
Lipoprotein Particle Profiles Associated with Telomere Length and Telomerase Complex Components

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Characteristics and Collection of Biological Samples
2.2. Lipoprotein Analysis by NMR Spectroscopy of Plasma Samples
2.3. Telomere Length Measurement
2.4. TERT and WRAP53 Expression
2.5. Statistical Analysis
3. Results
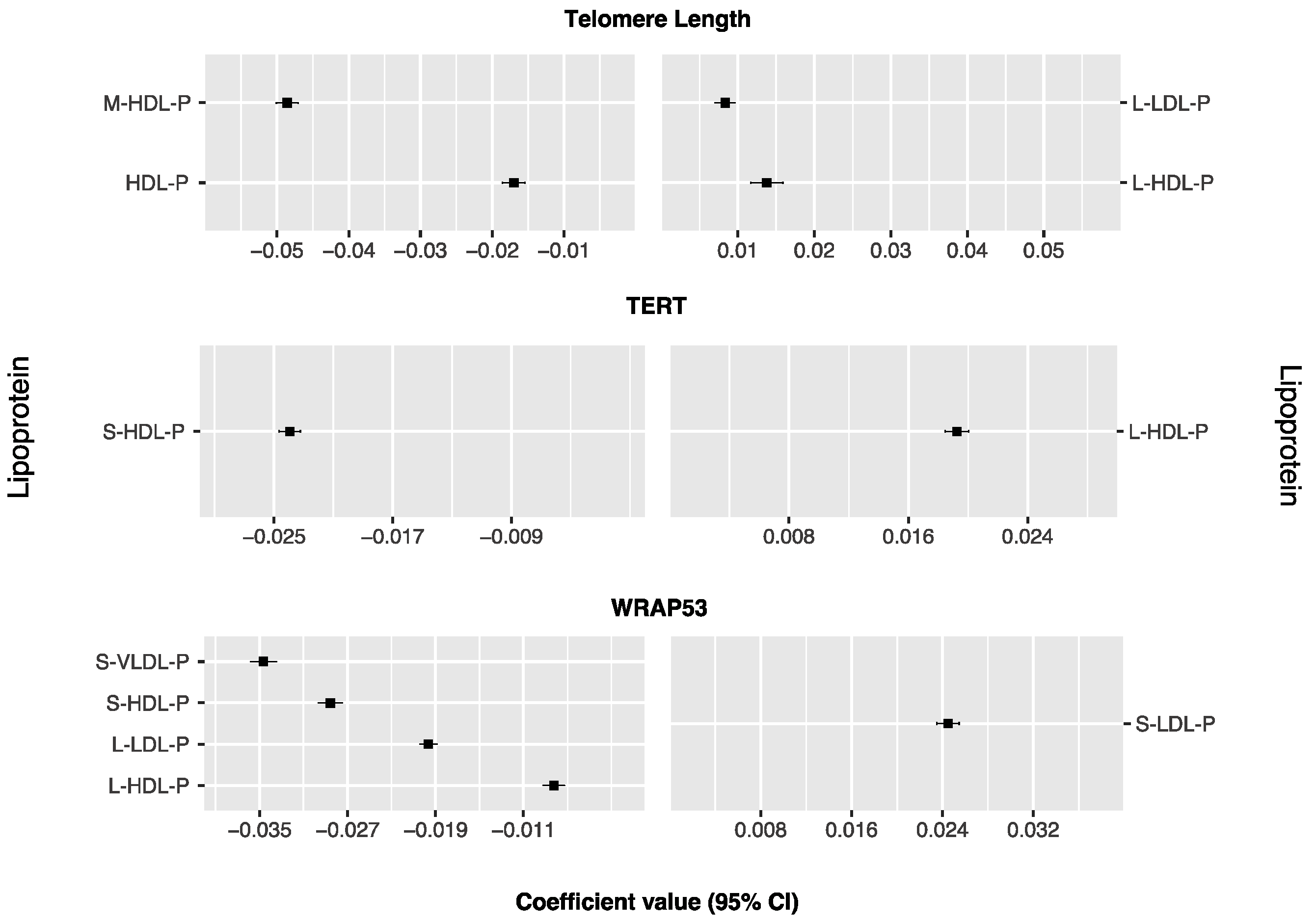
Associations of Lipoproteins and Their Subclasses with TL, TERT, and WRAP53
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vaiserman, A.; Krasnienkov, D. Telomere Length as a Marker of Biological Age: State-of-the-Art, Open Issues, and Future Perspectives. Front. Genet. 2021, 11, 1816. [Google Scholar] [CrossRef]
- Xu, X.; Hu, H.; Lin, Y.; Huang, F.; Ji, H.; Li, Y.; Lin, S.; Chen, X.; Duan, S. Differences in Leukocyte Telomere Length between Coronary Heart Disease and Normal Population: A Multipopulation Meta-Analysis. BioMed Res. Int. 2019, 2019, 5046867. [Google Scholar] [CrossRef]
- Haycock, P.C.; Heydon, E.E.; Kaptoge, S.; Butterworth, A.S.; Thompson, A.; Willeit, P. Leucocyte Telomere Length and Risk of Cardiovascular Disease: Systematic Review and Meta-Analysis. BMJ 2014, 349, g4227. [Google Scholar] [CrossRef] [PubMed]
- Aulinas, A.; Ramírez, M.J.; Barahona, M.J.; Valassi, E.; Resmini, E.; Mato, E.; Santos, A.; Crespo, I.; Bell, O.; Surrallés, J.; et al. Dyslipidemia and Chronic Inflammation Markers Are Correlated with Telomere Length Shortening in Cushing’s Syndrome. PLoS ONE 2015, 10, e0120185. [Google Scholar] [CrossRef]
- Baragetti, A.; Bonacina, F.; Da Dalt, L.; Moregola, A.; Zampoleri, V.; Pellegatta, F.; Grigore, L.; Pirillo, A.; Spina, R.; Cefalù, A.B.; et al. Genetically Determined Hypercholesterolaemia Results into Premature Leucocyte Telomere Length Shortening and Reduced Haematopoietic Precursors. Eur. J. Prev. Cardiol. 2022, 29, 721–729. [Google Scholar] [CrossRef]
- Tellechea, M.L.; Pirola, C.J. The Impact of Hypertension on Leukocyte Telomere Length: A Systematic Review and Meta-Analysis of Human Studies. J. Hum. Hypertens. 2017, 31, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Luk, A.O.; Shi, M.; Huang, C.; Jiang, G.; Yang, A.; Wu, H.; Lim, C.K.P.; Tam, C.H.T.; Fan, B.; et al. Shortened Leukocyte Telomere Length Is Associated with Glycemic Progression in Type 2 Diabetes: A Prospective and Mendelian Randomization Analysis. Diabetes Care 2022, 45, 701–709. [Google Scholar] [CrossRef]
