


Plant-Based Dietary Fibers and Polysaccharides as Modulators of Gut Microbiota in Intestinal and Lung Inflammation: Current State and Challenges

Abstract
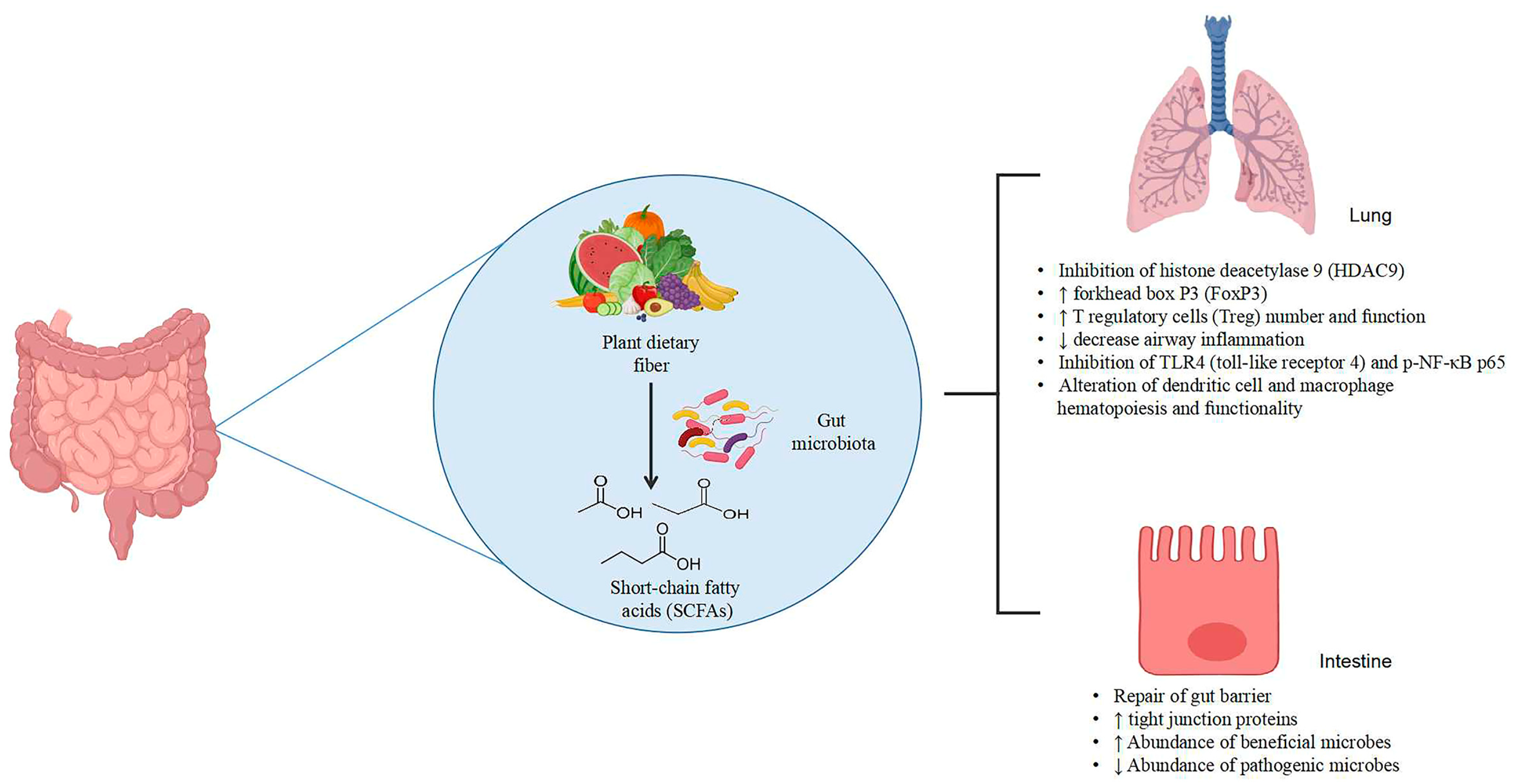
1. Introduction
2. Plant Dietary Fibers and Polysaccharides against Gut and Lung Inflammation
2.1. Different Plant Dietary Fibers and Polysaccharides Shape Gut Microbiota and Affect SCFA Production Differently
2.2. Plant Dietary Fibers and Polysaccharides Alter the Intestinal Barrier in Both Gut Inflammation and Lung Inflammation
2.3. Plant-Dietary-Fiber and Polysaccharide Supplementation in Gut and Lung Inflammation in Human Adults (Clinical Studies) and Adult Mice (Preclinical Studies)
2.4. Plant-Dietary-Fiber and Polysaccharide Supplementation in Maternal Rodent Models
2.5. Sex Differences in Lung Anti-Inflammatory Effect of Plant DFs and Polysaccharides
2.6. Combination of Plant Polysaccharides with Other Phytochemicals
2.7. Mechanisms of Polysaccharides and Dietary Fibers in Reducing Intestinal and Lung Inflammation
3. Issues and Challenges
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Molodecky, N.A.; Soon, I.S.; Rabi, D.M.; Ghali, W.A.; Ferris, M.; Chernoff, G.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Barkema, H.W.; et al. Increasing Incidence and Prevalence of the Inflammatory Bowel Diseases with Time, Based on Systematic Review. Gastroenterology 2012, 142, 46–54.e42. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Chronic Obstructive Pulmonary Disease (COPD). Available online: https://www.who.int/news-room/fact-sheets/detail/chronic-obstructive-pulmonary-disease-(copd) (accessed on 17 June 2023).
- Piovani, D.; Danese, S.; Peyrin-Biroulet, L.; Bonovas, S. Inflammatory bowel disease: Estimates from the global burden of disease 2017 study. Aliment. Pharm. Ther. 2020, 51, 261–270. [Google Scholar] [CrossRef]
- Barcik, W.; Boutin, R.C.T.; Sokolowska, M.; Finlay, B.B. The Role of Lung and Gut Microbiota in the Pathology of Asthma. Immunity 2020, 52, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cai, Y.; Garssen, J.; Henricks, P.A.J.; Folkerts, G.; Braber, S. The bidirectional gut–lung axis in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2023, 207, 1145–1160. [Google Scholar] [CrossRef]
- Kisiel, M.A.; Sedvall, M.; Malinovschi, A.; Franklin, K.A.; Gislason, T.; Shlunssen, V.; Johansson, A.; Modig, L.; Jogi, R.; Holm, M.; et al. Inflammatory bowel disease and asthma. Results from the RHINE study. Respir. Med. 2023, 216, 107307. [Google Scholar] [CrossRef] [PubMed]
- Antoine, M.H.; Mlika, M. Interstitial lung disease. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK541084/ (accessed on 7 June 2023).
