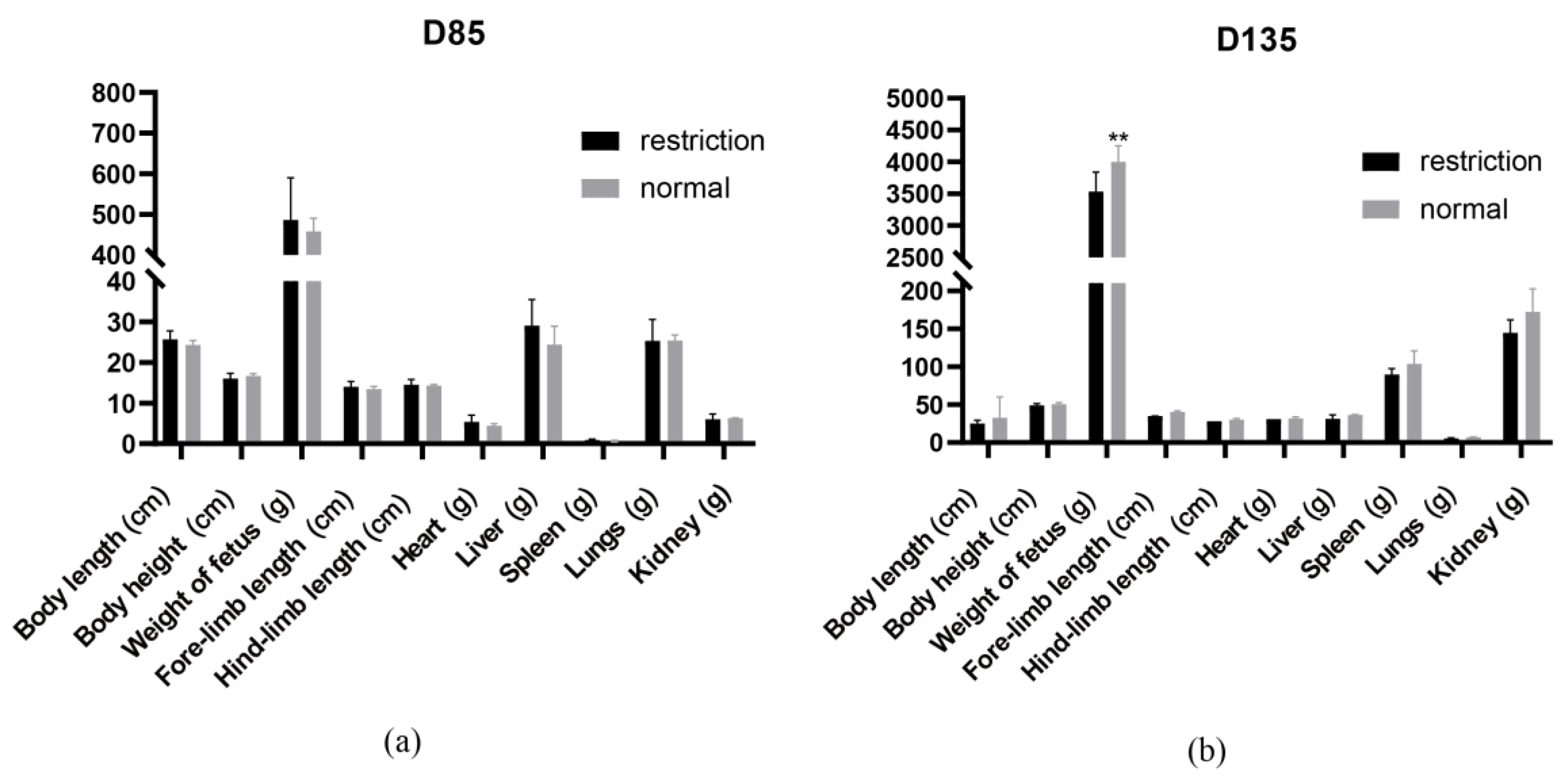
3.1. Maternal Dietary Restriction Nutrition Causes the Fetal Body Weight to Decrease Significantly
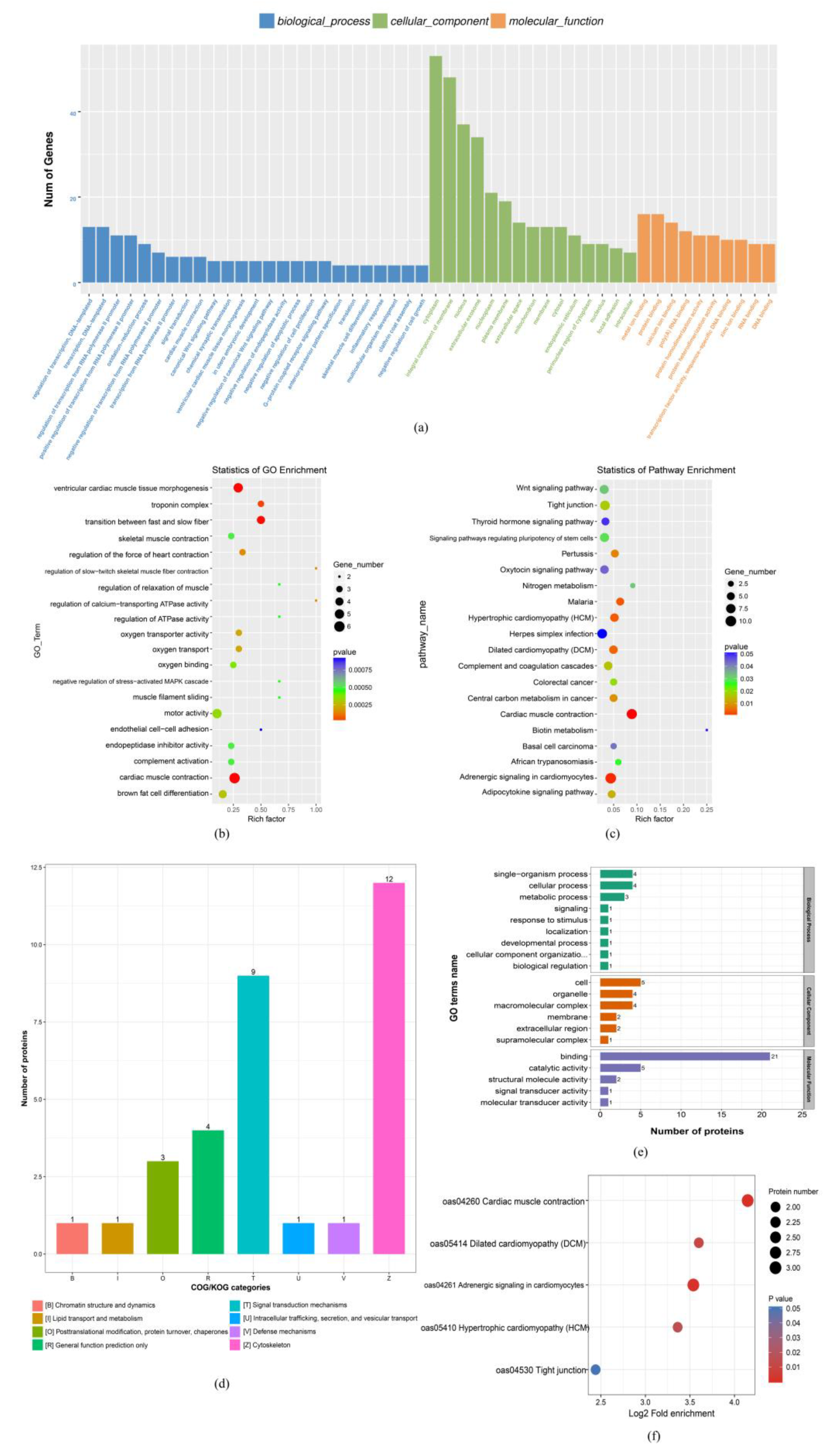
3.2. mRNA and Protein Expression Profiles
3.3. Integrating the mRNA and Protein Expression Profile Analyses
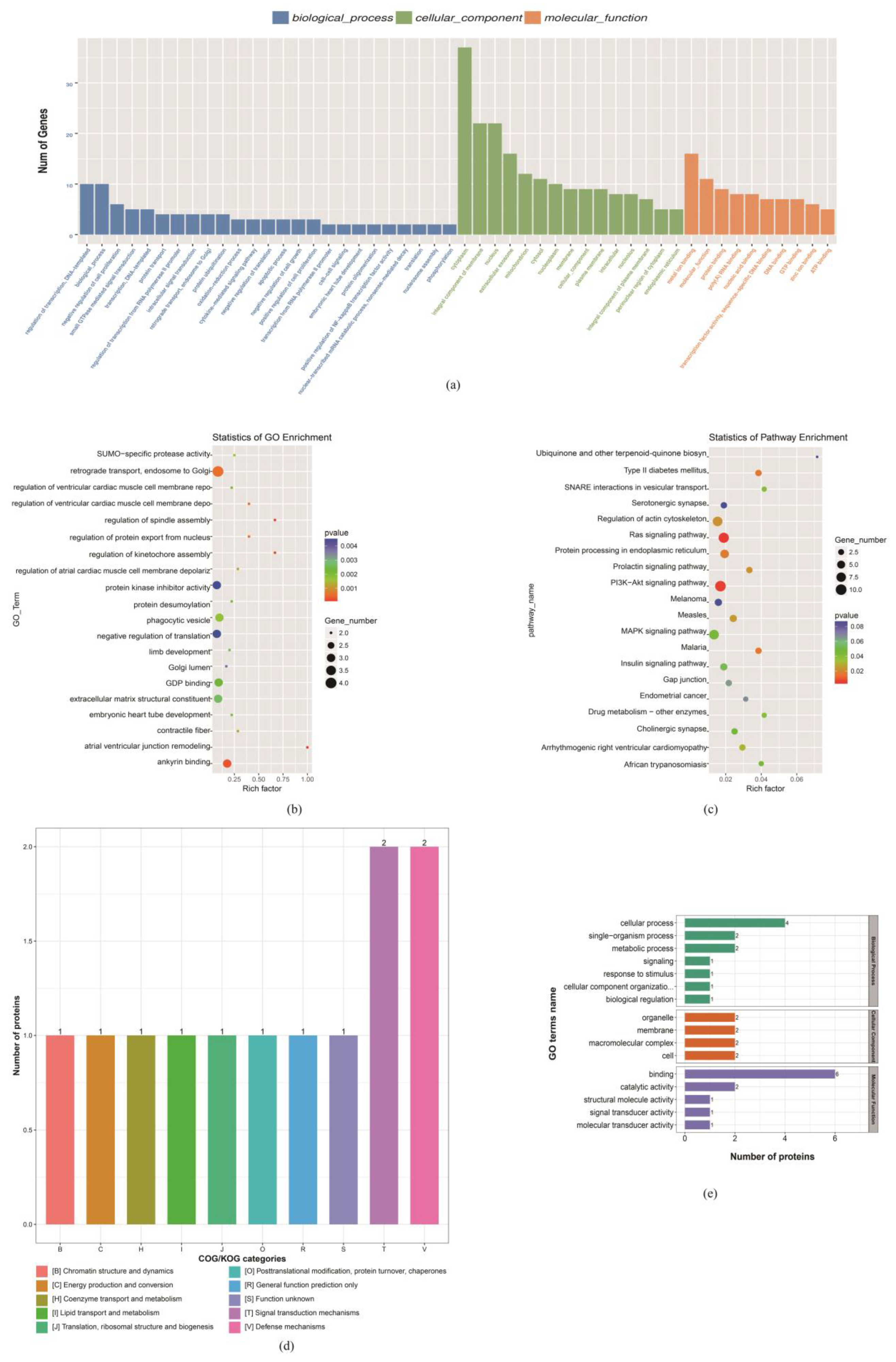
3.4. At GD 85, Maternal Dietary Restriction Alters the Cell Cycle of Sheep Embryonic Myoblasts
3.4.1. At the mRNA and Protein Levels, the Cell Cycle and Apoptosis-Related Gene Expression Changes

3.4.2. CDK2 May Play a Key Role in Myoblast Cell Cycle Changes in Response to Maternal Dietary Deficiency
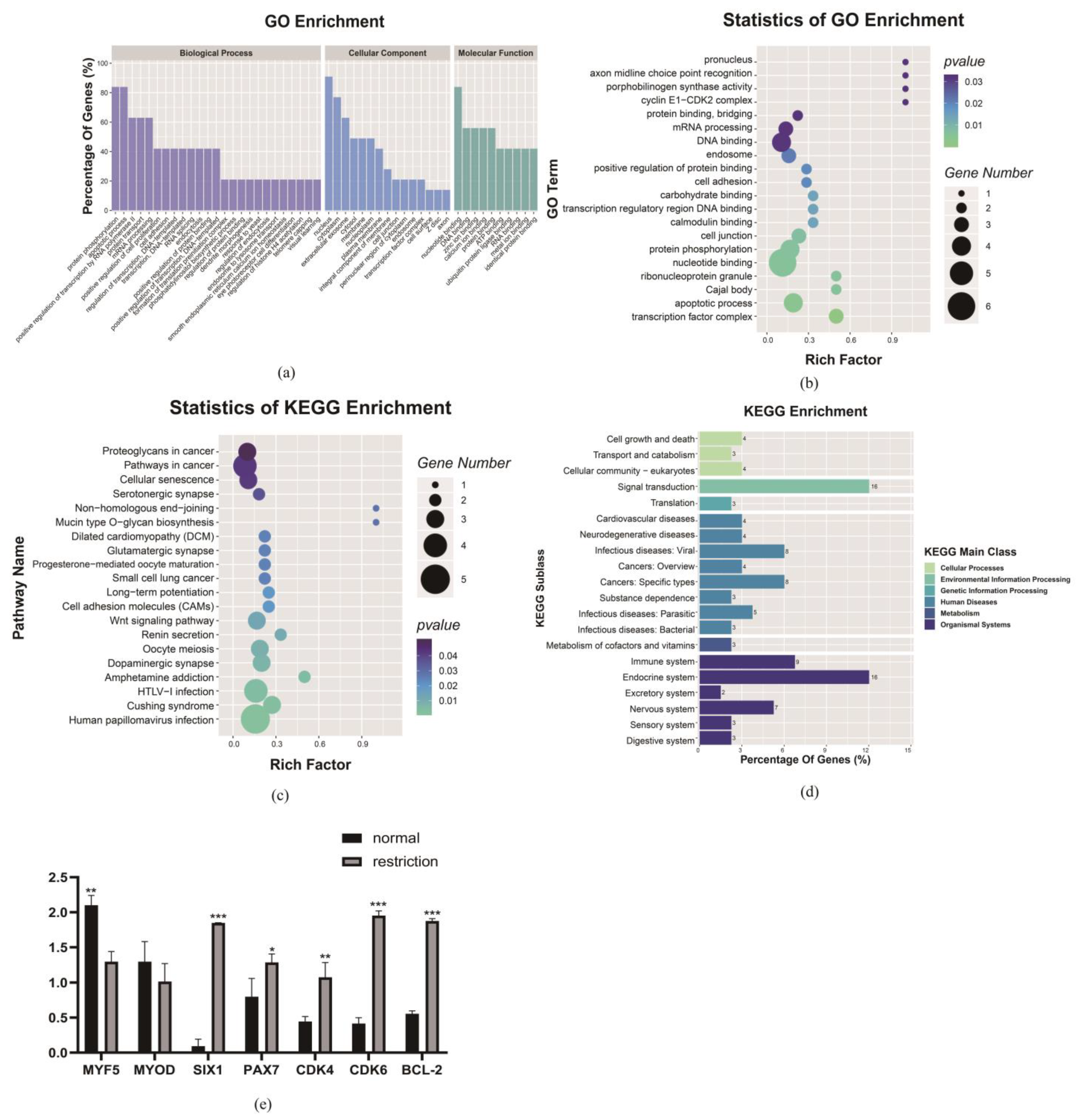
