
Effects of Insect Consumption on Human Health: A Systematic Review of Human Studies

Abstract
1. Introduction
2. Materials and Methods
2.1. Search Strategy and Eligibility Criteria
2.2. Data Extraction and Assessment of Risk of Bias
3. Results
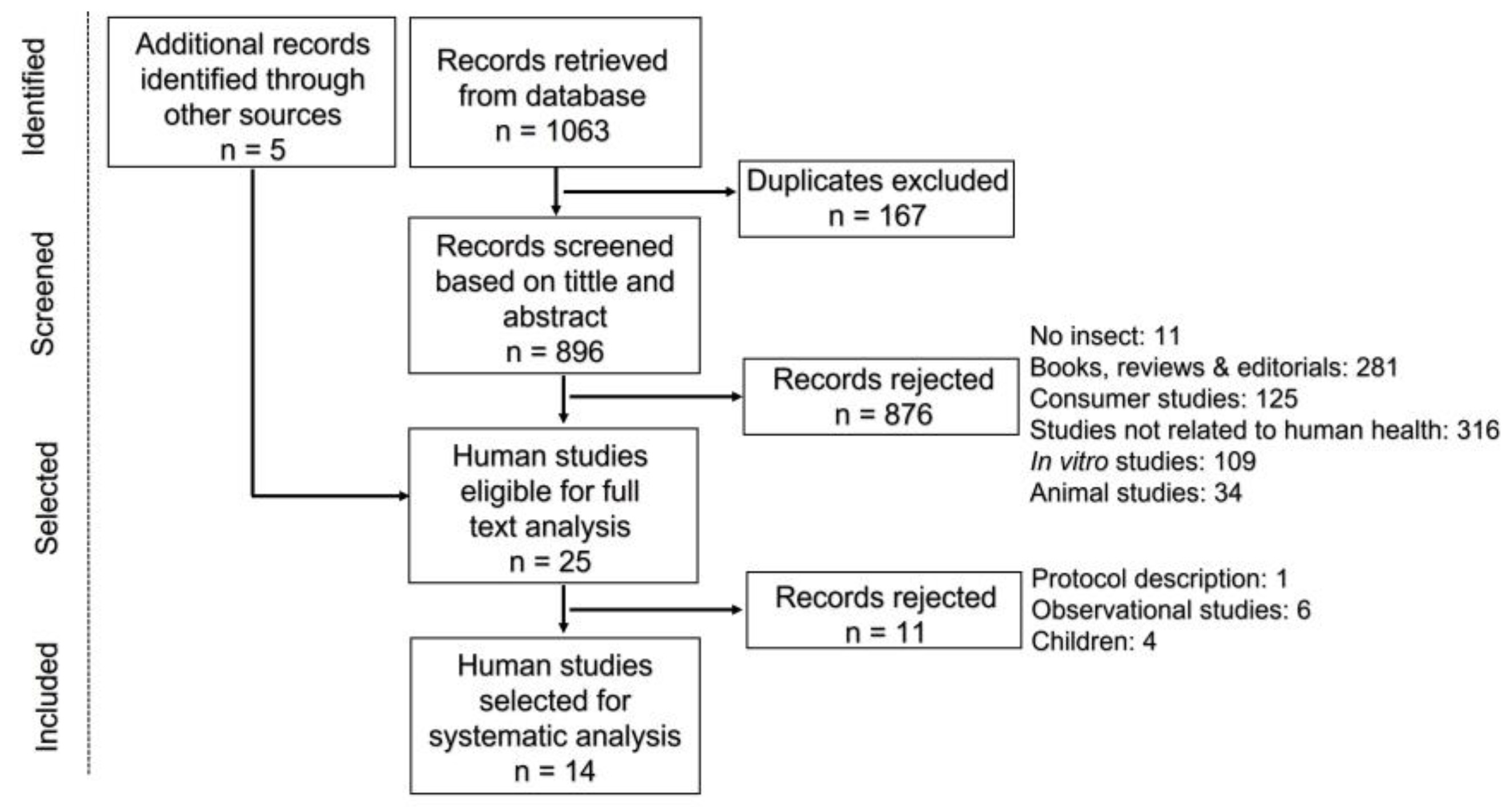
3.1. Selected Studies
3.2. Oral Interventions with Randomized Controlled Trials (RCTs)
3.2.1. Characteristics of the RCTs
3.2.2. Iron Absorption
3.2.3. Amino Acid Absorption and Muscle Protein Synthesis
3.2.4. Biomarkers of Metabolic Diseases
3.2.5. Appetite Regulation
3.2.6. Microbiota
3.2.7. Disease Treatment
3.3. Allergenicity Assessment Studies
3.3.1. Case Study
3.3.2. Epidemiological Studies
3.3.3. Cross-Reactivity Studies
4. Discussion
4.1. Beneficial Health Effects
4.1.1. Nutrient Absorption
4.1.2. Satiety
4.1.3. Metabolic and Inflammatory Biomarkers
4.1.4. Microbiota
4.2. Edible Insect Allergies
4.2.1. Tropomyosin and Arginine Kinase
4.2.2. Other Allergens
4.2.3. Effects of Food-Processing Technologies Used to Reduce Allergies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nowakowski, A.C.; Miller, A.C.; Miller, M.E.; Xiao, H.; Wu, X. Potential health benefits of edible insects. Crit. Rev. Food Sci. Nutr. 2022, 62, 3499–3508. [Google Scholar] [CrossRef]
- Ordoñez-Araque, R.; Egas-Montenegro, E. Edible insects: A food alternative for the sustainable development of the planet. Int. J. Gastron. Food Sci. 2021, 23, 100304. [Google Scholar] [CrossRef]
- Costa-Neto, E.M.; Dunkel, E.V. Insects as Food: History, Culture, and Modern Use around the World. In Insects as Sustainable Food Ingredients, 1st ed.; Dossey, A.T., Morales-Ramos, J.A., Rojas, M.G., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 29–60. [Google Scholar]
- Orkusz, A. Edible Insects versus Meat—Nutritional Comparison: Knowledge of Their Composition Is the Key to Good Health. Nutrients 2021, 13, 1207. [Google Scholar] [CrossRef]
- Mishyna, M.; Glumac, M. So different, yet so alike Pancrustacea: Health benefits of insects and shrimps. J. Funct. Foods 2021, 76, 104316. [Google Scholar] [CrossRef]
- Kuljanic, N.; Gregory-Manning, S. What If Insects Were on the Menu in Europe? Scientic Foresight Unit. European Parliamentary Research Service. 2020. Available online: https://www.europarl.europa.eu/RegData/etudes/ATAG/2020/641551/EPRS_ATA(2020)641551_EN.pdf (accessed on 2 June 2023).
