
Bioinformatics Analysis of the Molecular Networks Associated with the Amelioration of Aberrant Gene Expression by a Tyr–Trp Dipeptide in Brains Treated with the Amyloid-β Peptide
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Ethics Approval
2.2. DNA Microarray Analysis and Data Acquisition
2.3. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analyses
2.4. Gene Set Enrichment Analysis (GSEA)
2.5. Visualization of the Protein–Protein Interaction (PPI) Network and Module Detection
2.6. Ingenuity Pathways Analysis
2.7. Statistical Analyses
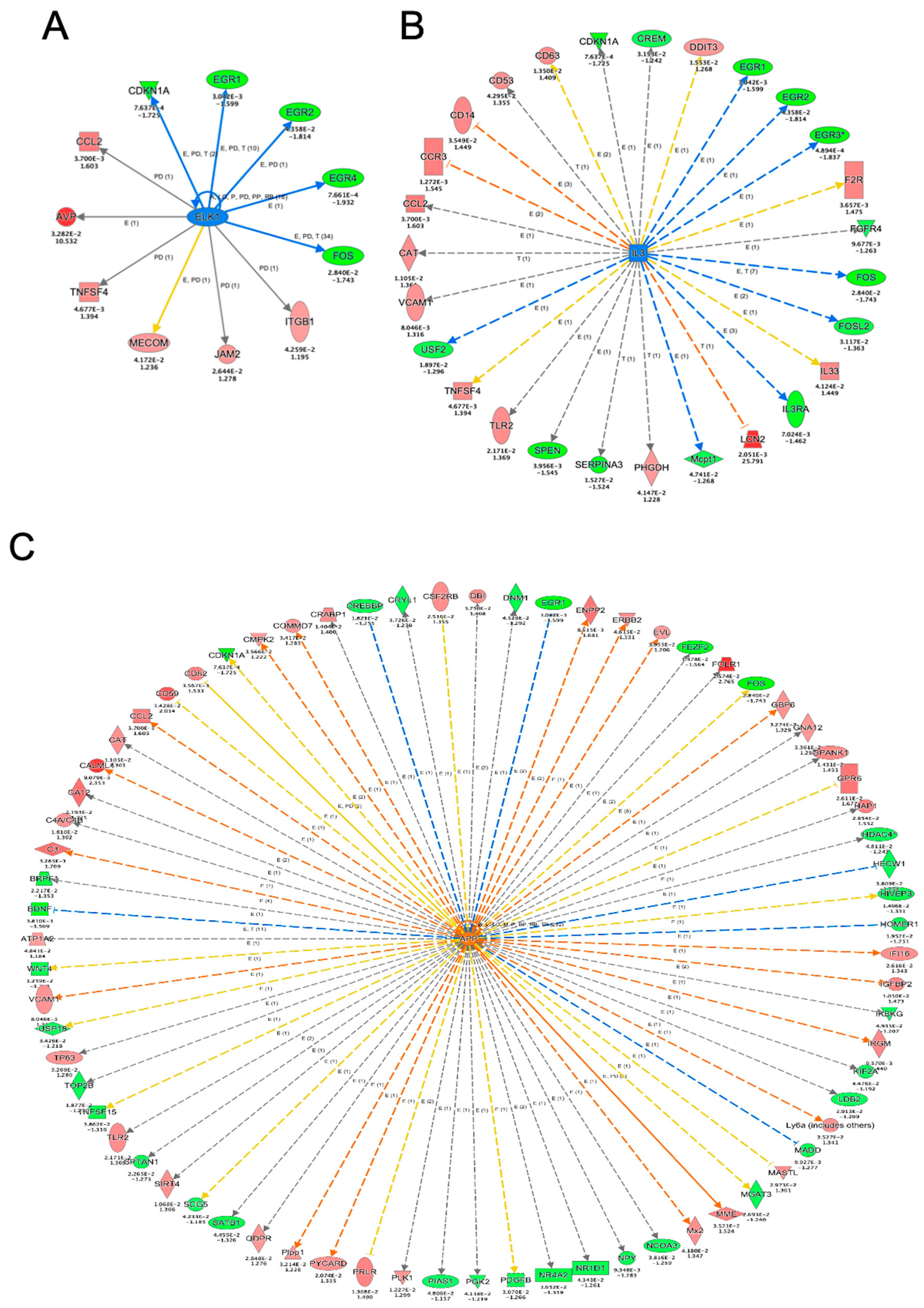
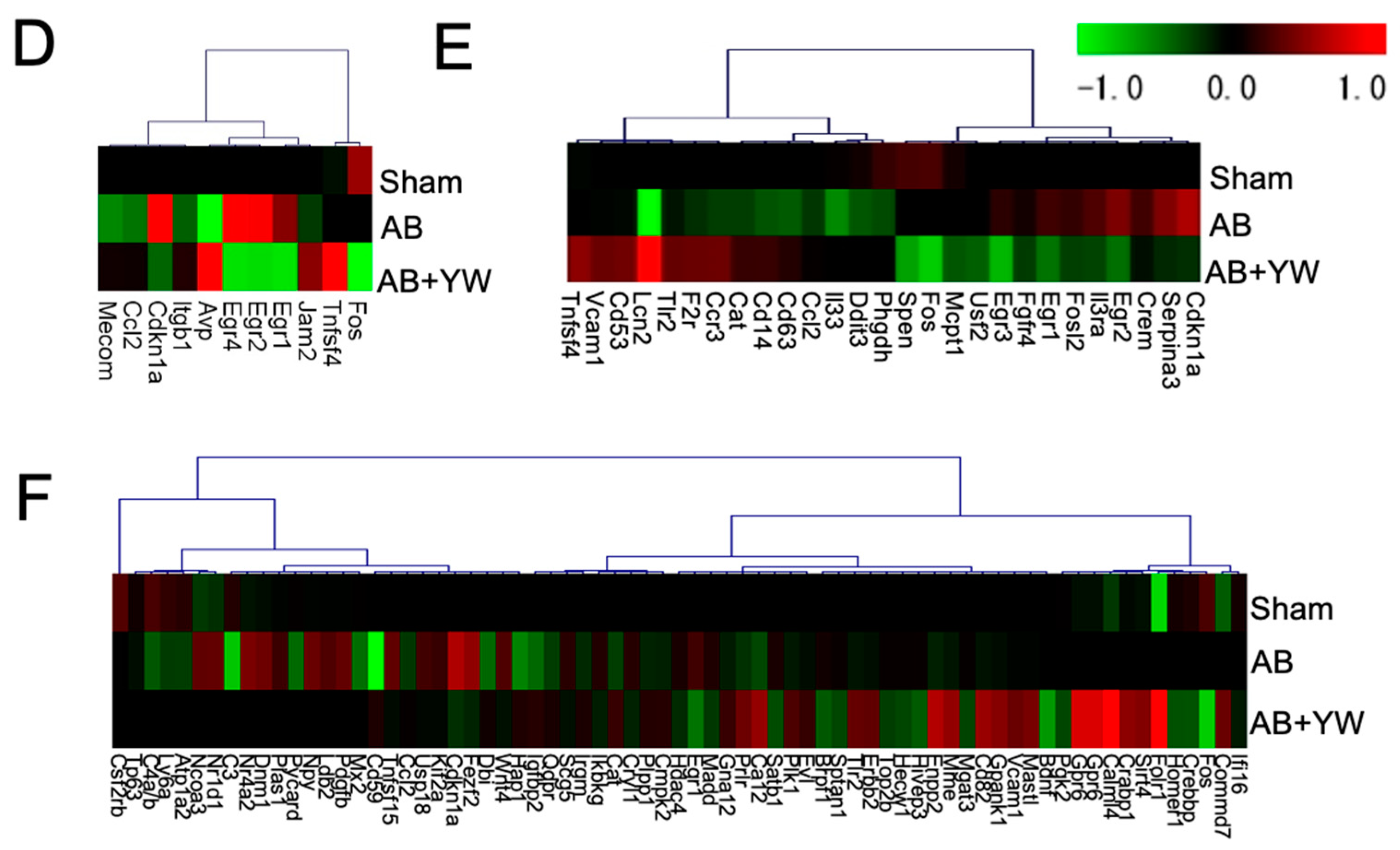
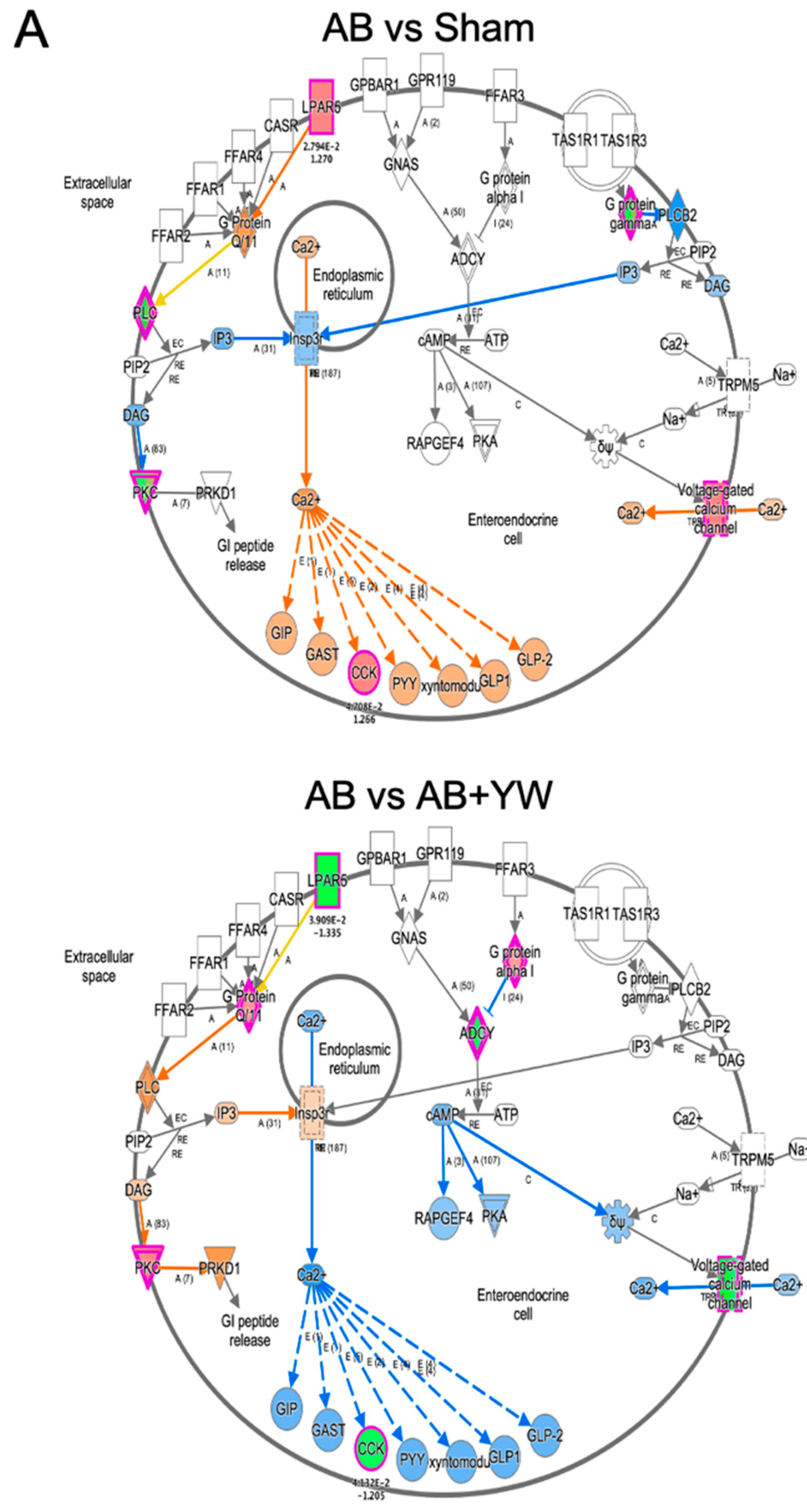
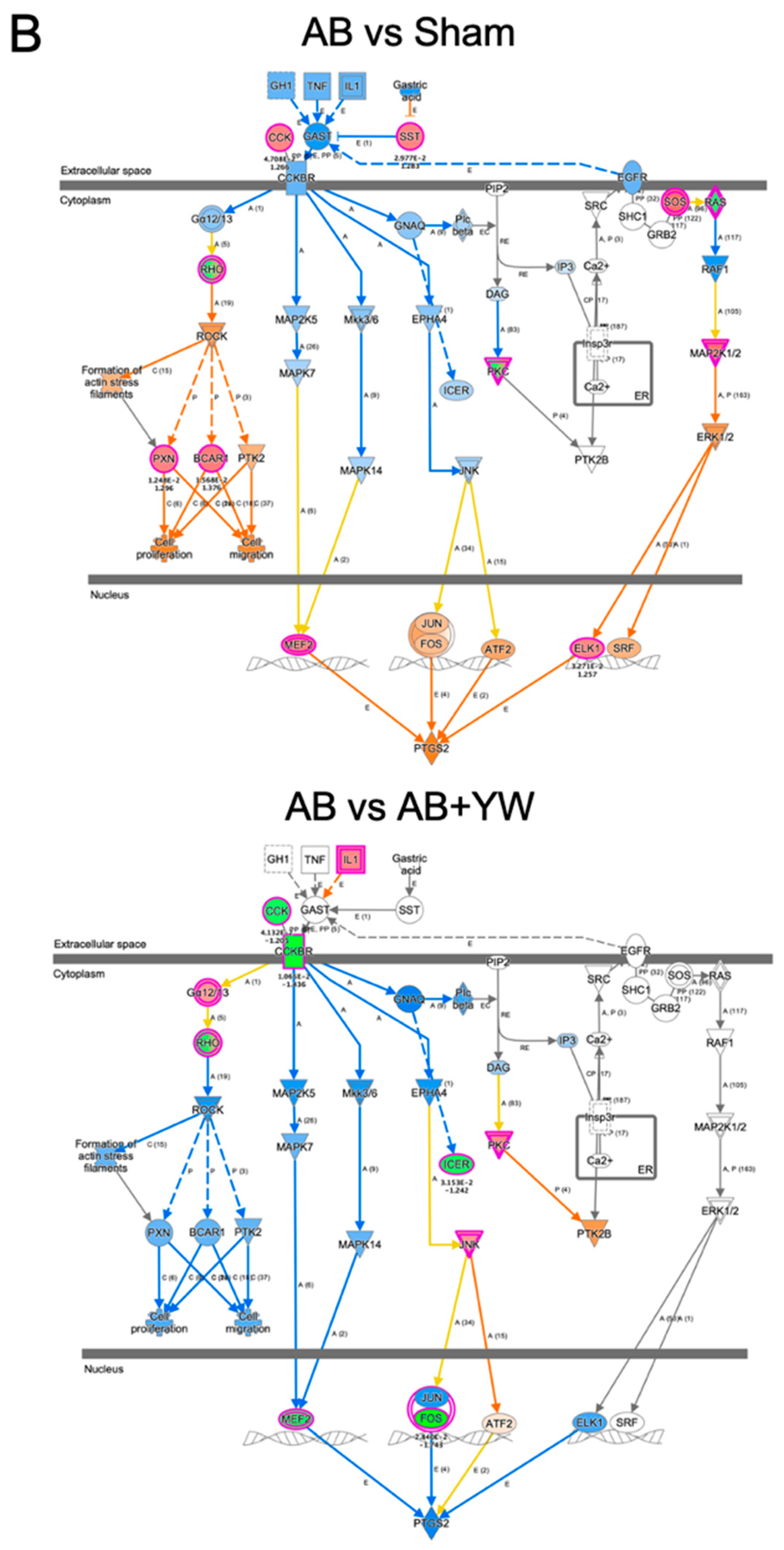
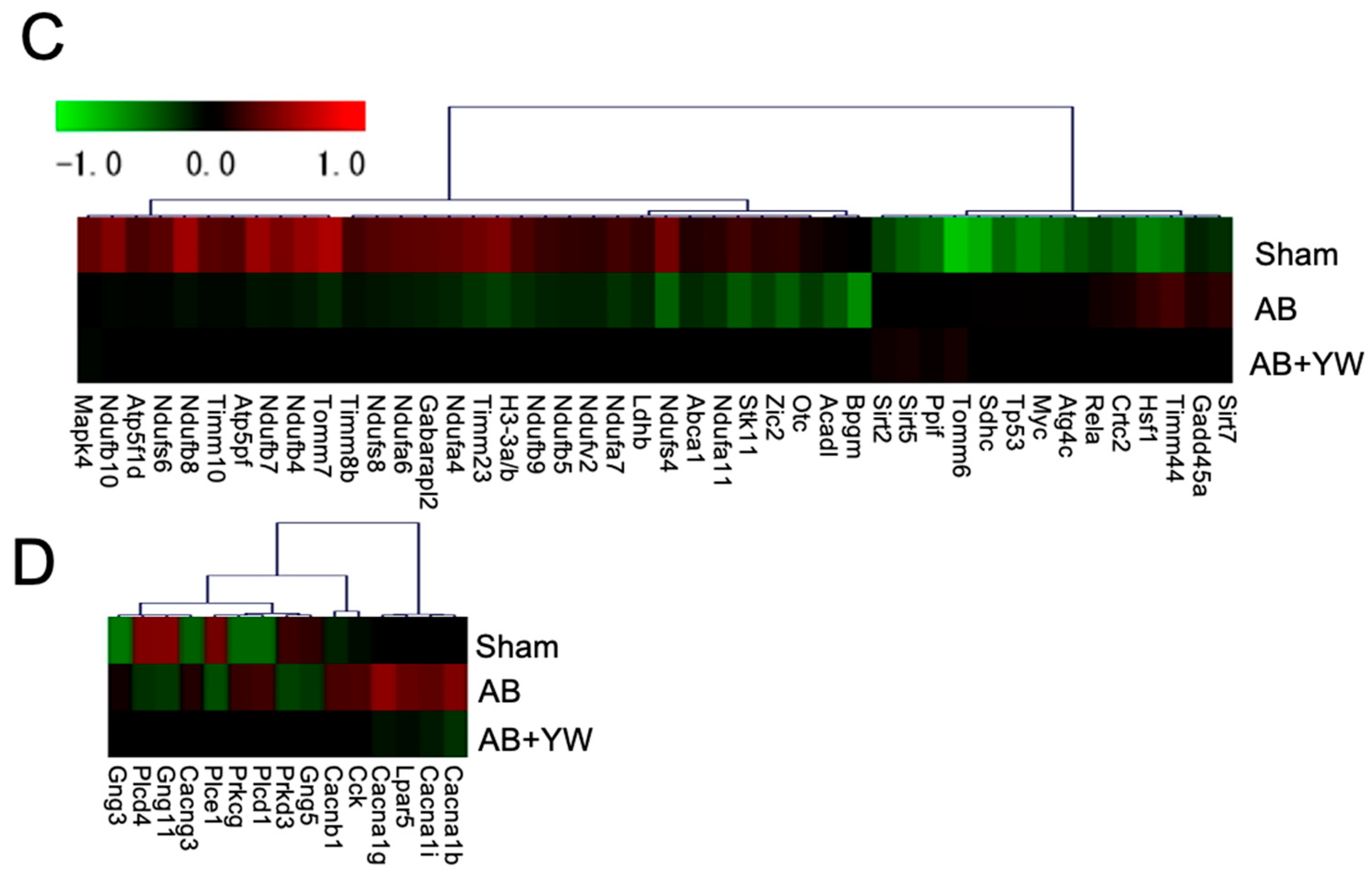
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peighambardoust, S.H.; Karami, Z.; Pateiro, M.; Lorenzo, J.M. A review on health-promoting, biological, and functional aspects of bioactive peptides in food application. Biomolecules 2021, 11, 631. [Google Scholar] [CrossRef]
- Wu, Y.H.; Samuel, C.; Chen, Y.; Wu, Y.H.S.; Chen, Y.C. Trends and applications of food protein-origin hydrolysates and bioactive peptides. J. Food Drug Anal. 2022, 30, 172–184. [Google Scholar] [CrossRef]
- Galland, F.; de Espindola, J.S.; Lopes, D.S.; Taccola, M.F.; Pacheco, M.T.B. Food-derived bioactive peptides: Mechanisms of action underlying inflammation and oxidative stress in the central nervous system. Food Chem. Adv. 2022, 1, 100087. [Google Scholar] [CrossRef]
- Wang, L.; Liu, S.; Xu, J.; Watanabe, N.; Mayo, K.H.; Li, J.; Li, X. Emodin inhibits aggregation of amyloid-β Peptide 1-42 and improves cognitive deficits in Alzheimer’s disease transgenic mice. J. Neurochem. 2021, 157, 1992–2007. [Google Scholar] [CrossRef] [PubMed]