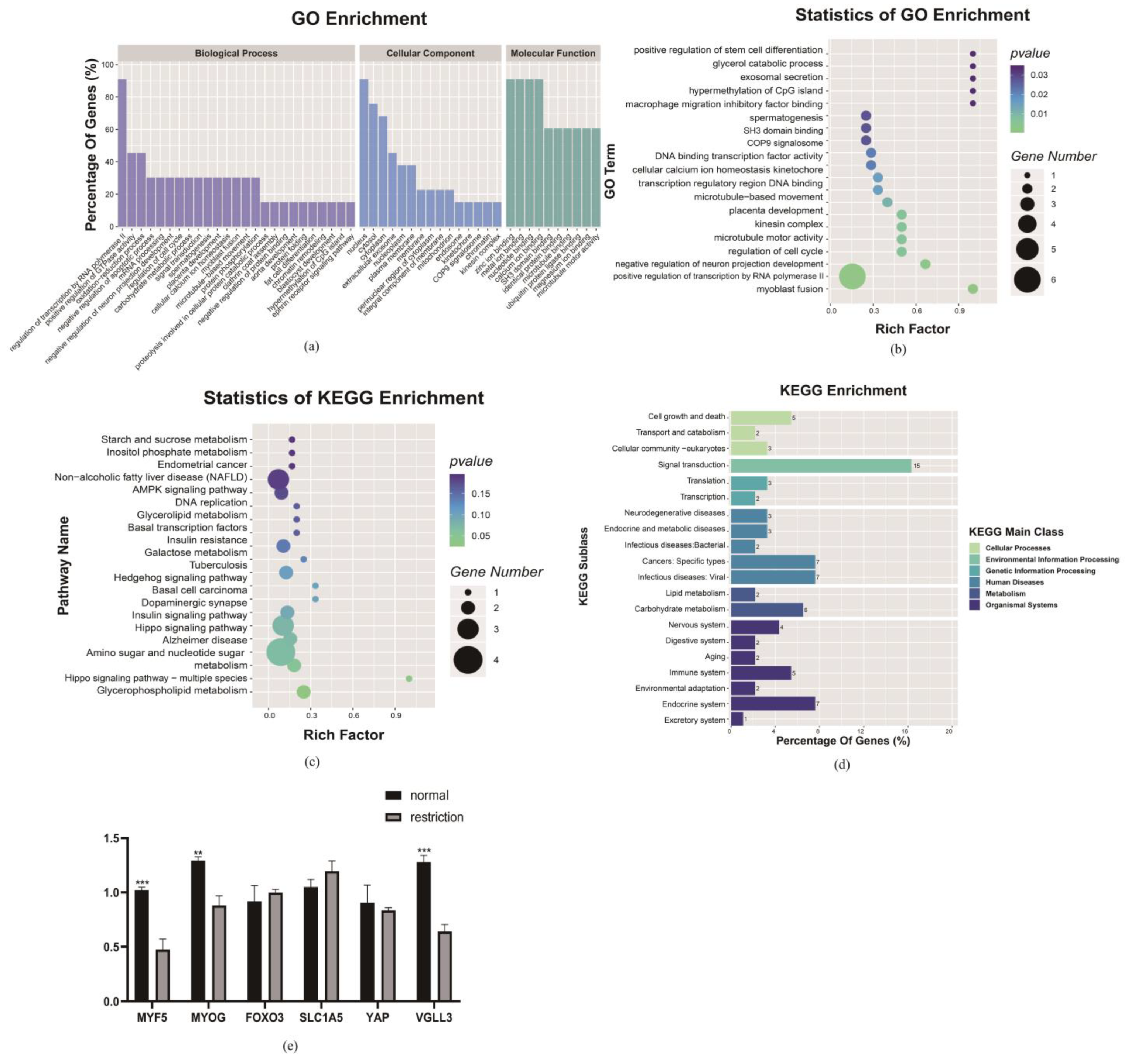
3.5. Maternal Dietary Restriction Prevents Muscle Fiber Differentiation and Maturation at GD 135
3.5.1. At the mRNA and Protein Levels, Genes Related to Muscle Fiber Protein Synthesis, Metabolism and Differentiation Change

3.5.2. Under the Maternal Dietary Restriction, TEAD1 Regulating the VGLL3 Gene May Be a Potential Reason for Affecting the Myofiber Differentiation and Maturation
3.6. mRNA Sequencing and Quantitative Protein Validation
3.7. CDK2 Knockdown Affects the Cell Cycle and Proliferation of Sheep Primary Embryonic Myoblasts
3.8. Low Nutrition Effect on the Sheep Primary Embryonic Myoblasts