- Rubtsova, M.P.; Vasilkova, D.P.; Malyavko, A.N.; Naraikina, Y.V.; Zvereva, M.I.; Dontsova, O.A. Telomere Lengthening and Other Functions of Telomerase. Acta Nat. 2012, 4, 44–61. [Google Scholar] [CrossRef]
- Narducci, M.L.; Grasselli, A.; Biasucci, L.M.; Farsetti, A.; Mulè, A.; Liuzzo, G.; La Torre, G.; Niccoli, G.; Mongiardo, R.; Pontecorvi, A.; et al. High Telomerase Activity in Neutrophils from Unstable Coronary Plaques. J. Am. Coll. Cardiol. 2007, 50, 2369–2374. [Google Scholar] [CrossRef] [PubMed]
- Richardson, G.D.; Breault, D.; Horrocks, G.; Cormack, S.; Hole, N.; Owens, W.A. Telomerase Expression in the Mammalian Heart. FASEB J. 2012, 26, 4832–4840. [Google Scholar] [CrossRef]
- Poch, E.; Carbonell, P.; Franco, S.; Díez-Juan, A.; Blasco, M.A.; Andrés, V. Short Telomeres Protect from Diet-Induced Atherosclerosis in Apolipoprotein E-Null Mice. FASEB J. 2004, 18, 418–420. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.; Taffet, G.E.; Youker, K.A.; Entman, M.L.; Overbeek, P.A.; Michael, L.H.; Schneider, M.D. Telomerase Reverse Transcriptase Promotes Cardiac Muscle Cell Proliferation, Hypertrophy, and Survival. Proc. Natl. Acad. Sci. USA 2001, 98, 10308. [Google Scholar] [CrossRef] [PubMed]
- Al Amri, T.; Bahijri, S.; Al-Raddadi, R.; Ajabnoor, G.; Al Ahmadi, J.; Jambi, H.; Borai, A.; Tuomilehto, J. The Association between Prediabetes and Dyslipidemia Among Attendants of Primary Care Health Centers in Jeddah, Saudi Arabia. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 2735. [Google Scholar] [CrossRef]
- Peng, J.; Zhao, F.; Yang, X.; Pan, X.; Xin, J.; Wu, M.; Peng, Y.G. Association between Dyslipidemia and Risk of Type 2 Diabetes Mellitus in Middle-Aged and Older Chinese Adults: A Secondary Analysis of a Nationwide Cohort. BMJ Open 2021, 11, e042821. [Google Scholar] [CrossRef] [PubMed]
- Rehkopf, D.H.; Needham, B.L.; Lin, J.; Blackburn, E.H.; Zota, A.R.; Wojcicki, J.M.; Epel, E.S. Leukocyte Telomere Length in Relation to 17 Biomarkers of Cardiovascular Disease Risk: A Cross-Sectional Study of US Adults. PLoS Med. 2016, 13, e1002188. [Google Scholar] [CrossRef]
- Chen, Y.F.; Zhou, K.W.; Yang, G.Z.; Chen, C. Association between Lipoproteins and Telomere Length in US Adults: Data from the NHANES 1999–2002. Lipids Health Dis. 2019, 18, 1–10. [Google Scholar] [CrossRef]
- Teis, A.; Cediel, G.; Amigó, N.; Julve, J.; Aranyó, J.; Andrés-Cordón, J.; Puig-Jové, C.; Castelblanco, E.; Gual-Capllonch, F.; Ferrer-Sistach, E.; et al. Particle Size and Cholesterol Content of Circulating HDL Correlate with Cardiovascular Death in Chronic Heart Failure. Sci. Rep. 2021, 11, 1–9. [Google Scholar] [CrossRef]
