
NAD+ Precursors and Intestinal Inflammation: Therapeutic Insights Involving Gut Microbiota
 ,
,  , , ,
, , ,  ,
,  and
and 
Abstract

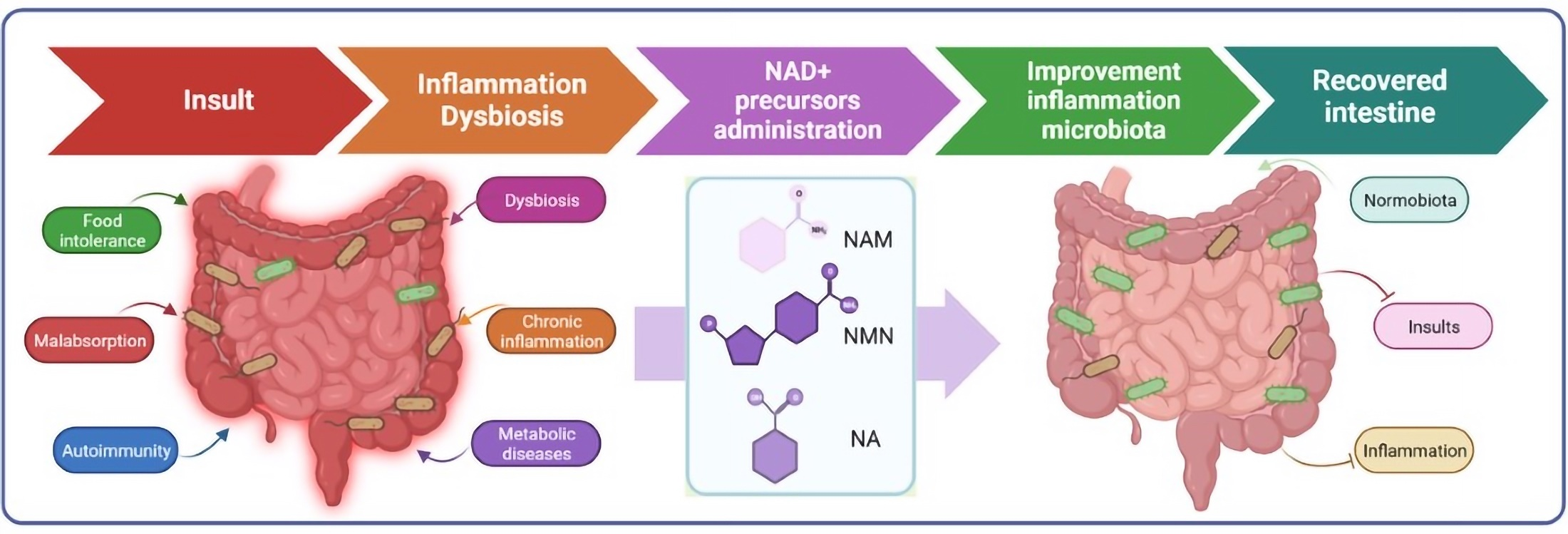
1. Background
2. Intestinal Inflammation and Dysbiosis in NAD+-Deficient States
2.1. Modifiable Factors
2.1.1. Pellagra, as an Example of Nutritional Deficiency
2.1.2. Chronic Alcoholism
2.1.3. Chemotherapy
2.2. Non-Modifiable Factors
2.2.1. Aging
2.2.2. Sex
2.2.3. Immunomodulatory Conditions
Inflammatory Bowel Disease (IBD)
Systemic Lupus Erythematosus (SLE)
Rheumatoid Arthritis (RA)
Multiple Sclerosis (MS)
2.2.4. Metabolic-Related Conditions
Obesity and Type 2 Diabetes
Polycystic Ovarian Syndrome (PCOS)
Cancer Cachexia (CC)
2.2.5. Linking Intestinal and Neurological Conditions: Autism Spectrum Disorder
3. NAD+-Increasing Therapies in the Treatment of Intestinal Inflammation

{kind=link}
{kind=link}
{kind=link}
| Form of B3 Supplement | Disease | Disease Model | Dose and Route Administration | Duration | Benefits in Gut Inflammation/Microbiota | References |
|---|---|---|---|---|---|---|
| NA | Colitis | Wistar rats (adult male 150–200 g) induced by intrarectal administration of iodoacetamide | 80 or 320 mg NA/kg was administered orally | Daily for 2 weeks |
| [127] |
| NA | Ulcerative colitis | Mutant mice with C57BL/6 genetic background 6- to 8-week-old male mice were administrated with DSS through drinking water (2%) for 6–9 days or pre-sensitized with 1% TNBS at day 1 and then challenged with 2.5% TNBS (100 µL) at day 8. | 600 mg NA/kg was administered via gavage | Daily for 6 (DP1) or 9 days (WT) |
| [128] |
| NA | Diarrheal disease | Weaned piglets (Duroc × Landrace × Yorkshire), 21 days old inoculated on the fourth day by oral administration of 4 × 109 cfu/mL enterotoxigenic E. coli (ETEC) K88 | 20 mL nicotinic acid solution with 40 mg NA | Daily for 3 days before E. coli inoculation |
| [129] |
| NA | Preventive | Weaned piglets (Duroc × Landrace × Yorkshire), 21 days old, 6.65 ± 0.02 kg body weight | Diet supplemented with 20.4 mg NA/kg | Daily for 14 days |
| [130] |
| NAM | Crohn’s disease | Human | 600 mg capsules | 2 doses daily for 4 weeks |
| [122] |
| NAM | Colitis | C57BL/6J mice (male, >18 weeks old) treated with 1.5% DSS in drinking water for 5 days | 30, 60 or 120 mg NAM/kg in drinking water or granules mixed with diet | Daily |
| [122] |
| NAM | Colitis | C57BL/6 WT mice (female, 6–8 weeks 20–28 g) induced by oral infection with C. rodentium or by DSS administration | Intraperitoneal injection of 250 mg/kg from day two before infection (preventive) or two days post infection (therapeutic) until the end of experiment | Daily for 12 days |
| [131] |
| NAM | Mild colitis | C57BL/6 mice Ace2-/y (male) with DSS | Trp1 diet and 0.4 g/L NAM in drinking water | Daily for 13 days |
| [132] |
| NMN | None | C57BL/6J mice (female, 12-week-old) | 0.1 mg/mL, 0.2 mg/mL, 0.4 mg/mL, and 0.6 mg/mL in drinking water | 15 weeks |
| [18] |
| NMN | Colitis | C57BL/6J mice (male, 8 weeks old) treated with drinking water containing 3% dextran sodium sulphate (DSS) | 1 mg NMN/g was administered via gavage | Daily for 3 weeks |
| [19] |
| β-NMN | Peritonitis | C57BL6 mice (male, 7–12 week-old) with cecal ligation and puncture (CLP) | Intraperitoneal injection at 185 mg β-NMN/kg, 4 days after CLP and for 3 days post thioglycolate treatment | Daily for 4 days |
| [133] |
| NR | None | C57BL/6N mice (male, 8–10-week) | 400 mg NR/kg by oral administration | Single |
| [134] |
| NR+ NRTBCl NRTOCl | None | Gallus gallus | Administration of 30 mg/mL NRCl, NRTBCl or NRTOCl via intra-amniotic | Day 17 of embryonic incubation |
| [135] |
3.1. Nicotinic Acid (NA)
3.2. Nicotinamide (NAM)
3.3. Nicotinamide Mononucleotide (NMN)
3.4. Nicotinamide Riboside (NR)
4. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bogan, K.L.; Brenner, C. Nicotinic Acid, Nicotinamide, and Nicotinamide Riboside: A Molecular Evaluation of NAD+ Precursor Vitamins in Human Nutrition. Annu. Rev. Nutr. 2008, 28, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Niacin and Nicotinic Acid. JAMA 1942, 118, 823. [CrossRef]
- Sydenstricker, V.P. The History of Pellagra, Its Recognition as a Disorder of Nutrition and Its Conquest. Am. J. Clin. Nutr. 1958, 6, 409–414. [Google Scholar] [CrossRef]
- Kleta, R.; Romeo, E.; Ristic, Z.; Ohura, T.; Stuart, C.; Arcos-Burgos, M.; Dave, M.H.; Wagner, C.A.; Camargo, S.R.M.; Inoue, S.; et al. Mutations in SLC6A19, Encoding B0AT1, Cause Hartnup Disorder. Nat. Genet. 2004, 36, 999–1002. [Google Scholar] [CrossRef]
- Seow, H.F.; Bröer, S.; Bröer, A.; Bailey, C.G.; Potter, S.J.; Cavanaugh, J.A.; Rasko, J.E.J. Hartnup Disorder Is Caused by Mutations in the Gene Encoding the Neutral Amino Acid Transporter SLC6A19. Nat. Genet. 2004, 36, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Elvehjem, C.A.; Madden, R.J.; Strong, F.M.; Woolley, D.W. The Isolation and Identification of the Anti-Black Tongue Factor*. Nutr. Rev. 1974, 32, 48–50. [Google Scholar] [CrossRef] [PubMed]
- Nozaki, J.I.; Dakeishi, M.; Ohura, T.; Inoue, K.; Manabe, M.; Wada, Y.; Koizumi, A. Homozygosity Mapping to Chromosome 5p15 of a Gene Responsible for Hartnup Disorder. Biochem. Biophys. Res. Commun. 2001, 284, 255–260. [Google Scholar] [CrossRef]