4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, G.; Bazer, F.W.; Cudd, T.A.; Meininger, C.J.; Spencer, T.E. Maternal Nutrition and Fetal Development. J. Nutr. 2004, 134, 2169–2172. [Google Scholar] [CrossRef] [PubMed]
- Ugwuoke, W.I.; Igwebuike, U.M.; Igbokwe, C.O. The effects of maternal dietary protein restriction during gestation in rats on postnatal growth of the body and internal organs of the offspring. Anim. Res. Int. 2020, 17, 3596–3602. [Google Scholar]
- Zeisel, S.H. Nutritional importance of choline for brain development. J. Am. Coll. Nutr. 2004, 23, 621s–626s. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.M.; Kirke, P.N.; Weir, D.G. The role of nutrition in neural tube defects. Annu. Rev. Nutr. 1990, 10, 277–295. [Google Scholar] [CrossRef] [PubMed]
- England-Mason, G.; Dewey, D. Strengthening research investigating maternal nutrition and children’s neurodevelopment: How can we do it better? Where do we go from here? Am. J. Clin. Nutr. 2021, 114, 1272–1274. [Google Scholar] [CrossRef] [PubMed]
- Lima, H.K. The Role of Maternal Nutrition on Fetal Development and Programming. Master of Science (MS), North Carolina State University, Raleigh, North Carolina USA, August 2014. Available online: http://www.lib.ncsu.edu/resolver/1840.16/9913 (accessed on 29 August 2014).