- Luo, Q.; Zhou, P.; Chang, S.; Huang, Z.; Zhu, Y. The gut-lung axis: Mendelian randomization identifies a causal association between inflammatory bowel disease and interstitial lung disease. Heart Lung 2023, 61, 120–126. [Google Scholar] [CrossRef]
- Cai, Z.; Wang, S.; Li, J. Treatment of inflammatory bowel disease: A comprehensive review. Front. Med. 2021, 8, 765474. [Google Scholar] [CrossRef]
- Gross, N.J.; Barnes, P.J. New therapies for asthma and chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2017, 195, 159–166. [Google Scholar] [CrossRef]
- Niu, Y.; Liu, W.; Fan, X.; Wen, D.; Wu, D.; Wang, H.; Liu, Z.; Li, B. Beyond cellulose: Pharmaceutical potential for bioactive plant polysaccharides in treating disease and gut dysbiosis. Front. Microbiol. 2023, 14, 1183130. [Google Scholar] [CrossRef]
- Lovegrove, A.; Edwards, C.H.; De Noni, I.; Patel, H.; El, S.N.; Grassby, T.; Zielke, C.; Ulmius, M.; Nilsson, L.; Butterworth, P.J.; et al. Role of polysaccharides in food, digestion, and health. Crit. Rev. Food Sci. Nutr. 2017, 57, 237–253. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Lu, W.; Li, W.; Pan, L.; Lu, M.; Igor, C.; Li, Z.; Zeng, W. Dietary fiber in plant cell walls—The healthy carbohydrates. Food Qual. Saf. 2022, 6, fyab037. [Google Scholar] [CrossRef]
- Stribling, P.; Ibrahim, F. Dietary fibre definition revisited—The case of low molecular weight carbohydrates. Clin. Nutr. ESPEN 2023, 55, 340–356. [Google Scholar] [CrossRef]
- Qiu, P.; Ishimoto, T.; Fu, L.; Zhang, J.; Zhang, Z.; Liu, Y. The Gut Microbiota in Inflammatory Bowel Disease. Front. Cell. Infect. Microbiol. 2022, 12, 733992. [Google Scholar] [CrossRef] [PubMed]
- Bowerman, K.L.; Rehman, S.F.; Vaughan, A.; Lachner, N.; Budden, K.F.; Kim, R.Y.; Wood, D.L.; Gellatly, S.L.; Shukla, S.D.; Wood, L.G.; et al. Disease-associated gut microbiome and metabolome changes in patients with chronic obstructive pulmonary disease. Nat. Commun. 2020, 11, 5886. [Google Scholar] [CrossRef]
- Russo, C.; Colaianni, V.; Ielo, G.; Valle, M.S.; Spicuzza, L.; Malaguarnera, L. Impact of lung microbiota on COPD. Biomedicines 2022, 10, 1337. [Google Scholar] [CrossRef] [PubMed]
- Magryś, A. Microbiota: A Missing Link in The Pathogenesis of Chronic Lung Inflammatory Diseases. Pol. J. Microbiol. 2021, 70, 25–32. [Google Scholar] [CrossRef]
- Price, C.E.; O’Toole, G.A. The Gut-Lung Axis in Cystic Fibrosis. J. Bacteriol. 2021, 203, 10–1128. [Google Scholar] [CrossRef]
- Yang, J.-Y.; Chen, S.-Y.; Wu, Y.-H.; Liao, Y.-L.; Yen, G.-C. Ameliorative effect of buckwheat polysaccharides on colitis via regulation of the gut microbiota. Int. J. Biol. Macromol. 2023, 227, 872–883. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, P.; Li, C.; Xu, F.; Chen, J. A polysaccharide from Rosa roxburghii Tratt fruit attenuates high-fat diet-induced intestinal barrier dysfunction and inflammation in mice by modulating the gut microbiota. Food Funct. 2022, 13, 530–547. [Google Scholar] [CrossRef]
- Luo, H.; Ying, N.; Zhao, Q.; Chen, J.; Xu, H.; Jiang, W.; Wu, Y.; Wu, Y.; Gao, H.; Zheng, H. A novel polysaccharide from Rubus chingii Hu unripe fruits: Extraction optimization, structural characterization and amelioration of colonic inflammation and oxidative stress. Food Chem. 2023, 421, 136152. [Google Scholar] [CrossRef]
- Lu, H.; Shen, M.; Chen, Y.; Yu, Q.; Chen, T.; Xie, J. Alleviative effects of natural plant polysaccharides against DSS-induced ulcerative colitis via inhibiting inflammation and modulating gut microbiota. Food Res. Int. 2023, 167, 112630. [Google Scholar] [CrossRef]
- Li, X.; Qiao, G.; Chu, L.; Lin, L.; Zheng, G. Smilax china L. Polysaccharide Alleviates Dextran Sulphate Sodium-Induced Colitis and Modulates the Gut Microbiota in Mice. Foods 2023, 12, 1632. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Zhou, Y.; Zhang, R.; Chen, Y.; Wang, F.; Zhang, M. Gut microbial fermentation promotes the intestinal anti-inflammatory activity of Chinese yam polysaccharides. Food Chem. 2023, 402, 134003. [Google Scholar] [CrossRef]
- Zeng, Z.; Xie, Z.; Chen, G.; Sun, Y.; Zeng, X.; Liu, Z. Anti-inflammatory and gut microbiota modulatory effects of polysaccharides from Fuzhuan brick tea on colitis in mice induced by dextran sulfate sodium. Food Funct. 2022, 13, 649–663. [Google Scholar] [CrossRef]
- Xiao, H.; Feng, J.; Peng, J.; Wu, P.; Chang, Y.; Li, X.; Wu, J.; Huang, H.; Deng, H.; Qiu, M.; et al. Fuc-S-A New Ultrasonic Degraded Sulfated α-l-Fucooligosaccharide-Alleviates DSS-Inflicted Colitis through Reshaping Gut Microbiota and Modulating Host-Microbe Tryptophan Metabolism. Mar. Drugs 2022, 21, 16. [Google Scholar] [CrossRef]
- Xu, A.; Zhao, Y.; Shi, Y.; Zuo, X.; Yang, Y.; Wang, Y.; Xu, P. Effects of oxidation-based tea processing on the characteristics of the derived polysaccharide conjugates and their regulation of intestinal homeostasis in DSS-induced colitis mice. Int. J. Biol. Macromol. 2022, 214, 402–413. [Google Scholar] [CrossRef]
- Wu, Q.; Fan, L.; Tan, H.; Zhang, Y.; Fang, Q.; Yang, J.; Cui, S.W.; Nie, S. Impact of pectin with various esterification degrees on the profiles of gut microbiota and serum metabolites. Appl. Microbiol. Biotechnol. 2022, 106, 3707–3720. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-J.; Li, Q.-M.; Zha, X.-Q.; Luo, J.-P. Dendrobium fimbriatum Hook polysaccharide ameliorates dextran-sodium-sulfate-induced colitis in mice via improving intestinal barrier function, modulating intestinal microbiota, and reducing oxidative stress and inflammatory responses. Food Funct. 2022, 13, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Zhong, R.; Dang, G.; Xia, B.; Wu, W.; Tang, S.; Tang, L.; Liu, L.; Liu, Z.; Chen, L.; et al. Pectin supplementation ameliorates intestinal epithelial barrier function damage by modulating intestinal microbiota in lipopolysaccharide-challenged piglets. J. Nutr. Biochem. 2022, 109, 109107. [Google Scholar] [CrossRef]