- Imai, H.; Moriyasu, K.; Nakahata, A.; Maebuchi, M.; Ichinose, T.; Furuya, S. Soy peptide ingestion augments the synthesis and metabolism of noradrenaline in the mouse brain. Biosci. Biotechnol. Biochem. 2017, 81, 1007–1013. [Google Scholar] [CrossRef] [PubMed]
- Hino, Y.; Koyanagi, A.; Maebuchi, M.; Ichinose, T.; Furuya, S. comparison of the effect of soy and casein-derived peptide administration on tyrosine and catecholamine metabolism in the mouse brain. J. Nutr. Sci. Vitaminol. 2018, 64, 329–334. [Google Scholar] [CrossRef]
- Ichinose, T.; Moriyasu, K.; Nakahata, A.; Tanaka, M.; Matsui, T.; Furuya, S. Orally administrated dipeptide Ser–Tyr efficiently stimulates noradrenergic turnover in the mouse brain. Biosci. Biotechnol. Biochem. 2015, 79, 1542–1547. [Google Scholar] [CrossRef]
- Hanh, V.T.; Kobayashi, Y.; Maebuchi, M.; Nakamori, T.; Tanaka, M.; Matsui, T. Quantitative mass spectrometric analysis of dipeptides in protein hydrolysate by a TNBS derivatization-aided standard addition method. Food Chem. 2016, 190, 345–350. [Google Scholar] [CrossRef]
- Ichinose, T.; Murasawa, H.; Ishijima, T.; Okada, S.; Abe, K.; Matsumoto, S.; Matsui, T.; Furuya, S. Tyr–Trp administration facilitates brain norepinephrine metabolism and ameliorates a short-term memory deficit in a mouse model of Alzheimer’s disease. PLoS ONE 2020, 15, e0232233. [Google Scholar] [CrossRef]
- Hamano, M.; Sayano, T.; Kusada, W.; Kato, H.; Furuya, S. Microarray data on altered transcriptional program of Phgdh-deficient mouse embryonic fibroblasts caused by ʟ-serine depletion. Data Brief 2016, 7, 1598–1601. [Google Scholar] [CrossRef]
- Hamano, M.; Esaki, K.; Moriyasu, K.; Yasuda, T.; Mohri, S.; Tashiro, K.; Hirabayashi, Y.; Furuya, S. Hepatocyte-specific Phgdh-deficient mice culminate in mild obesity, insulin resistance, and enhanced vulnerability to protein starvation. Nutrients 2021, 13, 3468. [Google Scholar] [CrossRef]
