

The Role of One-Carbon Metabolism in Healthy Brain Aging



{kind=link}
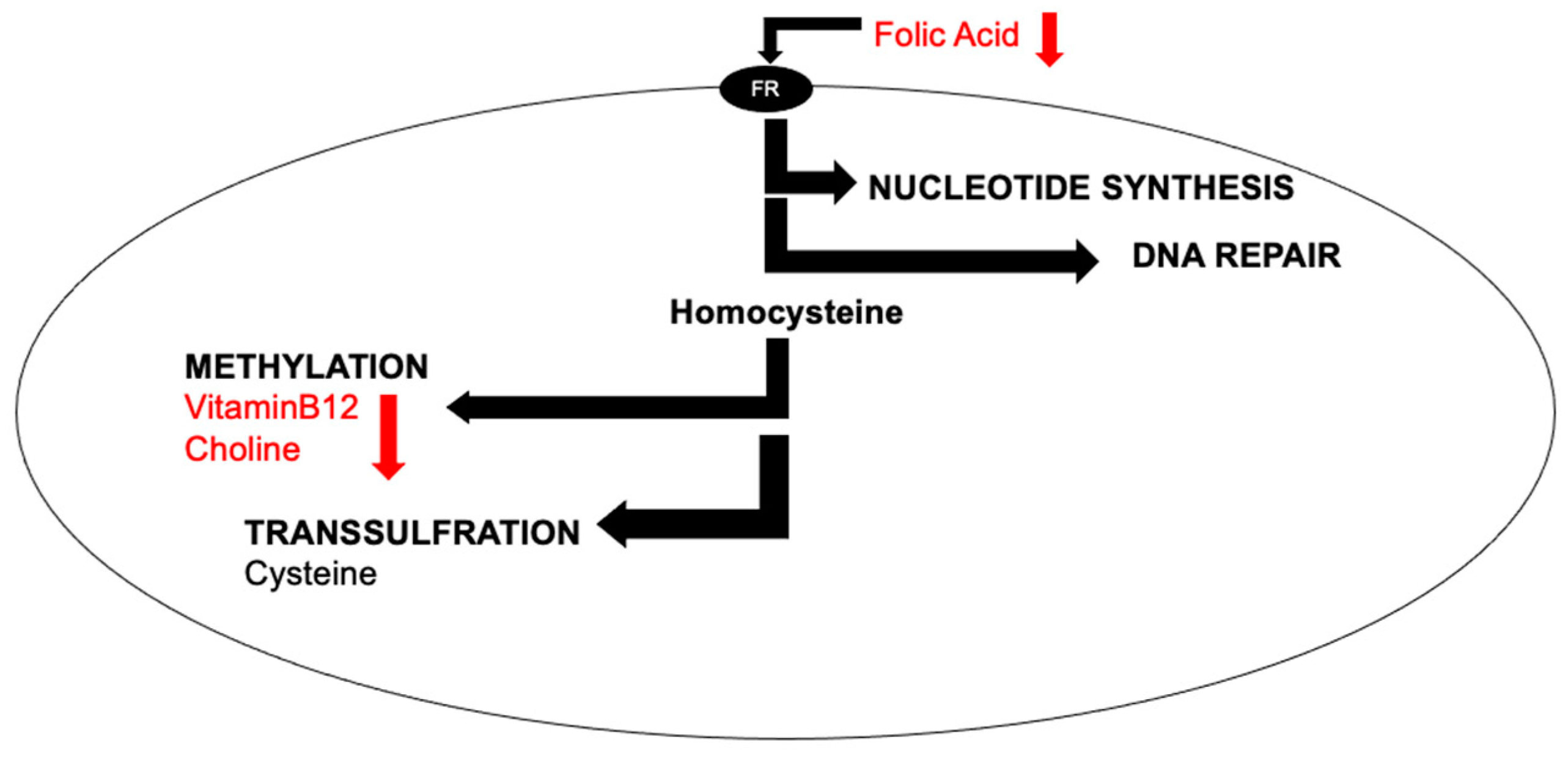
Abstract
:1. Introduction
2. Folic Acid
2.1. Functional Role of Folic Acid
2.2. Folic Acid’s Implications in Neurodegeneration
2.3. Folic Acid Interactions with Vitamin B12
3. Vitamin B12
4. Choline
5. Preclinical Studies of Folic, Choline, and Vitamin B12 Supplementation Using Model Systems
5.1. Brain Metabolism Is Affected by Folate and Vitamin B12 Status
5.2. Mechanisms of Choline Supplementation
6. Future Directions and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ageing and Health. Available online: https://www.who.int/news-room/fact-sheets/detail/ageing-and-health (accessed on 28 July 2023).