- Lv, H.; Jia, H.; Cai, W.; Cao, R.; Xue, C.; Dong, N. Rehmannia glutinosa polysaccharides attenuates colitis via reshaping gut microbiota and short-chain fatty acid production. J. Sci. Food Agric. 2023, 103, 3926–3938. [Google Scholar] [CrossRef] [PubMed]
- Li, L.-X.; Feng, X.; Tao, M.-T.; Paulsen, B.S.; Huang, C.; Feng, B.; Liu, W.; Yin, Z.-Q.; Song, X.; Zhao, X.; et al. Benefits of neutral polysaccharide from rhizomes of Polygonatum sibiricum to intestinal function of aged mice. Front. Nutr. 2022, 9, 992102. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Shen, Y.; Lin, J.; Yen, G. Rhinacanthus nasutus and okara polysaccharides attenuate colitis via inhibiting inflammation and modulating the gut microbiota. Phytother. Res. 2022, 36, 4631–4645. [Google Scholar] [CrossRef] [PubMed]
- Cen, L.; Yi, T.; Hao, Y.; Shi, C.; Shi, X.; Lu, Y.; Chen, D.; Zhu, H. Houttuynia cordata polysaccharides alleviate ulcerative colitis by restoring intestinal homeostasis. Chin. J. Nat. Med. 2022, 20, 914–924. [Google Scholar] [CrossRef]
- Zhou, X.; Lu, Q.; Kang, X.; Tian, G.; Ming, D.; Yang, J. Protective Role of a New Polysaccharide Extracted from Lonicera japonica Thunb in Mice with Ulcerative Colitis Induced by Dextran Sulphate Sodium. Biomed Res. Int. 2021, 2021, 8878633. [Google Scholar] [CrossRef]
- Guo, C.; Wang, Y.; Zhang, S.; Zhang, X.; Du, Z.; Li, M.; Ding, K. Crataegus pinnatifida polysaccharide alleviates colitis via modulation of gut microbiota and SCFAs metabolism. Int. J. Biol. Macromol. 2021, 181, 357–368. [Google Scholar] [CrossRef]
- Jin, M.-Y.; Wu, X.-Y.; Li, M.-Y.; Li, X.-T.; Huang, R.-M.; Sun, Y.-M.; Xu, Z.-L. Noni (Morinda citrifolia L.) fruit polysaccharides regulated IBD mice via targeting gut microbiota: Association of JNK/ERK/NF-κB signaling pathways. J. Agric. Food Chem. 2021, 69, 10151–10162. [Google Scholar] [CrossRef]
- Cui, L.; Guan, X.; Ding, W.; Luo, Y.; Wang, W.; Bu, W.; Song, J.; Tan, X.; Sun, E.; Ning, Q.; et al. Scutellaria baicalensis Georgi polysaccharide ameliorates DSS-induced ulcerative colitis by improving intestinal barrier function and modulating gut microbiota. Int. J. Biol. Macromol. 2021, 166, 1035–1045. [Google Scholar] [CrossRef]
- Yang, M.; Lee, U.; Cho, H.-R.; Lee, K.B.; Shin, Y.J.; Bae, M.-J.; Park, K.-Y. Effects of pear extracts on microbiome and immunocytokines to alleviate air pollution-related respiratory hypersensitivity. J. Med. Food 2023, 26, 211–214. [Google Scholar] [CrossRef]
- Yuan, G.; Wen, S.; Zhong, X.; Yang, X.; Xie, L.; Wu, X.; Li, X. Inulin alleviates offspring asthma by altering maternal intestinal microbiome composition to increase short-chain fatty acids. PLoS ONE 2023, 18, e0283105. [Google Scholar] [CrossRef]
- Wei, Y.; Qi, M.; Liu, C.; Li, L. Astragalus polysaccharide attenuates bleomycin-induced pulmonary fibrosis by inhibiting TLR4/NF-κB signaling pathway and regulating gut microbiota. Eur. J. Pharmacol. 2023, 944, 175594. [Google Scholar] [CrossRef]
- Ling, L.; Ren, A.; Lu, Y.; Zhang, Y.; Zhu, H.; Tu, P.; Li, H.; Chen, D. The synergistic effect and mechanisms of flavonoids and polysaccharides from Houttuynia cordata on H1N1-induced pneumonia in mice. J. Ethnopharmacol. 2023, 302, 115761. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Yuan, G.; Li, C.; Xiong, Y.; Zhong, X.; Li, X. High cellulose dietary intake relieves asthma inflammation through the intestinal microbiome in a mouse model. PLoS ONE 2022, 17, e0263762. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Zhou, L.; Li, H.; Shi, X.; Zhang, Y.; Lu, Y.; Zhu, H.; Chen, D. Intestinal microbiota metabolizing Houttuynia cordata polysaccharides in H1N1 induced pneumonia mice contributed to Th17/Treg rebalance in gut-lung axis. Int. J. Biol. Macromol. 2022, 221, 288–302. [Google Scholar] [CrossRef] [PubMed]
- Ming, K.; Zhuang, S.; Ma, N.; Nan, S.; Li, Q.; Ding, M.; Ding, Y. Astragalus polysaccharides alleviates lipopolysaccharides-induced inflammatory lung injury by altering intestinal microbiota in mice. Front. Microbiol. 2022, 13, 1033875. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, Q.; Ren, R.; Zhang, Q.; Yan, G.; Yin, D.; Zhang, M.; Yang, Y. Platycodon grandiflorus polysaccharides deeply participate in the anti-chronic bronchitis effects of platycodon grandiflorus decoction, a representative of “the lung and intestine are related”. Front. Pharmacol. 2022, 13, 927384. [Google Scholar] [CrossRef]
- Liu, J.-X.; Yuan, H.-Y.; Li, Y.-N.; Wei, Z.; Liu, Y.; Liang, J. Ephedra sinica polysaccharide alleviates airway inflammations of mouse asthma-like induced by PM2.5 and ovalbumin via the regulation of gut microbiota and short chain fatty acid. J. Pharm. Pharmacol. 2022, 74, 1784–1796. [Google Scholar] [CrossRef]
- Lu, J.; Zhu, B.; Zhou, F.; Ding, X.; Qian, C.; Ding, Z.; Ye, X. Polysaccharides from the aerial parts of Tetrastigma hemsleyanum Diels et Gilg induce bidirectional immunity and ameliorate LPS-induced acute respiratory distress syndrome in mice. Front. Pharmacol. 2022, 13, 838873. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, X.; Shen, J.; Xu, A.; Wang, Y.; Meng, Q.; Xu, P. Black Tea Alleviates Particulate Matter-Induced Lung Injury via the Gut-Lung Axis in Mice. J. Agric. Food Chem. 2021, 69, 15362–15373. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.O.; Kim, O.-H.; Kim, S.J.; Lee, S.H.; Yun, S.; Lim, S.E.; Yoo, H.J.; Shin, Y.; Lee, S.W. High-fiber diets attenuate emphysema development via modulation of gut microbiota and metabolism. Sci. Rep. 2021, 11, 7008. [Google Scholar] [CrossRef]