- Kawakami, Y.; Yoshida, K.; Hoon, J.; Suzuki, T.; Azuma, N.; Sakai, K.; Hashikawa, T.; Watanabe, M.; Yasuda, K.; Kuhara, S.; et al. Impaired neurogenesis in embryonic spinal cord of Phgdh knockout mice, a serine deficiency disorder model. Neurosci. Res. 2009, 63, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Takashima, Y.; Hamano, M.; Yoshii, K.; Hayano, A.; Fukai, J.; Iwadate, Y.; Kajiwara, K.; Hondoh, H.; Yamanaka, R. Reciprocal expression of the immune response genes CXCR3 and IFI44L as module hubs are associated with patient survivals in primary central nervous system lymphoma. Int. J. Clin. Oncol. 2023, 28, 468–481. [Google Scholar] [CrossRef]
- Nakamura, A.; Fujiwara, S.; Ishijima, T.; Okada, S.; Nakai, Y.; Matsumoto, I.; Misaka, T.; Abe, K. Neuron differentiation-related genes are up-regulated in the hypothalamus of odorant-inhaling rats subjected to acute restraint stress. J. Agric. Food Chem. 2010, 58, 7922–7929. [Google Scholar] [CrossRef]
- Okada, S.; Abuyama, M.; Yamamoto, R.; Kondo, T.; Narukawa, M.; Misaka, T. Dietary zinc status reversibly alters both the feeding behaviors of the rats and gene expression patterns in diencephalon. BioFactors 2012, 38, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Bolstad, B.M.; Irizarry, R.A.; Åstrand, M.; Speed, T.P. A comparison of normalization methods for high density array data based on variance and bias. Bioinformatics 2003, 19, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Smyth, G.K. Limma: Linear Models for Microarray Data. In Bioinformatics and Computational Biology Solution Using R and Bioconductor; Springer: Berlin/Heidelberg, Germany, 2006; pp. 397–420. [Google Scholar]
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: Open software development for computational biology and bioinformatics. Genome Biol. 2004, 5, R80. [Google Scholar] [CrossRef] [PubMed]
