

Improvement of Spatial Memory and Cognitive Function in Mice via the Intervention of Milk Fat Globule Membrane

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Determination of Physicochemical Composition of MFGM
2.3. Animals and Experimental Design
2.4. Morris Water Maze (MWM) Experimental Design
2.5. Histological Analysis of the Hippocampus
2.5.1. Hematoxylin and Eosin (H&E) Staining
2.5.2. Nissl Staining
2.6. Proteomics Analyses in Hippocampal Tissue of Mice
2.6.1. Tandem Mass Tag (TMT) Labeling
2.6.2. Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS) and Bioinformatics Analysis
2.7. Statistical Analysis
3. Results
3.1. Composition Analysis of MFGM
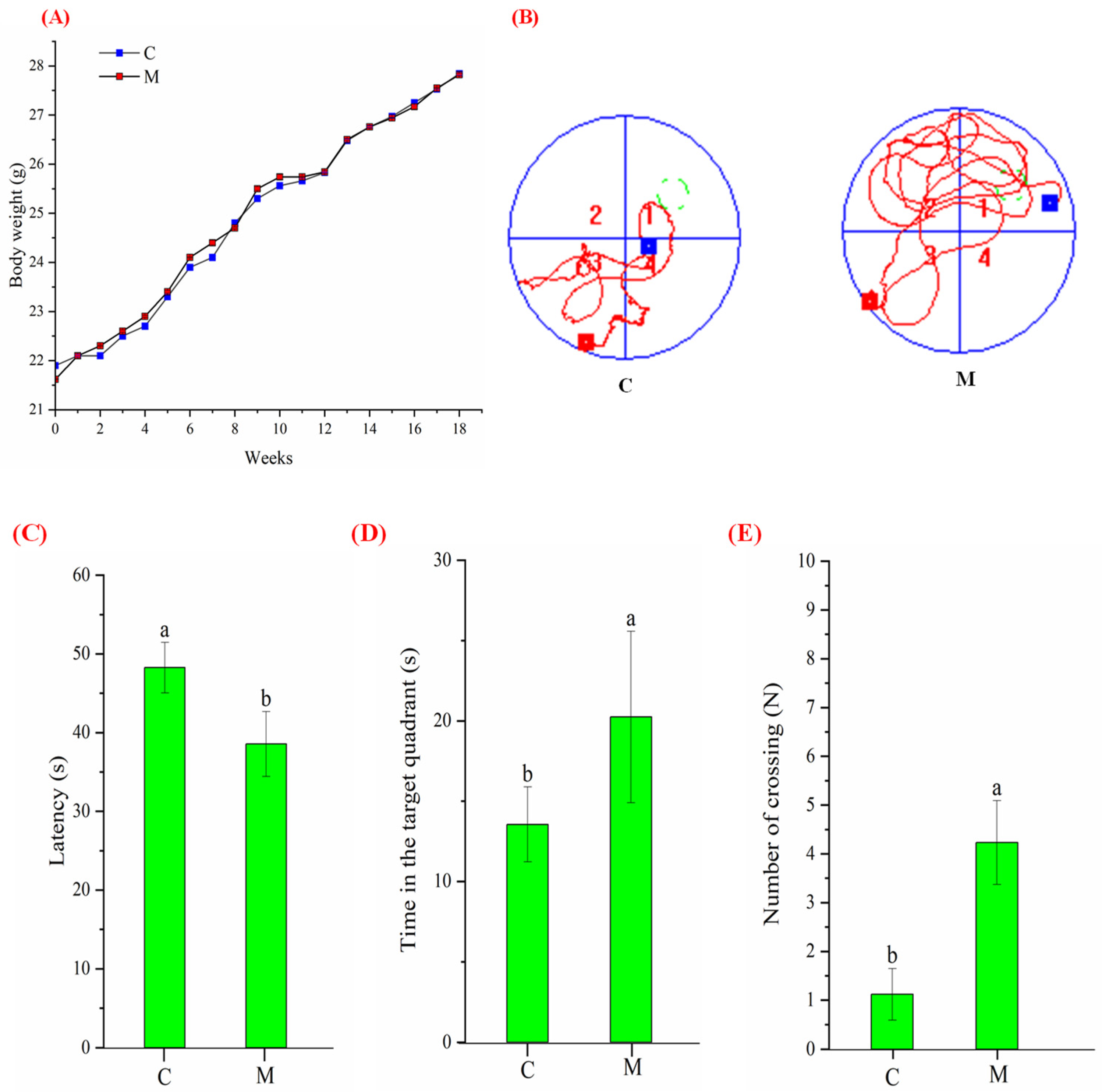
3.2. Growth and Morris Water Maze Performance
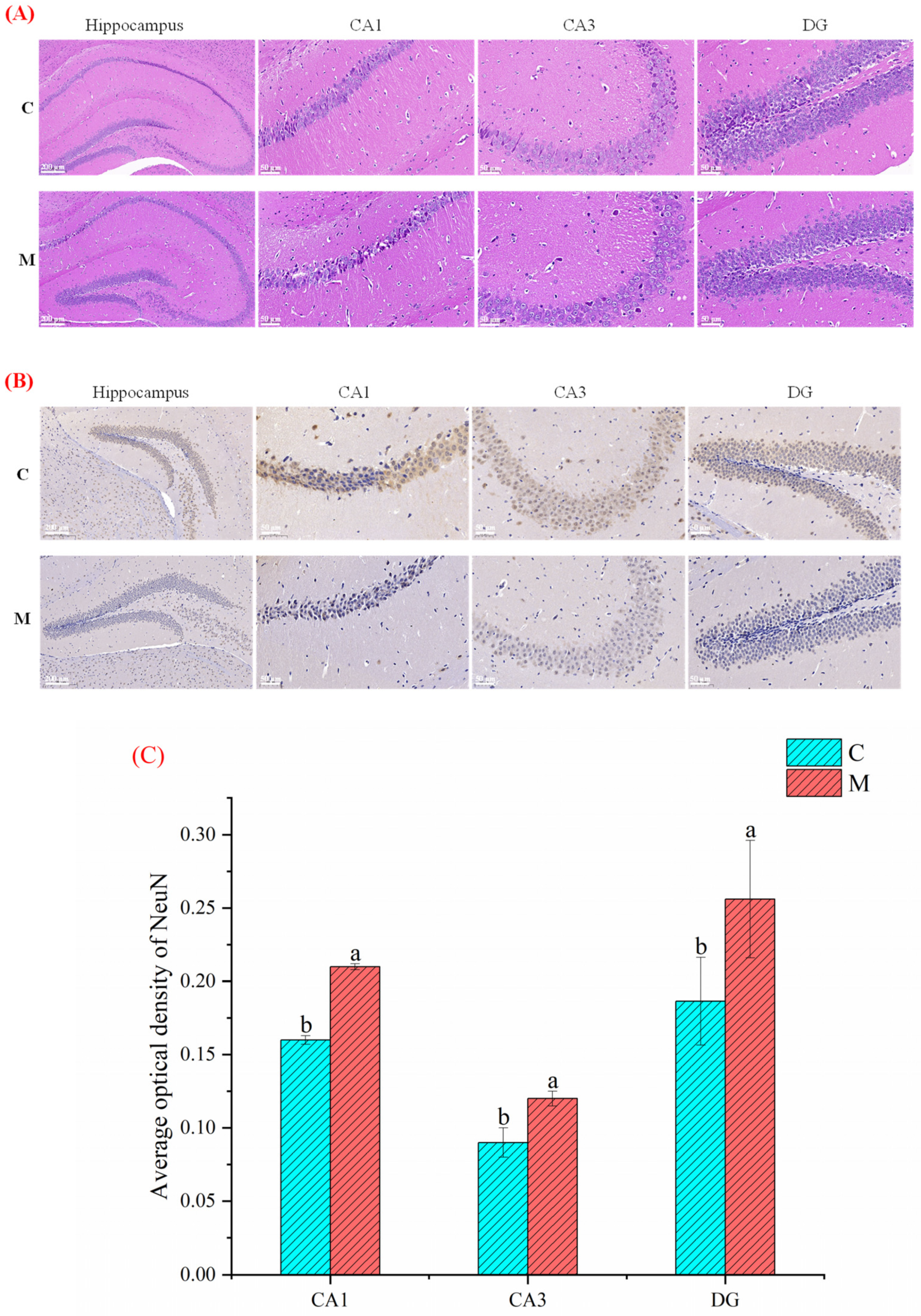
3.3. Effect of MFGM on Hippocampus Histological Analysis
3.4. Proteomic Analysis
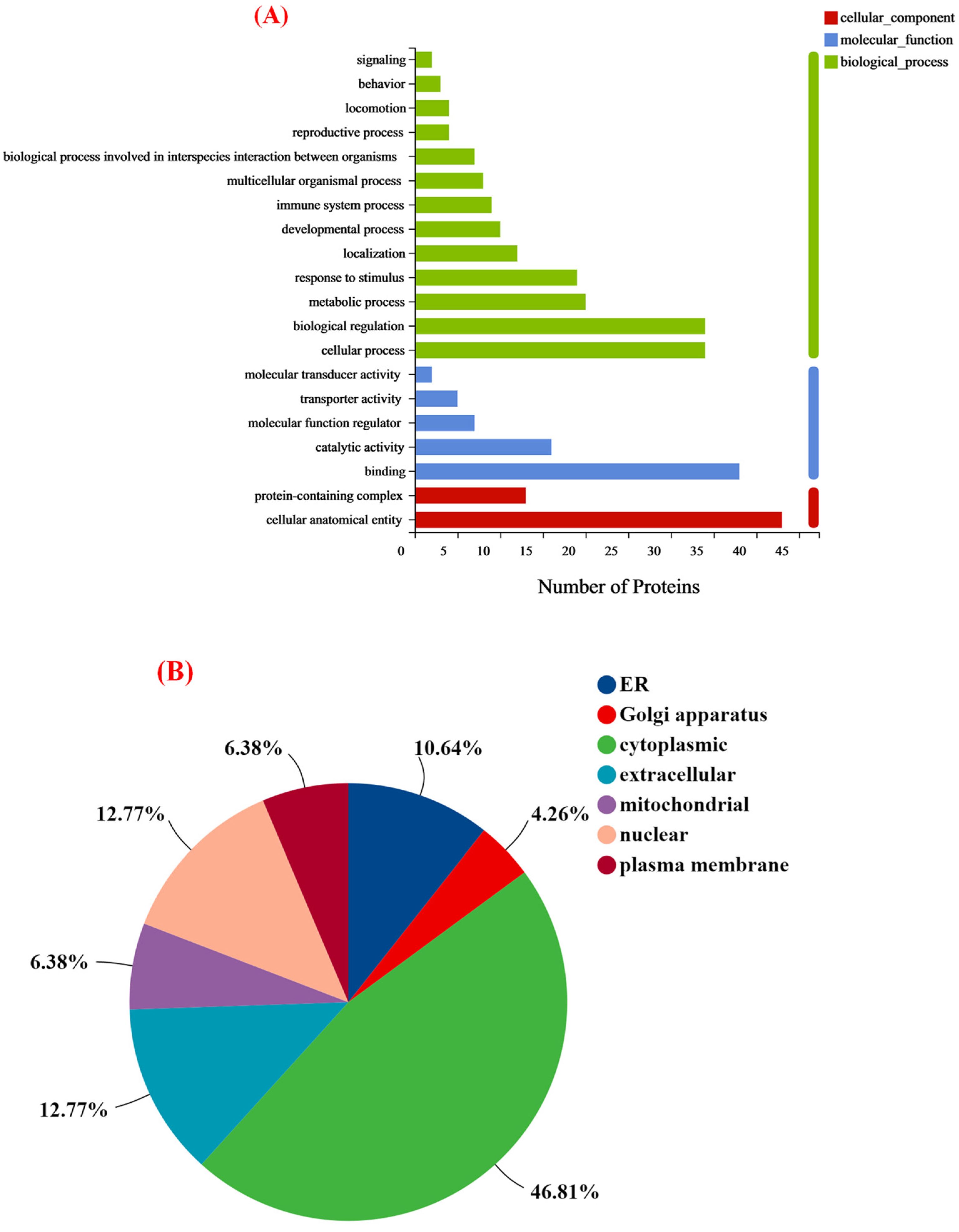
3.4.1. Full Protein Function Annotation
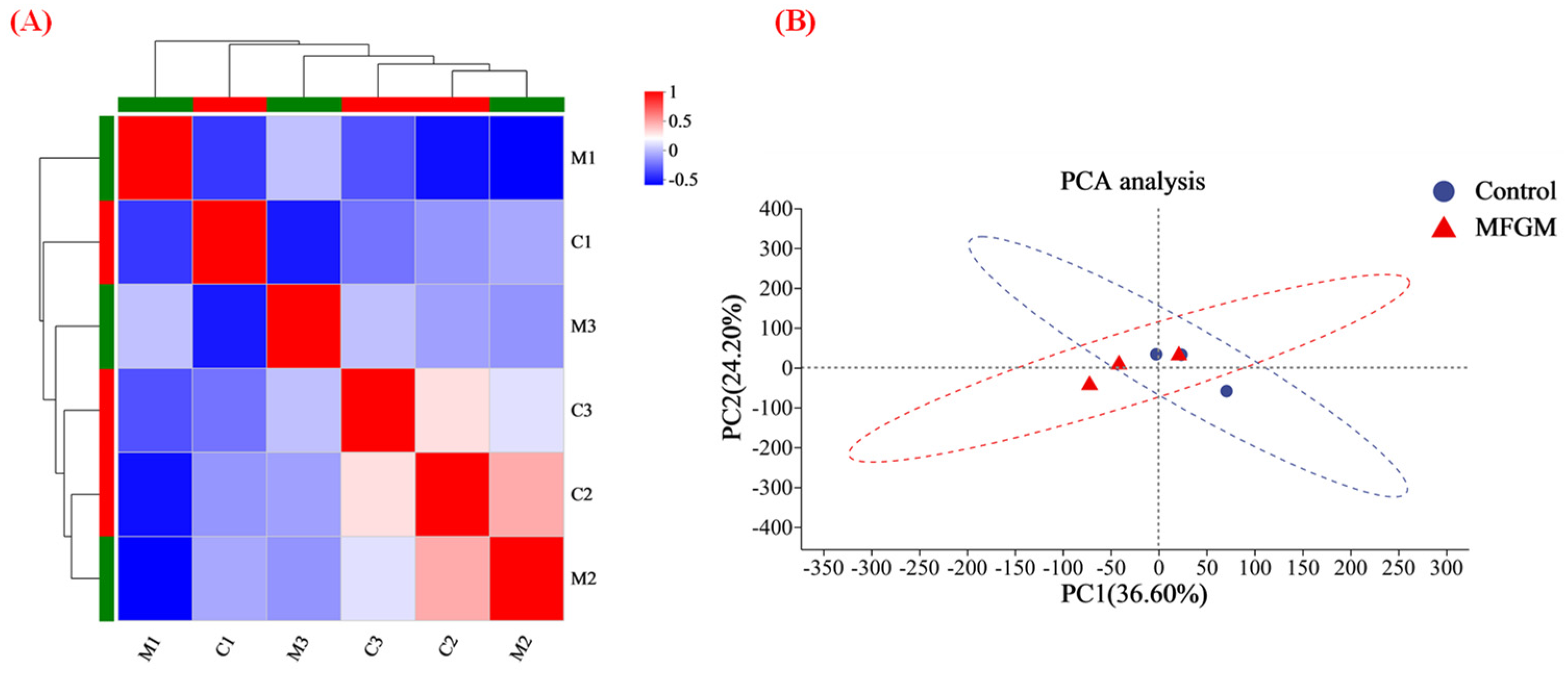
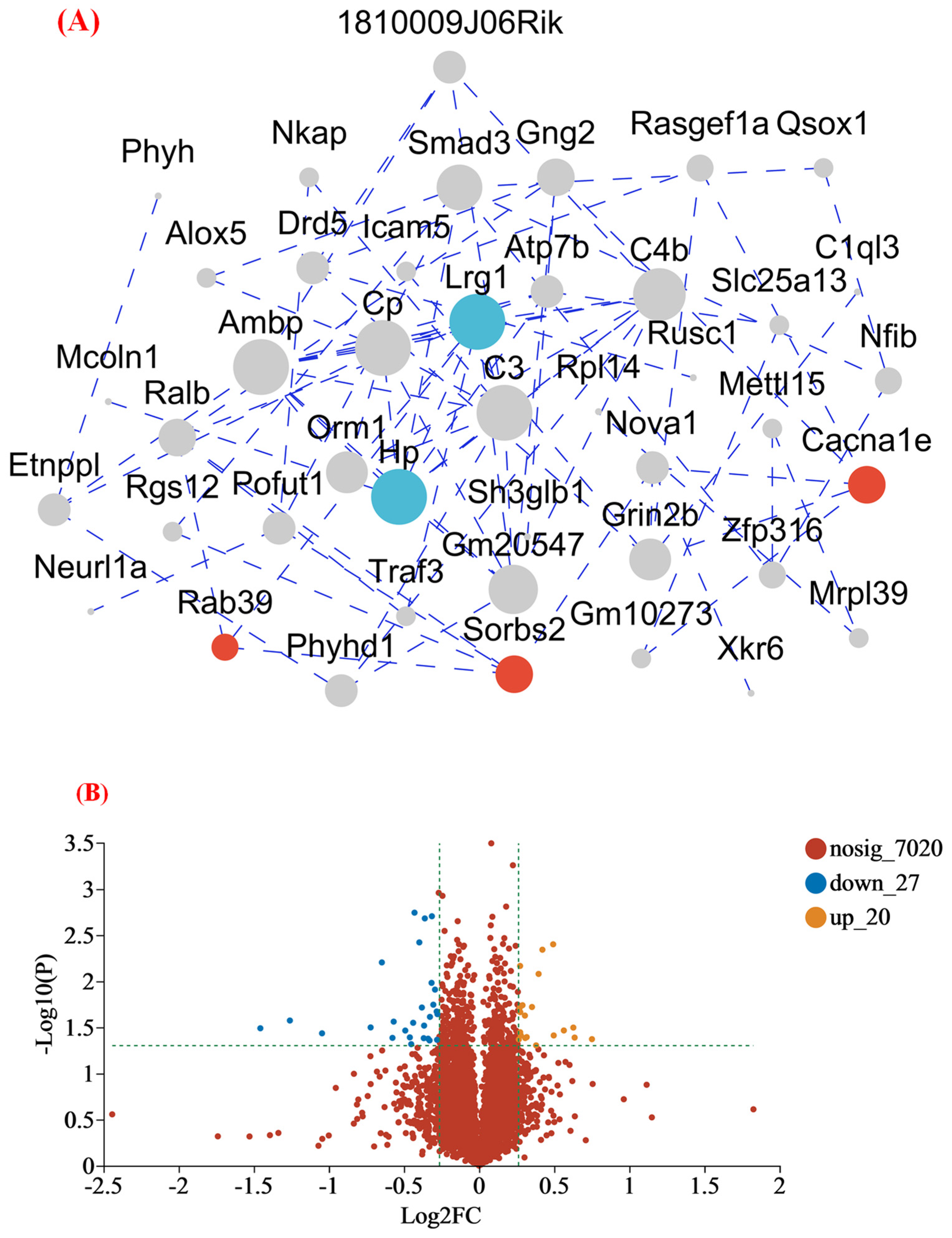
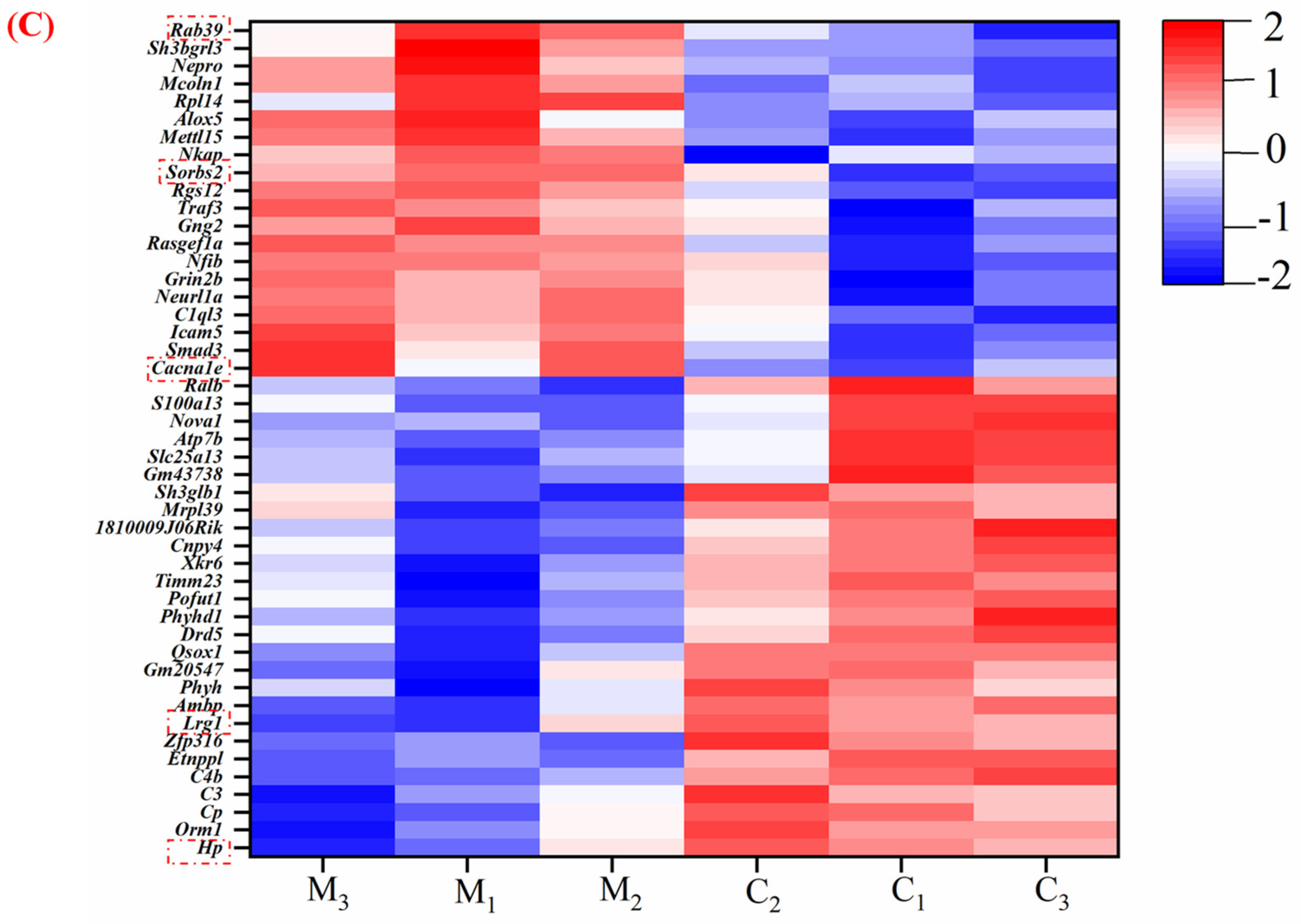
3.4.2. MFGM Altered the Hippocampus Proteomic Profiles
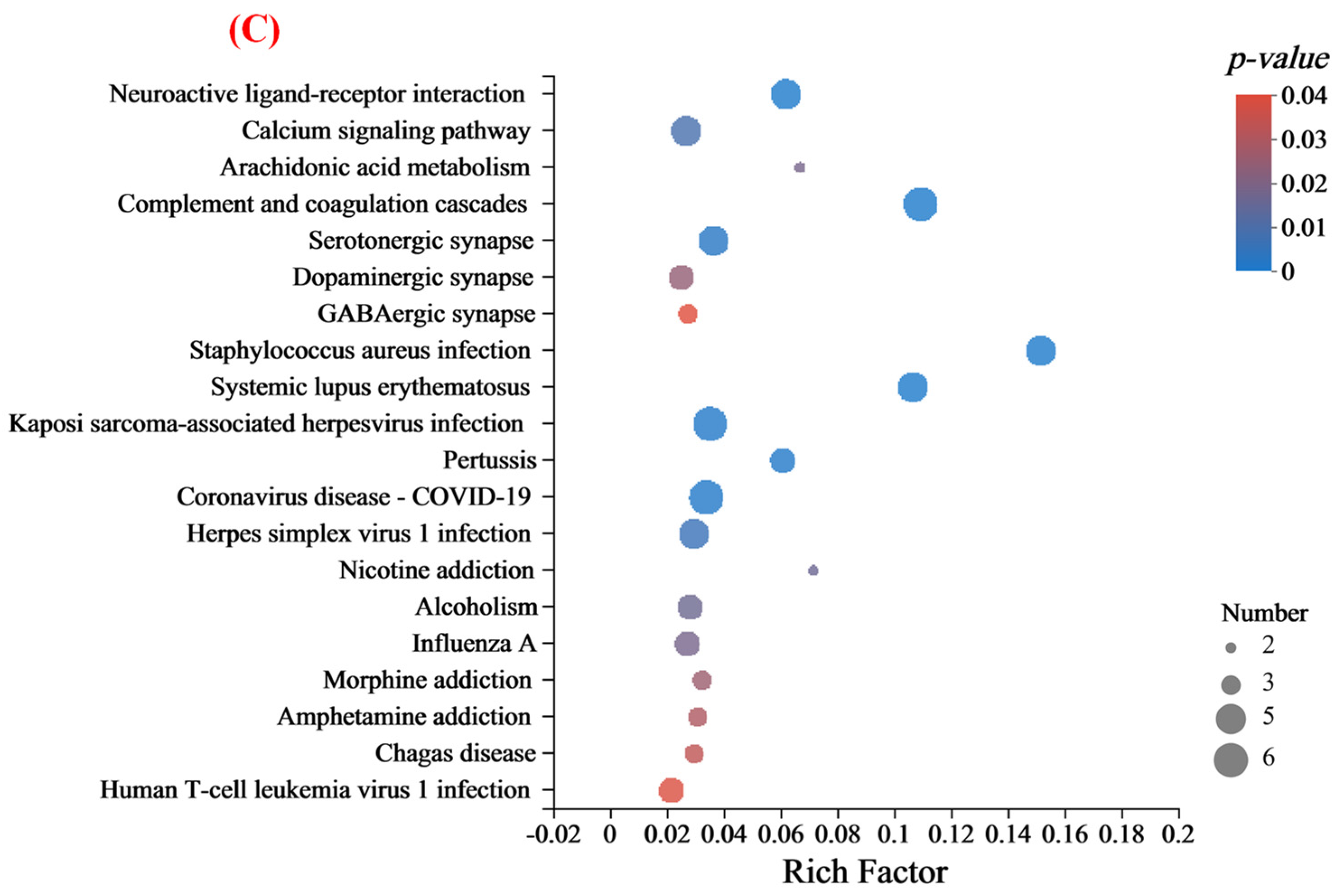
3.4.3. Functional Annotation of Differentially Expressed Proteins
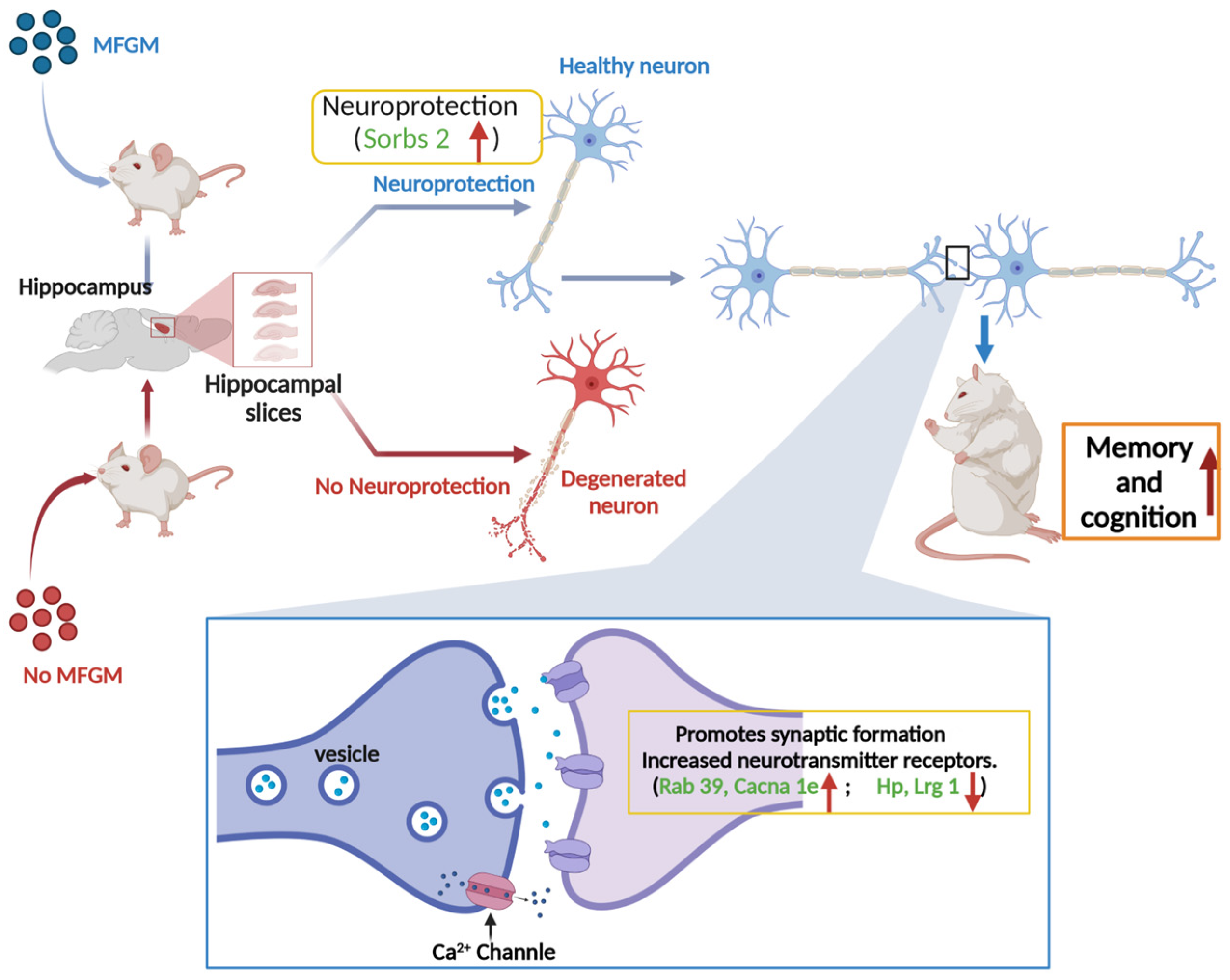
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Joels, M.; Pu, Z.; Wiegert, O.; Oitzl, M.S.; Krugers, H.J. Learning under stress: How does it work? Trends Cogn. Sci. 2006, 10, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, L.; Joels, M.; Roozendaal, B.; Wolf, O.T.; Oitzl, M.S. Stress effects on memory: An update and integration. Neurosci. Biobehav. Rev. 2012, 36, 1740–1749. [Google Scholar] [CrossRef]
- Pitman, R.K.; Rasmusson, A.M.; Koenen, K.C.; Shin, L.M.; Orr, S.P.; Gilbertson, M.W.; Milad, M.R.; Liberzon, I. Biological studies of post-traumatic stress disorder. Nat. Rev. Neurosci. 2012, 13, 769–787. [Google Scholar] [CrossRef]