- Rizvi, A.A.; Pantea Stoian, A.; Janez, A.; Rizzo, M.; Lipoproteins, M.; Iannuzzo, G.; Gentile, M. Lipoproteins and Cardiovascular Disease: An Update on the Clinical Significance of Atherogenic Small, Dense LDL and New Therapeutical Options. Biomedicines 2021, 9, 1579. [Google Scholar] [CrossRef]
- Soria-Florido, M.T.; Castañer, O.; Lassale, C.; Estruch, R.; Salas-Salvadó, J.; Martínez-González, M.Á.; Corella, D.; Ros, E.; Arós, F.; Elosua, R.; et al. Dysfunctional High-Density Lipoproteins Are Associated with a Greater Incidence of Acute Coronary Syndrome in a Population at High Cardiovascular Risk: A Nested Case-Control Study. Circulation 2020, 141, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Noureen, N.; Wu, S.; Lv, Y.; Yang, J.; Alfred Yung, W.K.; Gelfond, J.; Wang, X.; Koul, D.; Ludlow, A.; Zheng, S. Integrated Analysis of Telomerase Enzymatic Activity Unravels an Association with Cancer Stemness and Proliferation. Nat. Commun. 2021, 12, 139. [Google Scholar] [CrossRef]
- Zhu, Y.; Ding, L.; Chen, B.F.; Song, J.G.; Yao, Y.S. Oncogenic Activity of Wrap53 in Human Colorectal Cancer In Vitro and in Nude Mouse Xenografts. Med. Sci. Monit. 2018, 24, 6129. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Alonso, P.; Salas-Salvadó, J.; Baldrich-Mora, M.; Mallol, R.; Correig, X.; Bulló, M. Effect of Pistachio Consumption on Plasma Lipoprotein Subclasses in Pre-Diabetic Subjects. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 396–402. [Google Scholar] [CrossRef]
- Elosua, R.; Marrugat, J.; Molina, L.; Pons, S.; Pujol, E. Validation of the Minnesota Leisure Time Physical Activity Questionnaire in Spanish Men. The MARATHOM Investigators. Am. J. Epidemiol. 1994, 139, 1197–1209. [Google Scholar] [CrossRef]
- Mallol, R.; Amigó, N.; Rodríguez, M.A.; Heras, M.; Vinaixa, M.; Plana, N.; Rock, E.; Ribalta, J.; Yanes, O.; Masana, L.; et al. Liposcale: A Novel Advanced Lipoprotein Test Based on 2D Diffusion-Ordered 1H NMR Spectroscopy. J. Lipid Res. 2015, 56, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Cawthon, R.M. Telomere Length Measurement by a Novel Monochrome Multiplex Quantitative PCR Method. Nucleic Acids Res. 2009, 37, e21. [Google Scholar] [CrossRef] [PubMed]
- Canudas, S.; Hernández-Alonso, P.; Gali, S.; Muralidharan, J.; Morell-Azanza, L.; Zalba, G.; García-Gavilán, J.; Martí, A.; Salas-Salvadó, J.; Bulló, M. Pistachio Consumption Modulates DNA Oxidation and Genes Related to Telomere Maintenance: A Crossover Randomized Clinical Trial. Am. J. Clin. Nutr. 2019, 109, 1738–1745. [Google Scholar] [CrossRef]