- Dennis, G.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for annotation, visualization, and integrated discovery. Genome Biol. 2003, 4, P3. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING V11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [PubMed]
- Tsai, K.J.; Tsai, Y.C.; Shen, C.K.J. G-CSF rescues the memory impairment of animal models of Alzheimer’s disease. J. Exp. Med. 2007, 204, 1273–1280. [Google Scholar] [PubMed]
- Demuro, A.; Parker, I.; Stutzmann, G.E. Calcium signaling and amyloid toxicity in Alzheimer disease. J. Biol. Chem. 2010, 285, 12463–12468. [Google Scholar] [CrossRef] [PubMed]
- Wójtowicz, S.; Strosznajder, A.K.; Jeżyna, M.; Strosznajder, J.B. The novel role of PPAR alpha in the brain: Promising target in therapy of Alzheimer’s disease and other neurodegenerative disorders. Neurochem. Res. 2020, 45, 972–988. [Google Scholar] [CrossRef] [PubMed]
- Strosznajder, A.K.; Wójtowicz, S.; Jeżyna, M.J.; Sun, G.Y.; Strosznajder, J.B. Recent insights on the role of PPAR-β/δ in neuroinflammation and neurodegeneration, and its potential target for therapy. Neuromolecular Med. 2021, 23, 86–98. [Google Scholar] [CrossRef]
- Caccamo, A.; Maldonado, M.A.; Bokov, A.F.; Majumder, S.; Oddo, S. CBP gene Transfer increases BDNF levels and ameliorates learning and memory deficits in a mouse model of Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2010, 107, 22687–22692. [Google Scholar] [CrossRef]
- Gui, H.; Gong, Q.; Jiang, J.; Liu, M.; Li, H. Identification of the hub genes in Alzheimer’s disease. Comput. Math. Methods Med. 2021, 2021, 6329041. [Google Scholar]
- Tao, Y.; Han, Y.; Yu, L.; Wang, Q.; Leng, S.X.; Zhang, H. The predicted key molecules, functions, and pathways that bridge mild cognitive impairment (MCI) and Alzheimer’s disease (AD). Front. Neurol. 2020, 11, 233. [Google Scholar] [CrossRef]