- Digby, J.E.; Martinez, F.; Jefferson, A.; Ruparelia, N.; Chai, J.; Wamil, M.; Greaves, D.R.; Choudhury, R.P. Anti-Inflammatory Effects of Nicotinic Acid in Human Monocytes Are Mediated by GPR109A Dependent Mechanisms. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 669–676. [Google Scholar] [CrossRef]
- Kang, Y.H.; Tucker, S.A.; Quevedo, S.F.; Inal, A.; Korzenik, J.R.; Haigis, M.C. Metabolic Analyses Reveal Dysregulated NAD+ Metabolism and Altered Mitochondrial State in Ulcerative Colitis. PLoS ONE 2022, 17, e0273080. [Google Scholar] [CrossRef]
- Shats, I.; Williams, J.G.; Liu, J.; Makarov, M.V.; Wu, X.; Lih, F.B.; Deterding, L.J.; Lim, C.; Xu, X.; Randall, T.A.; et al. Bacteria Boost Mammalian Host NAD Metabolism by Engaging the Deamidated Biosynthesis Pathway. Cell Metab. 2020, 31, 564–579.e7. [Google Scholar] [CrossRef]
- Verde, J.; Chen, C.; Brahmstedt, C.; Lukas, M.; Kolar, M.; Laser, D.; Salky, B.; Harrison, M. Nad+ Depletion In The Intestinal Epithelium Results In Mitochondrial Dysfunction And Influences The Pathogenesis Of Experimental Colitis. Gastroenterology 2023, 164, S8. [Google Scholar] [CrossRef]
- Hui, S.; Heng, L.; Shaodong, W.; Fangyu, W.; Zhenkai, W. Pellagra Affecting a Patient with Crohn’s Disease. An. Bras. Dermatol. 2017, 92, 879–881. [Google Scholar] [CrossRef]
- Nagahisa, T.; Yamaguchi, S.; Kosugi, S.; Homma, K.; Miyashita, K.; Irie, J.; Yoshino, J.; Itoh, H. Intestinal Epithelial NAD+ Biosynthesis Regulates GLP-1 Production and Postprandial Glucose Metabolism in Mice. Endocrinology 2022, 163, bqac023. [Google Scholar] [CrossRef]
- Nagahisa, T.; Kosugi, S.; Yamaguchi, S. Interactions between Intestinal Homeostasis and NAD+ Biology in Regulating Incretin Production and Postprandial Glucose Metabolism. Nutrients 2023, 15, 1494. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, Y.; Wang, Y.; Chao, Y.; Zhang, J.; Jia, Y.; Tie, J.; Hu, D. Regulation of SIRT1 and Its Roles in Inflammation. Front. Immunol. 2022, 13, 831168. [Google Scholar] [CrossRef]
- Rotllan, N.; Camacho, M.; Tondo, M.; Diarte-Añazco, E.M.G.; Canyelles, M.; Méndez-Lara, K.A.; Benitez, S.; Alonso, N.; Mauricio, D.; Escolà-Gil, J.C.; et al. Therapeutic Potential of Emerging NAD+-Increasing Strategies for Cardiovascular Diseases. Antioxidants 2021, 10, 1939. [Google Scholar] [CrossRef] [PubMed]
- Ellinger, P.; Shattock, F.M. Nicotinamide Deficiency after Oral Penicillin. Br. Med. J. 1946, 2, 611–613. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Jiang, A.; Wang, X.; Zhou, Y.; Tang, W.; Ren, C.; Qian, X.; Zhou, Z.; Gong, A. NMN Maintains Intestinal Homeostasis by Regulating the Gut Microbiota. Front. Nutr. 2021, 8, 714604. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Wang, X.; Wang, S.; Wu, Z.; Zhou, Z.; Shao, G.; Ren, C.; Kuang, M.; Zhou, Y.; Jiang, A.; et al. Treatment of Inflammatory Bowel Disease: Potential Effect of NMN on Intestinal Barrier and Gut Microbiota. Curr. Res. Food Sci. 2022, 5, 1403–1411. [Google Scholar] [CrossRef]
- Kang, K.; Sun, Y.; Pan, D.; Chang, B.; Sang, L.X. Nicotinamide Ameliorates Dextran Sulfate Sodium-Induced Chronic Colitis in Mice through Its Anti-Inflammatory Properties and Modulates the Gut Microbiota. J. Immunol. Res. 2021, 2021, 5084713. [Google Scholar] [CrossRef]
- Peluso, A.A.; Lundgaard, A.T.; Babaei, P.; Mousovich-Neto, F.; Rocha, A.L.; Damgaard, M.V.; Bak, E.G.; Gnanasekaran, T.; Dollerup, O.L.; Trammell, S.A.J.; et al. Oral Supplementation of Nicotinamide Riboside Alters Intestinal Microbial Composition in Rats and Mice, but Not Humans. NPJ Aging 2023, 9, 7. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, D.; Dawe, R.S.; Mponda, K. Pellagra a Review Exploring Causes and Mechanisms, Including Isoniazid-Induced Pellagra. Photodermatol. Photoimmunol. Photomed. 2021, 37, 99–104. [Google Scholar] [CrossRef]
- Halsted, C.H.; Sheir, S.; Sourial, N.; Patwardhan, V.N. Small Intestinal Structure and Absorption in Egypt: Influence of Parasitism and Pellagra. Am. J. Clin. Nutr. 1969, 22, 744–754. [Google Scholar] [CrossRef]
- Fukamizu, Y.; Uchida, Y.; Shigekawa, A.; Sato, T.; Kosaka, H.; Sakurai, T. Safety Evaluation of β-Nicotinamide Mononucleotide Oral Administration in Healthy Adult Men and Women. Sci. Rep. 2022, 12, 14442. [Google Scholar] [CrossRef]
- Groth, B.; Venkatakrishnan, P.; Lin, S.J. NAD+ Metabolism, Metabolic Stress, and Infection. Front. Mol. Biosci. 2021, 8, 686412. [Google Scholar] [CrossRef] [PubMed]
- Rohm, T.V.; Fuchs, R.; Müller, R.L.; Keller, L.; Baumann, Z.; Bosch, A.J.T.; Schneider, R.; Labes, D.; Langer, I.; Pilz, J.B.; et al. Obesity in Humans Is Characterized by Gut Inflammation as Shown by Pro-Inflammatory Intestinal Macrophage Accumulation. Front. Immunol. 2021, 12, 668654. [Google Scholar] [CrossRef] [PubMed]
- Tan, F.; Cao, Y.; Zheng, L.; Wang, T.; Zhao, S.; Chen, J.; Pang, C.; Xia, W.; Xia, Z.; Li, N.; et al. Diabetes Exacerbated Sepsis-Induced Intestinal Injury by Promoting M1 Macrophage Polarization via MiR-3061/Snail1 Signaling. Front. Immunol. 2022, 13, 922614. [Google Scholar] [CrossRef]
- Glassner, K.L.; Abraham, B.P.; Quigley, E.M.M. The Microbiome and Inflammatory Bowel Disease. J. Allergy Clin. Immunol. 2020, 145, 16–27. [Google Scholar] [CrossRef]