- Van Huis, A.; Oonincx, D. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 43. [Google Scholar] [CrossRef]
- Araújo, R.R.S.; Benfica, T.A.R.S.; Ferraz, V.P.; Santos, E.M. Nutritional composition of insects Gryllus assimilis and Zophobas morio: Potential foods harvested in Brazil. J. Food Compost. Anal. 2019, 76, 22–26. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Cruz-Monterrosa, R.G.; Liceaga, A.M. Beyond Human Nutrition of Edible Insects: Health Benefits and Safety Aspects. Insects 2022, 13, 1007. [Google Scholar] [CrossRef]
- FAO Food and Agriculture Organization of the United Nations. Looking at Edible Insects from a Food Safety Perspective. Challenges and Opportunities for the Sector; FAO: Rome, Italy, 2021. [Google Scholar] [CrossRef]
- Imathiu, S. Benefits and food safety concerns associated with consumption of edible insects. NFS J. 2020, 18, 1–11. [Google Scholar] [CrossRef]
- Wendin, K.M.E.; Nyberg, M.E. Factors influencing consumer perception and acceptability of insect-based foods. Curr. Opin. Food Sci. 2021, 40, 67–71. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, D.; Zhou, S.; Duan, H.; Guo, J.; Yan, W. Nutritional Composition, Health Benefits, and Application Value of Edible Insects: A Review. Foods 2022, 11, 3961. [Google Scholar] [CrossRef]
- Costa-Neto, E.M.; Ramos-Elorduy, J. Los insectos comestibles de Brasil: Etnicidade, diversidade e importância en la alimentación. Bull. Entomol. Soc. Aragonesa 2006, 38, 423–442. [Google Scholar]
- Banjo, A.D.; Lawal, O.A.; Owolana, O.A.; Olubanjo, O.A.; Ashidi, J.S.; Dedeke, G.A.; Soewu, D.A.; Owa, D.A.; Sobowale, O.A. An ethno-zoological survey of insects and their allies among the remos (Ogun State) South Western Nigeria. IAJIKS 2009, 2, 104–111. [Google Scholar] [CrossRef]
- Roos, N.; Huis, A. Consuming insects: Are there health benefits? J. Insects as Food Feed 2017, 3, 225–229. [Google Scholar] [CrossRef]
- EFSA Scientific Committee. Scientific Opinion on a risk profile related to production and consumption of insects as food and feed. EFSA J. 2015, 1, 245–247. [Google Scholar] [CrossRef]
- Lange, K.; Nakamura, Y. Edible insects as future food: Chances and challenges. J. Future Foods 2021, 1, 38–46. [Google Scholar] [CrossRef]
- Sicherer, S.H.; Warren, C.M.; Dant, C.; Gupta, R.S.; Nadeau, K.C. Food Allergy from Infancy Through Adulthood. J. Allergy Clin. Immunol. Pract. 2020, 8, 1854–1864. [Google Scholar] [CrossRef]
- Taylor, G.; Wang, N. Entomophagy and allergies: A study of the prevalence of entomophagy and related allergies in a population living in North-Eastern Thailand. Biosci. Horiz. Int. J. Stud. Res. 2018, 11, hzy003. [Google Scholar] [CrossRef]
- Ji, K.; Chen, J.; Li, M.; Liu, Z.; Wang, C.; Zhan, Z.; Wu, X.; Xia, Q. Anaphylactic shock and lethal anaphylaxis caused by food consumption in China. Trends Food Sci Technol. 2009, 20, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Moher, D. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Syst. Rev. 2021, 10, 89. [Google Scholar] [CrossRef]
- Higgins, J.P.T.; Savović, J.; Page, M.J.; Elbers, R.G.; Sterne, J.A.C. Assessing Risk of Bias in a Randomized Trial. In Cochrane Handbook for Systematic Reviews of Interventions, version 6.3 (updated February 2022); Higgins, J.P.T., Thomas, J., Chandler, J., Cumpston, M., Li, T., Page, M.J., Welch, V.A., Eds.; Cochrane Training: London, UK, 2022; Available online: www.training.cochrane.org/handbook (accessed on 10 March 2023).