- Szabo, M.P.; Mishra, S.; Knupp, A.; Young, J.E. The role of Alzheimer’s disease risk genes in endolysosomal pathways. Neurobiol. Dis. 2022, 162, 105576. [Google Scholar] [CrossRef] [PubMed]
- Fang, E.F.; Scheibye-Knudsen, M.; Jahn, H.J.; Li, J.; Ling, L.; Guo, H.; Zhu, X.; Preedy, V.; Lu, H.; Bohr, V.A.; et al. A research agenda for aging in China in the 21st century. Ageing Res. Rev. 2015, 24, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Bietry, F.A.; Pfeil, A.M.; Reich, O.; Schwenkglenks, M.; Meier, C.R. Benzodiazepine Use and Risk of Developing Alzheimer’s Disease: A Case-Control Study Based on Swiss Claims Data. CNS Drugs 2017, 31, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Shelly, L.; Gray, M.A. Rebecca Hubbard. Anticholinergic Use With Incident Dementia—Reply. JAMA Intern. Med. 2015, 175, 1577. [Google Scholar] [CrossRef]
- Borda, M.G.; Jaramillo-Jimenez, A.; Oesterhus, R.; Santacruz, J.M.; Tovar-Rios, D.A.; Soennesyn, H.; Cano-Gutierrez, C.A.; Vik-Mo, A.O.; Aarsland, D. Benzodiazepines and antidepressants: Effects on cognitive and functional decline in Alzheimer’s disease and Lewy body dementia. Int. J. Geriatr. Psychiatry 2021, 36, 917–925. [Google Scholar] [CrossRef]
- Abd El-Salam, M.H.; El-Shibiny, S. Milk fat globule membrane: An overview with particular emphasis on its nutritional and health benefits. Int. J. Dairy Technol. 2020, 73, 639–655. [Google Scholar] [CrossRef]
- Mudd, A.T.; Alexander, L.S.; Berding, K.; Waworuntu, R.V.; Berg, B.M.; Donovan, S.M.; Dilger, R.N. Dietary Prebiotics, Milk Fat Globule Membrane, and Lactoferrin Affects Structural Neurodevelopment in the Young Piglet. Front. Pediatr. 2016, 4, 4. [Google Scholar] [CrossRef]
- Hernell, O.; Timby, N.; Domellof, M.; Lonnerdal, B. Clinical Benefits of Milk Fat Globule Membranes for Infants and Children. J. Pediatr. 2016, 173, S60–S65. [Google Scholar] [CrossRef]