- Elolimy, A.; Vailati-Riboni, M.; Liang, Y.; Loor, J.J. Cellular Mechanisms and Epigenetic Changes: Role of Nutrition in Livestock. Vet. Clin. N. Am. Food Anim. Pract. 2019, 35, 249–263. [Google Scholar] [CrossRef]
- Yang, W.; Han, N.; Jiao, M.; Chang, X.; Liu, J.; Zhou, Q.; Wang, H.J. Maternal diet quality during pregnancy and its influence on low birth weight and small for gestational age: A birth cohort in Beijing, China. Br. J. Nutr. 2022, 7, 1–34. [Google Scholar] [CrossRef]
- King, J.C. A Summary of Pathways or Mechanisms Linking Preconception Maternal Nutrition with Birth Outcomes. J. Nutr. 2016, 146, 1437s–1444s. [Google Scholar] [CrossRef]
- Zamora, A.N.; Peterson, K.E.; Téllez-Rojo, M.M.; Cantoral, A.; Song, P.X.K.; Mercado-García, A.; Solano-González, M.; Fossee, E.; Jansen, E.C. Third-Trimester Maternal Dietary Patterns Are Associated with Sleep Health among Adolescent Offspring in a Mexico City Cohort. J. Nutr. 2022, 152, 1487–1495. [Google Scholar] [CrossRef]
- Okubo, H.; Miyake, Y.; Sasaki, S.; Tanaka, K.; Murakami, K.; Hirota, Y.; Kanzaki, H.; Kitada, M.; Horikoshi, Y.; Ishiko, O.; et al. Maternal dietary patterns in pregnancy and fetal growth in Japan: The Osaka Maternal and Child Health Study. Br. J. Nutr. 2012, 107, 1526–1533. [Google Scholar] [CrossRef]
- Marangoni, F.; Cetin, I.; Verduci, E.; Canzone, G.; Giovannini, M.; Scollo, P.; Corsello, G.; Poli, A. Maternal Diet and Nutrient Requirements in Pregnancy and Breastfeeding. An Italian Consensus Document. Nutrients 2016, 8, 629. [Google Scholar] [CrossRef] [PubMed]
- Partridge, E.A.; Davey, M.G.; Hornick, M.A.; McGovern, P.E.; Mejaddam, A.Y.; Vrecenak, J.D.; Mesas-Burgos, C.; Olive, A.; Caskey, R.C.; Weiland, T.R.; et al. An extra-uterine system to physiologically support the extreme premature lamb. Nat. Commun. 2017, 8, 15112. [Google Scholar] [CrossRef] [PubMed]
- Aguilera-Castrejon, A.; Oldak, B.; Shani, T.; Ghanem, N.; Itzkovich, C.; Slomovich, S.; Tarazi, S.; Bayerl, J.; Chugaeva, V.; Ayyash, M.; et al. Ex utero mouse embryogenesis from pre-gastrulation to late organogenesis. Nature 2021, 593, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Heidari Kani, M.; Chan, E.C.; Young, R.C.; Butler, T.; Smith, R.; Paul, J.W. 3D Cell Culturing and Possibilities for Myometrial Tissue Engineering. Ann. Biomed. Eng. 2017, 45, 1746–1757. [Google Scholar] [CrossRef] [PubMed]
- Odle, J.; Jacobi, S.K.; Boyd, R.D.; Bauman, D.E.; Anthony, R.V.; Bazer, F.W.; Lock, A.L.; Serazin, A.C. The Potential Impact of Animal Science Research on Global Maternal and Child Nutrition and Health: A Landscape Review. Adv. Nutr. 2017, 8, 362–381. [Google Scholar] [CrossRef]
- Beede, K.A.; Limesand, S.W.; Petersen, J.L.; Yates, D.T. Real supermodels wear wool: Summarizing the impact of the pregnant sheep as an animal model for adaptive fetal programming. Anim. Front. 2019, 9, 34–43. [Google Scholar] [CrossRef]
- Barry, J.S.; Anthony, R.V. The pregnant sheep as a model for human pregnancy. Theriogenology 2008, 69, 55–67. [Google Scholar] [CrossRef]
- Wang, X.; Shi, T.; Zhao, Z.; Hou, H.; Zhang, L. Proteomic analyses of sheep (Ovis aries) embryonic skeletal muscle. Sci. Rep. 2020, 10, 1750. [Google Scholar] [CrossRef]
- Gauvin, M.C.; Pillai, S.M.; Reed, S.A.; Stevens, J.R.; Hoffman, M.L.; Jones, A.K.; Zinn, S.A.; Govoni, K.E. Poor maternal nutrition during gestation in sheep alters prenatal muscle growth and development in offspring. J. Anim. Sci. 2020, 98, skz388. [Google Scholar] [CrossRef]
- Li, C.; Jenkins, S.; Mattern, V.; Comuzzie, A.G.; Cox, L.A.; Huber, H.F.; Nathanielsz, P.W. Effect of moderate, 30 percent global maternal nutrient reduction on fetal and postnatal baboon phenotype. J. Med. Primatol. 2017, 46, 293–303. [Google Scholar] [CrossRef]
- Dwyer, C.M.; Stickland, N.C.; Fletcher, J.M. The influence of maternal nutrition on muscle fiber number development in the porcine fetus and on subsequent postnatal growth. J. Anim. Sci. 1994, 72, 911–917. [Google Scholar] [CrossRef]
- Hoffman, M.L.; Peck, K.N.; Forella, M.E.; Fox, A.R.; Govoni, K.E.; Zinn, S.A. The effects of poor maternal nutrition during gestation on postnatal growth and development of lambs. J. Anim. Sci. 2016, 94, 789–799. [Google Scholar] [CrossRef]
- Brameld, J.M.; Mostyn, A.; Dandrea, J.; Stephenson, T.J.; Dawson, J.M.; Buttery, P.J.; Symonds, M.E. Maternal nutrition alters the expression of insulin-like growth factors in fetal sheep liver and skeletal muscle. J. Endocrinol. 2000, 167, 429–437. [Google Scholar] [CrossRef]
- Sandoval, C.; Lambo, C.A.; Beason, K.; Dunlap, K.A.; Satterfield, M.C. Effect of maternal nutrient restriction on skeletal muscle mass and associated molecular pathways in SGA and Non-SGA sheep fetuses. Domest. Anim. Endocrinol. 2020, 72, 106443. [Google Scholar] [CrossRef]
- Raja, J.S.; Hoffman, M.L.; Govoni, K.E.; Zinn, S.A.; Reed, S.A. Restricted maternal nutrition alters myogenic regulatory factor expression in satellite cells of ovine offspring. Animal 2016, 10, 1200–1203. [Google Scholar] [CrossRef]