- Tashiro, H.; Kasahara, D.I.; Osgood, R.S.; Brown, T.; Cardoso, A.; Cho, Y.; Shore, S.A. Sex differences in the impact of dietary fiber on pulmonary responses to ozone. Am. J. Respir. Cell Mol. Biol. 2020, 62, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Zhu, H.; Li, H.; Zeng, D.; Shi, X.; Zhang, Y.; Lu, Y.; Ling, L.; Wang, C.; Chen, D. Regulating the balance of Th17/Treg cells in gut-lung axis contributed to the therapeutic effect of Houttuynia cordata polysaccharides on H1N1-induced acute lung injury. Int. J. Biol. Macromol. 2020, 158, 52–66. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Shi, C.; Zhou, X.; Wen, W.; Gao, X.; Wang, L.; He, B.; Yin, M.; Zhao, J.-Q. Lycium barbarum polysaccharide extracted from Lycium barbarum leaves ameliorates asthma in mice by reducing inflammation and modulating gut microbiota. J. Med. Food 2020, 23, 699–710. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, R.; Berthon, B.S.; Rogers, G.B.; Baines, K.J.; Leong LE, X.; Gibson, P.G.; Williams, E.J.; Wood, L.G. Soluble fibre supplementation with and without a probiotic in adults with asthma: A 7-day randomised, double blind, three way cross-over trial. EBioMedicine 2019, 46, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-Y.; Li, H.; Lu, X.-X.; Ling, L.-J.; Weng, H.-B.; Sun, W.; Chen, D.-F.; Zhang, Y.-Y. Houttuynia cordata polysaccharide alleviated intestinal injury and modulated intestinal microbiota in H1N1 virus infected mice. Chin. J. Nat. Med. 2019, 17, 187–197. [Google Scholar] [CrossRef]
- Zhu, H.; Lu, X.; Ling, L.; Li, H.; Ou, Y.; Shi, X.; Lu, Y.; Zhang, Y.; Chen, D. Houttuynia cordata polysaccharides ameliorate pneumonia severity and intestinal injury in mice with influenza virus infection. J. Ethnopharmacol. 2018, 218, 90–99. [Google Scholar] [CrossRef]
- Cho, Y.; Abu-Ali, G.; Tashiro, H.; Kasahara, D.I.; Brown, T.A.; Brand, J.D.; Mathews, J.A.; Huttenhower, C.; Shore, S.A. The microbiome regulates pulmonary responses to ozone in mice. Am. J. Respir. Cell Mol. Biol. 2018, 59, 346–354. [Google Scholar] [CrossRef]
- Verheijden KA, T.; Braber, S.; Leusink-Muis, T.; Jeurink, P.V.; Thijssen, S.; Kraneveld, A.D.; Garssen, J.; Folkerts, G.; Willemsen, L.E.M. The combination therapy of dietary galacto-oligosaccharides with budesonide reduces pulmonary Th2 driving mediators and mast cell degranulation in a murine model of house dust mite induced asthma. Front. Immunol. 2018, 9, 2419. [Google Scholar] [CrossRef]
- Halnes, I.; Baines, K.; Berthon, B.; MacDonald-Wicks, L.; Gibson, P.; Wood, L. Soluble fibre meal challenge reduces airway inflammation and expression of GPR43 and GPR41 in asthma. Nutrients 2017, 9, 57. [Google Scholar] [CrossRef]
- Zhang, Z.; Shi, L.; Pang, W.; Liu, W.; Li, J.; Wang, H.; Shi, G. Dietary Fiber Intake Regulates Intestinal Microflora and Inhibits Ovalbumin-Induced Allergic Airway Inflammation in a Mouse Model. PLoS ONE 2016, 11, e0147778. [Google Scholar] [CrossRef]
- Xu, Y.-Y.; Zhang, Y.-Y.; Ou, Y.-Y.; Lu, X.-X.; Pan, L.-Y.; Li, H.; Lu, Y.; Chen, D.-F. Houttuynia cordata Thunb. polysaccharides ameliorates lipopolysaccharide-induced acute lung injury in mice. J. Ethnopharmacol. 2015, 173, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Bernard, H.; Desseyn, J.-L.; Bartke, N.; Kleinjans, L.; Stahl, B.; Belzer, C.; Knol, J.; Gottrand, F.; Husson, M.O. Dietary pectin–derived acidic oligosaccharides improve the pulmonary bacterial clearance of Pseudomonas aeruginosa lung infection in mice by modulating intestinal microbiota and immunity. J. Infect. Dis. 2015, 211, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Verheijden, K.A.; Willemsen, L.E.; Braber, S.; Leusink-Muis, T.; Delsing, D.J.; Garssen, J.; Kraneveld, A.D.; Folkerts, G. Dietary galacto-oligosaccharides prevent airway eosinophilia and hyperresponsiveness in a murine house dust mite-induced asthma model. Respir. Res. 2015, 16, 17. [Google Scholar] [CrossRef] [PubMed]
- Hogenkamp, A.; Thijssen, S.; Van Vlies, N.; Garssen, J. Supplementing pregnant mice with a specific mixture of nondigestible oligosaccharides reduces symptoms of allergic asthma in male offspring. J. Nutr. 2015, 145, 640–646. [Google Scholar] [CrossRef]
- Thorburn, A.N.; McKenzie, C.I.; Shen, S.; Stanley, D.; Macia, L.; Mason, L.J.; Roberts, L.K.; Wong, C.H.Y.; Shim, R.; Robert, R.; et al. Evidence that asthma is a developmental origin disease influenced by maternal diet and bacterial metabolites. Nat. Commun. 2015, 6, 7320. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef]
- Chung, M.J.; Park, J.K.; Park, Y.I. Anti-inflammatory effects of low-molecular weight chitosan oligosaccharides in IgE–antigen complex-stimulated RBL-2H3 cells and asthma model mice. Int. Immunopharmacol. 2012, 12, 453–459. [Google Scholar] [CrossRef]
- Yasuda, A.; Inoue, K.-I.; Sanbongi, C.; Yanagisawa, R.; Ichinose, T.; Yoshikawa, T.; Takano, H. Dietary Supplementation with Fructooligosaccharides Attenuates Airway Inflammation Related to House Dust Mite Allergen in Mice. Int. J. Immunopath. Pharmacol. 2010, 23, 727–735. [Google Scholar] [CrossRef]
- Vos, A.P.; van Esch, B.C.; Stahl, B.; M’Rabet, L.; Folkerts, G.; Nijkamp, F.P.; Garssen, J. Dietary supplementation with specific oligosaccharide mixtures decreases parameters of allergic asthma in mice. Int. Immunopharmacol. 2007, 7, 1582–1587. [Google Scholar] [CrossRef]
