

Vitamin A- and D-Deficient Diets Disrupt Intestinal Antimicrobial Peptide Defense Involving Wnt and STAT5 Signaling Pathways in Mice
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Setup, Animals, and Diets
2.2. Measurement of Intestinal Permeability
2.3. Morphological Quantification by HE Staining and Immunofluorescence Staining
2.4. Generation of Standard Plasmids
2.5. Organoid Cell Culture
2.6. Assessment of Cell Viability by Measuring MTT Reduction
2.7. Treatment of Organoids
2.8. RNA Isolation and Quantitative Real-Time PCR
2.9. Statistical Analysis
3. Results
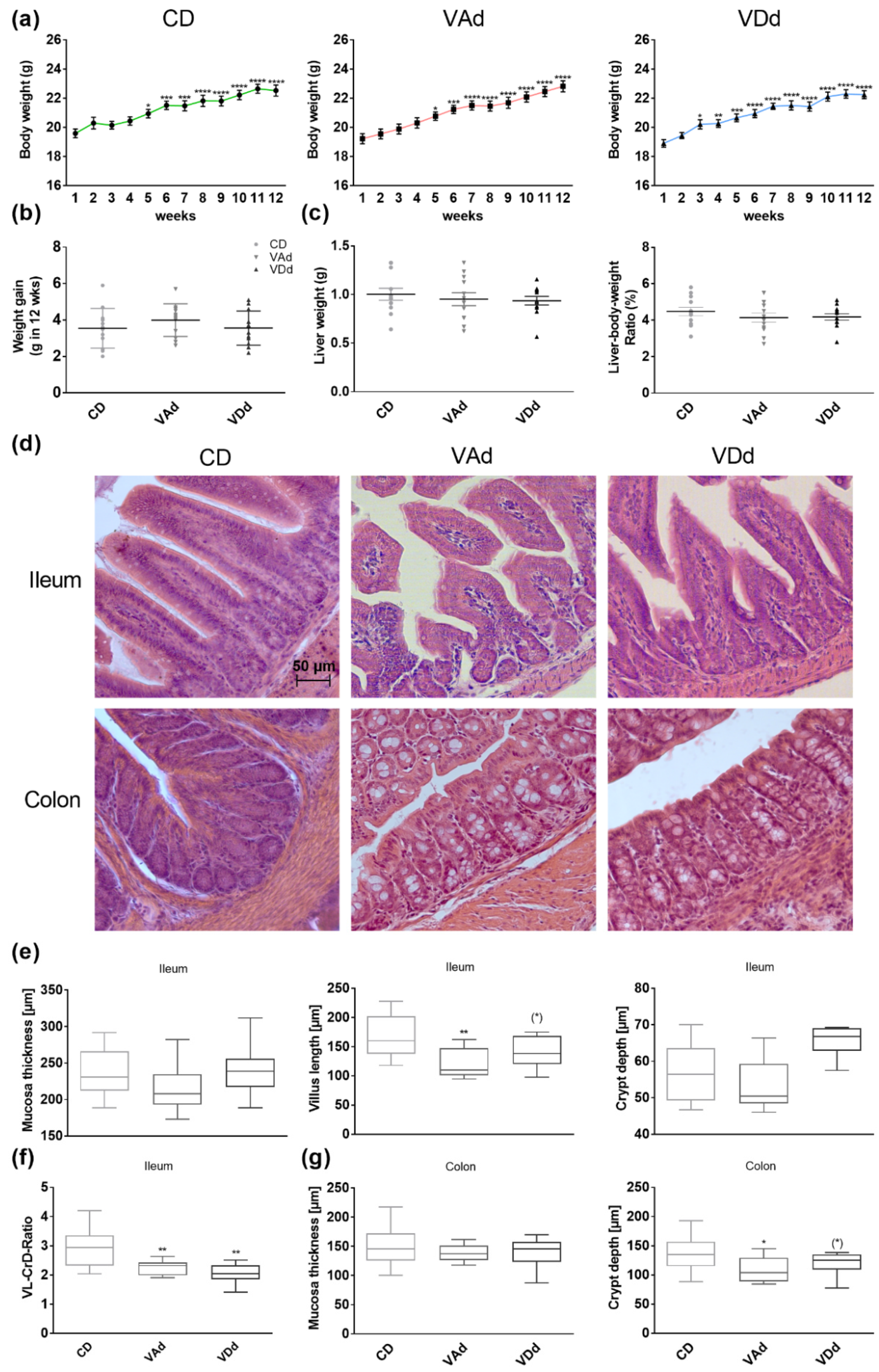
3.1. Vitamin-Deficient Diets Alter Gut Morphology in Mice
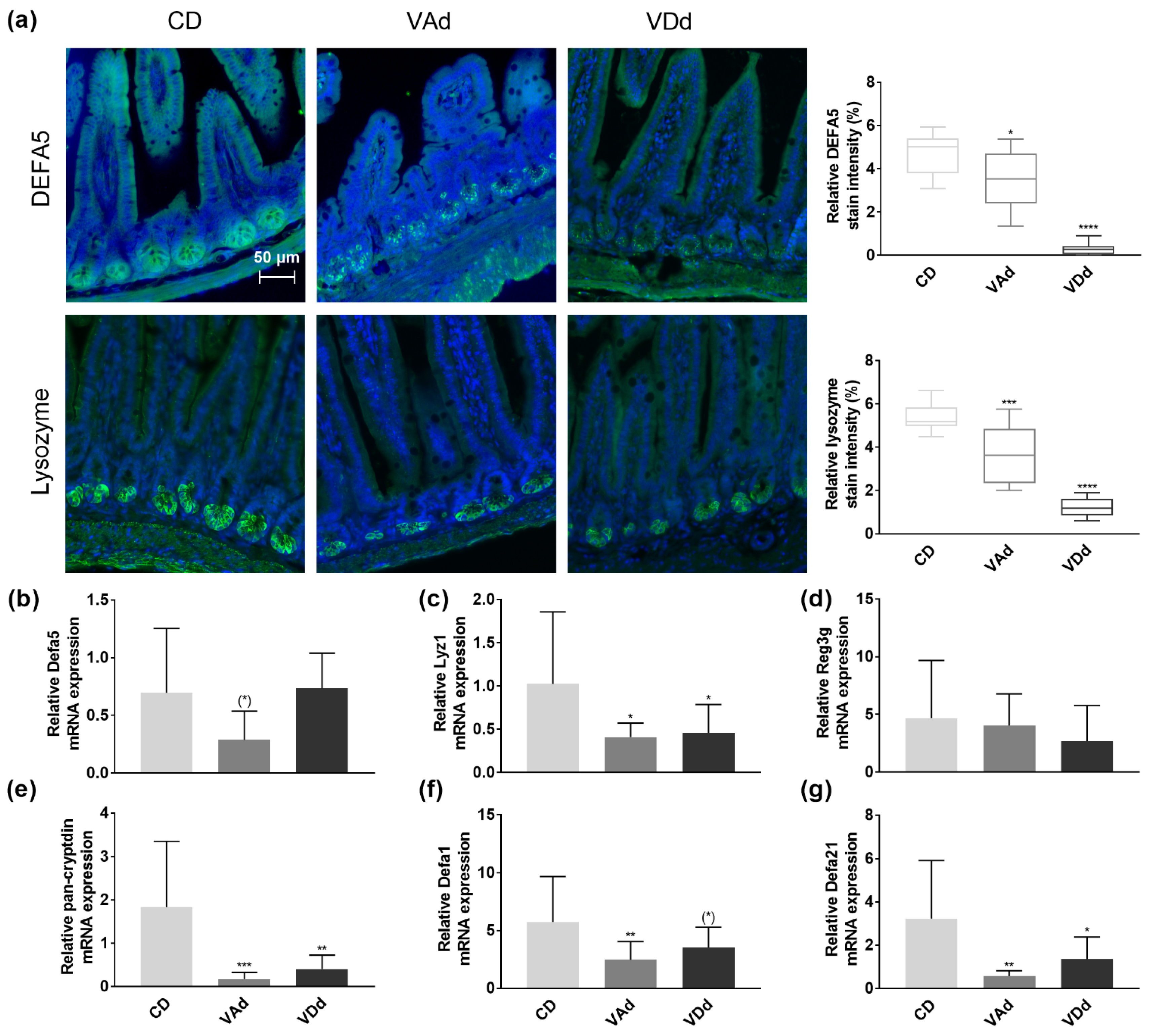
3.2. Vitamin-Deficient Fed Mice Exhibit Reduced Ileal AMP mRNA and Protein Level
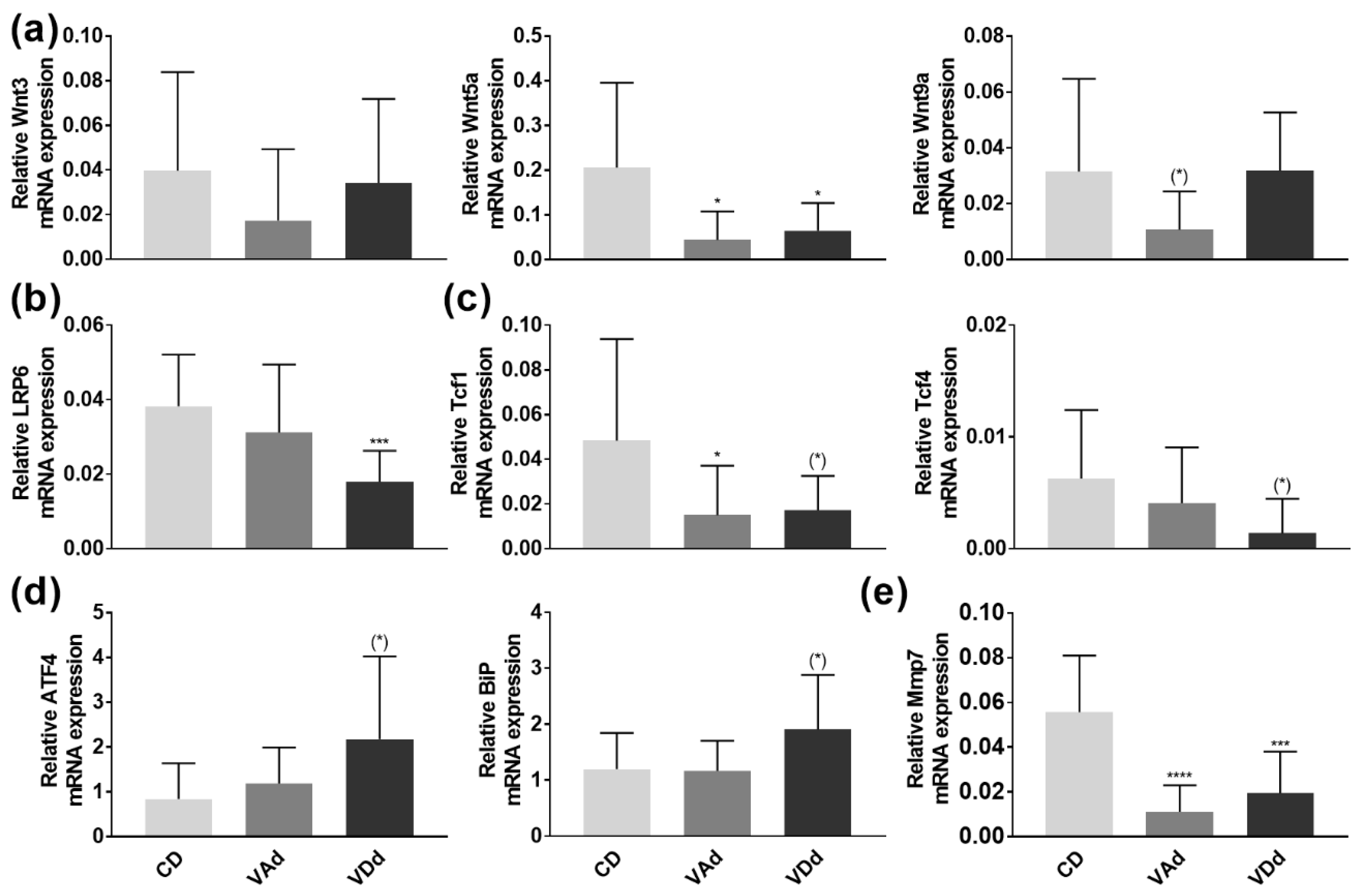
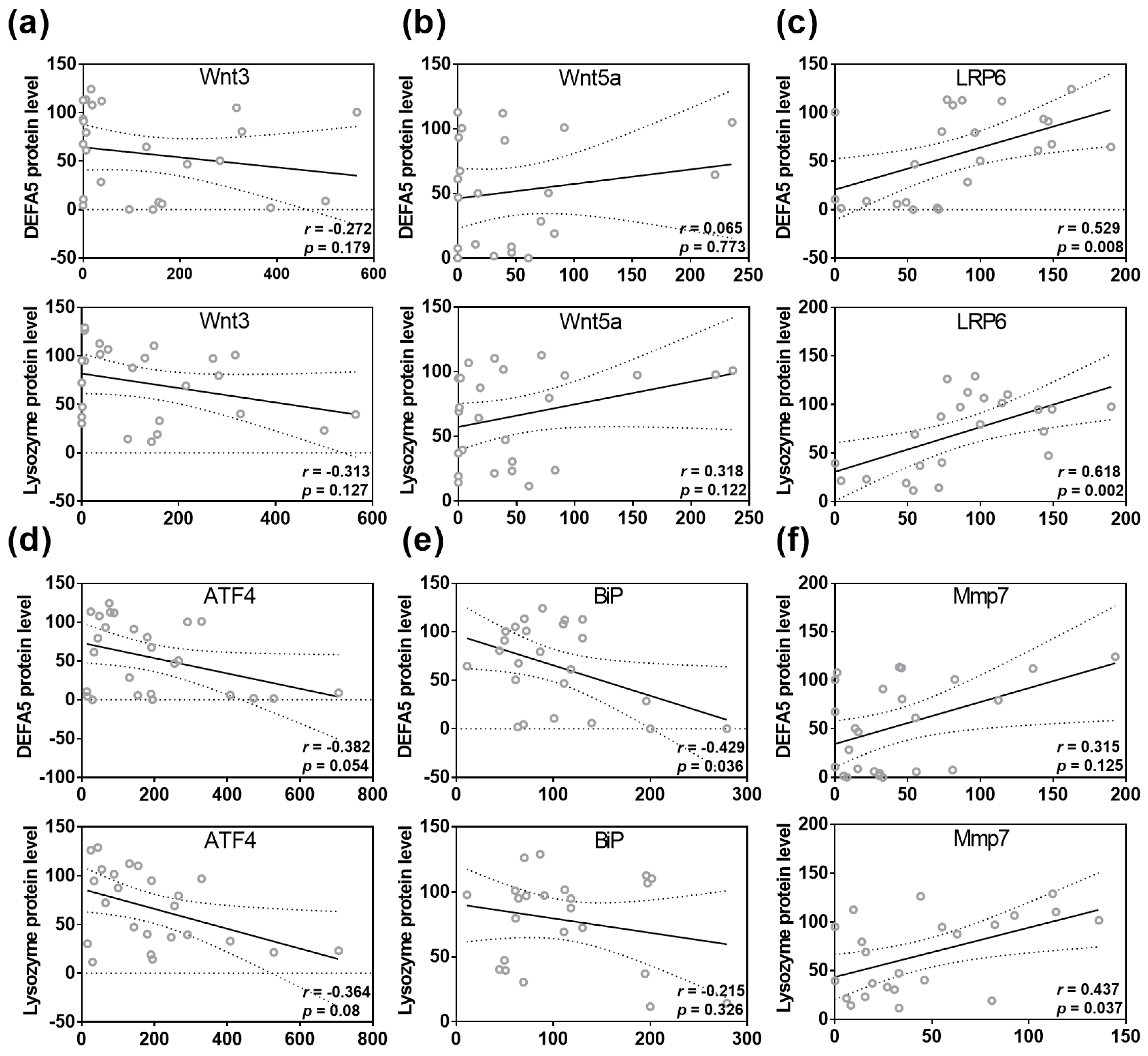
3.3. VA- and VD-Deficient Diets Deteriorate Ileal AMP Formation through Wnt, ER Stress and Mmp7
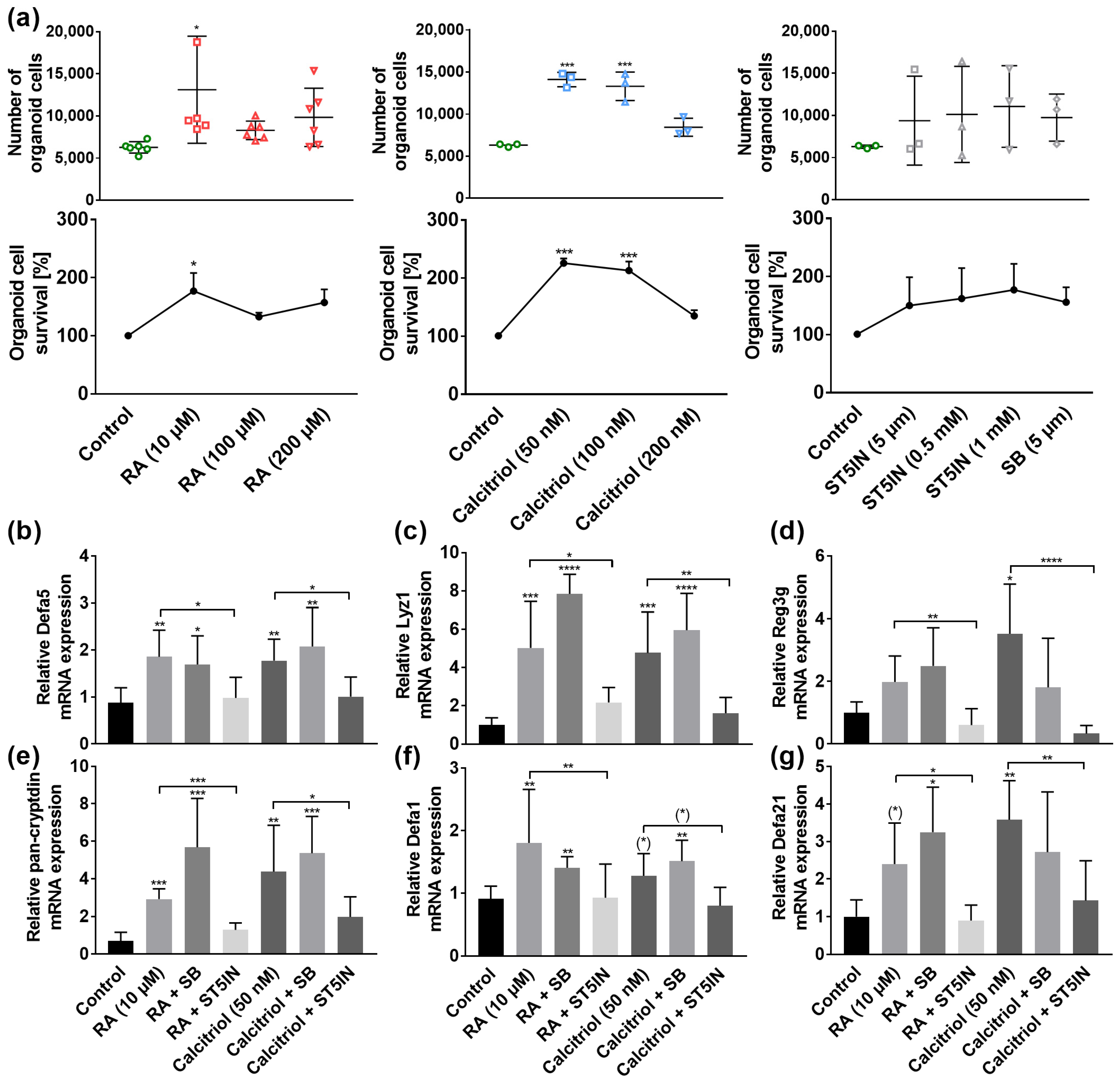
3.4. VA and VD Induce AMP Gene Expression In Vitro through STAT5 Signaling Pathway
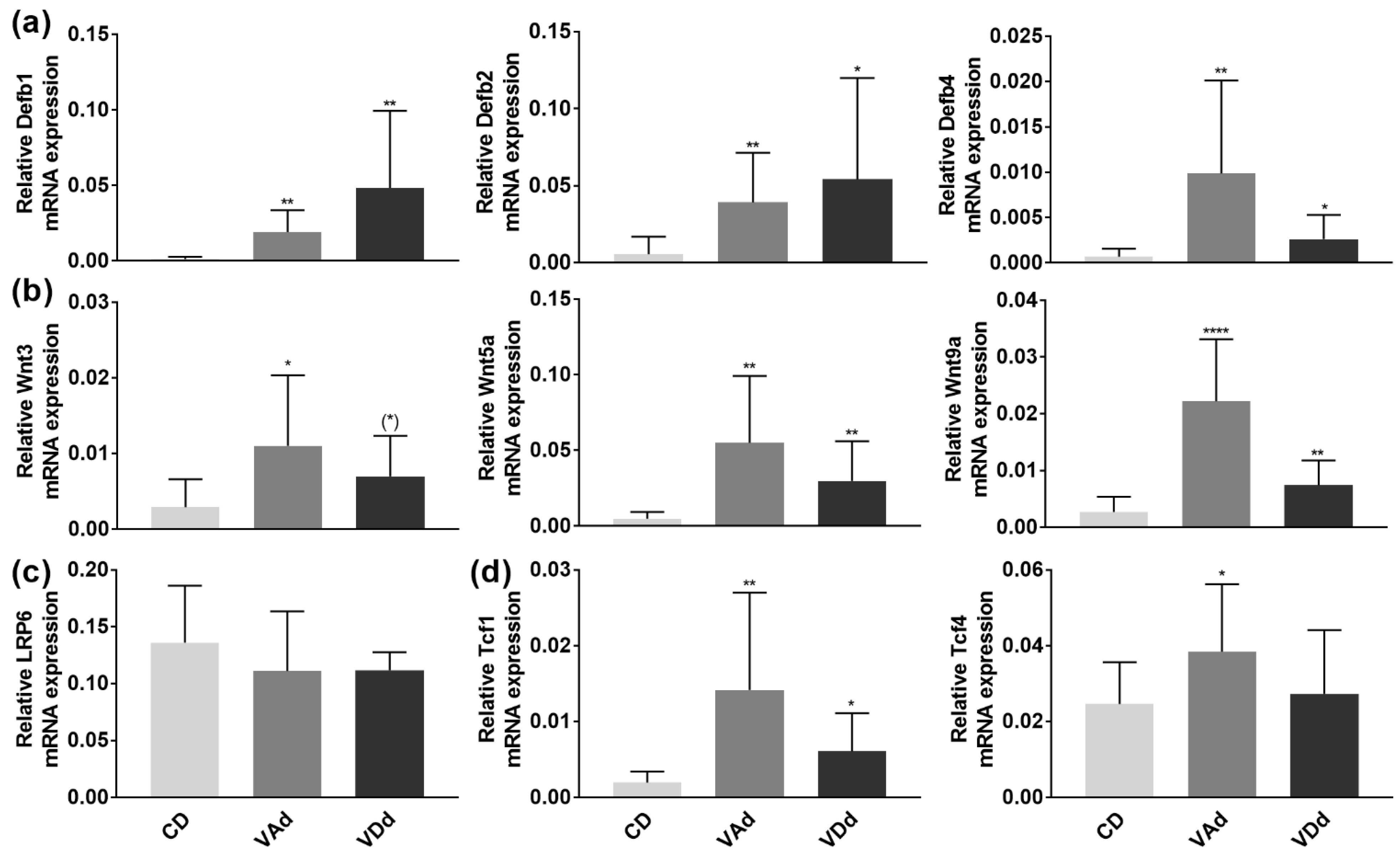
3.5. VA- and VD-Deficient Diets Increase Colonic β-Defensin Expression and Inflammation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hall, J.A.; Grainger, J.R.; Spencer, S.P.; Belkaid, Y. The Role of Retinoic Acid in Tolerance and Immunity. Immunity 2011, 35, 13–22. [Google Scholar] [CrossRef]
- Erkelens, M.N.; Goverse, G.; Konijn, T.; Molenaar, R.; Beijer, M.R.; Bossche, J.V.D.; de Goede, K.E.; Verberk, S.G.S.; de Jonge, W.J.; Haan, J.M.M.D.; et al. Intestinal Macrophages Balance Inflammatory Expression Profiles via Vitamin A and Dectin-1-Mediated Signaling. Front. Immunol. 2020, 11, 551. [Google Scholar] [CrossRef] [PubMed]
- Reifen, R.; Mor, A.; Nyska, A. Vitamin A Deficiency Aggravates Rotavirus Infection in CD-1 Mice Through Extensive Involvement of the Gut. Int. J. Vitam. Nutr. Res. 2004, 74, 355–361. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, H.; Wu, H.; Li, H.; Liu, L.; Guo, J.; Li, C.; Shih, D.Q.; Zhang, X. Protective role of 1,25(OH)2vitamin D3 in the mucosal injury and epithelial barrier disruption in DSS-induced acute colitis in mice. BMC Gastroenterol. 2012, 12, 57. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, D.N.; Clugston, R.D.; Blaner, W.S. Vitamin A metabolism: An update. Nutrients 2011, 3, 63–103. [Google Scholar] [CrossRef] [PubMed]
- Bikle, D. Nonclassic Actions of Vitamin D. J. Clin. Endocrinol. Metab. 2009, 94, 26–34. [Google Scholar] [CrossRef]
- Clemens, T.L.; Actams, J.S.; Nolan, J.M.; Holick, M.F. Measurement of circulating vitamin D in man. Clin. Chim. Acta 1982, 121, 301–308. [Google Scholar] [CrossRef]
- Chai, Z.; Lyu, Y.; Chen, Q.; Wei, C.-H.; Snyder, L.M.; Weaver, V.; Sebastian, A.; Albert, I.; Li, Q.; Cantorna, M.T.; et al. Transcriptional Profiling of the Small Intestine and the Colon Reveals Modulation of Gut Infection with Citrobacter rodentium According to the Vitamin A Status. Nutrients 2022, 14, 1563. [Google Scholar] [CrossRef]
- Levin, A.D.; Wadhera, V.; Leach, S.; Woodhead, H.J.; Lemberg, D.; Mendoza-Cruz, A.C.; Day, A.S. Vitamin D Deficiency in Children with Inflammatory Bowel Disease. Dig. Dis. Sci. 2011, 56, 830–836. [Google Scholar] [CrossRef]