- Timby, N.; Domellof, E.; Hernell, O.; Lonnerdal, B.; Domellof, M. Neurodevelopment, nutrition, and growth until 12 mo of age in infants fed a low-energy, low-protein formula supplemented with bovine milk fat globule membranes: A randomized controlled trial. Am. J. Clin. Nutr. 2014, 99, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wang, C.; Zhao, N.; Li, W.; Yang, Z.; Liu, X.; Le, W.; Zhang, X. Potential biomarkers of Parkinson’s disease revealed by plasma metabolic profiling. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2018, 1081–1082, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Cheng, S.L.; Fei, Q.; Gu, H.; Raftery, D.; Cao, B.; Sun, X.; Yan, J.; Zhang, C.; Wang, J. Metabolic profiling identifies phospholipids as potential serum biomarkers for schizophrenia. Psychiatry Res. 2019, 272, 18–29. [Google Scholar] [CrossRef]
- O’Mahony, S.M.; McVey Neufeld, K.A.; Waworuntu, R.V.; Pusceddu, M.M.; Manurung, S.; Murphy, K.; Strain, C.; Laguna, M.C.; Peterson, V.L.; Stanton, C.; et al. The enduring effects of early-life stress on the microbiota-gut-brain axis are buffered by dietary supplementation with milk fat globule membrane and a prebiotic blend. Eur. J. Neurosci. 2020, 51, 1042–1058. [Google Scholar] [CrossRef]
- Brink, L.R.; Gueniot, J.P.; Lonnerdal, B. Effects of milk fat globule membrane and its various components on neurologic development in a postnatal growth restriction rat model. J. Nutr. Biochem. 2019, 69, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Vieira, E.F.; Soares, C.; Machado, S.; Correia, M.; Ramalhosa, M.J.; Oliva-Teles, M.T.; Paula Carvalho, A.; Domingues, V.F.; Antunes, F.; Oliveira, T.A.C.; et al. Seaweeds from the Portuguese coast as a source of proteinaceous material: Total and free amino acid composition profile. Food Chem. 2018, 269, 264–275. [Google Scholar] [CrossRef]
- Ferraris, Q.; Hale, J.; Teigland, E.; Rao, A.; Qian, M.C. Phospholipid analysis in whey protein products using hydrophilic interaction high-performance liquid chromatography-evaporative light-scattering detection in an industry setting. J. Dairy Sci. 2020, 103, 11079–11085. [Google Scholar] [CrossRef]