- Stekhoven, D.J.; Bühlmann, P. MissForest—Non-Parametric Missing Value Imputation for Mixed-Type Data. Bioinformatics 2012, 28, 112–118. [Google Scholar] [CrossRef]
- White, I.R.; Royston, P.; Wood, A.M. Multiple Imputation Using Chained Equations: Issues and Guidance for Practice. Stat. Med. 2011, 30, 377–399. [Google Scholar] [CrossRef]
- Kuhn, M. Building Predictive Models in R Using the Caret Package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef]
- Huang, Y.; Dai, W.; Li, Y. Potential Associations of Testosterone/Estradiol Ratio, Leukocyte HTERT Expression and PBMC Telomerase Activity with Aging and the Presence of Coronary Artery Disease in Men. Exp. Gerontol. 2019, 117, 38–44. [Google Scholar] [CrossRef]
- Metcalfe, N.B.; Olsson, M. How Telomere Dynamics Are Influenced by the Balance between Mitochondrial Efficiency, Reactive Oxygen Species Production and DNA Damage. Mol. Ecol. 2022, 31, 6040–6052. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhu, Y.; Uppal, K.; Tran, V.L.T.; Yu, T.; Lin, J.; Matsuguchi, T.; Blackburn, E.; Jones, D.; Lee, E.T.; et al. Metabolic Profiles of Biological Aging in American Indians: The Strong Heart Family Study. Aging 2014, 6, 176. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Gardner, J.P.; Kimura, M.; Brimacombe, M.; Cao, X.; Srinivasan, S.R.; Berenson, G.S.; Aviv, A. Leukocyte Telomere Length Is Associated with HDL Cholesterol Levels: The Bogalusa Heart Study. Atherosclerosis 2009, 205, 620–625. [Google Scholar] [CrossRef]
- Bindu, G.H.; Rao, V.S.; Kakkar, V.V. Friend Turns Foe: Transformation of Anti-Inflammatory HDL to Proinflammatory HDL during Acute-Phase Response. Cholesterol 2011, 2011, 274629. [Google Scholar] [CrossRef]
- Soppert, J.; Lehrke, M.; Marx, N.; Jankowski, J.; Noels, H. Lipoproteins and Lipids in Cardiovascular Disease: From Mechanistic Insights to Therapeutic Targeting. Adv. Drug Deliv. Rev. 2020, 159, 4–33. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.E.; Sinning, C.; Kontush, A. HDL Particle Number and Size as Predictors of Cardiovascular Disease. Front. Pharmacol. 2015, 6, 218. [Google Scholar] [CrossRef]
- Pichler, G.; Amigo, N.; Tellez-Plaza, M.; Pardo-Cea, M.A.; Dominguez-Lucas, A.; Marrachelli, V.G.; Monleon, D.; Martin-Escudero, J.C.; Ascaso, J.F.; Chaves, F.J.; et al. LDL Particle Size and Composition and Incident Cardiovascular Disease in a South-European Population: The Hortega-Liposcale Follow-up Study. Int. J. Cardiol. 2018, 264, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.M.; Nova, E.; Anil, E.; Jackson, K.; Bateman, P.; Wolstencroft, E.; Williams, C.M.; Yaqoob, P. Differential Uptake of Subfractions of Triglyceride-Rich Lipoproteins by THP-1 Macrophages. Atherosclerosis 2005, 180, 233–244. [Google Scholar] [CrossRef]