- Soares-Mota, M.; Silva, T.A.; Gomes, L.M.; Pinto, M.A.; MC Mendonça, L.; Farias, M.L.F.; Nunes, T.; Ramalho, A.; Zaltman, C. High prevalence of vitamin A deficiency in Crohn’s disease patients according to serum retinol levels and the relative dose-response test. World J. Gastroenterol. 2015, 21, 1614–1620. [Google Scholar] [CrossRef]
- Chen, P.; Soares, A.M.; Lima, A.A.M.; Gamble, M.V.; Schorling, J.B.; Conway, M.; Barrett, L.J.; Blaner, W.S.; Guerrant, R.L. Association of vitamin A and zinc status with altered intestinal permeability: Analyses of cohort data from northeastern Brazil. J. Health Popul. Nutr. 2003, 21, 309–315. [Google Scholar] [PubMed]
- Ulitsky, A.; Ananthakrishnan, A.N.; Naik, A.; Skaros, S.; Zadvornova, Y.; Binion, D.G.; Issa, M. Vitamin D deficiency in patients with inflammatory bowel disease: Association with disease activity and quality of life, JPEN. J. Parenter. Enter. Nutr. 2011, 35, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Hodges, K.; Gill, R. Infectious diarrhea: Cellular and molecular mechanisms. Gut Microbes 2010, 1, 4–21. [Google Scholar] [CrossRef] [PubMed]
- Kosovac, K.; Brenmoehl, J.; Holler, E.; Falk, W.; Schoelmerich, J.; Hausmann, M.; Rogler, G. Association of the NOD2 genotype with bacterial translocation via altered cell-cell contacts in Crohn’s disease patients. Inflamm. Bowel Dis. 2010, 16, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.-D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal permeability—A new target for disease prevention and therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef]
- Damms-Machado, A.; Louis, S.; Schnitzer, A.; Volynets, V.; Rings, A.; Basrai, M.; Bischoff, S.C. Gut permeability is related to body weight, fatty liver disease, and insulin resistance in obese individuals undergoing weight reduction. Am. J. Clin. Nutr. 2017, 105, 127–135. [Google Scholar] [CrossRef]
- Kong, J.; Zhang, Z.; Musch, M.W.; Ning, G.; Sun, J.; Hart, J.; Bissonnette, M.; Li, Y.C. Novel role of the vitamin D receptor in maintaining the integrity of the intestinal mucosal barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G208–G216. [Google Scholar] [CrossRef]
- Lu, R.; Zhang, Y.-G.; Xia, Y.; Zhang, J.; Kaser, A.; Blumberg, R.; Sun, J. Paneth Cell Alertness to Pathogens Maintained by Vitamin D Receptors. Gastroenterology 2021, 160, 1269–1283. [Google Scholar] [CrossRef]
- Bischoff, S.C.; Kaden-Volynets, V.; Rosa, L.F.; Guseva, D.; Seethaler, B. Regulation of the gut barrier by carbohydrates from diet —Underlying mechanisms and possible clinical implications. Int. J. Med. Microbiol. 2021, 311, 151499. [Google Scholar] [CrossRef]
- Ouellette, A.J. Defensin-mediated innate immunity in the small intestine. Best Pract. Res. Clin. Gastroenterol. 2004, 18, 405–419. [Google Scholar] [CrossRef]
- Beisner, J.; Rosa, L.F.; Kaden-Volynets, V.; Stolzer, I.; Günther, C.; Bischoff, S.C. Prebiotic Inulin and Sodium Butyrate Attenuate Obesity-Induced Intestinal Barrier Dysfunction by Induction of Antimicrobial Peptides. Front. Immunol. 2021, 12, 678360. [Google Scholar] [CrossRef]
- Wehkamp, J.; Salzman, N.H.; Porter, E.; Nuding, S.; Weichenthal, M.; Petras, R.E.; Shen, B.; Schaeffeler, E.; Schwab, M.; Linzmeier, R.; et al. Reduced Paneth cell alpha-defensins in ileal Crohn’s disease. Proc. Natl. Acad. Sci. USA 2005, 102, 18129–18134. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.-T.; Nestel, F.P.; Bourdeau, V.; Nagai, Y.; Wang, Q.; Liao, J.; Tavera-Mendoza, L.; Lin, R.; Hanrahan, J.W.; Mader, S.; et al. Cutting Edge: 1,25-Dihydroxyvitamin D3 Is a Direct Inducer of Antimicrobial Peptide Gene Expression. J. Immunol. 2004, 173, 2909–2912. [Google Scholar] [CrossRef] [PubMed]