- Sonoyama, K.; Watanabe, H.; Watanabe, J.; Yamaguchi, N.; Yamashita, A.; Hashimoto, H.; Kishino, E.; Fujita, K.; Okada, M.; Mori, S.; et al. Allergic airway eosinophilia is suppressed in ovalbumin-sensitized brown norway rats fed raffinose and α-linked galactooligosaccharide1. J. Nutr. 2005, 135, 538–543. [Google Scholar] [CrossRef]
- Lee, J.C.; Pak, S.C.; Lee, S.H.; Na, C.S.; Lim, S.C.; Song, C.H.; Bai, Y.H.; Jang, C.H. Asian pear pectin administration during presensitization inhibits allergic response to ovalbumin in BALB/c mice. J. Altern. Complement. Med. 2004, 10, 527–534. [Google Scholar] [CrossRef]
- Watanabe, H.; Sonoyama, K.; Watanabe, J.; Yamaguchi, N.; Kikuchi, H.; Nagura, T.; Aritsuka, T.; Fukumoto, K.; Kasai, T. Reduction of allergic airway eosinophilia by dietary raffinose in Brown Norway rats. Br. J. Nutr. 2004, 92, 247–255. [Google Scholar] [CrossRef]
- Fan, L.; Zuo, S.; Tan, H.; Hu, J.; Cheng, J.; Wu, Q.; Nie, S. Preventive effects of pectin with various degrees of esterification on ulcerative colitis in mice. Food Funct. 2020, 11, 2886–2897. [Google Scholar] [CrossRef]
- Wang, Y.; Leong, L.E.X.; Keating, R.L.; Kanno, T.; Abell, G.C.J.; Mobegi, F.M.; Choo, J.M.; Wesselingh, S.L.; Mason, A.J.; Burr, L.D.; et al. Opportunistic bacteria confer the ability to ferment prebiotic starch in the adult cystic fibrosis gut. Gut Microbes 2019, 10, 367–381. [Google Scholar] [CrossRef]
- Wang, K.; Wu, L.; Dou, C.; Guan, X.; Wu, H.; Liu, H. Research Advance in Intestinal Mucosal Barrier and Pathogenesis of Crohn’s Disease. Gastroent. Res. Pract. 2016, 2016, 9686238. [Google Scholar] [CrossRef]
- Wang, Z.; Li, F.; Liu, J.; Luo, Y.; Guo, H.; Yang, Q.; Xu, C.; Ma, S.; Chen, H. Intestinal microbiota—An unmissable bridge to severe acute pancreatitis-associated acute lung injury. Front. Immunol. 2022, 13, 913178. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, P.; Chen, J.; Li, C.; Tian, Y.; Xu, F. Prebiotic properties of the polysaccharide from Rosa roxburghii Tratt fruit and its protective effects in high-fat diet-induced intestinal barrier dysfunction: A fecal microbiota transplantation study. Food Res. Int. 2023, 164, 112400. [Google Scholar] [CrossRef]
- Park, Y.; Subar, A.F.; Hollenbeck, A.; Schatzkin, A. Dietary Fiber Intake and Mortality in the NIH-AARP Diet and Health Study. Arch. Intern. Med. 2011, 171, 1061–1068. [Google Scholar] [CrossRef]
- Macfarlane, S.; Cleary, S.; Bahrami, B.; Reynolds, N.; Macfarlane, G.T. Synbiotic consumption changes the metabolism and composition of the gut microbiota in older people and modifies inflammatory processes: A randomised, double-blind, placebo-controlled crossover study. Aliment. Pharm. Ther. 2013, 38, 804–816. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Pattaroni, C.; Lopez-Mejia, I.C.; Riva, E.; Pernot, J.; Ubags, N.; Fajas, L.; Nicod, L.P.; Marsland, B.J. Dietary Fiber Confers Protection against Flu by Shaping Ly6c− Patrolling Monocyte Hematopoiesis and CD8+ T Cell Metabolism. Immunity 2018, 48, 992–1005.e8. [Google Scholar] [CrossRef]
- Zhou, F.; Lin, Y.; Chen, S.; Bao, X.; Fu, S.; Lv, Y.; Zhou, M.; Chen, Y.; Zhu, B.; Qian, C.; et al. Ameliorating role of Tetrastigma hemsleyanum polysaccharides in antibiotic-induced intestinal mucosal barrier dysfunction in mice based on microbiome and metabolome analyses. Int. J. Biol. Macromol. 2023, 241, 124419. [Google Scholar] [CrossRef]
- Hogenkamp, A.; Knippels LM, J.; Garssen, J.; van Esch, B.C.A.M. Supplementation of mice with specific nondigestible oligosaccharides during pregnancy or lactation leads to diminished sensitization and allergy in the female offspring. J. Nutr. 2015, 145, 996–1002. [Google Scholar] [CrossRef]
- Beauruelle, C.; Guilloux, C.-A.; Lamoureux, C.; Héry-Arnaud, G. The human microbiome, an emerging key-player in the sex gap in respiratory diseases. Front. Med. 2021, 8, 600879. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Hyun, J.E.; Thiesen, A.; Park, H.; Hotte, N.; Watanabe, H.; Higashiyama, T.; Madsen, K.L. Sex-Specific Differences in the Gut Microbiome in Response to Dietary Fiber Supplementation in IL-10-Deficient Mice. Nutrients 2020, 12, 2088. [Google Scholar] [CrossRef]
- Kalola, U.K.; Ambati, S. Budesonide. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK563201/ (accessed on 8 June 2023).
- Lee, G.R. The Balance of Th17 versus Treg Cells in Autoimmunity. Int. J. Mol. Sci. 2018, 19, 730. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Gang, X.; Yang, S.; Cui, M.; Sun, L.; Li, Z.; Wang, G. The Alterations in and the Role of the Th17/Treg Balance in Metabolic Diseases. Front. Immunol. 2021, 12, 678355. [Google Scholar] [CrossRef] [PubMed]
- Barnig, C.; Bezema, T.; Calder, P.C.; Charloux, A.; Frossard, N.; Garssen, J.; Haworth, O.; Dilevskaya, K.; Levi-Schaffer, F.; Lonsdorfer, E.; et al. Activation of Resolution Pathways to Prevent and Fight Chronic Inflammation: Lessons from Asthma and Inflammatory Bowel Disease. Front. Immunol. 2019, 10, 1699. [Google Scholar] [CrossRef]
- Cheng, H.; Guan, X.; Chen, D.; Ma, W. The Th17/Treg Cell Balance: A Gut Microbiota-Modulated Story. Microorganisms 2019, 7, 583. [Google Scholar] [CrossRef]
- Lai, H.-C.; Lin, T.-L.; Chen, T.-W.; Kuo, Y.-L.; Chang, C.-J.; Wu, T.-R.; Shu, C.C.; Tsai, Y.H.; Swift, S.; Lu, C.C. Gut microbiota modulates COPD pathogenesis: Role of anti-inflammatory Parabacteroides goldsteinii lipopolysaccharide. Gut 2022, 71, 309–321. [Google Scholar] [CrossRef]