- Stull, V.J.; Finer, E.; Bergmans, R.S.; Febvre, H.P.; Longhurst, C.; Manter, D.K.; Patz, J.A.; Weir, T.L. Impact of Edible Cricket Consumption on Gut Microbiota in Healthy Adults, a Double-blind, Randomized Crossover Trial. Sci. Rep. 2018, 8, 10762. [Google Scholar] [CrossRef]
- Vangsoe, M.T.; Thogersen, R.; Bertram, H.C.; Heckmann, L.L.; Hansen, M. Ingestion of Insect Protein Isolate Enhances Blood Amino Acid Concentrations Similar to Soy Protein in A Human Trial. Nutrients 2018, 10, 1357. [Google Scholar] [CrossRef] [PubMed]
- Vangsoe, M.T.; Joergensen, M.S.; Heckmann, L.L.; Hansen, M. Effects of Insect Protein Supplementation during Resistance Training on Changes in Muscle Mass and Strength in Young Men. Nutrients 2018, 10, 335. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Shi, Q.; Ying, S.; Zhu, D.; Chen, H.; Yang, X.; Xu, J.; Xu, F.; Tao, F.; Xu, B. Effects of compound Caoshi silkworm granules on stable COPD patients and their relationship with gut microbiota: A randomized controlled trial. Medicine 2020, 99, e20511. [Google Scholar] [CrossRef] [PubMed]
- Hermans, W.J.H.; Senden, J.M.; Churchward-Venne, T.A.; Paulussen, K.J.M.; Fuchs, C.J.; Smeets, J.S.J.; van Loon, J.J.A.; Verdijk, L.B.; van Loon, L.J.C. Insects are a viable protein source for human consumption: From insect protein digestion to postprandial muscle protein synthesis in vivo in humans: A double-blind randomized trial. Am. J. Clin. Nutr. 2021, 114, 934–944. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Lov, J.; Martin-Arrowsmith, P.W.; Gritsas, A.; Churchward-Venne, T.A. The acute effects of insect vs. beef-derived protein on postprandial plasma aminoacidemia, appetite hormones, appetite sensations, and energy intake in healthy young men. Eur. J. Clin. Nutr. 2022, 76, 1548–1556. [Google Scholar] [CrossRef]
- Miguéns-Gómez, A.; Sierra-Cruz, M.; Rodríguez-Gallego, E.; Beltrán-Debón, R.; Blay, M.T.; Terra, X.; Pinent, M.; Ardévol, A. Effect of an Acute Insect Preload vs. an Almond Preload on Energy Intake, Subjective Food Consumption and Intestinal Health in Healthy Young Adults. Nutrients 2022, 14, 1463. [Google Scholar] [CrossRef] [PubMed]
- Mwangi, M.N.; Oonincx, D.G.A.B.; Hummel, M.; Utami, D.A.; Gunawan, L.; Veenenbos, M.; Zeder, C.; Cercamondi, C.I.; Zimmermann, M.B.; van Loon, J.J.A.; et al. Absorption of iron from edible house crickets: A randomized crossover stable-isotope study in humans. Am. J. Clin. Nutr. 2022, 116, 1146–1156. [Google Scholar] [CrossRef] [PubMed]
- Skotnicka, M.; Mazurek, A.; Karwowska, K.; Folwarski, M. Satiety of Edible Insect-Based Food Products as a Component of Body Weight Control. Nutrients 2022, 14, 2147. [Google Scholar] [CrossRef]
- Beaumont, P.; Courtois, J.; Van der Brempt, X.; Tollenaere, S. Food-induced anaphylaxis to Tenebrio molitor and allergens implicated. Rev. Fr. Allergol. 2019, 59, 389–393. [Google Scholar] [CrossRef]
- Ndlovu, V.; Chimbari, M.; Sibanda, E.; Ndarukwa, P. A feasibility study to assess Imbrasia belina (mopane worm) sensitisation and related respiratory health outcomes in a rural community in Gwanda district, Zimbabwe. Pilot Feasibility Stud. 2021, 7, 55. [Google Scholar] [CrossRef]
- Verhoeckx, K.C.M.; van Broekhoven, S.; den Hartog-Jager, C.F.; Gaspari, M.; de Jong, G.A.H.; Wichers, H.J.; van Hoffen, E.; Houben, G.F.; Knulst, A.C. House dust mite (Der p 10) and crustacean allergic patients may react to food containing Yellow mealworm proteins. Food Chem. Toxicol. 2014, 65, 364–373. [Google Scholar] [CrossRef]
- Kamemura, N.; Sugimoto, M.; Tamehiro, N.; Adachi, R.; Tomonari, S.; Watanabe, T.; Mito, T. Cross-allergenicity of crustacean and the edible insect Gryllus bimaculatus in patients with shrimp allergy. Mol. Immunol. 2019, 106, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Lamberti, C.; Nebbia, S.; Cirrincione, S.; Brussino, L.; Giorgis, V.; Romito, A.; Marchese, C.; Manfredi, M.; Marengo, E.; Giuffrida, M.G.; et al. Thermal processing of insect allergens and IgE cross-recognition in Italian patients allergic to shrimp, house dust mite and mealworm. Food Res. Int. 2021, 148, 110567. [Google Scholar] [CrossRef] [PubMed]
- Stull, V.J. Impacts of insect consumption on human health. J. Insects Food Feed. 2021, 7, 695–713. [Google Scholar] [CrossRef]
- Ros-Baró, M.; Casas-Agustench, P.; Díaz-Rizzolo, D.A.; Batlle-Bayer, L.; Adrià-Acosta, F.; Aguilar-Martínez, A.; Medina, F.X.; Pujolà, M.; Bach-Faig, A. Edible Insect Consumption for Human and Planetary Health: A Systematic Review. Int. J. Environ. Res. Public Health 2022, 19, 11653. [Google Scholar] [CrossRef]
- Van Huis, A. Edible insects: Challenges and prospects. Entomol. Res. 2022, 52, 161–177. [Google Scholar] [CrossRef]
- Groen, B.B.; Horstman, A.M.; Hamer, H.M.; de Haan, M.; van Kranenburg, J.; Bierau, J.; Poeze, M.; Wodzig, W.K.; Rasmussen, B.B.; van Loon, L.J. Post-Prandial Protein Handling: You Are What You Just Ate. PLoS ONE 2015, 10, e0141582. [Google Scholar] [CrossRef]