- Gombart, A.F.; O’Kelly, J.; Saito, T.; Koeffler, H.P. Regulation of the CAMP gene by 1,25(OH)2D3 in various tissues. J. Steroid Biochem. Mol. Biol. 2007, 103, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Gombart, A.F.; Saito, T.; Koeffler, H.P. Exaptation of an ancient Alu short interspersed element provides a highly conserved vitamin D-mediated innate immune response in humans and primates. BMC Genom. 2009, 10, 321. [Google Scholar] [CrossRef] [PubMed]
- Yuk, J.-M.; Shin, D.-M.; Lee, H.-M.; Yang, C.-S.; Jin, H.S.; Kim, K.-K.; Lee, Z.-W.; Lee, S.-H.; Kim, J.-M.; Jo, E.-K. Vitamin D3 Induces Autophagy in Human Monocytes/Macrophages via Cathelicidin. Cell Host Microbe 2009, 6, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zhang, G.; Minton, J.E.; Ross, C.R.; Blecha, F. Regulation of Cathelicidin Gene Expression: Induction by Lipopolysaccharide, Interleukin-6, Retinoic Acid, and Salmonella enterica Serovar Typhimurium Infection. Infect. Immun. 2000, 68, 5552–5558. [Google Scholar] [CrossRef]
- Harder, J.; Meyer-Hoffert, U.; Wehkamp, K.; Schwichtenberg, L.; Schröder, J.M. Differential gene induction of human beta-defensins (hBD-1, -2, -3, and -4) in keratinocytes is inhibited by retinoic acid. J. Investig. Dermatol. 2004, 123, 3–522. [Google Scholar] [CrossRef]
- Gubatan, J.; Mehigan, G.A.; Villegas, F.; Mitsuhashi, S.; Longhi, M.S.; Malvar, G.; Csizmadia, E.; Robson, S.; Moss, A.C. Cathelicidin Mediates a Protective Role of Vitamin D in Ulcerative Colitis and Human Colonic Epithelial Cells. Inflamm. Bowel Dis. 2020, 26, 885–897. [Google Scholar] [CrossRef]
- Spruss, A.; Henkel, J.; Kanuri, G.; Blank, D.; Püschel, G.P.; Bischoff, S.C.; Bergheim, I. Female Mice Are More Susceptible to Nonalcoholic Fatty Liver Disease: Sex-Specific Regulation of the Hepatic AMP-Activated Protein Kinase-Plasminogen Activator Inhibitor 1 Cascade, but Not the Hepatic Endotoxin Response. Mol. Med. 2012, 18, 1346–1355. [Google Scholar] [CrossRef]
- Volynets, V.; Reichold, A.; Bárdos, G.; Rings, A.; Bleich, A.; Bischoff, S.C. Assessment of the Intestinal Barrier with Five Different Permeability Tests in Healthy C57BL/6J and BALB/cJ Mice. Dig. Dis. Sci. 2015, 61, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Hagenlocher, Y.; Hösel, A.; Bischoff, S.C.; Lorentz, A. Cinnamon extract reduces symptoms, inflammatory mediators and mast cell markers in murine IL-10(-/-) colitis. J. Nutr. Biochem. 2016, 30, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Vries, R.G.; Snippert, H.J.; Van De Wetering, M.; Barker, N.; Stange, D.E.; Van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single Lgr5 Stem Cells Build Crypt-Villus Structures in Vitro without a Mesenchymal Niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Li, X.; Lu, D.; Liu, S.; Suo, X.; Li, Q.; Li, N. Lysozyme improves gut performance and protects against enterotoxigenic Escherichia coli infection in neonatal piglets. Vet. Res. 2018, 49, 20. [Google Scholar] [CrossRef]
- Courth, L.F.; Ostaff, M.J.; Mailänder-Sánchez, D.; Malek, N.P.; Stange, E.F.; Wehkamp, J. Crohn’s disease-derived monocytes fail to induce Paneth cell defensins. Proc. Natl. Acad. Sci. USA 2015, 112, 14000–14005. [Google Scholar] [CrossRef]