- Khayer, N.; Mirzaie, M.; Marashi, S.A.; Jalessi, M. Rps27a might act as a controller of microglia activation in triggering neurodegenerative diseases. PLoS ONE 2020, 15, e0239219. [Google Scholar] [CrossRef]
- Yoshida, K.; Furuya, S.; Osuka, S.; Mitoma, J.; Shinoda, Y.; Watanabe, M.; Azuma, N.; Tanaka, H.; Hashikawa, T.; Itohara, S.; et al. Targeted disruption of the mouse 3-phosphoglycerate dehydrogenase gene causes severe neurodevelopmental defects and results in embryonic lethality. J. Biol. Chem. 2004, 279, 3573–3577. [Google Scholar] [CrossRef]
- Yang, J.H.; Wada, A.; Yoshida, K.; Miyoshi, Y.; Sayano, T.; Esaki, K.; Kinoshita, M.O.; Tomonaga, S.; Azuma, N.; Watanabe, M.; et al. Brain-specific Phgdh deletion reveals a pivotal role for l-serine biosynthesis in controlling the level of D-serine, an N-methyl-D-aspartate receptor co-agonist, in adult brain. J. Biol. Chem. 2010, 285, 41380–41390. [Google Scholar] [CrossRef] [PubMed]
- Hamano, M.; Haraguchi, Y.; Sayano, T.; Zyao, C.; Arimoto, Y.; Kawano, Y.; Moriyasu, K.; Udono, M.; Katakura, Y.; Ogawa, T.; et al. Enhanced vulnerability to oxidative stress and induction of inflammatory gene expression in 3-phosphoglycerate dehydrogenase-deficient fibroblasts. FEBS Open Bio 2018, 8, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Callahan, L.M.; Sul, J.Y.; Kim, T.K.; Barrett, L.; Kim, M.; Powers, J.M.; Federoff, H.; Eberwine, J. A neurotoxic phosphoform of Elk-1 associates with inclusions from multiple neurodegenerative diseases. PLoS ONE 2010, 5, e9002. [Google Scholar] [CrossRef]
- Frei, K.; Bodmer, S.; Schwerdel, C.; Fontana, A. Astrocyte-derived interleukin 3 as a growth factor for microglia cells and peritoneal macrophages. J. Immunol. 1986, 137, 3521–3527. [Google Scholar] [CrossRef]