- Paynter, N.P.; Sesso, H.D.; Conen, D.; Otvos, J.D.; Mora, S. Lipoprotein Subclass Abnormalities and Incident Hypertension in Initially Healthy Women. Clin Chem. 2011, 57, 1178–1187. [Google Scholar] [CrossRef]
- Karimi, B.; Yunesian, M.; Nabizadeh, R.; Mehdipour, P. Serum Level of Total Lipids and Telomere Length in the Male: A Cross-Sectional Study. Am. J. Mens. Health 2019, 13, 1557988319842973. [Google Scholar] [CrossRef]
- Van Der Spek, A.; Karamujić-Čomić, H.; Pool, R.; Bot, M.; Beekman, M.; Garmaeva, S.; Arp, P.P.; Henkelman, S.; Liu, J.; Alves, A.C.; et al. Fat Metabolism Is Associated with Telomere Length in Six Population-Based Studies. Hum. Mol. Genet. 2022, 31, 1159–1170. [Google Scholar] [CrossRef]
- Lin, J.; Epel, E. Stress and Telomere Shortening: Insights from Cellular Mechanisms. Ageing Res. Rev. 2022, 73, 101507. [Google Scholar] [CrossRef]
- Herrmann, W.; Herrmann, M. The Importance of Telomere Shortening for Atherosclerosis and Mortality. J. Cardiovasc. Dev. Dis. 2020, 7, 29. [Google Scholar] [CrossRef] [PubMed]
- Demissie, S.; Levy, D.; Benjamin, E.J.; Cupples, L.A.; Gardner, J.P.; Herbert, A.; Kimura, M.; Larson, M.G.; Meigs, J.B.; Keaney, J.F.; et al. Insulin Resistance, Oxidative Stress, Hypertension, and Leukocyte Telomere Length in Men from the Framingham Heart Study. Aging Cell 2006, 5, 325–330. [Google Scholar] [CrossRef]
- Landmesser, U.; Hazen, S. HDL-Cholesterol, Genetics, and Coronary Artery Disease: The Myth of the ‘Good Cholesterol’? Eur. Heart J. 2018, 39, 2179–2182. [Google Scholar] [CrossRef]
- Morgantini, C.; Natali, A.; Boldrini, B.; Imaizumi, S.; Navab, M.; Fogelman, A.M.; Ferrannini, E.; Reddy, S.T. Anti-Inflammatory and Antioxidant Properties of HDLs Are Impaired in Type 2 Diabetes. Diabetes 2011, 60, 2617. [Google Scholar] [CrossRef] [PubMed]
- Arsenault, B.J.; Lemieux, I.; Després, J.P.; Gagnon, P.; Wareham, N.J.; Stroes, E.S.G.; Kastelein, J.J.P.; Khaw, K.T.; Boekholdt, S.M. HDL Particle Size and the Risk of Coronary Heart Disease in Apparently Healthy Men and Women: The EPIC-Norfolk Prospective Population Study. Atherosclerosis 2009, 206, 276–281. [Google Scholar] [CrossRef]
- Bergstrand, S.; Böhm, S.; Malmgren, H.; Norberg, A.; Sundin, M.; Nordgren, A.; Farnebo, M. Biallelic Mutations in WRAP53 Result in Dysfunctional Telomeres, Cajal Bodies and DNA Repair, Thereby Causing Hoyeraal–Hreidarsson Syndrome. Cell Death Dis. 2020, 11, 238. [Google Scholar] [CrossRef] [PubMed]