- Odhiambo, J.F.; Pankey, C.L.; Ghnenis, A.B.; Ford, S.P. A Review of Maternal Nutrition during Pregnancy and Impact on the Offspring through Development: Evidence from Animal Models of Over- and Undernutrition. Int. J. Environ. Res. Public Health 2020, 17, 6926. [Google Scholar] [CrossRef] [PubMed]
- Campodonico-Burnett, W.; Hetrick, B.; Wesolowski, S.R.; Schenk, S.; Takahashi, D.L.; Dean, T.A.; Sullivan, E.L.; Kievit, P.; Gannon, M.; Aagaard, K.; et al. Maternal Obesity and Western-Style Diet Impair Fetal and Juvenile Offspring Skeletal Muscle Insulin-Stimulated Glucose Transport in Nonhuman Primates. Diabetes 2020, 69, 1389–1400. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Schlabritz-Loutsevitch, N.E.; Hubbard, G.B.; Han, V.; Nygard, K.; Cox, L.A.; McDonald, T.J.; Nathanielsz, P.W. Effects of maternal global nutrient restriction on fetal baboon hepatic insulin-like growth factor system genes and gene products. Endocrinology 2009, 150, 4634–4642. [Google Scholar] [CrossRef]
- Hoffman, M.L.; Rokosa, M.A.; Zinn, S.A.; Hoagland, T.A.; Govoni, K.E. Poor maternal nutrition during gestation in sheep reduces circulating concentrations of insulin-like growth factor-I and insulin-like growth factor binding protein-3 in offspring. Domest. Anim. Endocrinol. 2014, 49, 39–48. [Google Scholar] [CrossRef]
- Hart, B.; Morgan, E.; Alejandro, E.U. Nutrient sensor signaling pathways and cellular stress in fetal growth restriction. J. Mol. Endocrinol. 2019, 62, R155–R165. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.J.; Ford, S.P.; Nathanielsz, P.W.; Du, M. Effect of maternal nutrient restriction in sheep on the development of fetal skeletal muscle. Biol. Reprod. 2004, 71, 1968–1973. [Google Scholar] [CrossRef] [PubMed]
- Ford, S.P.; Zhang, L.; Zhu, M.; Miller, M.M.; Smith, D.T.; Hess, B.W.; Moss, G.E.; Nathanielsz, P.W.; Nijland, M.J. Maternal obesity accelerates fetal pancreatic beta-cell but not alpha-cell development in sheep: Prenatal consequences. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R835–R843. [Google Scholar] [CrossRef] [PubMed]
- Long, N.M.; George, L.A.; Uthlaut, A.B.; Smith, D.T.; Nijland, M.J.; Nathanielsz, P.W.; Ford, S.P. Maternal obesity and increased nutrient intake before and during gestation in the ewe results in altered growth, adiposity, and glucose tolerance in adult offspring. J. Anim. Sci. 2010, 88, 3546–3553. [Google Scholar] [CrossRef] [PubMed]
- Vonnahme, K.A.; Hess, B.W.; Hansen, T.R.; McCormick, R.J.; Rule, D.C.; Moss, G.E.; Murdoch, W.J.; Nijland, M.J.; Skinner, D.C.; Nathanielsz, P.W.; et al. Maternal undernutrition from early- to mid-gestation leads to growth retardation, cardiac ventricular hypertrophy, and increased liver weight in the fetal sheep. Biol. Reprod. 2003, 69, 133–140. [Google Scholar] [CrossRef]
- Ford, S.P.; Hess, B.W.; Schwope, M.M.; Nijland, M.J.; Gilbert, J.S.; Vonnahme, K.A.; Means, W.J.; Han, H.; Nathanielsz, P.W. Maternal undernutrition during early to mid-gestation in the ewe results in altered growth, adiposity, and glucose tolerance in male offspring. J. Anim. Sci. 2007, 85, 1285–1294. [Google Scholar] [CrossRef]
- Smith, A.M.; Pankey, C.L.; Odhiambo, J.F.; Ghnenis, A.B.; Nathanielsz, P.W.; Ford, S.P. Rapid Communication: Reduced maternal nutrition during early- to mid-gestation elevates newborn lamb plasma cortisol concentrations and eliminates the neonatal leptin surge. J. Anim. Sci. 2018, 96, 2640–2645. [Google Scholar] [CrossRef]
- Zhu, M.J.; Ford, S.P.; Means, W.J.; Hess, B.W.; Nathanielsz, P.W.; Du, M. Maternal nutrient restriction affects properties of skeletal muscle in offspring. J. Physiol. 2006, 575, 241–250. [Google Scholar] [CrossRef]
- Peñagaricano, F.; Wang, X.; Rosa, G.J.; Radunz, A.E.; Khatib, H. Maternal nutrition induces gene expression changes in fetal muscle and adipose tissues in sheep. BMC Genom. 2014, 15, 1034. [Google Scholar] [CrossRef]
- Namous, H.; Peñagaricano, F.; Del Corvo, M.; Capra, E.; Thomas, D.L.; Stella, A.; Williams, J.L.; Marsan, P.A.; Khatib, H. Integrative analysis of methylomic and transcriptomic data in fetal sheep muscle tissues in response to maternal diet during pregnancy. BMC Genom. 2018, 19, 123. [Google Scholar] [CrossRef]
- Ashmore, C.R.; Robinson, D.W.; Rattray, P.; Doerr, L. Biphasic development of muscle fibers in the fetal lamb. Exp. Neurol. 1972, 37, 241–255. [Google Scholar] [CrossRef]
- Ma, Y.; Zhu, M.J.; Uthlaut, A.B.; Nijland, M.J.; Nathanielsz, P.W.; Hess, B.W.; Ford, S.P. Upregulation of growth signaling and nutrient transporters in cotyledons of early to mid-gestational nutrient restricted ewes. Placenta 2011, 32, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Long, N.M.; Ford, S.P.; Nathanielsz, P.W. Maternal obesity eliminates the neonatal lamb plasma leptin peak. J. Physiol. 2011, 589, 1455–1462. [Google Scholar] [CrossRef] [PubMed]