- McAlpine, C.S.; Park, J.; Griciuc, A.; Kim, E.; Choi, S.H.; Iwamoto, Y.; Kiss, M.G.; Christie, K.A.; Vinegoni, C.; Poller, W.C.; et al. Astrocytic interleukin-3 programs microglia and limits Alzheimer’s disease. Nature 2021, 595, 701–706. [Google Scholar] [CrossRef]
- Rosa, P.; Zerbinati, C.; Crestini, A.; Canudas, A.M.; Ragona, G.; Confaloni, A.; Iuliano, L.; Calogero, A. Heme Oxygenase-1 and brain oxysterols metabolism are linked to Egr-1 expression in aged mice cortex, but not in hippocampus. Front. Aging Neurosci. 2018, 10, 363. [Google Scholar]
- Giri, R.K.; Selvaraj, S.K.; Kalra, V.K. Amyloid peptide-induced cytokine and chemokine expression in THP-1 monocytes is blocked by small inhibitory RNA duplexes for early growth Response-1 messenger RNA. J. Immunol. 2003, 170, 5281–5294. [Google Scholar]
- Hefter, D.; Ludewig, S.; Draguhn, A.; Korte, M. Amyloid, APP, and electrical activity of the brain. Neuroscientist 2020, 26, 231–251. [Google Scholar] [CrossRef]
- Hamano, T.; Shirafuji, N.; Yen, S.H.; Yoshida, H.; Kanaan, N.M.; Hayashi, K.; Ikawa, M.; Yamamura, O.; Fujita, Y.; Kuriyama, M.; et al. Rho-kinase ROCK inhibitors reduce oligomeric Tau protein. Neurobiol. Aging 2020, 89, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.; Huang, Y.; Jiang, Y.; Zou, L.; Liu, X.; Liu, S.; Chen, F.; Luo, J.; Zhu, Y. Ganoderma lucidum triterpenoids (GLTs) reduce neuronal apoptosis via inhibition of ROCK signal pathway in APP/PS1 transgenic Alzheimer’s disease mice. Oxid. Med. Cell Longev. 2020, 2020, 9894037. [Google Scholar] [CrossRef] [PubMed]
- Keringer, P.; Furedi, N.; Gaszner, B.; Miko, A.; Pakai, E.; Fekete, K.; Olah, E.; Kelava, L.; Romanovsky, A.A.; Rumbus, Z.; et al. The hyperthermic effect of central cholecystokinin is mediated by the cyclooxygenase-2 pathway. Am. J. Physiol. Endocrinol. Metab. 2022, 322, E10–E23. [Google Scholar] [CrossRef] [PubMed]