- Rudnicka, E.; Napierała, P.; Podfigurna, A.; Męczekalski, B.; Smolarczyk, R.; Grymowicz, M. The World Health Organization (WHO) Approach to Healthy Ageing. Maturitas 2020, 139, 6–11. [Google Scholar] [CrossRef]
- Clare, C.E.; Brassington, A.H.; Kwong, W.Y.; Sinclair, K.D. One-Carbon Metabolism: Linking Nutritional Biochemistry to Epigenetic Programming of Long-Term Development. Annu. Rev. Anim. Biosci. 2019, 7, 263–287. [Google Scholar] [CrossRef] [PubMed]
- Shiraki, N.; Shiraki, Y.; Tsuyama, T.; Obata, F.; Miura, M.; Nagae, G.; Aburatani, H.; Kume, K.; Endo, F.; Kume, S. Methionine Metabolism Regulates Maintenance and Differentiation of Human Pluripotent Stem Cells. Cell Metab. 2014, 19, 780–794. [Google Scholar] [CrossRef] [PubMed]
- Seshadri, S. Homocysteine and the Risk of Dementia. Clin. Chem. 2012, 58, 1059–1060. [Google Scholar] [CrossRef]
- Spence, J.D. Nutrition and Risk of Stroke. Nutrients 2019, 11, 647. [Google Scholar] [CrossRef]
- Castro, R.; Rivera, I.; Blom, H.J.; Jakobs, C.; de Almeida, I.T. Homocysteine Metabolism, Hyperhomocysteinaemia and Vascular Disease: An Overview. J. Inherit. Metab. Dis. 2006, 29, 3–20. [Google Scholar] [CrossRef]
- Osterhues, A.; Ali, N.S.; Michels, K.B. The Role of Folic Acid Fortification in Neural Tube Defects: A Review. Crit. Rev. Food Sci. Nutr. 2013, 53, 1180–1190. [Google Scholar] [CrossRef]
- Smith, A.D.; Refsum, H. Homocysteine, B Vitamins, and Cognitive Impairment. Annu. Rev. Nutr. 2016, 36, 211–239. [Google Scholar] [CrossRef]
- Yahn, G.; Abato, J.; Jadavji, N. Role of Vitamin B12 Deficiency in Ischemic Stroke Risk and Outcome. Neural Regen. Res. 2021, 16, 470. [Google Scholar] [CrossRef]
- Ahmed, S.; Bogiatzi, C.; Hackam, D.G.; Rutledge, A.C.; Sposato, L.A.; Khaw, A.; Mandzia, J.; Azarpazhoo, M.R.; Hachinski, V.; Spence, J.D. Vitamin B 12 Deficiency and Hyperhomocysteinaemia in Outpatients with Stroke or Transient Ischaemic Attack: A Cohort Study at an Academic Medical Centre. BMJ Open 2019, 9, e026564. [Google Scholar] [CrossRef]
- Smith, A.D.; Refsum, H.; Bottiglieri, T.; Fenech, M.; Hooshmand, B.; McCaddon, A.; Miller, J.W.; Rosenberg, I.H.; Obeid, R. Homocysteine and Dementia: An International Consensus Statement. J. Alzheimers Dis. 2018, 62, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Kaye, A.D.; Jeha, G.M.; Pham, A.D.; Fuller, M.C.; Lerner, Z.I.; Sibley, G.T.; Cornett, E.M.; Urits, I.; Viswanath, O.; Kevil, C.G. Folic Acid Supplementation in Patients with Elevated Homocysteine Levels. Adv. Ther. 2020, 37, 4149–4164. [Google Scholar] [CrossRef]
- Zhang, R.-S.; Tang, L.; Zhang, Y.; Shi, X.-L.; Shu, J.; Wang, L.; Zhang, X.; Xu, Y.-P.; Zou, J.-F.; Wang, R.; et al. Effect of Folic Acid Supplementation on the Change of Plasma S-Adenosylhomocysteine Level in Chinese Hypertensive Patients: A Randomized, Double-Blind, Controlled Clinical Trial. J. Clin. Biochem. Nutr. 2022, 71, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Nkemjika, S.; Ifebi, E.; Cowan, L.T.; Chun-Hai Fung, I.; Twum, F.; Liu, F.; Zhang, J. Association between Serum Folate and Cardiovascular Deaths among Adults with Hypertension. Eur. J. Clin. Nutr. 2020, 74, 970–978. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhu, X.; Guan, G.; Zhang, Y.; Hui, R.; Xing, Y.; Wang, J.; Zhu, L. Non-Linear Associations of Serum and Red Blood Cell Folate with Risk of Cardiovascular and All-Cause Mortality in Hypertensive Adults. Hypertens. Res. Off. J. Jpn. Soc. Hypertens. 2023, 46, 1504–1515. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhou, D.; Zhang, D.; Zhao, J.; Li, W.; Sun, Y.; Chen, Y.; Liu, H.; Wilson, J.X.; Qian, Z.; et al. Folic Acid Inhibits Aging-Induced Telomere Attrition and Apoptosis in Astrocytes In Vivo and In Vitro. Cereb. Cortex 2022, 32, 286–297. [Google Scholar] [CrossRef]
- Robinson, N.; Grabowski, P.; Rehman, I. Alzheimer’s Disease Pathogenesis: Is There a Role for Folate? Mech. Ageing Dev. 2018, 174, 86–94. [Google Scholar] [CrossRef]
- Smith, A.D. The Worldwide Challenge of the Dementias: A Role for B Vitamins and Homocysteine? Food Nutr. Bull. 2008, 29, S143–S172. [Google Scholar] [CrossRef]
- Locasale, J.W. Serine, Glycine and One-Carbon Units: Cancer Metabolism in Full Circle. Nat. Rev. Cancer 2013, 13, 572–583. [Google Scholar] [CrossRef]
- Chen, H.; Liu, S.; Ji, L.; Wu, T.; Ji, Y.; Zhou, Y.; Zheng, M.; Zhang, M.; Xu, W.; Huang, G. Folic Acid Supplementation Mitigates Alzheimer’s Disease by Reducing Inflammation: A Randomized Controlled Trial. Mediators Inflamm. 2016, 2016, 5912146. [Google Scholar] [CrossRef]
- Chen, H.; Liu, S.; Ge, B.; Zhou, D.; Li, M.; Li, W.; Ma, F.; Liu, Z.; Ji, Y.; Huang, G. Effects of Folic Acid and Vitamin B12 Supplementation on Cognitive Impairment and Inflammation in Patients with Alzheimer’s Disease: A Randomized, Single-Blinded, Placebo-Controlled Trial. J. Prev. Alzheimers Dis. 2021, 8, 249–256. [Google Scholar] [CrossRef]
- Christen, W.G.; Glynn, R.J.; Chew, E.Y.; Albert, C.M.; Manson, J.E. Folic Acid, Vitamin B6, and Vitamin B12 in Combination and Age-Related Macular Degeneration in a Randomized Trial of Women. Arch. Intern. Med. 2009, 169, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Merle, B.M.J.; Barthes, S.; Féart, C.; Cougnard-Grégoire, A.; Korobelnik, J.-F.; Rougier, M.-B.; Delyfer, M.-N.; Delcourt, C. B Vitamins and Incidence of Advanced Age-Related Macular Degeneration: The Alienor Study. Nutrients 2022, 14, 2821. [Google Scholar] [CrossRef]
- Deng, Y.; Wang, D.; Wang, K.; Kwok, T. High Serum Folate Is Associated with Brain Atrophy in Older Diabetic People with Vitamin B12 Deficiency. J. Nutr. Health Aging 2017, 21, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Evans, D.A.; Bienias, J.L.; Tangney, C.C.; Hebert, L.E.; Scherr, P.A.; Schneider, J.A. Dietary Folate and Vitamin B12 Intake and Cognitive Decline among Community-Dwelling Older Persons. Arch. Neurol. 2005, 62, 641–645. [Google Scholar] [CrossRef]
- Mills, J.L.; Molloy, A.M.; Reynolds, E.H. Do the Benefits of Folic Acid Fortification Outweigh the Risk of Masking Vitamin B12 Deficiency? BMJ 2018, 360, k724. [Google Scholar] [CrossRef] [PubMed]
- Vogiatzoglou, A.; Refsum, H.; Johnston, C.; Smith, S.M.; Bradley, K.M.; de Jager, C.; Budge, M.M.; Smith, A.D. Vitamin B12 Status and Rate of Brain Volume Loss in Community-Dwelling Elderly. Neurology 2008, 71, 826–832. [Google Scholar] [CrossRef] [PubMed]
- Douaud, G.; Refsum, H.; de Jager, C.A.; Jacoby, R.; Nichols, T.E.; Smith, S.M.; Smith, A.D. Preventing Alzheimer’s Disease-Related Gray Matter Atrophy by B-Vitamin Treatment. Proc. Natl. Acad. Sci. USA 2013, 110, 9523–9528. [Google Scholar] [CrossRef]
