
What Is the Evidence for Dietary-Induced DHA Deficiency in Human Brains?


Abstract
1. Introduction
2. Results
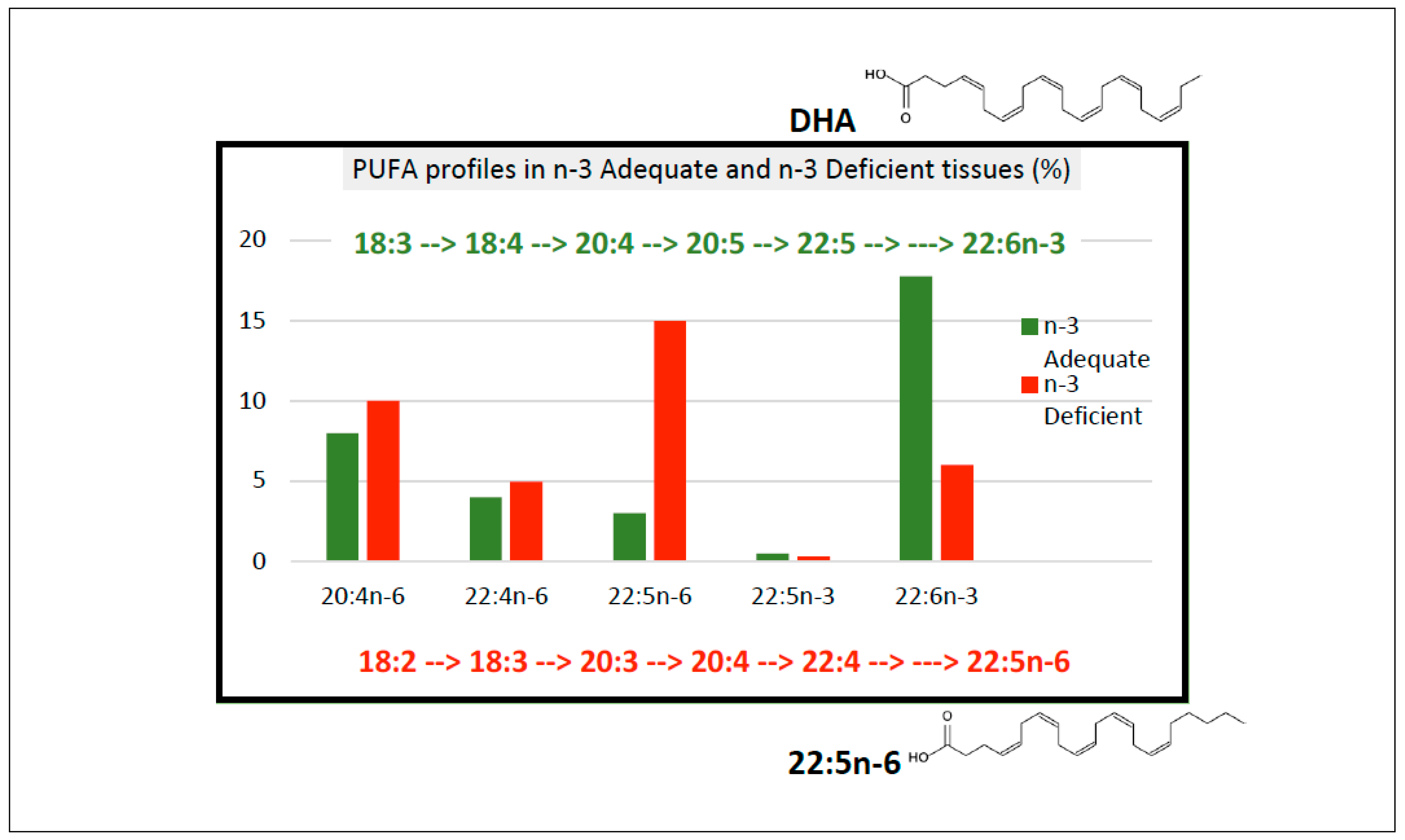
2.1. Postnatal Infant Brain Tissue Shows Evidence of Biochemical n-3 Deficiency
2.2. Do Human Diets Resemble Those Used to Induce n-3 PUFA Deficiency?

{kind=link}
{kind=link}
{kind=link}
| Reference | PUFA | LA | ALA | LA/ALA | LCn-3 Fatty Acids mg/Day |
|---|---|---|---|---|---|
| Harika et al. [47] (various countries) 1 | 4.7–8.9 1 | 2.6–8.6 | 0.14–1.0 | 3.9–29 | NR–419 |