- Fransen, F.; Sahasrabudhe, N.M.; Elderman, M.; Bosveld, M.; El Aidy, S.; Hugenholtz, F.; Theo, B.; Ben, K.; Simon, W.; van der Gaast-de Jongh, C.; et al. β2→1-Fructans Modulate the Immune System In Vivo in a Microbiota-Dependent and -Independent Fashion. Front. Immunol. 2017, 8, 154. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, B.O.; Birchenough, G.M.H.; Ståhlman, M.; Arike, L.; Johansson, M.E.V.; Hansson, G.C.; Bäckhed, F. Bifidobacteria or Fiber Protects against Diet-Induced Microbiota-Mediated Colonic Mucus Deterioration. Cell Host Microbe 2018, 23, 27–40.e7. [Google Scholar] [CrossRef] [PubMed]
- Dumas, A.; Bernard, L.; Poquet, Y.; Lugo-Villarino, G.; Neyrolles, O. The role of the lung microbiota and the gut-lung axis in respiratory infectious diseases. Cell. Microbiol. 2018, 20, e12966. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Paul, S.; Dutta, C. Geography, ethnicity or subsistence-specific variations in human microbiome composition and diversity. Front. Microbiol. 2017, 8, 1162. [Google Scholar] [CrossRef]


{kind=link}
| Gut Microbiota | Lung Microbiota | ||
|---|---|---|---|
| IBD | ↓ Bifidobacterium longum, ↓ Eubacterium rectale, ↓ Faecalibacterium prausnitzii, ↓ Roseburia intestinalis, ↑ Bacteroides, ↑ Ruminococcus torques, ↑ Ruminococcus, ↓ Christensenellaceae, ↓ Coriobacteriaceae, ↓ Clostridium leptum, ↑ Actinomyces spp., ↑ Veillonella spp., ↑ Escherichia coli, ↓ Eubacterium rectum, ↓ Akkermansia muciniphila | — | [16] |
| COPD | Streptococcus parasanguinis_B and Streptococcus salivarius, Streptococcus vestibularis, Streptococcus sp000187445, Lachnospiraceae | ↓ Veillonella, ↑ Actinomyces, ↑ Actinobacillus, ↑ Megasphaera, ↑ Selenomonas, ↑ Corynebacterium, ↑ Streptococcus pneumoniae, ↑ Gemella morbillorum, ↑ Prevotella histicola, ↑ Streptococcus gordonii | [17,18] |
| Asthma | ↑ Haemophilus, ↑ Streptococcus, ↑ Moraxella, ↑ Lactobacillus | ↑ Haemophilus spp., ↑ Moraxella catarrhalis, ↑ Streptococcus spp., Tropheryma | [19] |
| Cystic fibrosis | ↑ Escherichia, ↑ Shigella, ↑ Enterobacter, ↑ Clostridium, ↑ Veillonella, ↑ Enterococcus, ↑ Staphylococcus, ↓ Lachnospiraceae, ↓ Ruminococcu, ↓ Roseburia, ↓ Faecalibacterium, ↓ Eubacterium, ↓ Prevotella, ↓ Eggerthella, ↓ Alistipes | Pseudomonas aeruginosa, Staphylococcus aureus, H. influenzae, Burkholderia cepacia, Actinobacteria, Proteobacteria | [19,20] |
| Natural Source | Model | Effects on Inflammation Related Biomarkers | Intestinal Microbiota Modulation | References |
|---|---|---|---|---|
| Buckwheat | TNBS-induced colitis | ↓ IL-6, IL-1β, and TNF-α | ↑ F/B ratio, ↑ Oscillospiraceae, ↑ Oscillibacter, ↑ SCFAs | [21] |
| Rosa roxburghii Tratt polysaccharide | HFD-induced colitis | ↓ TNF-α, IL-6 and IL-1β, ↑ tight junction proteins | ↓ Desulfovibrionaceae, ↓ Enterobacteriaceae, ↑ Muribaculaceae, ↑ Bacteroidaceae | [22] |
| Rubus chingii Hu unripe fruit polysaccharide | HFD-induced colitis | ↓ IL-6, ↓ IL-1β, ↓ TNF-α | ↑ gut microbial diversity, ↓ Erysipelatoclostridium, ↓ Negativibacillus | [23] |
| Cyclocarya paliurus polysaccharide | DSS-induced colitis | ↑ IL-10, ↓ IL-1β, ↓ TNF-α | ↓ Akkermansia, ↓ Sutterella, ↓ AF12, ↓ Clostridiaceae_Clostridium, ↓ Helicobacter, ↓ Prevotella, ↑ Lactobacillus, ↑ Coprococcus | [24] |
| Smilax china L. polysaccharide | DSS-induced colitis | ↓ TNF-α, ↓ IL-6, ↓ IL-1β, ↑ IL-10 | ↑ Lachnospiraceae, ↑ Muribaculaceae, ↑ Blautia, ↑ Mucispirillum, ↓ Akkermansiaceae, ↓ Deferribacteraceae, ↓ Oscillibacter | [25] |
| Chinese yam polysaccharide | LPS-stimulated co-culture of Caco-2/Raw264.7 cells | ↓ NO, ↓ IL-1β, ↓ TNF-α | ↑ Bifidobacterium, ↑ Megasphaera. | [26] |
| Fuzhuan brick tea polysaccharide | DSS-induced colitis | ↓ IL-6, ↓ IL-1β, ↓ IFN-γ, ↓ TNF-α, ↑ tight junction proteins (Occludin, Claudin-1, and ZO-1), ↑ intestinal barrier function | ↑ Bacteroides, ↑ Parasutterella, ↑ Collinsella | [27] |
| Fuc-S (a sulfated α-L-Fucooligosaccharide) | DSS-induced colitis | ↓ TNF-α, ↓ IL-1β, ↓ IL-6, ↓ IL-17A | ↓ F/B ratio, ↑ Akkermansia, ↑ Prevotellaceae_UCG_001, ↓ Eubacterium xylanophilum, ↓ Intestinimonas, ↓ Ruminococcaceae UCG-014, ↓ Oscillibacter | [28] |
| Polysaccharide conjugates derived from dried fresh tea leaves, green tea, and black tea | DSS-induced colitis | ↓ IL-6, ↓ IFN-γ, ↓ IL-1β, ↓ TNF-α, ↑ IL-10 | ↑ Bacteroides, ↑ Muribaculaceae, ↓ Helicobacter, ↓ Enterococcus | [29] |
| Pectin with various esterification degrees | DSS-induced colitis | — | ↑ Lactobacillus, ↑ Bifidobacterium | [30] |
| Rosa roxburghii Tratt | HFD-induced intestinal barrier dysfunction and inflammation | ↓ TNF-α, ↓IL-6, ↓IL-1β; ↑ tight junction proteins (ZO-1, claudin-1, and occludin); ↓ intestinal permeability; ↓ colonic oxidative stress | ↓ F/B ratio, ↑ Ruminococcaceae, ↑ Muribaculaceae, ↑ Akkermansiaceae | [22] |
| Dendrobium fimbriatum Hook | DSS-induced colitis | ↓ IL-1β, ↓ IL-6, ↓ IL-17A, ↓ IL-17F, ↓ IL-21, ↓ IL-23, ↑ IL-5, ↑ IL-10, ↑ IL-22, ↑ IFN-γ, ↑ TNF-α, ↑ TGFβ | ↑ Romboutsia, ↑ Lactobacillus, ↑ Odoribacter, ↓ Parasutterella, ↓ Burkholderia-Caballeronia-Paraburkholderia, ↓ Acinetobacter | [31] |
| Pectin | LPS-induced inflammation in piglets | ↓ TNF-α, ↑ IL-10 | ↓ Helicobacter, ↑ Olsenella, ↑ Bacteroides, ↑ Proteus, ↑ Eubacterium | [32] |
| Rehmannia glutinosa polysaccharide | DSS-induced colitis | ↑tight junction proteins, ↓ IL-10, ↓ IL-6, ↓ TNF-α, ↑ IL-10 | ↓ Bacteroidaceae, ↑ Lactobacillus, ↑ Alistipes, ↑ Lachnospiraceae_NK4A13 | [33] |
| Polygonatum sibiricum polysaccharide | Aged mouse model | ↓ IL-23, ↓ IL-6, ↓ IL-1β, ↓ TNFα, ↓ IL-17, ↓ IL-12, ↓ IL-6, ↑ IL4, ↑ IL-10 | ↑ Bifidobacterium, ↑ Lactobacillus, ↓ Escherichia coli | [34] |
| Rhinacanthus nasutus and okra | AA-induced colitis | ↓ IL1β, ↓ IL-2, ↓ IL-6, ↑ IL-10 | ↑ Muribaculaceae, ↓ Bacteroidaceae, ↓ Tannerellaceae | [35] |
| Houttuynia cordata polysaccharides | DSS-induced colitis | ↓ TNF-α, ↓ IL-1β, ↓ IL-6 | ↑ Firmicutes, ↑ Bacteroides, ↓ Proteobacteria | [36] |