- Ueno, A.; Hamano, T.; Enomoto, S.; Shirafuji, N.; Nagata, M.; Kimura, H.; Ikawa, M.; Yamamura, O.; Yamanaka, D.; Ito, T.; et al. Influences of Vitamin B12 Supplementation on Cognition and Homocysteine in Patients with Vitamin B12 Deficiency and Cognitive Impairment. Nutrients 2022, 14, 1494. [Google Scholar] [CrossRef]
- Song, H.; Bharadwaj, P.K.; Raichlen, D.A.; Habeck, C.G.; Huentelman, M.J.; Hishaw, G.A.; Trouard, T.P.; Alexander, G.E. Association of Homocysteine-Related Subcortical Brain Atrophy with White Matter Lesion Volume and Cognition in Healthy Aging. Neurobiol. Aging 2023, 121, 129–138. [Google Scholar] [CrossRef]
- Jiang, B.; Yao, G.; Yao, C.; Zheng, N. The effect of folate and VitB12 in the treatment of MCI patients with hyperhomocysteinemia. J. Clin. Neurosci. 2020, 81, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Vogiatzoglou, A.; Smith, A.D.; Nurk, E.; Drevon, C.A.; Ueland, P.M.; Vollset, S.E.; Nygaard, H.A.; Engedal, K.; Tell, G.S.; Refsum, H. Cognitive Function in an Elderly Population: Interaction Between Vitamin B12 Status, Depression, and Apolipoprotein E4: The Hordaland Homocysteine Study. Psychosom. Med. 2012, 75, 20–29. [Google Scholar] [CrossRef] [PubMed]
- de Jager, C.A.; Oulhaj, A.; Jacoby, R.; Refsum, H.; Smith, A.D. Cognitive and Clinical Outcomes of Homocysteine-Lowering B-Vitamin Treatment in Mild Cognitive Impairment: A Randomized Controlled Trial. Int. J. Geriatr. Psychiatry 2012, 27, 592–600. [Google Scholar] [CrossRef] [PubMed]
- Kwok, T.; Wu, Y.; Lee, J.; Lee, R.; Yung, C.Y.; Choi, G.; Lee, V.; Harrison, J.; Lam, L.; Mok, V. A Randomized Placebo-Controlled Trial of Using B Vitamins to Prevent Cognitive Decline in Older Mild Cognitive Impairment Patients. Clin. Nutr. 2019, 39, 2399–2405. [Google Scholar] [CrossRef]
- Aisen, P.S.; Schneider, L.S.; Sano, M.; Diaz-Arrastia, R.; van Dyck, C.H.; Weiner, M.F.; Bottiglieri, T.; Jin, S.; Stokes, K.T.; Thomas, R.G.; et al. High Dose B Vitamin Supplementation and Cognitive Decline in Alzheimer’s Disease: A Randomized Controlled Trial. J. Am. Med. Assoc. 2008, 300, 1774–1783. [Google Scholar] [CrossRef]
- Ford, A.H.; Flicker, L.; Alfonso, H.; Thomas, J.; Clarnette, R.; Martins, R.; Almeida, O.P. Vitamins B12, B6, and Folic Acid for Cognition in Older Men. Neurology 2010, 75, 1540–1547. [Google Scholar] [CrossRef]
- Dangour, A.D.; Allen, E.; Clarke, R.; Elbourne, D.; Fletcher, A.E.; Letley, L.; Richards, M.; Whyte, K.; Uauy, R.; Mills, K. Effects of Vitamin B-12 Supplementation on Neurologic and Cognitive Function in Older People: A Randomized Controlled Trial. Am. J. Clin. Nutr. 2015, 102, 639–647. [Google Scholar] [CrossRef]
- Poly, C.; Massaro, J.M.; Seshadri, S.; Wolf, P.A.; Cho, E.; Krall, E.; Jacques, P.F.; Au, R. The Relation of Dietary Choline to Cognitive Performance and White-Matter Hyperintensity in the Framingham Offspring Cohort 1–4. Am. J. Clin. Nutr. 2011, 94, 1584–1591. [Google Scholar] [CrossRef]
- Liu, L.; Qiao, S.; Zhuang, L.; Xu, S.; Chen, L.; Lai, Q.; Wang, W. Choline Intake Correlates with Cognitive Performance among Elder Adults in the United States. Behav. Neurol. 2021, 2021, 2962245. [Google Scholar] [CrossRef]