- Demirtas, B.; Ozcan, M. The effect of maternal undernutrition on muscle development in the ovine fetus. Turk. J. Vet. Anim. Sci. 2012, 36, 297–303. [Google Scholar] [CrossRef]
- Yan, X.; Huang, Y.; Zhao, J.X.; Long, N.M.; Uthlaut, A.B.; Zhu, M.J.; Ford, S.P.; Nathanielsz, P.W.; Du, M. Maternal Obesity-Impaired Insulin Signaling in Sheep and Induced Lipid Accumulation and Fibrosis in Skeletal Muscle of Offspring. Biol. Reprod. 2011, 85, 172–178. [Google Scholar] [CrossRef]
- Maloney, C.A.; Lilley, C.; Czopek, A.; Hay, S.M.; Rees, W.D. Interactions between protein and vegetable oils in the maternal diet determine the programming of the insulin axis in the rat. Br. J. Nutr. 2007, 97, 912–920. [Google Scholar] [CrossRef]
- Hindi, S.M.; Tajrishi, M.M.; Kumar, A. Signaling mechanisms in mammalian myoblast fusion. Sci. Signal. 2013, 6, re2. [Google Scholar] [CrossRef]
- Chu, C.Y.; Lim, R.W. Involvement of p27(kip1) and cyclin D3 in the regulation of cdk2 activity during skeletal muscle differentiation. Biochim. Biophys. Acta 2000, 1497, 175–185. [Google Scholar] [CrossRef]
- Clegg, C.H.; Linkhart, T.A.; Olwin, B.B.; Hauschka, S.D. Growth factor control of skeletal muscle differentiation: Commitment to terminal differentiation occurs in G1 phase and is repressed by fibroblast growth factor. J. Cell Biol. 1987, 105, 949–956. [Google Scholar] [CrossRef]
- Guo, K.; Wang, J.; Andrés, V.; Smith, R.C.; Walsh, K. MyoD-induced expression of p21 inhibits cyclin-dependent kinase activity upon myocyte terminal differentiation. Mol. Cell Biol. 1995, 15, 3823–3829. [Google Scholar] [CrossRef]
- Campaner, S.; Doni, M.; Hydbring, P.; Verrecchia, A.; Bianchi, L.; Sardella, D.; Schleker, T.; Perna, D.; Tronnersjö, S.; Murga, M.; et al. Cdk2 suppresses cellular senescence induced by the c-myc oncogene. Nat. Cell Biol. 2010, 12, 54–59. [Google Scholar] [CrossRef]
- David, A.; Liu, F.; Tibelius, A.; Vulprecht, J.; Wald, D.; Rothermel, U.; Ohana, R.; Seitel, A.; Metzger, J.; Ashery-Padan, R.; et al. Lack of centrioles and primary cilia in STIL(−/−) mouse embryos. Cell Cycle 2014, 13, 2859–2868. [Google Scholar] [CrossRef] [PubMed]
- Stearns, T. Centrosome duplication. a centriolar pas de deux. Cell 2001, 105, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Tarapore, P.; Okuda, M.; Fukasawa, K. A mammalian in vitro centriole duplication system: Evidence for involvement of CDK2/cyclin E and nucleophosmin/B23 in centrosome duplication. Cell Cycle 2002, 1, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Tarapore, P.; Horn, H.F.; Tokuyama, Y.; Fukasawa, K. Direct regulation of the centrosome duplication cycle by the p53-p21Waf1/Cip1 pathway. Oncogene 2001, 20, 3173–3184. [Google Scholar] [CrossRef]
- Gupta, M.; Gupta, S.K.; Hoffman, B.; Liebermann, D.A. Gadd45a and Gadd45b protect hematopoietic cells from UV-induced apoptosis via distinct signaling pathways, including p38 activation and JNK inhibition. J. Biol. Chem. 2006, 281, 17552–17558. [Google Scholar] [CrossRef]
- Yan, Y.; Frisén, J.; Lee, M.H.; Massagué, J.; Barbacid, M. Ablation of the CDK inhibitor p57Kip2 results in increased apoptosis and delayed differentiation during mouse development. Genes Dev. 1997, 11, 973–983. [Google Scholar] [CrossRef]
- Eapen, S.A.; Netherton, S.J.; Sarker, K.P.; Deng, L.; Chan, A.; Riabowol, K.; Bonni, S. Identification of a novel function for the chromatin remodeling protein ING2 in muscle differentiation. PLoS ONE 2012, 7, e40684. [Google Scholar] [CrossRef]
- Archambeau, J.; Blondel, A.; Pedeux, R. Focus-ING on DNA Integrity: Implication of ING Proteins in Cell Cycle Regulation and DNA Repair Modulation. Cancers 2019, 12, 58. [Google Scholar] [CrossRef]
- Nagashima, M.; Shiseki, M.; Miura, K.; Hagiwara, K.; Linke, S.P.; Pedeux, R.; Wang, X.W.; Yokota, J.; Riabowol, K.; Harris, C.C. DNA damage-inducible gene p33ING2 negatively regulates cell proliferation through acetylation of p53. Proc. Natl. Acad. Sci. USA 2001, 98, 9671–9676. [Google Scholar] [CrossRef]
- D’Assoro, A.B.; Barrett, S.L.; Folk, C.; Negron, V.C.; Boeneman, K.; Busby, R.; Whitehead, C.; Stivala, F.; Lingle, W.L.; Salisbury, J.L. Amplified centrosomes in breast cancer: A potential indicator of tumor aggressiveness. Breast Cancer Res. Treat. 2002, 75, 25–34. [Google Scholar] [CrossRef]
- Mourtada-Maarabouni, M.; Kirkham, L.; Farzaneh, F.; Williams, G.T. Regulation of apoptosis by fau revealed by functional expression cloning and antisense expression. Oncogene 2004, 23, 9419–9426. [Google Scholar] [CrossRef] [PubMed]
- Gahlmann, R.; Wade, R.; Gunning, P.; Kedes, L. Differential expression of slow and fast skeletal muscle troponin C. Slow skeletal muscle troponin C is expressed in human fibroblasts. J. Mol. Biol. 1988, 201, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Schreier, T.; Kedes, L.; Gahlmann, R. Cloning, structural analysis, and expression of the human slow twitch skeletal muscle/cardiac troponin C gene. J. Biol. Chem. 1990, 265, 21247–21253. [Google Scholar] [CrossRef] [PubMed]