- Aisen, P.S. Evaluation of selective COX-2 inhibitors for the treatment of Alzheimer’s disease. J. Pain Symptom Manag. 2002, 23, S35–S40. [Google Scholar] [CrossRef]
- Fu, Y.; Young, J.F.; Løkke, M.M.; Lametsch, R.; Aluko, R.E.; Therkildsen, M. Revalorisation of bovine collagen as a potential precursor of angiotensin I-converting enzyme (ACE) inhibitory peptides based on in silico and in vitro protein digestions. J. Funct. Foods 2016, 24, 196–206. [Google Scholar] [CrossRef]
- Kitagaki, H. Medical application of substances derived from non-pathogenic fungi Aspergillus oryzae and A. luchuensis-containing koji. J. Fungi 2021, 7, 243. [Google Scholar] [CrossRef] [PubMed]
- Ayabe, T.; Ano, Y.; Ohya, R.; Kitaoka, S.; Furuyashiki, T. The lacto-tetrapeptide Gly–Thr–Trp–Tyr, β-Lactolin, improves spatial memory functions via dopamine release and D1 receptor activation in the hippocampus. Nutrients 2019, 11, 2469. [Google Scholar] [CrossRef] [PubMed]
- Ano, Y.; Ayabe, T.; Ohya, R.; Kondo, K.; Kitaoka, S.; Furuyashiki, T. Tryptophan-tyrosine dipeptide, the core sequence of β-Lactolin, improves memory by modulating the dopamine system. Nutrients 2019, 11, 348. [Google Scholar]
- Chakrabarti, S.; Guha, S.; Majumder, K. Food-derived bioactive peptides in human health: Challenges and opportunities. Nutrients 2018, 10, 1738. [Google Scholar] [CrossRef]
- Canet, G.; Zussy, C.; Hernandez, C.; Maurice, T.; Desrumaux, C.; Givalois, L. The pathomimetic oAβ25–35 model of Alzheimer’s disease: Potential for screening of new therapeutic agents. Pharmacol. Ther. 2023, 245, 108398. [Google Scholar] [CrossRef]
- D’Agostino, G.; Russo, R.; Avagliano, C.; Cristiano, C.; Meli, R.; Calignano, A. Palmitoylethanolamide protects against the amyloid-β25-35-induced learning and memory impairment in mice, an experimental model of Alzheimer disease. Neuropsychopharmacology 2012, 37, 1784–1792. [Google Scholar] [CrossRef]