- Tucker, L.B.; Velosky, A.G.; McCabe, J.T. Applications of the Morris water maze in translational traumatic brain injury research. Neurosci. Biobehav. Rev. 2018, 88, 187–200. [Google Scholar] [CrossRef]
- Geng, M.; Zhao, F.; Lu, H.; Fang, L.; Wang, J.; Liu, C.; Min, W. Insights into the hippocampus proteome and phosphorylation modification alterations in C57BL/6 revealed the memory improvement mechanisms of a walnut-derived peptide. Food Res. Int. 2022, 156, 111311. [Google Scholar] [CrossRef]
- Wang, K.; Shi, J.; Gao, S.; Hong, H.; Tan, Y.; Luo, Y. Oyster protein hydrolysates alleviated chronic alcohol-induced liver injury in mice by regulating hepatic lipid metabolism and inflammation response. Food Res. Int. 2022, 160, 111647. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Yu, Y.; Wang, C.; Zhu, Q.; Wu, J.; Ke, W.; Ji, D.; Niu, C.; Yang, X.; Wei, Y. Hippocampal proteomic analysis reveals the disturbance of synaptogenesis and neurotransmission induced by developmental exposure to organophosphate flame retardant triphenyl phosphate. J. Hazard. Mater. 2021, 404, 124111. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Wu, Z.; Liu, C.; Han, D.; Feng, C.; Wang, S.; Wang, J. Milk Fat Globule Membrane Supplementation Promotes Neonatal Growth and Alleviates Inflammation in Low-Birth-Weight Mice Treated with Lipopolysaccharide. Biomed. Res. Int 2019, 2019, 4876078. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.C.; Gong, H.; Du, M.; Mao, X.Y. Supplementation of milk polar lipids to obese dams improves neurodevelopment and cognitive function in male offspring. FASEB J. 2021, 35, e21454. [Google Scholar] [CrossRef]