- Nakazaki, E.; Mah, E.; Sanoshy, K.; Citrolo, D.; Watanabe, F. Citicoline and Memory Function in Healthy Older Adults: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. J. Nutr. 2021, 151, 2153–2160. [Google Scholar] [CrossRef]
- Lippelt, D.P.; van der Kint, S.; van Herk, K.; Naber, M. No Acute Effects of Choline Bitartrate Food Supplements on Memory in Healthy, Young, Human Adults. PLoS ONE 2016, 11, e0157714. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.J.; Andrews, N.; Ball, D.; Bellantuono, I.; Gray, J.; Hachoumi, L.; Holmes, A.; Latcham, J.; Petrie, A.; Potter, P.; et al. Does Age Matter? The Impact of Rodent Age on Study Outcomes. Lab. Anim. 2017, 51, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Holtze, S.; Gorshkova, E.; Braude, S.; Cellerino, A.; Dammann, P.; Hildebrandt, T.B.; Hoeflich, A.; Hoffmann, S.; Koch, P.; Terzibasi Tozzini, E.; et al. Alternative Animal Models of Aging Research. Front. Mol. Biosci. 2021, 8, 660959. [Google Scholar] [CrossRef] [PubMed]
- Fotenos, A.F.; Snyder, A.Z.; Girton, L.E.; Morris, J.C.; Buckner, R.L. Normative Estimates of Cross-Sectional and Longitudinal Brain Volume Decline in Aging and AD. Neurology 2005, 64, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Deery, H.A.; Di Paolo, R.; Moran, C.; Egan, G.F.; Jamadar, S.D. Lower Brain Glucose Metabolism in Normal Ageing Is Predominantly Frontal and Temporal: A Systematic Review and Pooled Effect Size and Activation Likelihood Estimates Meta-analyses. Hum. Brain Mapp. 2022, 44, 1251–1277. [Google Scholar] [CrossRef]
- Zhou, D.; Sun, Y.; Qian, Z.; Wang, Z.; Zhang, D.; Li, Z.; Zhao, J.; Dong, C.; Li, W.; Huang, G. Long-Term Dietary Folic Acid Supplementation Attenuated Aging-Induced Hippocampus Atrophy and Promoted Glucose Uptake in 25-Month-Old Rats with Cognitive Decline. J. Nutr. Biochem. 2023, 117, 109328. [Google Scholar] [CrossRef]
- Serra, M.; Chan, A.; Dubey, M.; Gilman, V.; Shea, T.B. Folate and S-Adenosylmethionine Modulate Synaptic Activity in Cultured Cortical Neurons: Acute Differential Impact on Normal and Apolipoprotein-Deficient Mice. Phys. Biol. 2008, 5, 044002. [Google Scholar] [CrossRef]
- Chan, A.; Tchantchou, F.; Graves, V.; Rozen, R.; Shea, T.B. Dietary and Genetic Compromise in Folate Availability Reduces Acetylcholine, Cognitive Performance and Increases Aggression: Critical Role of S-Adenosyl Methionine. J. Nutr. Health Aging 2008, 12, 252–261. [Google Scholar] [CrossRef]
- Kruman, I.I.; Mouton, P.R.; Emokpae, R.; Cutler, R.G.; Mattson, M.P. Folate Deficiency Inhibits Proliferation of Adult Hippocampal Progenitors. Neuroreport 2005, 16, 1055–1059. [Google Scholar] [CrossRef]
- Wei, W.; Liu, Y.-H.; Zhang, C.-E.; Qang, Q.; Wei, Z.; Mousseau, D.D.; Wang, J.-Z.; Tian, Q.; Liu, G.-P. Folate/Vitamin-B12 Prevents Chronic Hyperhomocysteinemia-Induced Tau Hyperphosphorylation and Memory Deficits in Aged Rats. J. Alzheimers Dis. 2011, 27, 639–650. [Google Scholar] [CrossRef]
- Bahnfleth, C.L.; Strupp, B.J.; Caudill, M.A.; Canfield, R.L. Prenatal Choline Supplementation Improves Child Sustained Attention: A 7-year Follow-up of a Randomized Controlled Feeding Trial. FASEB J. 2022, 36, e22054. [Google Scholar] [CrossRef] [PubMed]
- Mellott, T.J.; Williams, C.L.; Meck, W.H.; Blusztajn, J.K. Prenatal Choline Supplementation Advances Hippocampal Development and Enhances MAPK and CREB Activation. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2004, 18, 545–547. [Google Scholar] [CrossRef]