- Wehrli, M.; Dougan, S.T.; Caldwell, K.; O’Keefe, L.; Schwartz, S.; Vaizel-Ohayon, D.; Schejter, E.; Tomlinson, A.; DiNardo, S. arrow encodes an LDL-receptor-related protein essential for Wingless signalling. Nature 2000, 407, 527–530. [Google Scholar] [CrossRef]
- Guo, X.; Tang, R.; Yang, S.; Lu, Y.; Luo, J.; Liu, Z. Rutin and Its Combination With Inulin Attenuate Gut Dysbiosis, the Inflammatory Status and Endoplasmic Reticulum Stress in Paneth Cells of Obese Mice Induced by High-Fat Diet. Front. Microbiol. 2018, 9, 2651. [Google Scholar] [CrossRef]
- Wilson, C.L.; Ouellette, A.J.; Satchell, D.P.; Ayabe, T.; López-Boado, Y.S.; Stratman, J.L.; Hultgren, S.J.; Matrisian, L.M.; Parks, W.C. Regulation of Intestinal α-Defensin Activation by the Metalloproteinase Matrilysin in Innate Host Defense. Science 1999, 286, 113–117. [Google Scholar] [CrossRef]
- Ayabe, T.; Satchell, D.P.; Pesendorfer, P.; Tanabe, H.; Wilson, C.L.; Hagen, S.J.; Ouellette, A.J. Activation of Paneth Cell α-Defensins in Mouse Small Intestine. J. Biol. Chem. 2002, 277, 5219–5228. [Google Scholar] [CrossRef]
- Piskunov, A.; Rochette-Egly, C. A retinoic acid receptor RARα pool present in membrane lipid rafts forms complexes with G protein αQ to activate p38MAPK. Oncogene 2011, 31, 3333–3345. [Google Scholar] [CrossRef]
- Alsayed, Y.; Uddin, S.; Mahmud, N.; Lekmine, F.; Kalvakolanu, D.V.; Minucci, S.; Bokoch, G.; Platanias, L.C. Activation of Rac1 and the p38 Mitogen-activated Protein Kinase Pathway in Response to All-trans-retinoic Acid. J. Biol. Chem. 2001, 276, 4012–4019. [Google Scholar] [CrossRef] [PubMed]
- Noy, N. Vitamin A Transport and Cell Signaling by the Retinol-Binding Protein Receptor STRA6. In Sub-Cellular Biochemistry; Springer: Dordrecht, The Netherlands, 2016; Volume 81, pp. 77–93. [Google Scholar] [CrossRef]
- Schroeder, B.O.; Wu, Z.; Nuding, S.; Groscurth, S.; Marcinowski, M.; Beisner, J.; Buchner, J.; Schaller, M.; Stange, E.F.; Wehkamp, J. Reduction of disulphide bonds unmasks potent antimicrobial activity of human β-defensin 1. Nature 2011, 469, 419–423. [Google Scholar] [CrossRef]
- Hu, C.-Q.; Bo, Q.-L.; Chu, L.-L.; Hu, Y.-D.; Fu, L.; Wang, G.-X.; Lu, Y.; Liu, X.-J.; Wang, H.; Xu, D.-X. Vitamin D Deficiency Aggravates Hepatic Oxidative Stress and Inflammation during Chronic Alcohol-Induced Liver Injury in Mice. Oxidative Med. Cell. Longev. 2020, 2020, 5715893. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, J.; Wang, Y.; Wang, L.; Yin, Y.; Yin, L.; Yang, H.; Yin, Y. Dietary vitamin A affects growth performance, intestinal development, and functions in weaned piglets by affecting intestinal stem cells. J. Anim. Sci. 2020, 98, 2. [Google Scholar] [CrossRef] [PubMed]
- White, J.C.; Shankar, V.N.; Highland, M.; Epstein, M.L.; DeLuca, H.F.; Clagett-Dame, M. Defects in embryonic hindbrain development and fetal resorption resulting from vitamin A deficiency in the rat are prevented by feeding pharmacological levels of all-trans-retinoic acid. Proc. Natl. Acad. Sci. USA 1998, 95, 13459–13464. [Google Scholar] [CrossRef]
- See, A.W.-M.; Kaiser, M.E.; White, J.C.; Clagett-Dame, M. A nutritional model of late embryonic vitamin A deficiency produces defects in organogenesis at a high penetrance and reveals new roles for the vitamin in skeletal development. Dev. Biol. 2007, 316, 171–190. [Google Scholar] [CrossRef]