- Hedström, E.; Pederiva, C.; Farnebo, J.; Nodin, B.; Jirström, K.; Brennan, D.J.; Farnebo, M. Downregulation of the Cancer Susceptibility Protein WRAP53β in Epithelial Ovarian Cancer Leads to Defective DNA Repair and Poor Clinical Outcome. Cell Death Dis. 2015, 6, e1892. [Google Scholar] [CrossRef] [PubMed]
- Gadelha, R.B.; Machado, C.B.; Pessoa, F.M.C.d.P.; Pantoja, L.d.C.; Barreto, I.V.; Ribeiro, R.M.; de Moraes Filho, M.O.; de Moraes, M.E.A.; Khayat, A.S.; Moreira-Nunes, C.A. The Role of WRAP53 in Cell Homeostasis and Carcinogenesis Onset. Curr. Issues Mol. Biol. 2022, 44, 5498–5515. [Google Scholar] [CrossRef]


{kind=link}
| Variable | Subjects (n = 54) |
|---|---|
| Age, years | 55 (6.25) |
| Female, n (%) | 25 (46) |
| Weight, kg | 75.70 [71.00, 82.15] |
| Body mass index, kg/m2 | 28.86 (2.55) |
| Dyslipidemia, n (%) | 27 (50) |
| Acetylsalicylic Acid, n (%) | 3 (0.01) |
| Statins, n (%) | 5 (9.3) |
| Leisure-time physical activity (Kcal/day) | 347.40 (145.33) |
| Fasting plasma glucose (mg/dL) | 110.33 (4.28) |
| Telomere length | 0.76 [0.63, 0.88] |
| TERT | 8.57 [7.37, 11.05] |
| WRAP53 | 3.46 [2.93, 3.72] |
| Metabolites | r (95% CI) | p-Value |
|---|---|---|
| Telomere length | 0.347 (0.088, 0.563) | 0.010 * |
| TERT | 0.316 (0.052, 0.538) | 0.020 * |
| WRAP53 | 0.379 (0.124, 0.587) | 0.005 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novau-Ferré, N.; Rojas, M.; Gutierrez-Tordera, L.; Arcelin, P.; Folch, J.; Papandreou, C.; Bulló, M. Lipoprotein Particle Profiles Associated with Telomere Length and Telomerase Complex Components. Nutrients 2023, 15, 2624. https://doi.org/10.3390/nu15112624
Novau-Ferré N, Rojas M, Gutierrez-Tordera L, Arcelin P, Folch J, Papandreou C, Bulló M. Lipoprotein Particle Profiles Associated with Telomere Length and Telomerase Complex Components. Nutrients. 2023; 15(11):2624. https://doi.org/10.3390/nu15112624
Chicago/Turabian StyleNovau-Ferré, Nil, Melina Rojas, Laia Gutierrez-Tordera, Pierre Arcelin, Jaume Folch, Christopher Papandreou, and Mònica Bulló. 2023. "Lipoprotein Particle Profiles Associated with Telomere Length and Telomerase Complex Components" Nutrients 15, no. 11: 2624. https://doi.org/10.3390/nu15112624
APA StyleNovau-Ferré, N., Rojas, M., Gutierrez-Tordera, L., Arcelin, P., Folch, J., Papandreou, C., & Bulló, M. (2023). Lipoprotein Particle Profiles Associated with Telomere Length and Telomerase Complex Components. Nutrients, 15(11), 2624. https://doi.org/10.3390/nu15112624