- Vrieze, A.; Holleman, F.; Zoetendal, E.G.; De Vos, W.M.; Hoekstra, J.B.L.; Nieuwdorp, M. The Environment within: How Gut Microbiota May Influence Metabolism and Body Composition. Diabetologia 2010, 53, 606–613. [Google Scholar] [CrossRef]
- Garrido, A.; Djouder, N. NAD+ Deficits in Age-Related Diseases and Cancer. Trends Cancer 2017, 3, 593–610. [Google Scholar] [CrossRef]
- Susai, N.; Kuroita, T.; Kuronuma, K.; Yoshioka, T. Analysis of the Gut Microbiome to Validate a Mouse Model of Pellagra. Biosci. Microbiota Food Health 2022, 41, 73–82. [Google Scholar] [CrossRef]
- Chellappa, K.; McReynolds, M.R.; Lu, W.; Zeng, X.; Makarov, M.; Hayat, F.; Mukherjee, S.; Bhat, Y.R.; Lingala, S.R.; Shima, R.T.; et al. NAD Precursors Cycle between Host Tissues and the Gut Microbiome. Cell Metab. 2022, 34, 1947–1959.e5. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.T.; Rodionov, D.A.; Peterson, S.N.; Osterman, A.L. B Vitamins and Their Role in Immune Regulation and Cancer. Nutrients 2020, 12, 3380. [Google Scholar] [CrossRef] [PubMed]
- Hill, L.J.; Williams, A.C. Meat Intake and the Dose of Vitamin B3—Nicotinamide: Cause of the Causes of Disease Transitions, Health Divides, and Health Futures? Int. J. Tryptophan Res. 2017, 10, 1178646917704662. [Google Scholar] [CrossRef]
- Parker, R.; Schmidt, M.S.; Cain, O.; Gunson, B.; Brenner, C. Nicotinamide Adenine Dinucleotide Metabolome Is Functionally Depressed in Patients Undergoing Liver Transplantation for Alcohol-Related Liver Disease. Hepatol. Commun. 2020, 4, 1183–1192. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A.B. Pellagra and Alcoholism: A Biochemical Perspective. Alcohol Alcohol. 2014, 49, 238–250. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhong, W. Targeting the Gut Barrier for the Treatment of Alcoholic Liver Disease. Liver Res. 2017, 1, 197–207. [Google Scholar] [CrossRef]
- French, S.W. Chronic Alcohol Binging Injures the Liver and Other Organs by Reducing NAD+ Levels Required for Sirtuin’s Deacetylase Activity. Exp. Mol. Pathol. 2016, 100, 303–306. [Google Scholar] [CrossRef]
- Bishehsari, F.; Magno, E.; Swanson, G.; Desai, V.; Voigt, R.M.; Forsyth, C.B.; Keshavarzian, A. Alcohol and Gut-Derived Inflammation. Alcohol Res. 2017, 38, 163–171. [Google Scholar] [PubMed]
- Engen, P.A.; Green, S.J.; Voigt, R.M.; Forsyth, C.B.; Keshavarzian, A. Alcohol Effects on the Composition of Intestinal Microbiota. Alcohol. Gastrointest. Microbiome Res. 2015, 37, 223–236. [Google Scholar] [PubMed]
- Banerjee, P.; Olmsted-Davis, E.A.; Deswal, A.; Nguyen, M.T.; Koutroumpakis, E.; Palaskas, N.L.; Lin, S.H.; Kotla, S.; Reyes-Gibby, C.; Yeung, S.J.; et al. Cancer treatment-induced NAD+ depletion in premature senescence and late cardiovascular complications. J. Cardiovasc. Aging 2022, 2, 28. [Google Scholar] [CrossRef]
- Stevens, H.P.; Ostlere, L.S.; Begent, R.H.J.; Dooley, J.S.; Rustin, M.H.A. Pellagra Secondary to 5-fluorouracil. Br. J. Dermatol. 1993, 128, 578–580. [Google Scholar] [CrossRef] [PubMed]
- Boyonoski, A.C.; Spronck, J.C.; Gallacher, L.M.; Jacobs, R.M.; Shah, G.M.; Poirier, G.G.; Kirkland, J.B. Pharmacological Intakes of Niacin Increase Bone Marrow Poly(ADP-Ribose) and the Latency of Ethylnitrosourea-Induced Carcinogenesis in Rats. J. Nutr. 2002, 132, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Wen, X.S.; Xian, C.J. Chemotherapy-Induced Intestinal Microbiota Dysbiosis Impairs Mucosal Homeostasis by Modulating Toll-like Receptor Signaling Pathways. Int. J. Mol. Sci. 2021, 22, 9474. [Google Scholar] [CrossRef] [PubMed]
- Sonis, S.T. The pathobiology of mucositis. Nat. Rev. Cancer 2004, 4, 277–284. [Google Scholar] [CrossRef]
- Mouchiroud, L.; Houtkooper, R.H.; Moullan, N.; Katsyuba, E.; Ryu, D.; Cantó, C.; Mottis, A.; Jo, Y.S.; Viswanathan, M.; Schoonjans, K.; et al. The NAD+/Sirtuin Pathway Modulates Longevity through Activation of Mitochondrial UPR and FOXO Signaling. Cell 2013, 154, 430–441. [Google Scholar] [CrossRef]
- Reiten, O.K.; Wilvang, M.A.; Mitchell, S.J.; Hu, Z.; Fang, E.F. Preclinical and Clinical Evidence of NAD+ Precursors in Health, Disease, and Ageing. Mech. Ageing Dev. 2021, 199, 111567. [Google Scholar] [CrossRef]
- Zhu, X.H.; Lu, M.; Lee, B.Y.; Ugurbil, K.; Chen, W. In Vivo NAD Assay Reveals the Intracellular NAD Contents and Redox State in Healthy Human Brain and Their Age Dependences. Proc. Natl. Acad. Sci. USA 2015, 112, 2876–2881. [Google Scholar] [CrossRef]
- Feuz, M.B.; Meyer-Ficca, M.L.; Meyer, R.G. Beyond Pellagra-Research Models and Strategies Addressing the Enduring Clinical Relevance of NAD Deficiency in Aging and Disease. Cells 2023, 12, 500. [Google Scholar] [CrossRef]
- Beneke, S.; Bürkle, A. Poly(ADP-Ribosyl)Ation in Mammalian Ageing. Nucleic Acids Res. 2007, 35, 7456–7465. [Google Scholar] [CrossRef]
- Niu, K.M.; Bao, T.; Gao, L.; Ru, M.; Li, Y.; Jiang, L.; Ye, C.; Wang, S.; Wu, X. The Impacts of Short-Term NMN Supplementation on Serum Metabolism, Fecal Microbiota, and Telomere Length in Pre-Aging Phase. Front. Nutr. 2021, 8, 756243. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, J.; Baur, J.A.; Imai, S. ichiro NAD+ Intermediates: The Biology and Therapeutic Potential of NMN and NR. Cell Metab. 2018, 27, 513–528. [Google Scholar] [CrossRef]
- Navarro, M.N.; Gómez de las Heras, M.M.; Mittelbrunn, M. Nicotinamide Adenine Dinucleotide Metabolism in the Immune Response, Autoimmunity and Inflammageing. Br. J. Pharmacol. 2022, 179, 1839–1856. [Google Scholar] [CrossRef] [PubMed]