- Glenn, M.J.; Kirby, E.D.; Gibson, E.M.; Wong-Goodrich, S.J.; Mellott, T.J.; Blusztajn, J.K.; Williams, C.L. Age-Related Declines in Exploratory Behavior and Markers of Hippocampal Plasticity Are Attenuated by Prenatal Choline Supplementation in Rats. Brain Res. 2008, 1237, 110–123. [Google Scholar] [CrossRef]
- Sandstrom, N.J.; Loy, R.; Williams, C.L. Prenatal Choline Supplementation Increases NGF Levels in the Hippocampus and Frontal Cortex of Young and Adult Rats. Brain Res. 2002, 947, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.-K.; Scott, A.C.; Penney, T.B.; Williams, C.L.; Meck, W.H. Prenatal-Choline Supplementation Differentially Modulates Timing of Auditory and Visual Stimuli in Aged Rats. Brain Res. 2008, 1237, 167–175. [Google Scholar] [CrossRef]
- Konno, M.; Asai, A.; Kawamoto, K.; Nishida, N.; Satoh, T.; Doki, Y.; Mori, M.; Ishii, H. The One-Carbon Metabolism Pathway Highlights Therapeutic Targets for Gastrointestinal Cancer (Review). Int. J. Oncol. 2017, 50, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Franco, C.N.; Seabrook, L.J.; Nguyen, S.T.; Leonard, J.T.; Albrecht, L.V. Simplifying the B Complex: How Vitamins B6 and B9 Modulate One Carbon Metabolism in Cancer and Beyond. Metabolites 2022, 12, 961. [Google Scholar] [CrossRef]
- Lapchak, P.A.; Araujo, D.M. Advances in Ischemic Stroke Treatment: Neuroprotective and Combination Therapies. Expert Opin. Emerg. Drugs 2007, 12, 97–112. [Google Scholar] [CrossRef]
- Arranz-Romera, A.; Esteban-Pérez, S.; Garcia-Herranz, D.; Aragón-Navas, A.; Bravo-Osuna, I.; Herrero-Vanrell, R. Combination Therapy and Co-Delivery Strategies to Optimize Treatment of Posterior Segment Neurodegenerative Diseases. Drug Discov. Today 2019, 24, 1644–1653. [Google Scholar] [CrossRef]
- Mau, K.J.; Jadavji, N.M. A New Perspective on Parkinson’s Disease: Pathology Begins in the Gastrointestinal Tract. J. Young Investig. 2017, 33, 1–8. [Google Scholar] [CrossRef]
- Hörder, H.; Johansson, L.; Guo, X.; Grimby, G.; Kern, S.; Östling, S.; Skoog, I. Midlife Cardiovascular Fitness and Dementia: A 44-Year Longitudinal Population Study in Women. Neurology 2018, 90, e1298–e1305. [Google Scholar] [CrossRef] [PubMed]
- Suvila, K.; Lima, J.A.C.; Yano, Y.; Tan, Z.S.; Cheng, S.; Niiranen, T.J. Early-but Not Late-Onset Hypertension Is Related to Midlife Cognitive Function. Hypertension 2021, 77, 972–979. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Virdi, S.; McKee, A.M.; Nuthi, M.; Jadavji, N.M. The Role of One-Carbon Metabolism in Healthy Brain Aging. Nutrients 2023, 15, 3891. https://doi.org/10.3390/nu15183891
Virdi S, McKee AM, Nuthi M, Jadavji NM. The Role of One-Carbon Metabolism in Healthy Brain Aging. Nutrients. 2023; 15(18):3891. https://doi.org/10.3390/nu15183891
Chicago/Turabian StyleVirdi, Sapna, Abbey M. McKee, Manogna Nuthi, and Nafisa M. Jadavji. 2023. "The Role of One-Carbon Metabolism in Healthy Brain Aging" Nutrients 15, no. 18: 3891. https://doi.org/10.3390/nu15183891
APA StyleVirdi, S., McKee, A. M., Nuthi, M., & Jadavji, N. M. (2023). The Role of One-Carbon Metabolism in Healthy Brain Aging. Nutrients, 15(18), 3891. https://doi.org/10.3390/nu15183891