- Schverer, M.; O’Mahony, S.M.; O’Riordan, K.J.; Donoso, F.; Roy, B.L.; Stanton, C.; Dinan, T.G.; Schellekens, H.; Cryan, J.F. Dietary phospholipids: Role in cognitive processes across the lifespan. Neurosci. Biobehav. Rev. 2020, 111, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.E.; Broadbent, N.J.; Squire, L.R. Hippocampus and remote spatial memory in rats. Hippocampus 2005, 15, 260–272. [Google Scholar] [CrossRef]
- Dinel, A.L.; Lucas, C.; Guillemet, D.; Laye, S.; Pallet, V.; Joffre, C. Chronic Supplementation with a Mix of Salvia officinalis and Salvia lavandulaefolia Improves Morris Water Maze Learning in Normal Adult C57Bl/6J Mice. Nutrients 2020, 12, 1777. [Google Scholar] [CrossRef]
- Kourosh-Arami, M.; Komaki, A.; Joghataei, M.T.; Mohsenzadegan, M. Phospholipase Cbeta3 in the hippocampus may mediate impairment of memory by long-term blockade of orexin 1 receptors assessed by the Morris water maze. Life Sci. 2020, 257, 118046. [Google Scholar] [CrossRef]
- Lv, H.; Tang, L.; Guo, C.; Jiang, Y.; Gao, C.; Wang, Y.; Jian, C. Intranasal insulin administration may be highly effective in improving cognitive function in mice with cognitive dysfunction by reversing brain insulin resistance. Cogn. Neurodyn. 2020, 14, 323–338. [Google Scholar] [CrossRef]
- Ma, X.; Huang, M.; Zheng, M.; Dai, C.; Song, Q.; Zhang, Q.; Li, Q.; Gu, X.; Chen, H.; Jiang, G.; et al. ADSCs-derived extracellular vesicles alleviate neuronal damage, promote neurogenesis and rescue memory loss in mice with Alzheimer’s disease. J. Control. Release 2020, 327, 688–702. [Google Scholar] [CrossRef]
- Benesty, J.; Chen, J.; Huang, Y.; Cohen, I. Pearson Correlation Coefficient. In Noise Reduction in Speech Processing; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–4. [Google Scholar] [CrossRef]
- Joshi, Y.B.; Giannopoulos, P.F.; Chu, J.; Sperow, M.; Kirby, L.G.; Abood, M.E.; Pratico, D. Absence of ALOX5 gene prevents stress-induced memory deficits, synaptic dysfunction and tauopathy in a mouse model of Alzheimer’s disease. Hum. Mol. Genet. 2014, 23, 6894–6902. [Google Scholar] [CrossRef] [PubMed]