- Covarrubias, A.J.; Perrone, R.; Grozio, A.; Verdin, E. NAD+ Metabolism and Its Roles in Cellular Processes during Ageing. Nat. Rev. Mol. Cell Biol. 2020, 22, 119–141. [Google Scholar] [CrossRef] [PubMed]
- Schultz, M.B.; Bochaton, T.; Bonkowski, M.; Li, J.; Lokitiyakul, D.; Colville, A.; Gomes, A.; Sinclair, D.; Boucher, N.A.; Steinhauser, K.E.; et al. NAD+ Depletion as A Cause of Inflammaging. Innov. Aging 2018, 2, 746. [Google Scholar] [CrossRef]
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic Inflammation in Ageing, Cardiovascular Disease, and Frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef]
- Zhu, X.; Shen, W.; Wang, Y.; Jaiswal, A.; Ju, Z.; Sheng, Q. Nicotinamide Adenine Dinucleotide Replenishment Rescues Colon Degeneration in Aged Mice. Signal Transduct. Target. Ther. 2017, 2, 17017. [Google Scholar] [CrossRef]
- Shintouo, C.M.; Mets, T.; Beckwee, D.; Bautmans, I.; Ghogomu, S.M.; Souopgui, J.; Leemans, L.; Meriki, H.D.; Njemini, R. Is Inflammageing Influenced by the Microbiota in the Aged Gut? A Systematic Review. Exp. Gerontol. 2020, 141, 111079. [Google Scholar] [CrossRef]
- Kim, S.; Jazwinski, S.M. The Gut Microbiota and Healthy Aging: A Mini-Review. Gerontology 2018, 64, 513–520. [Google Scholar] [CrossRef]
- Yurkovetskiy, L.; Burrows, M.; Khan, A.A.; Graham, L.; Volchkov, P.; Becker, L.; Antonopoulos, D.; Umesaki, Y.; Chervonsky, A.V. Gender Bias in Autoimmunity Is Influenced by Microbiota. Immunity 2013, 39, 400–412. [Google Scholar] [CrossRef]
- Angum, F.; Khan, T.; Kaler, J.; Siddiqui, L.; Hussain, A.; Angum, F.; Khan, T.; Kaler, J.; Siddiqui, L.; Hussain, A. The Prevalence of Autoimmune Disorders in Women: A Narrative Review. Cureus 2020, 12, e8094. [Google Scholar] [CrossRef]
- Brown, K.; Thomson, C.A.; Wacker, S.; Drikic, M.; Groves, R.; Fan, V.; Lewis, I.A.; McCoy, K.D. Microbiota Alters the Metabolome in an Age- and Sex- Dependent Manner in Mice. Nat. Commun. 2023, 14, 1348. [Google Scholar] [CrossRef] [PubMed]
- Schwarzmann, L.; Pliquett, R.U.; Simm, A.; Bartling, B. Sex-Related Differences in Human Plasma NAD+/NADH Levels Depend on Age. Biosci. Rep. 2021, 41, BSR20200340. [Google Scholar] [CrossRef] [PubMed]
- Escalante-Covarrubias, Q.; Mendoza-Viveros, L.; González-Suárez, M.; Sitten-Olea, R.; Velázquez-Villegas, L.A.; Becerril-Pérez, F.; Pacheco-Bernal, I.; Carreño-Vázquez, E.; Mass-Sánchez, P.; Bustamante-Zepeda, M.; et al. Time-of-Day Defines NAD+ Efficacy to Treat Diet-Induced Metabolic Disease by Synchronizing the Hepatic Clock in Mice. Nature Comm. 2023, 14, 1685. [Google Scholar] [CrossRef]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal Permeability--a New Target for Disease Prevention and Therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef]
- Hussein, I.H.; Dosh, L.; Al Qassab, M.; Jurjus, R.; El Masri, J.; Abi Nader, C.; Rappa, F.; Leone, A.; Jurjus, A. Highlights on two decades with microbiota and inflammatory bowel disease from etiology to therapy. Transpl. Immunol. 2023, 78, 101835. [Google Scholar] [CrossRef]
- Quaglio, A.E.V.; Grillo, T.G.; De Oliveira, E.C.S.; Di Stasi, L.C.; Sassaki, L.Y. Gut Microbiota, Inflammatory Bowel Disease and Colorectal Cancer. World J. Gastroenterol. 2022, 28, 4053–4060. [Google Scholar] [CrossRef]
- Nishida, A.; Inoue, R.; Inatomi, O.; Bamba, S.; Naito, Y.; Andoh, A. Gut Microbiota in the Pathogenesis of Inflammatory Bowel Disease. Clin. J. Gastroenterol. 2018, 11, 1–10. [Google Scholar] [CrossRef]
- Xue, X.; Miao, Y.; Wei, Z. Nicotinamide Adenine Dinucleotide Metabolism: Driving or Counterbalancing Inflammatory Bowel Disease? FEBS Lett. 2023, 597, 1179–1192. [Google Scholar] [CrossRef]
- Luo, X.M.; Edwards, M.R.; Mu, Q.; Yu, Y.; Vieson, M.D.; Reilly, C.M.; Ahmed, S.A.; Bankole, A.A. Gut Microbiota in Human Systemic Lupus Erythematosus and a Mouse Model of Lupus. Appl. Environ. Microbiol. 2018, 84, e02288-17. [Google Scholar] [CrossRef] [PubMed]
- Grammatikos, A.P.; Tsokos, G.C. Immunodeficiency and Autoimmunity: Lessons from Systemic Lupus Erythematosus. Trends Mol. Med. 2012, 18, 101–108. [Google Scholar] [CrossRef]
- Charoensappakit, A.; Sae-khow, K.; Leelahavanichkul, A. Gut Barrier Damage and Gut Translocation of Pathogen Molecules in Lupus, an Impact of Innate Immunity (Macrophages and Neutrophils) in Autoimmune Disease. Int. J. Mol. Sci. 2022, 23, 8223. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Singh, K.; Lin, A.; Meadows, A.M.; Wu, K.; Shing, V.; Bley, M.; Hassanzadeh, S.; Huffstutler, R.D.; Schmidt, M.S.; et al. Boosting NAD+ Blunts TLR4-Induced Type I IFN in Control and Systemic Lupus Erythematosus Monocytes. J. Clin. Investig. 2022, 132, e139828. [Google Scholar] [CrossRef] [PubMed]
- Buang, N.; Tapeng, L.; Gray, V.; Sardini, A.; Whilding, C.; Lightstone, L.; Cairns, T.D.; Pickering, M.C.; Behmoaras, J.; Ling, G.S.; et al. Type I Interferons Affect the Metabolic Fitness of CD8+ T Cells from Patients with Systemic Lupus Erythematosus. Nat. Commun. 2021, 12, 1980. [Google Scholar] [CrossRef]
- Guerreiro, C.S.; Calado, Â.; Sousa, J.; Fonseca, J.E. Diet, Microbiota, and Gut Permeability-The Unknown Triad in Rheumatoid Arthritis. Front. Med. 2018, 5, 349. [Google Scholar] [CrossRef]