| Lonicera japonica Thunb | DSS-induced colitis | ↑ IL-2, ↑ TNF-α, ↑ IFN-γ | ↑ Bifidobacterium, ↑ Lactobacilli, ↓ Escherichia coli, ↓ Enterococcus | [37] |
| Crataegus pinnatifida | DSS-induced colitis | ↓ IL-1β, ↓ IL-6, ↓ TNF-α | ↑ Alistipes, ↑ Odoribacter | [38] |
| Morinda citrifolia L. | DSS-induced colitis | ↓ TNF-α, ↓ IL-17 | ↑ Dubosiella, ↑ Muribaculaceae, ↑ Ruminococcaceae_UGG-014, ↑ Ruminococcus_1, ↓ Bilophila, ↓ Campylobacter, ↓ Escherichia-Shigella, ↓ Ochrobactrum, ↓ Vibrio | [39] |
| Scutellaria baicalensis Georgi. | DSS-induced colitis | ↓IL-6, ↓IL-1β, ↓TNF-α | ↑ Bifidobacterium, ↑ Firmicutes, ↑ Lactobacillus, ↑ Roseburia | [40] |
| Polysaccharide Intervention | Disease | Anti-Inflammatory Outcomes in Lungs | Effects on Intestinal Microflora | Reference |
|---|---|---|---|---|
| Pear extract | Preclinical asthma mouse model and randomized, double-blind clinical studies | ↓ pro-inflammatory cytokines, including IgE, IL-4, IL-5, and IL-13 | ↑ Bifidobacterium and Eubacterium | [41] |
| Inulin (10%) | OVA- and Al (OH)3-induced asthma in SD rats | Attenuation of the asthmatic inflammatory response in the offspring | ↑ SCFA-producing bacteria (mainly Bifidobacterium) in maternal intestinal microflora; alteration in intestinal microflora composition of offspring | [42] |
| Astragalus membranaceus polysaccharide (25, 50, and 100 mg/kg) | Bleomycin-induced pulmonary fibrosis | ↓ damage and collagen deposition in lung tissue; ↓ inflammatory cytokines TNF-α, IL-6, and IL-1β levels; ↓ apoptosis | Restoration of gut microbiota homeostasis; ↑ Lactobacillus and Akkermansia; ↓ Lachnoclostridium, Clostridium, and Erysipelatoclostridium | [43] |
| Houttuynia cordata polysaccharides (80 mg/kg) and flavonoids (100 mg/kg) either alone or in combination | Influenza virus H1N1-infected mice | Combined therapy showed more potent effect than monotherapy; inhibition of inflammatory-cell infiltration and production of chemokines or pro-inflammatory cytokines such as MCP-1, IL-8, TNF-α, IL-6, and IL-1 β; | Restoration of microflora composition, ↑ Bacteroidetes-to-Firmicutes (B/F) ratio | [44] |
| Cellulose-rich diet (30%) | OVA- and Al(OH)3-induced asthma in C57BL/6J mice | ↓ inflammatory cell infiltration around the bronchus and blood vessels; normalization of airway epithelial structure; ↓IL-4; ↓IgE | ↑Peptostreptococcaceae | [45] |
| Houttuynia cordata polysaccharides (40 mg/kg) | H1N1-induced pneumonia in antibiotic-treated BALB/c mice (termed BALB/c-ABX mice) | Significant amelioration of inflammation in lungs of BALB/c mice | Attenuation of pathological change in intestine; ↓ Bacteroidetes at phylum level; ↓ Bacteroides and ↑ f_Lachnoospiraceae at genus level, ↑ gut microbial diversity; ↑ acetate | [46] |
| Astragalus polysaccharides | Lipopolysaccharide-induced inflammatory lung injury | Alleviation of histopathological abnormalities in lung tissues; ↓ neutrophils infiltration; inhibition of LPS-induced lung inflammation | Change in colonic microbiota composition; ↑ short-chain fatty acid (SCFA)-producing genera such as Oscillospira, Akkermansia, and Coprococcus | [47] |
| Platycodon grandiflorus polysaccharide (75, 150, and 300 mg/kg) and platycodin D alone and together | Chronic bronchitis in SD rats induced via smoking | Improvement in histopathological abnormalities; ↓ excess mucus secretion; improved immunological imbalance in lungs of CB model rat | — | [48] |
| Ephedra sinica polysaccharide | PM- and OVA-induced asthma in mice | ↓ eosinophils in BALF; ↓ serum Ig-E, IL-6, TNF-α, and IL-1β; ↓ airway inflammation | ↑ Bacteroides, Lactobacillus, Prevotella, Butyricicoccus, and Paraprevotella; ↓ Enterococcus and Ruminococcus; ↑ acetic acid, propionic acid, butyric acid, isobutyric acid, valeric acid, isovaleric acid, and isohexanic acid | [49] |
| Polysaccharides from Tetrastigma hemsleyanum Diels et Gilg | LPS-induced ARDS in Balb/c mice | ↓ IL-6, ↓ TNF-α, inhibition of pulmonary inflammation via TLR2/TLR4 pathway | — | [50] |
| Polysaccharide-rich ethanol precipitate fraction of black tea | Particulate matter (PM)-induced lung injury in BALB/c mice | ↓ oxidative stress and inflammation in the lungs; ↓ IL-6, CXCL1, CXCL15, and MDA | ↑ Lachnospiraceae, ↓ Lactobacillaceae | [51] |
| High-cellulose (20%) or high-pectin diet (20) | Cigarette-smoke (CS)-exposed C57BL/6 mouse emphysema model | ↓ Alveolar destruction and inflammation in BALF; ↓ macrophages and neutrophils in BALF; ↓ mRNA expression of IFN-γ, IL-1β, IL-6, IL-8, IL-18, TNF-α, TGF-β | ↑ SCFAs, bile acids, sphingolipids; ↑ Bacteroidetes; ↓ Lactobacillaceae; ↓ Defluviitaleaceae; ↓ Oscillospiraceae | [52] |
| Cellulose- or pectin-enriched diet | Ozone-induced airway hyperresponsiveness in C57BL/6 mice | ↓ ozone-induced AHR, neutrophilic airway inflammation, and airway injury in female but not male mice; cellulose-based diets ameliorated ozone-induced airway hyperresponsiveness in male but not female mice | ↓ Firmicutes, ↑ Proteobacteria and Verrucomicrobia in pectin-fed mice; ↑ Proteus and Lactobacillus, and ↓ Parabacteroides, Clostridiales, and Lachnospiraceae in pectin- versus cellulose-fed mice | [53] |
| Houttuynia cordata polysaccharides | H1N1-induced acute lung injury in C57BL/6 mice | Restoration of Th17/Treg cells balance in the lung, ↓ CCL20 expression | Restoration of Th17/Treg-cell balance of gut mucosa-associated lymphoid tissue (GALT) | [54] |
| Lycium barbarum polysaccharide | Asthma (OVA-induced mouse model) | ↓ lung injury; ↓ TNF, IL-4, IL-6, MCP-1, and IL-17A in plasma and BALF | ↑ Lactobacillus and Bifidobacterium; ↓ Firmicutes, Actinobacteria, Alistipes, and Clostridiales | [55] |