- Niu, M.; Zheng, N.; Wang, Z.; Gao, Y.; Luo, X.; Chen, Z.; Fu, X.; Wang, Y.; Wang, T.; Liu, M.; et al. RAB39B Deficiency Impairs Learning and Memory Partially Through Compromising Autophagy. Front. Cell Dev. Biol. 2020, 8, 598622. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Gao, X.; Li, C.; Feliciano, C.; Wang, D.; Zhou, D.; Mei, Y.; Monteiro, P.; Anand, M.; Itohara, S.; et al. Impaired Dendritic Development and Memory in Sorbs2 Knock-Out Mice. J. Neurosci. 2016, 36, 2247–2260. [Google Scholar] [CrossRef]
- Kessi, M.; Chen, B.; Peng, J.; Yan, F.; Yang, L.; Yin, F. Calcium channelopathies and intellectual disability: A systematic review. Orphanet J. Rare Dis. 2021, 16, 219. [Google Scholar] [CrossRef]
- Corbeel, L.; Freson, K. Rab proteins and Rab-associated proteins: Major actors in the mechanism of protein-trafficking disorders. Eur. J. Pediatr. 2008, 167, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Durand-de Cuttoli, R.; Chauhan, P.S.; Petriz Reyes, A.; Faure, P.; Mourot, A.; Ellis-Davies, G.C.R. Optofluidic control of rodent learning using cloaked caged glutamate. Proc. Natl. Acad. Sci. USA 2020, 117, 6831–6835. [Google Scholar] [CrossRef] [PubMed]
- Andrade, A.; Brennecke, A.; Mallat, S.; Brown, J.; Gomez-Rivadeneira, J.; Czepiel, N.; Londrigan, L. Genetic Associations between Voltage-Gated Calcium Channels and Psychiatric Disorders. Int. J. Mol. Sci. 2019, 20, 3537. [Google Scholar] [CrossRef]
- Guerrero-Berroa, E.; Ravona-Springer, R.; Heymann, A.; Schmeidler, J.; Levy, A.; Leroith, D.; Beeri, M.S. Haptoglobin genotype modulates the relationships of glycaemic control with cognitive function in elderly individuals with type 2 diabetes. Diabetologia 2015, 58, 736–744. [Google Scholar] [CrossRef]
- Miao, Y.; Wang, M.; Cai, X.; Zhu, Q.; Mao, L. Leucine rich alpha-2-glycoprotein 1 (Lrg1) silencing protects against sepsis-mediated brain injury by inhibiting transforming growth factor beta1 (TGFbeta1)/SMAD signaling pathway. Bioengineered 2022, 13, 7316–7327. [Google Scholar] [CrossRef]