- Cheng, M.; Zhao, Y.; Cui, Y.; Zhong, C.; Zha, Y.; Li, S.; Cao, G.; Li, M.; Zhang, L.; Ning, K.; et al. Stage-Specific Roles of Microbial Dysbiosis and Metabolic Disorders in Rheumatoid Arthritis. Ann. Rheum. Dis. 2022, 81, 1669–1677. [Google Scholar] [CrossRef]
- Zaiss, M.M.; Joyce Wu, H.J.; Mauro, D.; Schett, G.; Ciccia, F. The Gut–Joint Axis in Rheumatoid Arthritis. Nat. Rev. Rheumatol. 2021, 17, 224–237. [Google Scholar] [CrossRef]
- Perez-Sanchez, C.; Escudero-Contreras, A.; Cerdó, T.; Sánchez-Mendoza, L.M.; Llamas-Urbano, A.; Arias-de la Rosa, I.; Pérez-Rodriguez, M.; Muñoz-Barrera, L.; Abalos-Aguilera, M.D.C.; Barbarroja, N.; et al. Preclinical Characterization of Pharmacological NAD+ Boosting as a Promising Therapeutic Approach in Rheumatoid Arthritis. Arthritis Rheumatol. 2023. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F.; Russell, A.L. Niacinamide Therapy for Osteoarthritis—Does It Inhibit Nitric Oxide Synthase Induction by Interleukin-1 in Chondrocytes? Med. Hypotheses 1999, 53, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, W. THE USE OF VITAMIN THERAPY TO REVERSE CERTAIN CONCOMITANTS OF AGING†. J. Am. Geriatr. Soc. 1955, 3, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Ghezzi, L.; Cantoni, C.; Pinget, G.V.; Zhou, Y.; Piccio, L. Targeting the Gut to Treat Multiple Sclerosis. J. Clin. Investig. 2021, 131, e143774. [Google Scholar] [CrossRef]
- Penberthy, W.; Tsunoda, I. The Importance of NAD in Multiple Sclerosis. Curr. Pharm. Des. 2009, 15, 64–99. [Google Scholar] [CrossRef]
- Khan, S.; Luck, H.; Winer, S.; Winer, D.A. Emerging Concepts in Intestinal Immune Control of Obesity-Related Metabolic Disease. Nat. Commun. 2021, 12, 2598. [Google Scholar] [CrossRef]
- Luck, H.; Khan, S.; Kim, J.H.; Copeland, J.K.; Revelo, X.S.; Tsai, S.; Chakraborty, M.; Cheng, K.; Tao Chan, Y.; Nøhr, M.K.; et al. Gut-Associated IgA+ Immune Cells Regulate Obesity-Related Insulin Resistance. Nat. Commun. 2019, 10, 3650. [Google Scholar] [CrossRef]
- Tilg, H.; Zmora, N.; Adolph, T.E.; Elinav, E. The Intestinal Microbiota Fuelling Metabolic Inflammation. Nat. Rev. Immunol. 2019, 20, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Scheithauer, T.P.M.; Rampanelli, E.; Nieuwdorp, M.; Vallance, B.A.; Verchere, C.B.; Van Raalte, D.H.; Herrema, H. Gut Microbiota as a Trigger for Metabolic Inflammation in Obesity and Type 2 Diabetes. Front. Immunol. 2020, 11, 571731. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Jiang, Y.; Wang, M.; Melaku, M.; Liu, L.; Zhao, Y.; Everaert, N.; Yi, B.; Zhang, H. Intestinal Dysbiosis in Nonalcoholic Fatty Liver Disease (NAFLD): Focusing on the Gut-Liver Axis. Crit. Rev. Food Sci. Nutr. 2023, 63, 1689–1706. [Google Scholar] [CrossRef] [PubMed]
- Piras, C.; Mussap, M.; Noto, A.; De Giacomo, A.; Cristofori, F.; Spada, M.; Fanos, V.; Atzori, L.; Francavilla, R. Alteration of the Intestinal Permeability Are Reflected by Changes in the Urine Metabolome of Young Autistic Children: Preliminary Results. Metabolites 2022, 12, 104. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Cacicedo, J.M.; Ido, Y. Impaired Nicotinamide Adenine Dinucleotide (NAD+) Metabolism in Diabetes and Diabetic Tissues: Implications for Nicotinamide-related Compound Treatment. J. Diabetes Investig. 2020, 11, 1403–1419. [Google Scholar] [CrossRef]
- Gariani, K.; Menzies, K.J.; Ryu, D.; Wegner, C.J.; Wang, X.; Ropelle, E.R.; Moullan, N.; Zhang, H.; Perino, A.; Lemos, V.; et al. Eliciting the Mitochondrial Unfolded Protein Response by Nicotinamide Adenine Dinucleotide Repletion Reverses Fatty Liver Disease in Mice. Hepatology 2016, 63, 1190–1204. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, J.; Mills, K.F.; Yoon, M.J.; Imai, S.I. Nicotinamide Mononucleotide, a Key NAD+ Intermediate, Treats the Pathophysiology of Diet- and Age-Induced Diabetes in Mice. Cell Metab. 2011, 14, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Gulshan, M.; Yaku, K.; Okabe, K.; Mahmood, A.; Sasaki, T.; Yamamoto, M.; Hikosaka, K.; Usui, I.; Kitamura, T.; Tobe, K.; et al. Overexpression of Nmnat3 Efficiently Increases NAD and NGD Levels and Ameliorates Age-associated Insulin Resistance. Aging Cell 2018, 17, e12798. [Google Scholar] [CrossRef] [PubMed]
- Jukarainen, S.; Heinonen, S.; Rämö, J.T.; Rinnankoski-Tuikka, R.; Rappou, E.; Tummers, M.; Muniandy, M.; Hakkarainen, A.; Lundbom, J.; Lundbom, N.; et al. Obesity Is Associated With Low NAD/SIRT Pathway Expression in Adipose Tissue of BMI-Discordant Monozygotic Twins. J. Clin. Endocrinol. Metab. 2016, 101, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An Obesity-Associated Gut Microbiome with Increased Capacity for Energy Harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Ciofi, C.; De Boer, M.; Bruford, M.W.; Hecht, M.K.; Wallace, B.; Macintyre, R.J.; Lenk, P.; Eidenmueller, B.; Staudter, H.; Wicker, R.; et al. Human Gut Microbes Associated with Obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of Gut Microbiota in Type 2 Diabetes Pathophysiology. EBioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef] [PubMed]
- Pezzino, S.; Sofia, M.; Faletra, G.; Mazzone, C.; Litrico, G.; La Greca, G.; Latteri, S. Gut–Liver Axis and Non-Alcoholic Fatty Liver Disease: A Vicious Circle of Dysfunctions Orchestrated by the Gut Microbiome. Biology 2022, 11, 1622. [Google Scholar] [CrossRef]