- Stepanichev, M.Y.; Moiseeva, Y.V.; Lazareva, N.A.; Onufriev, M.V.; Gulyaeva, N.V. Single intracerebroventricular administration of amyloid-beta (25–35) peptide induces impairment in short-term rather than long-term memory in rats. Brain Res. Bull. 2003, 61, 197–205. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamano, M.; Ichinose, T.; Yasuda, T.; Ishijima, T.; Okada, S.; Abe, K.; Tashiro, K.; Furuya, S. Bioinformatics Analysis of the Molecular Networks Associated with the Amelioration of Aberrant Gene Expression by a Tyr–Trp Dipeptide in Brains Treated with the Amyloid-β Peptide. Nutrients 2023, 15, 2731. https://doi.org/10.3390/nu15122731
Hamano M, Ichinose T, Yasuda T, Ishijima T, Okada S, Abe K, Tashiro K, Furuya S. Bioinformatics Analysis of the Molecular Networks Associated with the Amelioration of Aberrant Gene Expression by a Tyr–Trp Dipeptide in Brains Treated with the Amyloid-β Peptide. Nutrients. 2023; 15(12):2731. https://doi.org/10.3390/nu15122731
Chicago/Turabian StyleHamano, Momoko, Takashi Ichinose, Tokio Yasuda, Tomoko Ishijima, Shinji Okada, Keiko Abe, Kosuke Tashiro, and Shigeki Furuya. 2023. "Bioinformatics Analysis of the Molecular Networks Associated with the Amelioration of Aberrant Gene Expression by a Tyr–Trp Dipeptide in Brains Treated with the Amyloid-β Peptide" Nutrients 15, no. 12: 2731. https://doi.org/10.3390/nu15122731
APA StyleHamano, M., Ichinose, T., Yasuda, T., Ishijima, T., Okada, S., Abe, K., Tashiro, K., & Furuya, S. (2023). Bioinformatics Analysis of the Molecular Networks Associated with the Amelioration of Aberrant Gene Expression by a Tyr–Trp Dipeptide in Brains Treated with the Amyloid-β Peptide. Nutrients, 15(12), 2731. https://doi.org/10.3390/nu15122731