- Boirie, Y.; Dangin, M.; Gachon, P.; Vasson, M.P.; Maubois, J.L.; Beaufrère, B. Slow and fast dietary proteins differently modulate postprandial protein accretion. Proc. Natl. Acad. Sci. USA 1997, 94, 14930–14935. [Google Scholar] [CrossRef]
- Landi, F.; Calvani, R.; Tosato, M.; Martone, A.M.; Ortolani, E.; Savera, G.; D’Angelo, E.; Sisto, A.; Marzetti, E. Protein Intake and Muscle Health in Old Age: From Biological Plausibility to Clinical Evidence. Nutrients 2016, 8, 295. [Google Scholar] [CrossRef]
- Bruttomesso, D.; Tessari, P. A High-Fiber Diet Decreases Postabsorptive Protein Turnover but Does Not Alter Insulin Sensitivity in Men with Type 1 Diabetes Mellitus. J. Nutr. 2019, 149, 596–604. [Google Scholar] [CrossRef]
- Gorissen, S.H.M.; Trommelen, J.; Kouw, I.W.K.; Holwerda, A.M.; Pennings, B.; Groen, B.B.L.; Wall, B.T.; Churchward-Venne, T.A.; Horstman, A.M.H.; Koopman, R.; et al. Protein Type, Protein Dose, and Age Modulate Dietary Protein Digestion and Phenylalanine Absorption Kinetics and Plasma Phenylalanine Availability in Humans. J. Nutr. 2020, 150, 2041–2050. [Google Scholar] [CrossRef]
- Trommelen, J.; Holwerda, A.; Pinckaers, P.; Van Loon, L. Comprehensive assessment of post-prandial protein handling by the application of intrinsically labelled protein in vivo in human subjects. Proc. Nutr. Soc. 2021, 80, 221–229. [Google Scholar] [CrossRef]
- Gorissen, S.H.; Rémond, D.; van Loon, L.J. The muscle protein synthetic response to food ingestion. Meat Sci. 2015, 109, 96–100. [Google Scholar] [CrossRef]
- Kouřimská, L.; Adámková, A. Nutritional and sensory quality of edible insects. NFS J. 2016, 4, 22–26. [Google Scholar] [CrossRef]
- Ahmad, R.S.; Imran, A.; Hussain, M.B. Nutritional Composition of Meat. In Meat Science and Nutrition, 1st ed.; Arshad, M.S., Ed.; InTech Open: London, UK, 2018. [Google Scholar] [CrossRef]
- Morell, P.; Fiszman, S. Revisiting the role of protein-induced satiation and satiety. Food Hydrocoll. 2017, 68, 199–210. [Google Scholar] [CrossRef]
- Bowen, J.; Noakes, M.; Clifton, P.M. Appetite Regulatory Hormone Responses to Various Dietary Proteins Differ by Body Mass Index Status Despite Similar Reductions in ad Libitum Energy Intake. J. Clin. Endocr. 2006, 91, 2913–2919. [Google Scholar] [CrossRef]
- Kim, T.K.; Yong, H.I.; Kim, Y.B.; Kim, H.W.; Choi, Y.S. Edible Insects as a Protein Source: A Review of Public Perception, Processing Technology, and Research Trends. Food Sci. Anim. Resour. 2019, 39, 521–540. [Google Scholar] [CrossRef]
- Roncolini, A.; Milanović, V.; Aquilanti, L.; Cardinali, F.; Garofalo, C.; Sabbatini, R.; Clementi, F.; Belleggia, L.; Pasquini, M.; Mozzon, M. Lesser mealworm (Alphitobius diaperinus) powder as a novel baking ingredient for manufacturing high-protein, mineral-dense snacks. Food Res. Int. 2020, 131, 109031. [Google Scholar] [CrossRef]
- Leidy, H.J.; Armstrong, C.L.; Tang, M.; Mattes, R.D.; Campbell, W.W. The influence of higher protein intake and greater eating frequency on appetite control in overweight and obese men. Obesity 2010, 18, 1725–1732. [Google Scholar] [CrossRef]
- Rigamonti, A.E.; Tamini, S.; Cicolini, S.; De Col, A.; Caroli, D.; Mai, S.; Rondinelli, E.; Saezza, A.; Cella, S.G.; Sartorio, A. Evaluation of an Amino Acid Mix on the Secretion of Gastrointestinal Peptides, Glucometabolic Homeostasis, and Appetite in Obese Adolescents Administered with a Fixed-Dose or ad Libitum Meal. J. Clin. Med. 2020, 9, 3054. [Google Scholar] [CrossRef]
- Köhler, R.; Kariuki, L.; Lambert, C.; Biesalski, H.K. Protein, amino acid and mineral composition of some edible insects from Thailand. J. Asia Pac. Entomol. 2019, 22, 372–378. [Google Scholar] [CrossRef]
- Park, S.A.; Lee, G.H.; Lee, H.Y.; Hoang, T.H.; Chae, H.J. Glucose-lowering effect of Gryllus bimaculatus powder on streptozotocin-induced diabetes through the AKT/mTOR pathway. Food Sci. Nutr. 2019, 8, 402–409. [Google Scholar] [CrossRef]
- Escobar-Ortiz, A.; Hernández-Saavedra, D.; Lizardi-Mendoza, J.; Pérez-Ramírez, I.F.; Mora-Izaguirre, O.; Ramos-Gómez, M.; Reynoso-Camacho, R. Consumption of cricket (Acheta domesticus) flour decreases insulin resistance and fat accumulation in rats fed with high-fat and -fructose diet. J. Food Biochem. 2022, 46, e14269. [Google Scholar] [CrossRef]
- Galland, L. Diet and inflammation. Nutr. Clin. Pract. 2010, 25, 634–640. [Google Scholar] [CrossRef]
- Hess, J.M.; Stephensen, C.B.; Kratz, M.; Bolling, B.W. Exploring the Links between Diet and Inflammation: Dairy Foods as Case Studies. Adv. Nutr. 2021, 12, 1S–13S. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Reyes, A.; Rosell, C.M.; Rodrigo, D.; Ibarra-Herrera, C.C. Benefits and Challenges in the Incorporation of Insects in Food Products. Front. Nutr. 2021, 8, 687712. [Google Scholar] [CrossRef]
- Quah, Y.; Tong, S.R.; Bojarska, J.; Giller, K.; Tan, S.A.; Ziora, Z.M.; Esatbeyoglu, T.; Chai, T.T. Bioactive Peptide Discovery from Edible Insects for Potential Applications in Human Health and Agriculture. Molecules 2023, 28, 1233. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes. Nutrients 2010, 2, 355–374. [Google Scholar] [CrossRef]