- Sedighi, M.; Razavi, S.; Navab-Moghadam, F.; Khamseh, M.E.; Alaei-Shahmiri, F.; Mehrtash, A.; Amirmozafari, N. Comparison of Gut Microbiota in Adult Patients with Type 2 Diabetes and Healthy Individuals. Microb. Pathog. 2017, 111, 362–369. [Google Scholar] [CrossRef]
- Ren, Z.; Xu, Y.; Li, T.; Sun, W.; Tang, Z.; Wang, Y.; Zhou, K.; Li, J.; Ding, Q.; Liang, K.; et al. NAD+ and Its Possible Role in Gut Microbiota: Insights on the Mechanisms by Which Gut Microbes Influence Host Metabolism. Anim. Nutr. 2022, 10, 360. [Google Scholar] [CrossRef] [PubMed]
- Albillos, A.; De Gottardi, A.; Rescigno, M. The Gut-Liver Axis in Liver Disease: Pathophysiological Basis for Therapy. J. Hepatol. 2020, 72, 558–577. [Google Scholar] [CrossRef]
- Jornayvaz, R.; Gariani, K.; Song, Q.; Zhang, X. The Role of Gut–Liver Axis in Gut Microbiome Dysbiosis Associated NAFLD and NAFLD-HCC. Biomedicines 2022, 10, 524. [Google Scholar] [CrossRef]
- Dall, M.; Hassing, A.S.; Treebak, J.T. NAD+ and NAFLD—Caution, Causality and Careful Optimism. J. Physiol. 2022, 600, 1135–1154. [Google Scholar] [CrossRef] [PubMed]
- Tremellen, K.; Pearce, K. Dysbiosis of Gut Microbiota (DOGMA)—A Novel Theory for the Development of Polycystic Ovarian Syndrome. Med. Hypotheses 2012, 79, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Aflatounian, A.; Paris, V.R.; Richani, D.; Edwards, M.C.; Cochran, B.J.; Ledger, W.L.; Gilchrist, R.B.; Bertoldo, M.J.; Wu, L.E.; Walters, K.A. Declining Muscle NAD+ in a Hyperandrogenism PCOS Mouse Model: Possible Role in Metabolic Dysregulation. Mol. Metab. 2022, 65, 101583. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Jiang, Y.; Xi, H.; Chen, L.; Feng, X. Exploration of the Relationship between Gut Microbiota and Polycystic Ovary Syndrome (PCOS): A Review. Geburtshilfe Frauenheilkd. 2020, 80, 161–171. [Google Scholar] [CrossRef]
- Rizk, M.G.; Thackray, V.G. Intersection of Polycystic Ovary Syndrome and the Gut Microbiome. J. Endocr. Soc. 2021, 5, bvaa177. [Google Scholar] [CrossRef]
- Park, J.M.; Han, Y.M.; Lee, H.J.; Park, Y.J.; Hahm, K.B. Nicotinamide Riboside Vitamin B3 Mitigated C26 Adenocarcinoma–Induced Cancer Cachexia. Front. Pharmacol. 2021, 12, 665493. [Google Scholar] [CrossRef]
- Herremans, K.M.; Riner, A.N.; Cameron, M.E.; Trevino, J.G. The Microbiota and Cancer Cachexia. Int. J. Mol. Sci. 2019, 20, 6267. [Google Scholar] [CrossRef]
- Okamura, A.; Ohmura, Y.; Islam, M.M.; Tagawa, M.; Horitsu, K.; Moriyama, Y.; Fujimura, S. Increased Hepatic Nicotinamide N-Methyltransferase Activity as a Marker of Cancer Cachexia in Mice Bearing Colon 26 Adenocarcinoma. Jpn. J. Cancer Res. 1998, 89, 649–656. [Google Scholar] [CrossRef]
- Samant, S.A.; Pillai, V.B.; Gupta, M.P. Cellular Mechanisms Promoting Cachexia and How They Are Opposed by Sirtuins. Can. J. Physiol. Pharmacol. 2019, 97, 235–245. [Google Scholar] [CrossRef]
- Costa, R.G.F.; Caro, P.L.; De Matos-Neto, E.M.; Lima, J.D.C.C.; Radloff, K.; Alves, M.J.; Camargo, R.G.; Pessoa, A.F.M.; Simoes, E.; Gama, P.; et al. Cancer Cachexia Induces Morphological and Inflammatory Changes in the Intestinal Mucosa. J. Cachexia Sarcopenia Muscle 2019, 10, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Beltrà, M.; Pöllänen, N.; Fornelli, C.; Tonttila, K.; Hsu, M.Y.; Zampieri, S.; Moletta, L.; Corrà, S.; Porporato, P.E.; Kivelä, R.; et al. NAD+ Repletion with Niacin Counteracts Cancer Cachexia. Nat. Commun. 2023, 14, 1849. [Google Scholar] [CrossRef]
- Gujar, A.D.; Le, S.; Mao, D.D.; Dadey, D.Y.A.; Turski, A.; Sasaki, Y.; Aum, D.; Luo, J.; Dahiya, S.; Yuan, L.; et al. An NAD+-Dependent Transcriptional Program Governs Self-Renewal and Radiation Resistance in Glioblastoma. Proc. Natl. Acad. Sci. USA 2016, 113, E8247–E8256. [Google Scholar] [CrossRef]
- Chalkiadaki, A.; Guarente, L. The Multifaceted Functions of Sirtuins in Cancer. Nat. Rev. Cancer 2015, 15, 608–624. [Google Scholar] [CrossRef] [PubMed]
- Essa, M.M.; Subash, S.; Braidy, N.; Al-Adawi, S.; Lim, C.K.; Manivasagam, T.; Guillemin, G.J. Role of NAD+, Oxidative Stress, and Tryptophan Metabolism in Autism Spectrum Disorders. Int. J. Tryptophan Res. 2013, 6 (Suppl. 1), 15–28. [Google Scholar] [CrossRef] [PubMed]
- Brister, D.; Rose, S.; Delhey, L.; Tippett, M.; Jin, Y.; Gu, H.; Frye, R.E. Metabolomic Signatures of Autism Spectrum Disorder. J. Pers. Med. 2022, 12, 1727. [Google Scholar] [CrossRef] [PubMed]
- Giulivi, C.; Zhang, Y.F.; Omanska-Klusek, A.; Ross-Inta, C.; Wong, S.; Hertz-Picciotto, I.; Tassone, F.; Pessah, I.N. Mitochondrial Dysfunction in Autism. JAMA 2010, 304, 2389–2396. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.B.; Audhya, T.; McDonough-Means, S.; Rubin, R.A.; Quig, D.; Geis, E.; Gehn, E.; Loresto, M.; Mitchell, J.; Atwood, S.; et al. Nutritional and Metabolic Status of Children with Autism vs. Neurotypical Children, and the Association with Autism Severity. Nutr. Metab. 2011, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Gerner, R.R.; Klepsch, V.; Macheiner, S.; Arnhard, K.; Adolph, T.E.; Grander, C.; Wieser, V.; Pfister, A.; Moser, P.; Hermann-Kleiter, N.; et al. NAD Metabolism Fuels Human and Mouse Intestinal Inflammation. Gut 2018, 67, 1813–1823. [Google Scholar] [CrossRef]