- Gloria, N.F.; Soares, N.; Brand, C.; de Oliveira, F.L.; Borojevic, R.; Teodoro, A.J. Lycopene and beta-carotene induce cell-cycle arrest and apoptosis in human breast cancer cell lines. Anticancer. Res. 2014, 34, 1377–1386. [Google Scholar]
- Wolbach, S.B.; Howe, P.R. Tissue Changes Following Deprivation of Fat-Soluble A Vitamin. J. Exp. Med. 1925, 42, 753–777. [Google Scholar] [CrossRef]
- Moore, T. Vitamin A and carotene: The absence of the liver oil vitamin A from carotene. VI. The conversion of carotene to vitamin A in vivo. Biochem. J. 1930, 24, 692–702. [Google Scholar] [CrossRef]
- Shen, Q.; Wang, X.; Bai, H.; Tan, X.; Liu, X. Effects of high-dose all-trans retinoic acid on longitudinal bone growth of young rats. Growth Horm. IGF Res. 2022, 62, 101446. [Google Scholar] [CrossRef]
- Yeung, C.-Y.; Chiau, J.-S.C.; Cheng, M.-L.; Chan, W.-T.; Jiang, C.-B.; Chang, S.-W.; Liu, C.-Y.; Chang, C.-W.; Lee, H.-C. Effects of Vitamin D-Deficient Diet on Intestinal Epithelial Integrity and Zonulin Expression in a C57BL/6 Mouse Model. Front. Med. 2021, 8, 649818. [Google Scholar] [CrossRef] [PubMed]
- Assa, A.; Vong, L.; Pinnell, L.J.; Avitzur, N.; Johnson-Henry, K.C.; Sherman, P.M. Vitamin D Deficiency Promotes Epithelial Barrier Dysfunction and Intestinal Inflammation. J. Infect. Dis. 2014, 210, 1296–1305. [Google Scholar] [CrossRef] [PubMed]
- Stojanović, O.; Altirriba, J.; Rigo, D.; Spiljar, M.; Evrard, E.; Roska, B.; Fabbiano, S.; Zamboni, N.; Maechler, P.; Rohner-Jeanrenaud, F.; et al. Dietary excess regulates absorption and surface of gut epithelium through intestinal PPARα. Nat. Commun. 2021, 12, 7031. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yuan, Y.; Tao, Y.; Wang, W. Effects of vitamin A deficiency on mucosal immunity and response to intestinal infection in rats. Nutrition 2011, 27, 227–232. [Google Scholar] [CrossRef]
- Wang, P.-F.; Yao, D.-H.; Hu, Y.-Y.; Li, Y. Vitamin D Improves Intestinal Barrier Function in Cirrhosis Rats by Upregulating Heme Oxygenase-1 Expression. Biomol. Ther. 2019, 27, 222–230. [Google Scholar] [CrossRef]
- Li, W.; Lin, Y.; Luo, Y.; Wang, Y.; Lu, Y.; Li, Y.; Guo, H. Vitamin D Receptor Protects against Radiation-Induced Intestinal Injury in Mice via Inhibition of Intestinal Crypt Stem/Progenitor Cell Apoptosis. Nutrients 2021, 13, 2910. [Google Scholar] [CrossRef]
- Amit-Romach, E.; Uni, Z.; Cheled, S.; Berkovich, Z.; Reifen, R. Bacterial population and innate immunity-related genes in rat gastrointestinal tract are altered by vitamin A-deficient diet. J. Nutr. Biochem. 2009, 20, 70–77. [Google Scholar] [CrossRef]
- Su, D.; Nie, Y.; Zhu, A.; Chen, Z.; Wu, P.; Zhang, L.; Luo, M.; Sun, Q.; Cai, L.; Lai, Y.; et al. Vitamin D Signaling through Induction of Paneth Cell Defensins Maintains Gut Microbiota and Improves Metabolic Disorders and Hepatic Steatosis in Animal Models. Front. Physiol. 2016, 7, 498. [Google Scholar] [CrossRef]
- Lee, P.; Hsieh, Y.; Huo, T.; Yang, U.; Lin, C.; Li, C.; Huang, Y.; Hou, M.; Lin, H.; Lee, K. Active Vitamin D 3 Treatment Attenuated Bacterial Translocation via Improving Intestinal Barriers in Cirrhotic Rats. Mol. Nutr. Food Res. 2020, 65, e2000937. [Google Scholar] [CrossRef]