- Townsend, P.J.; Yacoub, M.H.; Barton, P.J. Assignment of the human cardiac/slow skeletal muscle troponin C gene (TNNC1) between D3S3118 and GCT4B10 on the short arm of chromosome 3 by somatic cell hybrid analysis. Ann. Hum. Genet. 1997, 61, 375–377. [Google Scholar] [CrossRef]
- Kretsinger, R.H.; Nockolds, C.E. Carp muscle calcium-binding protein. II. Structure determination and general description. J. Biol. Chem. 1973, 248, 3313–3326. [Google Scholar] [CrossRef]
- Li, M.X.; Hwang, P.M. Structure and function of cardiac troponin C (TNNC1): Implications for heart failure, cardiomyopathies, and troponin modulating drugs. Gene 2015, 571, 153–166. [Google Scholar] [CrossRef]
- Wang, F.; Wang, H.; Wu, H.; Qiu, H.; Zeng, C.; Sun, L.; Liu, B. TEAD1 controls C2C12 cell proliferation and differentiation and regulates three novel target genes. Cell Signal. 2013, 25, 674–681. [Google Scholar] [CrossRef]
- Qiu, H.; Wang, F.; Liu, C.; Xu, X.; Liu, B. TEAD1-dependent expression of the FoxO3a gene in mouse skeletal muscle. BMC Mol. Biol. 2011, 12, 1. [Google Scholar] [CrossRef]
- Zhao, B.; Lei, Q.Y.; Guan, K.L. The Hippo-YAP pathway: New connections between regulation of organ size and cancer. Curr. Opin. Cell Biol. 2008, 20, 638–646. [Google Scholar] [CrossRef]
- Osman, I.; He, X.; Liu, J.; Dong, K.; Wen, T.; Zhang, F.; Yu, L.; Hu, G.; Xin, H.-B.; Zhang, W.; et al. TEAD1 (TEA Domain Transcription Factor 1) Promotes Smooth Muscle Cell Proliferation Through Upregulating SLC1A5 (Solute Carrier Family 1 Member 5)-Mediated Glutamine Uptake. Circ. Res. 2019, 124, 1309–1322. [Google Scholar] [CrossRef]
- Wen, T.; Liu, J.; He, X.; Dong, K.; Hu, G.; Yu, L.; Yin, Q.; Osman, I.; Peng, J.; Zheng, Z.; et al. Transcription factor TEAD1 is essential for vascular development by promoting vascular smooth muscle differentiation. Cell Death Differ. 2019, 26, 2790–2806. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Ye, X.; Yu, J.; Li, L.; Li, W.; Li, S.; Yu, J.; Lin, J.D.; Wang, C.-Y.; Chinnaiyan, A.M.; et al. TEAD mediates YAP-dependent gene induction and growth control. Genes Dev. 2008, 22, 1962–1971. [Google Scholar] [CrossRef] [PubMed]
- Hansen, C.G.; Moroishi, T.; Guan, K.L. YAP and TAZ: A nexus for Hippo signaling and beyond. Trends Cell Biol. 2015, 25, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Wackerhage, H.; Del Re, D.P.; Judson, R.N.; Sudol, M.; Sadoshima, J. The Hippo signal transduction network in skeletal and cardiac muscle. Sci. Signal. 2014, 7, re4. [Google Scholar] [CrossRef] [PubMed]
- Watt, K.I.; Turner, B.J.; Hagg, A.; Zhang, X.; Davey, J.R.; Qian, H.; Beyer, C.; Winbanks, C.E.; Harvey, K.F.; Gregorevic, P. The Hippo pathway effector YAP is a critical regulator of skeletal muscle fibre size. Nat. Commun. 2015, 6, 6048. [Google Scholar] [CrossRef] [PubMed]
- Maeda, T.; Chapman, D.L.; Stewart, A.F. Mammalian vestigial-like 2, a cofactor of TEF-1 and MEF2 transcription factors that promotes skeletal muscle differentiation. J. Biol. Chem. 2002, 277, 48889–48898. [Google Scholar] [CrossRef]
- Hélias-Rodzewicz, Z.; Pérot, G.; Chibon, F.; Ferreira, C.; Lagarde, P.; Terrier, P.; Coindre, J.M.; Aurias, A. YAP1 and VGLL3, encoding two cofactors of TEAD transcription factors, are amplified and overexpressed in a subset of soft tissue sarcomas. Genes Chromosom. Cancer 2010, 49, 1161–1171. [Google Scholar] [CrossRef]
- Southard, S.; Kim, J.R.; Low, S.; Tsika, R.W.; Lepper, C. Myofiber-specific TEAD1 overexpression drives satellite cell hyperplasia and counters pathological effects of dystrophin deficiency. Elife 2016, 5, e15461. [Google Scholar] [CrossRef]
- Figeac, N.; Mohamed, A.D.; Sun, C.; Schönfelder, M.; Matallanas, D.; Garcia-Munoz, A.; Missiaglia, E.; Collie-Duguid, E.; De Mello, V.; Pobbati, A.V.; et al. VGLL3 operates via TEAD1, TEAD3 and TEAD4 to influence myogenesis in skeletal muscle. J. Cell Sci. 2019, 132, jcs225946. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Shang, M.; Hu, W.; Zhang, L. Multi-Omics Analysis Reveals the Potential Effects of Maternal Dietary Restriction on Fetal Muscle Growth and Development. Nutrients 2023, 15, 1051. https://doi.org/10.3390/nu15041051
Wang X, Shang M, Hu W, Zhang L. Multi-Omics Analysis Reveals the Potential Effects of Maternal Dietary Restriction on Fetal Muscle Growth and Development. Nutrients. 2023; 15(4):1051. https://doi.org/10.3390/nu15041051
Chicago/Turabian StyleWang, Xinyue, Mingyu Shang, Wenping Hu, and Li Zhang. 2023. "Multi-Omics Analysis Reveals the Potential Effects of Maternal Dietary Restriction on Fetal Muscle Growth and Development" Nutrients 15, no. 4: 1051. https://doi.org/10.3390/nu15041051
APA StyleWang, X., Shang, M., Hu, W., & Zhang, L. (2023). Multi-Omics Analysis Reveals the Potential Effects of Maternal Dietary Restriction on Fetal Muscle Growth and Development. Nutrients, 15(4), 1051. https://doi.org/10.3390/nu15041051