- Hong, S.M.; Lee, A.-Y.; Hwang, S.-M.; Ha, Y.-J.; Kim, M.-J.; Min, S.; Hwang, W.; Yoon, G.; Kwon, S.M.; Woo, H.G.; et al. NAMPT Mitigates Colitis Severity by Supporting Redox-Sensitive Activation of Phagocytosis in Inflammatory Macrophages. Redox Biol. 2022, 50, 102237. [Google Scholar] [CrossRef]
- Cheng, C.Y.; Gutierrez, N.M.; Marzuki, M.B.; Lu, X.; Foreman, T.W.; Paleja, B.; Lee, B.; Balachander, A.; Chen, J.; Tsenova, L.; et al. Host Sirtuin 1 Regulates Mycobacterial Immunopathogenesis and Represents a Therapeutic Target against Tuberculosis. Sci. Immunol. 2017, 2, eaaj1789. [Google Scholar] [CrossRef] [PubMed]
- Wätzig, G.; Seegert, D. A Composition Containing Nicotinic Acid and/or Nicotinamide and/or Tryptophan for Positively Influencing the Intestinal Microbiota. European Patent 3831379 A1, 9 June 2021. [Google Scholar]
- Magnúsdóttir, S.; Ravcheev, D.; De Crécy-Lagard, V.; Thiele, I. Systematic Genome Assessment of B-Vitamin Biosynthesis Suggests Cooperation among Gut Microbes. Front. Genet. 2015, 6, 148. [Google Scholar] [CrossRef]
- Jin, L.H.; Fang, Z.P.; Fan, M.J.; Huang, W. Dong Bile-Ology: From Bench to Bedside. J. Zhejiang Univ. Sci. B 2019, 20, 414–427. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, T.; Moschetta, A.; Lee, Y.K.; Peng, L.; Zhao, G.; Downes, M.; Yu, R.T.; Shelton, J.M.; Richardson, J.A.; Repa, J.J.; et al. Regulation of Antibacterial Defense in the Small Intestine by the Nuclear Bile Acid Receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 3920–3925. [Google Scholar] [CrossRef]
- Kurdi, P.; Kawanishi, K.; Mizutani, K.; Yokota, A. Mechanism of Growth Inhibition by Free Bile Acids in Lactobacilli and Bifidobacteria. J. Bacteriol. 2006, 188, 1979–1986. [Google Scholar] [CrossRef]
- Salem, H.A.; Wadie, W. Effect of Niacin on Inflammation and Angiogenesis in a Murine Model of Ulcerative Colitis. Sci. Rep. 2017, 7, 7139. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kong, D.; Wang, Q.; Wu, W.; Tang, Y.; Bai, T.; Guo, L.; Wei, L.; Zhang, Q.; Yu, Y.; et al. Niacin Ameliorates Ulcerative Colitis via Prostaglandin D2-Mediated D Prostanoid Receptor 1 Activation. EMBO Mol. Med. 2017, 9, 571–588. [Google Scholar] [CrossRef]
- Zhen, R.; Feng, J.; He, D.; Chen, Y.; Chen, T.; Cai, W.; Xiong, Y.; Qiu, Y.; Jiang, Z.; Wang, L.; et al. Effects of Niacin on Resistance to Enterotoxigenic Escherichia Coli Infection in Weaned Piglets. Front. Nutr. 2022, 9, 865311. [Google Scholar] [CrossRef]
- Liu, S.; Zhu, X.; Qiu, Y.; Wang, L.; Shang, X.; Gao, K.; Yang, X.; Jiang, Z. Effect of Niacin on Growth Performance, Intestinal Morphology, Mucosal Immunity and Microbiota Composition in Weaned Piglets. Animals 2021, 11, 2186. [Google Scholar] [CrossRef]
- Bettenworth, D.; Nowacki, T.M.; Ross, M.; Kyme, P.; Schwammbach, D.; Kerstiens, L.; Thoennissen, G.B.; Bokemeyer, C.; Hengst, K.; Berdel, W.E.; et al. Nicotinamide Treatment Ameliorates the Course of Experimental Colitis Mediated by Enhanced Neutrophil-Specific Antibacterial Clearance. Mol. Nutr. Food Res. 2014, 58, 1474–1490. [Google Scholar] [CrossRef]
- Hashimoto, T.; Perlot, T.; Rehman, A.; Trichereau, J.; Ishiguro, H.; Paolino, M.; Sigl, V.; Hanada, T.; Hanada, R.; Lipinski, S.; et al. ACE2 Links Amino Acid Malnutrition to Microbial Ecology and Intestinal Inflammation. Nature 2012, 487, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Cros, C.; Margier, M.; Cannelle, H.; Charmetant, J.; Hulo, N.; Laganier, L.; Grozio, A.; Canault, M. Nicotinamide Mononucleotide Administration Triggers Macrophages Reprogramming and Alleviates Inflammation During Sepsis Induced by Experimental Peritonitis. Front. Mol. Biosci. 2022, 9, 895028. [Google Scholar] [CrossRef] [PubMed]
- Yaku, K.; Palikhe, S.; Izumi, H.; Yoshida, T.; Hikosaka, K.; Hayat, F.; Karim, M.; Iqbal, T.; Nitta, Y.; Sato, A.; et al. BST1 Regulates Nicotinamide Riboside Metabolism via Its Glycohydrolase and Base-Exchange Activities. Nat. Commun. 2021, 12, 6767. [Google Scholar] [CrossRef]
- Kolba, N.; Zarei, A.; Cheng, J.; Agarwal, N.; Dadmohammadi, Y.; Khazdooz, L.; Abbaspourrad, A.; Tako, E. Alterations in Intestinal Brush Border Membrane Functionality and Bacterial Populations Following Intra-Amniotic Administration (Gallus Gallus) of Nicotinamide Riboside and Its Derivatives. Nutrients 2022, 14, 3130. [Google Scholar] [CrossRef] [PubMed]
- Altschul, R.; Hoffer, A. Effects of Salts of Nicotinic Acid on Serum Cholesterol. Br. Med. J. 1958, 2, 713–714. [Google Scholar] [CrossRef]
- Lukasova, M.; Malaval, C.; Gille, A.; Kero, J.; Offermanns, S. Nicotinic Acid Inhibits Progression of Atherosclerosis in Mice through Its Receptor GPR109A Expressed by Immune Cells. J. Clin. Investig. 2011, 121, 1163–1173. [Google Scholar] [CrossRef]
- Santoru, M.L.; Piras, C.; Murgia, A.; Palmas, V.; Camboni, T.; Liggi, S.; Ibba, I.; Lai, M.A.; Orrù, S.; Blois, S.; et al. Cross Sectional Evaluation of the Gut-Microbiome Metabolome Axis in an Italian Cohort of IBD Patients. Sci. Rep. 2017, 7, 9523. [Google Scholar] [CrossRef]
- Santoru, M.L.; Piras, C.; Murgia, F.; Spada, M.; Tronci, L.; Leoni, V.P.; Serreli, G.; Deiana, M.; Atzori, L. Modulatory Effect of Nicotinic Acid on the Metabolism of Caco-2 Cells Exposed to IL-1β and LPS. Metabolites 2020, 10, 204. [Google Scholar] [CrossRef] [PubMed]