- Filteau, S.M.; Rollins, N.C.; Coutsoudis, A.; Sullivan, K.R.; Willumsen, J.F.; Tomkins, A.M. The effect of antenatal vitamin A and beta-carotene supplementation on gut integrity of infants of HIV-infected South African women. J. Pediatr. Gastroenterol. Nutr. 2001, 32, 464–470. [Google Scholar] [CrossRef]
- Wu, P.; Zhang, R.; Luo, M.; Zhang, T.; Pan, L.; Xu, S.; Pan, L.; Ren, F.; Ji, C.; Hu, R.; et al. Impaired 25-hydroxylation of vitamin D in liver injury suppresses intestinal Paneth cell defensins, leading to gut dysbiosis and liver fibrogenesis. Am. J. Physiol. Liver Physiol. 2020, 319, G685–G695. [Google Scholar] [CrossRef]
- Goswami, S.; Angkasekwinai, P.; Shan, M.; Greenlee, K.J.; Barranco, W.T.; Polikepahad, S.; Seryshev, A.; Song, L.-Z.; Redding, D.; Singh, B.; et al. Divergent functions for airway epithelial matrix metalloproteinase 7 and retinoic acid in experimental asthma. Nat. Immunol. 2009, 10, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Seifart, C.; Muyal, J.P.; Plagens, A.; Yildirim, A.; Kohse, K.; Grau, V.; Sandu, S.; Reinke, C.; Tschernig, T.; Vogelmeier, C.; et al. All-trans retinoic acid results in irregular repair of septa and fails to inhibit proinflammatory macrophages. Eur. Respir. J. 2011, 38, 425–439. [Google Scholar] [CrossRef] [PubMed]
- Adachi, Y.; Itoh, F.; Yamamoto, H.; Iku, S.; Matsuno, K.; Arimura, Y.; Imai, K. Retinoic acids reduce matrilysin (matrix metalloproteinase 7) and inhibit tumor cell invasion in human colon cancer. Tumour Biol. 2001, 22, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Park, E.Y.; Wilder, E.T.; Lane, M.A. Retinol Inhibits the Invasion of Retinoic Acid–Resistant Colon Cancer Cells In Vitro and Decreases Matrix Metalloproteinase mRNA, Protein, and Activity Levels. Nutr. Cancer 2007, 57, 66–77. [Google Scholar] [CrossRef]
- Gröschel, C.; Aggarwal, A.; Tennakoon, S.; Höbaus, J.; Prinz-Wohlgenannt, M.; Marian, B.; Heffeter, P.; Berger, W.; Kállay, E. Effect of 1,25-dihydroxyvitamin D3 on the Wnt pathway in non-malignant colonic cells. J. Steroid Biochem. Mol. Biol. 2016, 155, 224–2300. [Google Scholar] [CrossRef] [PubMed]
- Pálmer, H.G.; Sancho, J.M.G.; Espada, J.; Berciano, M.T.; Puig, I.; Baulida, J.; Quintanilla, M.; Cano, A.; De Herreros, A.G.; Lafarga, M.; et al. Vitamin D3 promotes the differentiation of colon carcinoma cells by the induction of E-cadherin and the inhibition of β-catenin signaling. J. Cell Biol. 2001, 154, 369–388. [Google Scholar] [CrossRef]
- Zhang, T.; Ahn, K.; Emerick, B.; Modarai, S.R.; Opdenaker, L.M.; Palazzo, J.; Schleiniger, G.; Fields, J.Z.; Boman, B.M. APC mutations in human colon lead to decreased neuroendocrine maturation of ALDH+ stem cells that alters GLP-2 and SST feedback signaling: Clue to a link between WNT and retinoic acid signalling in colon cancer development. PLoS ONE 2020, 15, e0239601. [Google Scholar] [CrossRef]
- Liao, X.-D.; Zhou, C.-H.; Zhang, J.; Shen, J.-L.; Wang, Y.-J.; Jin, Y.-C.; Li, S.-L. Effect of all-trans retinoic acid on casein and fatty acid synthesis in MAC-T cells. Asian-Australas. J. Anim. Sci. 2020, 33, 1012–1022. [Google Scholar] [CrossRef]
- Si, J.; Collins, S.J. IL-3–induced enhancement of retinoic acid receptor activity is mediated through Stat5, which physically associates with retinoic acid receptors in an IL-3—Dependent manner. Blood 2002, 100, 4401–4409. [Google Scholar] [CrossRef]