- Illahi, M.I.; Amjad, S.; Alam, S.M.; Ahmed, S.T.; Fatima, M.; Shahid, M.A. Serum Tumor Necrosis Factor-Alpha as a Competent Biomarker for Evaluation of Disease Activity in Early Rheumatoid Arthritis. Cureus 2021, 13, e15314. [Google Scholar] [CrossRef]
- Lebeer, S.; Claes, I.; Tytgat, H.L.P.; Verhoeven, T.L.A.; Marien, E.; Ossowski, I.; Reunanen, J.; Palva, A.; de Vos, W.M.; De Keersmaecker, S.C.J.; et al. Functional Analysis of Lactobacillus rhamnosus GG Pili in Relation to Adhesion and Immunomodulatory Interactions with Intestinal Epithelial Cells. Appl. Environ. Microbiol. 2012, 78, 185–193. [Google Scholar] [CrossRef]
- Turroni, F.; Peano, C.; Pass, D.A.; Foroni, E.; Severgnini, M.; Claesson, M.J.; Kerr, C.; Hourihane, J.; Murray, D.; Fuligni, F.; et al. Diversity of Bifidobacteria within the infant gut microbiota. PLoS ONE 2012, 7, e36957. [Google Scholar] [CrossRef]
- Saeb, A.; Grundmann, S.M.; Gessner, D.K.; Schuchardt, S.; Most, E.; Wen, G.; Eder, K.; Ringseis, R. Feeding of cuticles from Tenebrio molitor larvae modulates the gut microbiota and attenuates hepatic steatosis in obese Zucker rats. Food Funct. 2022, 13, 1421–1436. [Google Scholar] [CrossRef]
- Verbeke, K.A.; Boobis, A.R.; Chiodini, A.; Edwards, C.A.; Franck, A.; Kleerebezem, M.; Nauta, A.; Raes, J.; van Tol, E.A.; Tuohy, K.M. Towards microbial fermentation metabolites as markers for health benefits of prebiotics. Nutr. Res. Rev. 2015, 28, 42–66. [Google Scholar] [CrossRef]
- Vinolo, M.A.; Ferguson, G.J.; Kulkarni, S.; Damoulakis, G.; Anderson, K.; Bohlooly-Y, M.; Stephens, L.; Hawkins, P.T.; Curi, R. SCFAs induce mouse neutrophil chemotaxis through the GPR43 receptor. PLoS ONE 2011, 6, e21205. [Google Scholar] [CrossRef]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef]
- De Martinis, M.; Sirufo, M.M.; Suppa, M.; Ginaldi, L. New Perspectives in Food Allergy. Int. J. Mol. Sci. 2020, 21, 1474. [Google Scholar] [CrossRef]
- Peters, R.L.; Krawiec, M.; Koplin, J.J.; Santos, A.F. Update on food allergy. Pediatr. Allergy Immunol. 2021, 32, 647–657. [Google Scholar] [CrossRef]
- Warren, C.M.; Jiang, J.; Gupta, R.S. Epidemiology and Burden of Food Allergy. Curr. Allergy Asthma Rep. 2020, 20, 6. [Google Scholar] [CrossRef]
- De Marchi, L.; Wangorsch, A.; Zoccatelli, G. Allergens from Edible Insects: Cross-reactivity and Effects of Processing. Curr. Allergy Asthma Rep. 2021, 21, 35. [Google Scholar] [CrossRef]
- Popescu, F.D. Cross-reactivity between aeroallergens and food allergens. World J. Methodol. 2015, 5, 31–50. [Google Scholar] [CrossRef]
- Ribeiro, J.C.; Sousa-Pinto, B.; Fonseca, J.; Caldas Fonseca, S.; Cunha, L.M. Edible insects and food safety: Allergy. J. Insects Food Feed. 2021, 7, 833–847. [Google Scholar] [CrossRef]
- Worm, M.; Jappe, U.; Kleine-Tebbe, J.; Seschäfer, C.; Reese, I.; Saloga, J.; Treudler, R.; Zuberbier, T.; Wassmann, A.; Fuchs, T.; et al. Food allergies resulting from immunological cross-reactivity with inhalant allergens: Guidelines from the German Society for Allergology and Clinical Immunology (DGAKI), the German Dermatology Society (DDG), the Association of German Allergologists (AeDA) and the Society for Pediatric Allergology and Environmental Medicine (GPA). Allergo J. Int. 2014, 23, 1–16. [Google Scholar] [CrossRef]
- Celi, G.; Brusca, I.; Scala, E.; Villalta, D.; Pastorello, E.; Farioli, L.; Cortellini, G.; Deleonardi, G.; Galati, P.; Losappio, L.; et al. House dust mite allergy and shrimp allergy: A complex interaction. Eur. Ann. Allergy Clin. Immunol. 2020, 52, 205–209. [Google Scholar] [CrossRef]
- Reese, G.; Ayuso, R.; Lehrer, S.B. Tropomyosin: An invertebrate pan-allergen. Int. Arch. Allergy Immunol. 1999, 119, 247–258. [Google Scholar] [CrossRef]
- Rougé, P.; Barre, A. Allergy to Edible Insects: A Computational Identification of the IgE-Binding Cross-Reacting Allergen Repertoire of Edible Insects. In Future Foods, 1st ed.; Mikkola, H., Ed.; InTech Open: London, UK, 2017; Available online: https://www.intechopen.com/chapters/54930 (accessed on 2 June 2023). [CrossRef]
- Srinroch, C.; Srisomsap, C.; Chokchaichamnankit, D.; Punyarit, P.; Phiriyangkul, P. Identification of novel allergen in edible insect, Gryllus bimaculatus and its cross-reactivity with Macrobrachium spp. allergens. Food Chem. 2015, 184, 160–166. [Google Scholar] [CrossRef]
- Delgado Calvo-Flores, L.; Garino, C.; Moreno, F.J.; Broll, H. Insects in food and their relevance regarding allergenicityassessment. EFSA J. 2022, 20, e200909. [Google Scholar] [CrossRef]
- Barre, A.; Pichereaux, C.; Velazquez, E.; Maudouit, A.; Simplicien, M.; Garnier, L.; Bienvenu, F.; Bienvenu, J.; Burlet-Schiltz, O.; Auriol, C.; et al. Insights into the Allergenic Potential of the Edible Yellow Mealworm (Tenebrio molitor). Foods 2019, 8, 515. [Google Scholar] [CrossRef]
- De Las Marinas, M.D.; Cerdá, J.C.; López-Matas, M.A.; González-Ruiz, A.; Martorell, C.; Felix, R.; Alvariño, M.; Carnés, J. Hexamerin-like protein 2, a cricket allergen involved in occupational and food allergy. Clin. Exp. Allergy 2021, 51, 858–860. [Google Scholar] [CrossRef]