- Jin, J.; Sun, H.; Liu, D.; Wang, H.; Liu, Q.; Chen, H.; Zhong, D.; Li, G. LRG1 Promotes Apoptosis and Autophagy through the TGFbeta-smad1/5 Signaling Pathway to Exacerbate Ischemia/Reperfusion Injury. Neuroscience 2019, 413, 123–134. [Google Scholar] [CrossRef]
- Akiba, C.; Nakajima, M.; Miyajima, M.; Ogino, I.; Miura, M.; Inoue, R.; Nakamura, E.; Kanai, F.; Tada, N.; Kunichika, M.; et al. Leucine-rich alpha2-glycoprotein overexpression in the brain contributes to memory impairment. Neurobiol. Aging 2017, 60, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Papassotiropoulos, A.; de Quervain, D.J. Failed drug discovery in psychiatry: Time for human genome-guided solutions. Trends Cogn. Sci. 2015, 19, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Cheng, S.; Xia, Y.; Peng, B.; Tang, Q.; Tu, B. Effects of subchronic benzo(a)pyrene exposure on neurotransmitter receptor gene expression in the rat hippocampus related with spatial learning and memory change. Toxicology 2011, 289, 83–90. [Google Scholar] [CrossRef]
- Berridge, M.J. The Inositol Trisphosphate/Calcium Signaling Pathway in Health and Disease. Physiol. Rev. 2016, 96, 1261–1296. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Liu, H.; He, L.; Ma, S.; Chen, X.; Wang, K.; Zhao, F.; Qi, F.; Guan, S.; Liu, Z. Prenatal chronic stress impairs the learning and memory ability via inhibition of the NO/cGMP/PKG pathway in the Hippocampus of offspring. Behav. Brain Res. 2022, 433, 114009. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition | Content |
|---|---|
| Crude protein | 66.47 ± 6.56% |
| Fat | 14.98 ± 0.20% |
| Moisture | 5.79 ± 0.14% |
| Glutamate | 16.42 ± 0.01% |
| Proline | 10.83 ± 0.01% |
| Aspartate | 10.53 ± 0.04% |
| Lysine | 9.44 ± 0.02% |
| Leucine | 8.32 ± 0.02% |
| Valine | 6.72 ± 0.01% |
| Tyrosine | 5.54 ± 0.03% |
| Phenylalanine | 5.08 ± 0.01% |
| Isoleucine | 4.69 ± 0.03% |
| Serine | 4.18 ± 0.04% |