- Yang, M.; Yang, B.; Gan, H.; Li, X.; Xu, J.; Yu, J.; Gao, L.; Li, F. Anti-inflammatory effect of 1,25-dihydroxyvitamin D3 is associated with crosstalk between signal transducer and activator of transcription 5 and the vitamin D receptor in human monocytes. Exp. Ther. Med. 2015, 9, 1739–1744. [Google Scholar] [CrossRef]
- Yang, M.; Shen, Z.; Chen, D.; Gan, H.; Shen, Q.; Yang, B.; Du, X. Effects of 1,25-(OH)(2)D(3) on the expressions of vitamin D receptor, STAT5 and cytoskeletal rearrangement in human monocytes incubated with sera from type 2 diabetes patients and diabetic nephropathy patients with uremia. Inflamm. Res. 2012, 61, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Xie, X.; Zhao, Y.; Zhou, M.; Yang, Y.; Zhang, X. Calcitriol attenuates renal tubular epithelial cells apoptosis via inhibiting p38MAPK signaling in diabetic nephropathy. Acta Diabetol. 2020, 57, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- Timoneda, J.; Rodríguez-Fernández, L.; Zaragozá, R.; Marín, M.P.; Cabezuelo, M.T.; Torres, L.; Viña, J.R.; Barber, T. Vitamin A deficiency and the lung. Nutrients 2018, 10, 1132. [Google Scholar] [CrossRef]
- Harder, J.; Bartels, J.; Christophers, E.; Schröder, J.-M. A peptide antibiotic from human skin. Nature 1997, 387, 861. [Google Scholar] [CrossRef] [PubMed]
- Tarver, A.P.; Clark, D.P.; Diamond, G.; Russell, J.P.; Erdjument-Bromage, H.; Tempst, P.; Cohen, K.S.; Jones, D.E.; Sweeney, R.W.; Wines, M.; et al. Enteric β-Defensin: Molecular Cloning and Characterization of a Gene with Inducible Intestinal Epithelial Cell Expression Associated with Cryptosporidium parvum Infection. Infect. Immun. 1998, 66, 1045–1056. [Google Scholar] [CrossRef]
- Otte, J.-M.; Werner, I.; Brand, S.; Chromik, A.M.; Schmitz, F.; Kleine, M.; Schmidt, W.E. Human beta defensin 2 promotes intestinal wound healing in vitro. J. Cell. Biochem. 2008, 104, 2286–2297. [Google Scholar] [CrossRef]
- Diamond, G.; Russell, J.P.; Bevins, C.L. Inducible expression of an antibiotic peptide gene in lipopolysaccharide-challenged tracheal epithelial cells. Proc. Natl. Acad. Sci. USA 1996, 93, 5156–5160. [Google Scholar] [CrossRef]
- Ooi, J.H.; Li, Y.; Rogers, C.J.; Cantorna, M.T. Vitamin D Regulates the Gut Microbiome and Protects Mice from Dextran Sodium Sulfate-Induced Colitis. J. Nutr. 2013, 143, 1679–1686. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, Y.-G.; Lu, R.; Xia, Y.; Zhou, D.; Petrof, E.O.; Claud, E.C.; Chen, D.; Chang, E.B.; Carmeliet, G.; et al. Intestinal epithelial vitamin D receptor deletion leads to defective autophagy in colitis. Gut 2014, 64, 1082–1094. [Google Scholar] [CrossRef]
- Wang, F.; Graham, W.V.; Wang, Y.; Witkowski, E.D.; Schwarz, B.T.; Turner, J.R. Interferon-γ and Tumor Necrosis Factor-α Synergize to Induce Intestinal Epithelial Barrier Dysfunction by Up-Regulating Myosin Light Chain Kinase Expression. Am. J. Pathol. 2005, 166, 409–419. [Google Scholar] [CrossRef]
- Capaldo, C.T.; Nusrat, A. Cytokine regulation of tight junctions. Biochim. Biophys. Acta Biomembr. 2009, 1788, 864–871. [Google Scholar] [CrossRef] [PubMed]