- Ganseman, E.; Ieven, T.; Frans, G.; Coorevits, L.; Pörtner, N.; Martens, E.; Bullens, D.M.; Schrijvers, R.; Breynaert, C.; Proost, P. Alpha-amylase as the culprit in an occupational mealworm allergy case. Front. Allergy 2022, 3, 992195. [Google Scholar] [CrossRef]
- Liguori, B.; Sancho, A.I.; Poulsen, M.; Bøgh, K.L. Novel foods: Allergenicity assessment of insect proteins. EFSA J. 2022, 20, e200910. [Google Scholar] [CrossRef]


{kind=link}
| Author, Year, (Country) | Type of RCT | Test | Control | Participants | Intervention | Outcomes Data Collection | Results |
|---|---|---|---|---|---|---|---|
| Iron Absorption | |||||||
| Mwangi et al., 2022 (Netherlands) [31] | Crossover, acute (1 day) | 800–900 g porridge meal with low phytate or high phytate maize flour + 100 g of [57Fe]-labeled ground Cr or 50 g of unlabeled ground Cr. | Placebo = 800–900 g porridge meal with low phytate or high phytate maize flour. Fe content matched by the addition of FeSO4 | 20 Females, iron-depleted, mean age 24.7 ± 2.9 years; mean BMI 21.6 ± 1.9 kg/m2 | Administration of different meals on consecutive days, after overnight fast, with a 14-day interval between insect and placebo meals. Meals ingested in two portions of 400–450 g each, served at breakfast and ≥3 h after. Lunch is provided after the second serving. | Hemoglobin, serum ferritin, STR, fractional iron absorption. Blood samples collected on days 0, 23, and 39 after the ingestion. | ↔ Hemoglobin ↔ Serum ferritin. ↑ STR for insect compared to placebo. ↔ Fractional iron absorption in high phytate meals. ↓ Fractional iron absorption for low phytate meal with labeled or unlabelled insect, compared to low phytate meal placebo. |
| Amino acid Absorption and muscle protein synthesis | |||||||
| Hermans et al., 2021 (Netherlands) [28] | Parallel, acute (1 day) | 64 g of Lw protein powder in 300 mL water (30 g protein, 312 kcal). | Alternate protein source: 40 g of dried milk protein concentrate in 300 mL water (30 g protein, 142 kcal) | 24 Males, healthy, mean age 23.0 ± 3.0 years; BMI 23.1 ± 2.7 Kg/m2 | Exercise (one leg exercised and one leg at rest) after overnight fast, followed by administration of insect or milk protein. | Plasma amino acids (EAA, NEAA, TAA, Leu, Phe, Tyr), glucose, insulin, muscle protein synthesis rate. Blood samples collected before and after exercise, and at postprandial states (20 to 300 min); muscle biopsy collected for rest and exercised leg after exercise, and 120, 300 min after ingestion of insect or control. | ↔ Glucose and insulin. ↓ Peak Leu, Phe, EAA, NEAA, TAA for insect compared to milk protein. ↑Peak Tyr for insect, compared to milk protein. ↔ AUC EAA, NEAA and TAA. Amino acid plasma levels peaked at 30 min for milk and at 60 min for insect. ↔ Protein synthesis rate at rest and after exercise. |
| Vangsoe et al., 2018 (Denmark) [25] | Crossover, Acute (1 day) | 30.5 g of protein isolate from Lw (25 g protein) in 400 mL water (100 Kcal). | Placebo = 400 mL of water Alternative protein sources: C1 = 25 g of soy protein isolate in 400 mL of water C2 = 25 g of whey protein isolate in 400 mL of water | Six Males, healthy, mean age 24 ± 1 years. | Administration of products on four different days, after overnight fast, with one-day intervals between each intervention. Ingestion of insect, soy, whey, and placebo products within 1 min. | Plasma amino acids (EAA, BCAA, Leu) and insulin. Blood samples collected at pre-prandial (0 min) and postprandial states (20 to 120 min). | ↑ AUC of EAA, BCAA, and Leu for all protein sources compared to placebo. ↑ AUC of EAA, BCAA, and Leu for whey, compared to soy and insect. Amino acid plasma levels peaked at 60 min for whey and soy and at 120 min for insect. ↓ Insulin at 20 and 40 min for insect compared to whey and soy. |
| Vangsoe et al., 2018 (Denmark) [26] | Parallel, chronic (8 weeks) | Protein bar of banana, ginger, and oats, supplemented with 0.4 g of Lw protein/kg body weight (137 kcal). | Placebo = Isocaloric bar with no insect protein (3.6 g protein). | 18 Males, healthy, mean age 24.2 ± 2.6 years; body weight 79.9 ± 9.0 kg, height 186.6 ± 6.6 cm | Two insect or placebo bars per day: one bar 1h after resistance training and one bar 1 h before sleep on training days, 4 days a week. | Body composition (BW, FM, BMC, FBFM), muscle strength (1 RM leg and bench press), energy, and macronutrient intake. Body composition (DXA) and muscle strength measured 1 week before the start and 2 days after the last training session; 3-day dietary records before and during the intervention. | ↔ BW, FM, FBFM, and strength. ↑ Protein intake for insect compared to placebo (2.3 g/kg/day for test, 1.7 g/kg/day for control) ↓ CHO intake for insect compared to placebo (4.8 g/kg/day, for test, 5.8 g/kg/day for control) ↔ Energy and fat intake |
| Amino acid Absorption and Appetite Regulation | |||||||