- Lopez, L.R.; Barlogio, C.J.; Broberg, C.A.; Wang, J.; Arthur, J.C. A NadA Mutation Confers Nicotinic Acid Auxotrophy in Pro-Carcinogenic Intestinal Escherichia Coli NC101. Front. Microbiol. 2021, 12, 670005. [Google Scholar] [CrossRef]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The Biological Stress of Early Weaned Piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef]
- Pan, L.; Zhao, P.F.; Ma, X.K.; Shang, Q.H.; Xu, Y.T.; Long, S.F.; Wu, Y.; Yuan, F.M.; Piao, X.S. Probiotic Supplementation Protects Weaned Pigs against Enterotoxigenic Escherichia Coli K88 Challenge and Improves Performance Similar to Antibiotics. J. Anim. Sci. 2017, 95, 2627–2639. [Google Scholar] [CrossRef] [PubMed]
- Boudry, G.; Péron, V.; Le Huërou-Luron, I.; Lallès, J.P.; Sève, B. Weaning Induces Both Transient and Long-Lasting Modifications of Absorptive, Secretory, and Barrier Properties of Piglet Intestine. J. Nutr. 2004, 134, 2256–2262. [Google Scholar] [CrossRef] [PubMed]
- Spreeuwenberg, M.A.M.; Verdonk, J.M.A.J.; Gaskins, H.R.; Verstegen, M.W.A. Small Intestine Epithelial Barrier Function Is Compromised in Pigs with Low Feed Intake at Weaning. J. Nutr. 2001, 131, 1520–1527. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yi, Z.; Wang, Q.; Yin, L.; Li, J.; Xie, J.; Yang, H.; Yin, Y. Effect of Different Levels of Niacin on Serum Biochemical Parameters, Antioxidant Status, Cytokine Levels, Inflammatory Gene Expression and Colonic Microbial Composition in Weaned Piglets. Animals 2022, 12, 3018. [Google Scholar] [CrossRef]
- Hiippala, K.; Jouhten, H.; Ronkainen, A.; Hartikainen, A.; Kainulainen, V.; Jalanka, J.; Satokari, R. The Potential of Gut Commensals in Reinforcing Intestinal Barrier Function and Alleviating Inflammation. Nutrients 2018, 10, 988. [Google Scholar] [CrossRef] [PubMed]
- Ru, M.; Wang, W.; Zhai, Z.; Wang, R.; Li, Y.; Liang, J.; Kothari, D.; Niu, K.; Wu, X. Nicotinamide Mononucleotide Supplementation Protects the Intestinal Function in Aging Mice and D-Galactose Induced Senescent Cells. Food Funct. 2022, 13, 7507–7519. [Google Scholar] [CrossRef]
- Zhao, X.; Ji, K.; Zhang, M.; Huang, H.; Wang, F.; Liu, Y.; Liu, Q. NMN Alleviates Radiation-Induced Intestinal Fibrosis by Modulating Gut Microbiota. Int. J. Radiat. Biol. 2023, 99, 823–834. [Google Scholar] [CrossRef]
- Fang, D.; Xu, T.; Sun, J.; Shi, J.; Li, F.; Yin, Y.; Wang, Z.; Liu, Y. Nicotinamide Mononucleotide Ameliorates Sleep Deprivation-Induced Gut Microbiota Dysbiosis and Restores Colonization Resistance against Intestinal Infections. Adv. Sci. 2023, 10, e2207170. [Google Scholar] [CrossRef]
- Rodionov, D.A.; Li, X.; Rodionova, I.A.; Yang, C.; Sorci, L.; Dervyn, E.; Martynowski, D.; Zhang, H.; Gelfand, M.S.; Osterman, A.L. Transcriptional Regulation of NAD Metabolism in Bacteria: Genomic Reconstruction of NiaR (YrxA) Regulon. Nucleic Acids Res. 2008, 36, 2032–2046. [Google Scholar] [CrossRef]
- Engels, C.; Ruscheweyh, H.J.; Beerenwinkel, N.; Lacroix, C.; Schwab, C. The Common Gut Microbe Eubacterium Hallii Also Contributes to Intestinal Propionate Formation. Front. Microbiol. 2016, 7, 713. [Google Scholar] [CrossRef]


| Disease | Characterization in Humans or Disease Model | NAD Deficiency or Metabolic Alteration | References |
|---|---|---|---|
| Pellagra | Human | Primary pellagra is characterized by a cellular deficiency of NAD+ caused by insufficient intake of dietary precursors | [6,22] |
| Alcoholism | Human | Secondary pellagra as a result of malnutrition, which is also linked to NAD+ deficiency | [6] |
| Chemotherapy | Human | Chemotherapy can induce pellagra symptoms due to excess NAD+ consumption | [6,22] |
| Inflammatory bowel disease | Human | Increased cellular NAD consumption in IBD. | [23,67] |
| Autism spectrum disorder | Human | Irritable bowel syndrome, and gastroesophageal reflux are common comorbidities in ASD; increased gut permeability related to disruptions in the tryptophan-NA metabolic pathway | [88] |
| Obesity | Mouse model | Abrogation of NAMPT specifically in intestinal epithelial cells led to reduced local NAD+ levels and impaired intestinal physiology. | [13] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niño-Narvión, J.; Rojo-López, M.I.; Martinez-Santos, P.; Rossell, J.; Ruiz-Alcaraz, A.J.; Alonso, N.; Ramos-Molina, B.; Mauricio, D.; Julve, J. NAD+ Precursors and Intestinal Inflammation: Therapeutic Insights Involving Gut Microbiota. Nutrients 2023, 15, 2992. https://doi.org/10.3390/nu15132992
Niño-Narvión J, Rojo-López MI, Martinez-Santos P, Rossell J, Ruiz-Alcaraz AJ, Alonso N, Ramos-Molina B, Mauricio D, Julve J. NAD+ Precursors and Intestinal Inflammation: Therapeutic Insights Involving Gut Microbiota. Nutrients. 2023; 15(13):2992. https://doi.org/10.3390/nu15132992
Chicago/Turabian StyleNiño-Narvión, Julia, Marina Idalia Rojo-López, Patricia Martinez-Santos, Joana Rossell, Antonio J. Ruiz-Alcaraz, Núria Alonso, Bruno Ramos-Molina, Didac Mauricio, and Josep Julve. 2023. "NAD+ Precursors and Intestinal Inflammation: Therapeutic Insights Involving Gut Microbiota" Nutrients 15, no. 13: 2992. https://doi.org/10.3390/nu15132992
APA StyleNiño-Narvión, J., Rojo-López, M. I., Martinez-Santos, P., Rossell, J., Ruiz-Alcaraz, A. J., Alonso, N., Ramos-Molina, B., Mauricio, D., & Julve, J. (2023). NAD+ Precursors and Intestinal Inflammation: Therapeutic Insights Involving Gut Microbiota. Nutrients, 15(13), 2992. https://doi.org/10.3390/nu15132992