- Pinto, D.; Gregorieff, A.; Begthel, H.; Clevers, H. Canonical Wnt signals are essential for homeostasis of the intestinal epithelium. Genes Dev. 2003, 17, 1709–1713. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.-H.; Lafita-Navarro, M.C.; Zacharias, L.; Borenstein-Auerbach, N.; Kim, M.; Barnes, S.; Kim, J.; Shay, J.; DeBerardinis, R.J.; Conacci-Sorrell, M. Induction of LEF1 by MYC activates the WNT pathway and maintains cell proliferation. Cell Commun. Signal. 2019, 17, 129. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Huang, J.; Lou, K.; Ouyang, H. Long noncoding RNASEH1-AS1 exacerbates the progression of non-small cell lung cancer by acting as a ceRNA to regulate microRNA-516a-5p/FOXK1 and thereby activating the Wnt/β-catenin signaling pathway. Cancer Med. 2022, 7, 1589–1604. [Google Scholar] [CrossRef]
- Xiong, X.; Hasani, S.; Young, L.E.A.; Rivas, D.R.; Skaggs, A.T.; Martinez, R.; Wang, C.; Weiss, H.L.; Gentry, M.S.; Sun, R.C.; et al. Activation of Drp1 promotes fatty acids-induced metabolic reprograming to potentiate Wnt signaling in colon cancer. Cell Death Differ. 2022, 29, 1913–1927. [Google Scholar] [CrossRef]
- Sana, S.; Kayani, M.A. Role of Vitamin D deficiency and mRNA expression of VDR and RXR in haematological cancers. Mol. Biol. Rep. 2021, 48, 4431–4439. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CD, Ssniff ® EF R/M Control | Ssniff® EF R/M Vitamin-A-Free Vitamin A < 120 IE | Ssniff® EF R/M Vitamin-D-Free Vitamin D < 5 IE | |
|---|---|---|---|
| Diet specification | CD | VAd | VDd |
| ME [MJ/kg] | 18.0 | 18.0 | 18.0 |
| Carbohydrates [kJ%] | 61 | 61 | 61 |
| Mono- and Disaccharides [%] | 10.8 | 10.8 | 12.9 |
| Protein [kJ%] | 30 | 30 | 30 |
| Fat [kJ%] | 9 | 9 | 9 |
| Vitamin A [IE] | 15,000 | <120 | 15,000 |
| Vitamin D [IE] | 1500 | 1500 | <5 |
| Minerals [%] | 2.9 | 2.9 | 1.63 |
| Sodium [%] | 0.19 | 0.19 | 0.19 |
| Starch [%] | 46.8 | 46.8 | 46.8 |
| Crude ash [%] | 5.6 | 5.6 | 4.0 |
| Crude fiber [%] | 5.0 | 5.0 | 5.0 |
| Crude protein [%] | 20.8 | 20.8 | 20.8 |
| Crude fat [%] | 4.2 | 4.2 | 4.2 |
| Treatment | PPARG | NOD2 | Tnfα | Myd88 | IL6 | IL1β |
|---|---|---|---|---|---|---|
| CD | 0.086 ± 0.011 | 0.003 ± 0.0001 | 1.14 ± 0.3 | 3.1 ± 1.3 | 0.61 ± 0.34 | 0.8 ± 0.22 |
| VAd | 0.124 ± 0.18 * | 0.015 ± 0.003 ** | 7.04 ± 1.73 ** | 16.64 ± 6.22 * | 15.77 ± 4.7 ** | 8.61 ± 2.91 * |
| VDd | 0.083 ± 0.017 | 0.01 ± 0.003 * | 8.11 ± 1.68 *** | 27.54 ± 10.02 * | 8.28 ± 3.5 * | 3.14 ± 1.83 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filipe Rosa, L.; Petersen, P.P.; Görtz, L.F.; Stolzer, I.; Kaden-Volynets, V.; Günther, C.; Bischoff, S.C. Vitamin A- and D-Deficient Diets Disrupt Intestinal Antimicrobial Peptide Defense Involving Wnt and STAT5 Signaling Pathways in Mice. Nutrients 2023, 15, 376. https://doi.org/10.3390/nu15020376
Filipe Rosa L, Petersen PP, Görtz LF, Stolzer I, Kaden-Volynets V, Günther C, Bischoff SC. Vitamin A- and D-Deficient Diets Disrupt Intestinal Antimicrobial Peptide Defense Involving Wnt and STAT5 Signaling Pathways in Mice. Nutrients. 2023; 15(2):376. https://doi.org/10.3390/nu15020376
Chicago/Turabian StyleFilipe Rosa, Louisa, Patricia P. Petersen, Lisa F. Görtz, Iris Stolzer, Valentina Kaden-Volynets, Claudia Günther, and Stephan C. Bischoff. 2023. "Vitamin A- and D-Deficient Diets Disrupt Intestinal Antimicrobial Peptide Defense Involving Wnt and STAT5 Signaling Pathways in Mice" Nutrients 15, no. 2: 376. https://doi.org/10.3390/nu15020376
APA StyleFilipe Rosa, L., Petersen, P. P., Görtz, L. F., Stolzer, I., Kaden-Volynets, V., Günther, C., & Bischoff, S. C. (2023). Vitamin A- and D-Deficient Diets Disrupt Intestinal Antimicrobial Peptide Defense Involving Wnt and STAT5 Signaling Pathways in Mice. Nutrients, 15(2), 376. https://doi.org/10.3390/nu15020376