| Soluble fiber (inulin 12 g/day), soluble fiber + probiotic (inulin 12 g/day + multi-strain probiotic >25 billion CFU) | Randomized, double-blind, three-way cross-over trial involving asthmatic patients | No differences between groups in asthma control or airway inflammation | No differences between groups in SCFA levels | [56] |
| Houttuynia cordata polysaccharide (40 mg/kg/day) | H1N1 virus-infected mice | ↓ lung inflammation via inhibition of TLR signaling pathway, and IL-1β production and promotion of IL-10 production | ↓ intestinal barrier damage; ↑ ZO-1 expression; ↓ relative abundance of pathogenic bacterial genera Vibrio and Bacillus | [57] |
| Houttuynia cordata polysaccharide (20 mg/kg; 40 mg/kg) | Influenza A virus (IAV) H1N1-mediated pneumonia in BALB/c mice | Amelioration of pulmonary injury; inhibition of TNF-α, IL-6, IFN-α, RANTES, MCP-1, MIP-1α; and IP-10 production | ↓ intestinal goblet cells; ↑ intestinal physical and immune barrier; ↑ tight junction protein (ZO-1) in intestine | [58] |
| Pectin (30%) | Asthma (ozone-exposed mice) | ↓ ozone-induced airway hyperresponsiveness | ↑ serum short-chain fatty acids | [59] |
| GOS (1 or 2.5 w/w%) alone or with budesonide | HDM-induced asthma in BALB/c mice | Budesonide or GOS: ↓ eosinophils in BALF GOS + budesonide: ↓ CCL17, CCL22, and IL-33 protein levels; ↓ allergic inflammatory response | — | [60] |
| Soluble-fiber meal containing probiotic yoghurt; inulin (3.5 g); and the probiotics Lactobacillus acidophilus, Bifidobacterium lactis, and Lactobacillus rhamnosus | Human subjects with stable asthma | Airway inflammation biomarkers, including sputum total cell count, neutrophils, macrophages, lymphocytes, sputum IL-8, and eNO, significantly decreased; no decrease in airway eosinophils | — | [61] |
| Soluble pectin/insoluble cellulose (4%) | Asthma (female BALB/c mice; OVA induction) | ↓ eosinophil inflammation, ↓ frequency of allergic symptoms, ↓ BALF and NALF total cells and eosinophils, ↓ IL-4 in BALF, ↑ IFN-γ and IL-10 in BALF | ↑ Bifidobacteria | [62] |
| Houttuynia cordata Thunb. polysaccharides (40, 80, and 160 mg/kg) | Lipopolysaccharide-induced acute lung injury in Balb/c mice | ↓ pro-inflammatory cytokine (TNF-α, interleukin-6, and interleukin-1β) production | — | [63] |
| Citrus pectin-derived acidic oligosaccharides (5%) | Pseudomonas aeruginosa lung infection in BALB/c mice | ↑ M1 macrophage activation, ↑ IL-10 release, ↓ TNF-α release | ↑ Escherichia coli, Allobaculum species, Sutturella wadsworthia, Bacteroides vulgatus, Bifidobacterium species, Clostridium difficile, Clostridium ramosum, Clostridium sphenoides; ↑ production of butyrate and propionate | [64] |
| GOS (1%) | HDM-induced asthma in BALB/c mice | ↓ AHR development, ↓ BALF eosinophils, ↓ BALF leukocytes, ↓ CCL5 and IL-13 | — | [65] |
| scGOS/lcFOS (in 9:1 ratio) | OVA-induced asthma in female BALB/c and male C57BL/6 | ↓ OVA-induced AHR; no significant change in the concentrations of IFN-γ, IL-4, IL-5, IL-10, IL-12(p70), IL-13, IL-17, and TNF-α in BAL fluid and plasma | — | [66] |
| High-fiber diet | HDM-induced asthma in female C57BL/6 and BALB/C mice | ↓ total BALF leukocytes, eosinophils, macrophages, and lymphocytes; ↓ IL-4, -5, -13, −10, and IFN-γ; ↓ airway hyperresponsiveness | ↑ Bacteroidetes; ↓ Firmicutes | [67] |
| Apple pectin (30%) | HDM-induced asthma in BALB/c mice | ↓ allergic airway inflammation | Changes in intestinal and lung microbiota; increment in SCFAs; ↑ Bacterioidetes; ↑ Bifidobacteriaceae; ↓ Firmicutes | [68] |
| Chitosan oligosaccharides | OVA-induced asthma in mice | ↓ mRNA and protein levels of IL-4, IL-5, IL-13, TNF-α in lung tissue and BALF | — | [69] |
| FOS (2.5%) | HDM-induced airway inflammation in male C3H/HeN mice | ↓ BALF eosinophils, ↓ IL-5 | — | [70] |
| 1% w/w of 9:1 scGOS: lcFOS; 1% w/w of 83% scGOS/lcFOS + 17% pAOS | OVA-induced asthma in male BALB/c mice | ↓ OVA-induced airway inflammation and hyperresponsiveness, ↓ BALF inflammatory cells | — | [71] |
| Raffinose (50 g/kg) and GOS (50 g/kg) | Allergic airway eosinophilia (OVA-sensitized brown Norway rats) | ↓ IL-4, ↓ IL-5 | ↑ total anaerobic bacteria in the colon | [72] |
| Asian pear pectin (100 µg) | Asthma (OVA-sensitized murine model) | ↓ asthmatic reactions in sensitized mice, ↓ IFN-γ, ↓ IL-5 | — | [73] |
| Raffinose (50 g/kg) | Allergic airway eosinophilia (OVA-sensitized brown Norway rats) | ↓ IL-4 and IL-5 mRNA expression, ↓ mucus-producing cells, = IFN-γ mRNA expression | ↑ bifidobacteria | [74] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Y.; Song, M.; Wu, S.; Zhao, H.; Zhang, Y. Plant-Based Dietary Fibers and Polysaccharides as Modulators of Gut Microbiota in Intestinal and Lung Inflammation: Current State and Challenges. Nutrients 2023, 15, 3321. https://doi.org/10.3390/nu15153321
Shen Y, Song M, Wu S, Zhao H, Zhang Y. Plant-Based Dietary Fibers and Polysaccharides as Modulators of Gut Microbiota in Intestinal and Lung Inflammation: Current State and Challenges. Nutrients. 2023; 15(15):3321. https://doi.org/10.3390/nu15153321
Chicago/Turabian StyleShen, Yu, Mingming Song, Shihao Wu, Hongbo Zhao, and Yu Zhang. 2023. "Plant-Based Dietary Fibers and Polysaccharides as Modulators of Gut Microbiota in Intestinal and Lung Inflammation: Current State and Challenges" Nutrients 15, no. 15: 3321. https://doi.org/10.3390/nu15153321
APA StyleShen, Y., Song, M., Wu, S., Zhao, H., & Zhang, Y. (2023). Plant-Based Dietary Fibers and Polysaccharides as Modulators of Gut Microbiota in Intestinal and Lung Inflammation: Current State and Challenges. Nutrients, 15(15), 3321. https://doi.org/10.3390/nu15153321