| Threonine | 3.78 ± 0.02% |
| Histidine | 3.40 ± 0.03% |
| Arginine | 3.14 ± 0.01% |
| Alanine | 2.92 ± 0.01% |
| Methionine | 2.59 ± 0.02% |
| Glycine | 2.27 ± 0.02% |
| Cysteine | 0.14 ± 0.01% |
| Phosphatidylcholine (PC) | 1.97 ± 0.02% |
| Phosphatidylethanolamine (PE) | 2.61 ± 0.01% |
| Phosphatidylinositol (PI) | 0.14 ± 0.01% |
| Phosphatidylserine (PS) | 0.88 ± 0.03% |
| Sphingomyelin (SM) | 1.86 ± 0.02% |
| KEGG Description | DEPs Number | Pathway ID | p Value |
|---|---|---|---|
| Staphylococcus aureus infection | 5 | mmu05150 | <0.01 |
| Complement and coagulation cascades | 6 | mmu04610 | <0.01 |
| Systemic lupus erythematosus | 5 | mmu05322 | <0.01 |
| Neuroactive ligand-receptor interaction | 5 | mmu04080 | <0.01 |
| Pertussis | 4 | mmu05133 | 0.03 |
| Coronavirus disease-COVID-19 | 6 | mmu05171 | 0.03 |
| Kaposi sarcoma-associated herpesvirus infection | 6 | mmu05167 | 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Zou, X.; Feng, R.; Zhan, X.; Hong, H.; Luo, Y.; Tan, Y. Improvement of Spatial Memory and Cognitive Function in Mice via the Intervention of Milk Fat Globule Membrane. Nutrients 2023, 15, 534. https://doi.org/10.3390/nu15030534
Zhou Y, Zou X, Feng R, Zhan X, Hong H, Luo Y, Tan Y. Improvement of Spatial Memory and Cognitive Function in Mice via the Intervention of Milk Fat Globule Membrane. Nutrients. 2023; 15(3):534. https://doi.org/10.3390/nu15030534
Chicago/Turabian StyleZhou, Yongjie, Xiaoxiao Zou, Ruifang Feng, Xin Zhan, Hui Hong, Yongkang Luo, and Yuqing Tan. 2023. "Improvement of Spatial Memory and Cognitive Function in Mice via the Intervention of Milk Fat Globule Membrane" Nutrients 15, no. 3: 534. https://doi.org/10.3390/nu15030534
APA StyleZhou, Y., Zou, X., Feng, R., Zhan, X., Hong, H., Luo, Y., & Tan, Y. (2023). Improvement of Spatial Memory and Cognitive Function in Mice via the Intervention of Milk Fat Globule Membrane. Nutrients, 15(3), 534. https://doi.org/10.3390/nu15030534