| Dai et al., 2022 (Canada) [29] | Crossover, acute | 400 mL beverage with 25 g of Cr-derived protein. | Alternative protein source: 400 mL beverage with 25 g of beef-derived protein | 20 Males, healthy, mean age 23 ± 4 years. | Administration of beverages after overnight fast, with a 7-day interval between insect and beef. Beverages drank in 5 min; ad libitum meal after 300 min. | Plasma amino acids (BCAA, EAA, NEAA, TAA, Leu), glucose, insulin, perceived hunger, fullness, desire to eat, PFC, satiety hormones (GLP-1, PYY). Self-filled rating questionnaires (VAS) and blood samples, before ingestion and postprandial every 15 min, until 300 min. | ↔ Glucose, GLP-1, and PYY. ↓ Insulin for insect. ↑ AUC of Leu, BCAA and EAA for insect ↓ AUC of NEAA and TAA for insect. Amino acid plasma levels peaked at approx 60 to 80 min for beef and at approx 90 to 100 min for insect. ↓ Hunger for beef. ↔ Fullness, desire to eat, and PFC. ↔ Energy intake after ad libitum meal. |
| Miguéns-Gómez et al., 2022 (Spain) [30] | Crossover, acute (1 day) | 170 g Cocoa milkshake with 25 g of Lw powder (20 g protein; 194.1 kcal). | Placebo = 145 g Cocoa milkshake (5.7 g protein; 66.7 kcal) Alternative protein source: 205 g Cocoa milkshake with 60 g almond flour (20.1 g protein; 443.2 kcal) | 12 Females, 17 Males, healthy, 22 to 33 years, BMI < 40 kg/m2 | Administration of a preload of insect, almond, and placebo on separate days, after overnight fasting, with 6–7-day intervals; 1 h and 4 h after preload, ad libitum breakfast and lunch. | Subjective desire to eat, PFC, feelings of indigestion, energy and protein intake after ad libitum meal. Self-filled rating questionnaires (VAS) before ingestion and every 1 h after, until 8 h. | ↓Desire to eat, for insect and almond compared to placebo, 1 h after preload. ↓PFC, for insect and almond compared to placebo, 1 h and 5 h after preload. ↔ Desire to eat and PFC for insect compared to almond. ↔ Cumulative energy and protein intake (breakfast + lunch). ↑ Sensation of indigestion for insect, compared to almond and placebo. |
| Skotnicka et al., 2022 (Poland) [32] | Crossover, acute (1 day) | 240 kcal pancake of wheat flour, egg and milk + 10%, 20% or 30% flour of Tm, Cr, or Lw. Total protein (g) of each pancake was: Tm, 8.2, 9.3, 10.2; Cr, 9.6, 12.0, 14.4; Lw, 9.6, 11, 12.6. | Placebo = 240 kcal pancake (7.6g protein from milk and egg). | 41 Females, 33 Males, healthy, 20 to 28 years, BMI 18.5 to 25 kg/m2 | Administration of insect or placebo products on separate days, after overnight fasting, with 1-day intervals (eight samples in total). Pancakes are eaten within 5 min. | Subjective feelings of hunger and satiety. Self-filled subjective rating questionnaires (VAS) before and after ingestion at 30-minute intervals for 180 min. Pearson correlation coefficients determined for satiety levels and physico-chemical parameters of products. | ↔ Hunger for 10%, 20%, 30% Tm and Cr, 10% Lw, placebo between men and women. ↑ Hunger for 20% and 30% Lw in women compared to men. ↓ AUC of Hunger for 20% and 30% Cr, and 30% Lw. ↔ Satiety for 10% and 30% Tm,10, 20 and 20% Cr, 10% Lw, and placebo between men and women. ↑ Satiety for 20% Tm, 20% and 30% Lw in women compared to men. ↑ AUC of satiety for 30% Lw, 20% and 30% Cr. Physico-chemical characteristics related to induction of satiety were: protein (positive relation) > carbohydrates, water (negative relation) > dietary fiber (positive relation). |
| Microbiota | |||||||
| Stull, et al., 2018 (USA) [24] | Crossover, Chronic (14 days) | Breakfast meal with Cr powder: muffins (15g Cr) and shake (10 g Cr), 21.7 g protein; 569.3 kcal. | Breakfast meal: pumpkin muffins and a chocolate shake (9.3 g protein, 495.3 kcal). | Eleven Females, nine Males, healthy, mean age 26.4 ± 6.3; mean BMI 23.4 ± 2.5 | Administration of insect or placebo products after overnight fasting in separate weeks, with 14-day washout intervals. | Plasma glucose, Na, K, CO2, Cl, Ca, urea nitrogen, creatine, ALP, ALT, AST, bilirubin, albumin, total protein; Systemic inflammation: plasma cytokines and chemokines; Mucosal immunity: fecal sIgA; Microbial metabolism: fecal SCFAs and bile acids; Microbiota; GI function: feelings related to digestive health. Collection of blood and stool after overnight fast, before intervention, after insect and control interventions. | Blood chemistry: ↔ Plasma Na, K, Cl, CO2, Ca, glucose, urea nitrogen, creatine, ALP, ALT, AST, bilirubin, albumin, total protein Inflammation: ↓ TNF-α for insect, ↔ for all other cytokines and chemokines Mucosal immunity: ↔ sIgA Microbial metabolism: ↓ SCFA acetate and propionate for insect; ↔ Bile acids Microbiota: ↓ Probiotic Lactobacillus spp. and Leuconostoc; ↑ Probiotic Bifidobacterium animalis, for insect ↔ GI function. |
| Disease treatment | |||||||
| Hu, et al., 2020 (China) [27] | Parallel, chronic (3 months) | Routine medication (β2 receptor agonist and inhaled glucocorticoids) + 30 g compound Caoshi silkworm Granules + 5 g Astragalus in granules. | Routine medication | Test: twelve Males, eight Females Control: eight Males, twelve Females; COPD, mean age 64.9 ± 8.2; mean BMI 21.3 ± 4.0 | Daily ingestion of medication or medication + granules. | Respiratory symptoms, activity and impact (SGRQ), lung function (FEV1, FVC, FEV1/FVC). Data collected before intervention and at the end of month 3. | ↓ SGRQ scores on symptoms, activity, and impact for insect. ↔ Pulmonary function 3 months after the intervention. |
| Author, Year, (Country) | Type | Test/Case | Control | Participants/Case | Intervention/Event | Outcomes Data Collection | Results |
|---|---|---|---|---|---|---|---|
| Cross-reactivity studies with patients’ sera | |||||||
| Lamberti et al., 2021 (Italy) [37] | Specific IgE to allergen extracts | Thermal processing of buffalo worm (Alphitobius diaperinus), mealworm larvae (Tenebrio molitor), cricket (Gryllodes sigillatus), grasshopper (Locusta migratoria), and silkworm larvae (Bombyx mori) proteins extracts. | Protein extracts from the mentioned raw insects. | Patients allergic to: house dust mites (HDM) (n = 28); shrimp (n = 8); mealworm (n = 2); Control patients: not allergic to either shrimps or HDM (n = 3); all adults. | Patients’ sera exposure to the proteins extracted from the insects, with or without thermal processing. | Insects’ protein profiles after each thermal treatment; screening in patients’ sera for the immunorecognition of the extracted insects’ proteins. | 71% of HDM and 87% of shrimp allergic patients recognizing at least one insect protein extract; tropomyosin and larval cuticle protein (LCP) as the most cross-reactive proteins; partial reduction of cross-allergenicity by thermal processing. |
| Kamemura et al., 2019 (Japan) [36] | Specific IgE to allergen extracts | Gryllus bimaculatus (cricket) allergen extracts. | Patients without shrimp allergy (n = 6). | Patients allergic to shrimp (n = 9). | Patients’ sera exposure to Gryllus and shrimp allergens extracts. | Identification of allergenic proteins in Gryllus and shrimp; estimation of allergen-specific IgE levels for shrimp- and Gryllus in patients sera. | Strong correlation between shrimp- and Gryllus-specific IgE responses; tropomyosin as the major allergen in shrimp and Gryllus. |
| Verhoeckx et al., 2014 (Netherlands) [35] | Specific IgE to allergen extracts; Basophil activation test | Yellow mealworm (Tenebrio molitor) protein extracts. | Patients allergic to grass pollen, peanuts, fish, or eggs and/or milk; also not allergic to crustaceans or HDM (n = 15). | Patients allergic to crustaceans and HDM (n = 7). | Patients’ sera exposure to different yellow mealworm protein fractions. | Identification of cross-reactive proteins of the yellow mealworm; indirect basophil activation. | IgE from HDM- and crustacean allergic patients cross-reaction with yellow mealworm proteins; induction of basophil activation; tropomyosin and arginine kinase as major cross-reactive proteins; moderate stability of cross-allergens after static pepsin digestion model test. |
| Epidemiological study | |||||||
| Ndlovu et al., 2021 (Zimbabwe) [34] | RCT Parallel, acute (1 day) | In-house preparation of mopane worm (Imbrasia belina) inhalant allergen extracts and other 10 inhalant allergen extracts locally relevant. | No control. | Patients ≥ 10 years old (total of 29 households) from a mopane worm harvesting rural community (n = 17, 13 females and 4 males). | Exposure by skin prick to the allergens. | Allergen sensitization patterns assessed by skin prick test, lung function (spirometry), and fractional exhaled nitric oxide levels (allergic airway inflammation). | Prevalence of sensitization to Imbrasia belina of 50%; respiratory health symptoms amongst participants sensitized to mopane worm; prevalence from 22 to 72% for other allergens including cockroach, mosquito, and HDM. |
| Case report | |||||||
| Beaumont et al., 2019 (France) [33] | Case study | Severe food anaphylaxis induced by mealworm (Tenebrio molitor). | Not applicable. | Patients allergic to HDM but not to crustaceans; 31 years old. | Consumption of 1 cooked larvae, probably fried. | Prick-tests; serum proteomic analysis (identification of T. molitor proteins to which he was sensitized). | Sensitization to hexamerin, tropomyosin epitopes, α-amylase (identified as an allergen in mealworms and with structural homology with HDM), and to larva cuticle proteins A1A and A2B (known T. molitor allergens). |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cunha, N.; Andrade, V.; Ruivo, P.; Pinto, P. Effects of Insect Consumption on Human Health: A Systematic Review of Human Studies. Nutrients 2023, 15, 3076. https://doi.org/10.3390/nu15143076
Cunha N, Andrade V, Ruivo P, Pinto P. Effects of Insect Consumption on Human Health: A Systematic Review of Human Studies. Nutrients. 2023; 15(14):3076. https://doi.org/10.3390/nu15143076
Chicago/Turabian StyleCunha, Nair, Vanda Andrade, Paula Ruivo, and Paula Pinto. 2023. "Effects of Insect Consumption on Human Health: A Systematic Review of Human Studies" Nutrients 15, no. 14: 3076. https://doi.org/10.3390/nu15143076
APA StyleCunha, N., Andrade, V., Ruivo, P., & Pinto, P. (2023). Effects of Insect Consumption on Human Health: A Systematic Review of Human Studies. Nutrients, 15(14), 3076. https://doi.org/10.3390/nu15143076