| Pinto et al. [43] (omnivores UK) | 6.0 | 3.5 | 0.34 | 10.2 | 590 |
| Miyake et al. [49] (pregnant women Japan) | 6.7 | 5.4 | 0.9 | 6.3 | 460 |
| Lakshmipriya et al. [50] (M & F India) 2 | 5.2 | 5.0 | 0.22 | 22.7 | 5 |
| Pinto et al. [43] (vegans UK) | 5.6 | 5.2 | 0.4 | 13.0 | 10 |
| Agren et al. [44] (vegans Finland) | 9.7 | 8.1 | 1.6 | 5.1 | 0 |
| Mann et al. [42] (vegans Australia) | 6.4 | 6.1 | 0.3 | 18.7 | 0 |
| Chamorro et al. [45] (vegan males USA) | 13.9 | 11.1 | 2.4 | 4.6 | 0 |
2.3. Do Tissue Samples from Humans Show Evidence of n-3 PUFA Deficiency?
2.3.1. Plasma and Red Blood Cells in Adults
2.3.2. Cord Blood and Umbilical Cord Artery DHA Levels
2.3.3. Adipose Tissue
2.3.4. Milk DHA
2.3.5. Are the Low Levels of DHA in Some Breast Milk Samples Sufficient to Support the Accretion of DHA in the Infant Brain?
2.3.6. Do Post-Mortem Brain Samples Show Evidence of DHA Deficiency?
Biochemical Indication of n-3 PUFA Deficiency in Infant Human Brain
Biochemical Indication of n-3 PUFA Deficiency in Young Children and Adolescents
Biochemical Indication of n-3 PUFA Deficiency in Adult Human Brains
Variability in the PUFA Proportions in Brain Regions
3. Discussion
3.1. Plasma, Red Blood Cells, Adipose, Milk and Artery Data
3.2. Brain Data
3.2.1. What Are the Implications and Limitations of the Brain Data?
3.2.2. What Are the Most Likely Places and Conditions to Find Dietary Insufficiency of n-3 Fatty Acids?
3.2.3. What Are the Conditions and Situations Where Brain DHA Levels Are Vulnerable to Depletion?
3.2.4. Unknowns
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| Authors, Year | Country | Disease | Sample Numbers Patients; Controls | PM Interval hrs P; C | Brain Region (Brain Lipid Fraction Analysed) | Patient C22:6n-3 | Patient C22:5n-6 | Control C22:6n-3 | Control C22:5n-6 |
|---|---|---|---|---|---|---|---|---|---|
| Horrobin et al. [84] | Canada | Schizophrenia | 7; 7 | 20; 18 | Frontal Cortex; Cerebellar cortex (PE fraction) | FC: 34.2 ± 4.4 CC: 30.6 ± 3.2 | 2.8 ± 0.8 2.2 ± 0.4 | 34.0 ± 3.5 29.1 ± 2.4 | 2.3 ± 0.8 1.9 ± 0.6 |
| Soderberg et al. [74] | Sweden | Ageing and Alzheimer’s | 8–10; 8–10 | 24; 24 | Frontal GM, WM, Hippocampus, Pons (PE fraction) | 12.6 2.0 7.9 4.1 | NR | 23.5 4.6 16.9 5.7 | NR |
| Skinner et al. [85] | UK | Alzheimer’s | 15; 10 | NR | Anterior frontal near the pole. Parahippocampal gyrus (Total Lipids) | 17.1 ± 4.1 17.33 ± 0.86 | NR | 17.6 ± 1.3 18.64 ± 2.06 | NR |
| Yao et al. [86] | US | Schizophrenia | 11; 21 | 21; 18 | Caudate region (Total Lipids) | 8.64 ± 2.82 | NR | Control1 9.66 ± 1.63, Control2 10.53 ± 2.27 | NR |
| Landen et al. [87] | Sweden | Schizophrenia | 11; 13 | 40; 75 | Anterior portion of the left gyrus cinguli (Phospholipids) | 15.8 ± 0.69 | 0.75 ± 0.21 | 15.9 ± 0.66 | 0.58 ± 0.13 |
| Lalovic et al. [88] | Canada | Completed suicide with or without major depression (MD) | 16, 23; 19 | 27, 25; 24 | Orbitofrontal cortex (BA 11). Ventral prefrontal cortex (BA 47) (Total Lipids) | Su14.48 ± 2.42, MD14.64 ± 2.17 Su14.66 ± 2.28, MD14.77 ± 1.54 | NR | 15.12 ± 1.09 14.42 ± 2.99 | NR |
| Igarashi et al. [89] | US | Bipolar disorder | 8; 6 | 21; 27 | Prefrontal cortex (Total Lipids) | 8.8 ± 2.1 mmol/g | 1.2 ± 0.6 | 10.2 ± 1.8 | 1.0 ± 0.3 |
| McNamara et al. [90] | US | Suicide | 20; 20 | 21; 20 | Prefrontal cortex (BA 10) (Total Lipids) | 11.3 ± 0.4 (values estimated from figure) | 1.8 ± 0.1 | 11.2 ± 0.6 | 1.6 ± 0.1 |
| Conklin et al. [91] | US | Major depressive disorder and bipolar disorder | 12; 14 | 14; 14 | Anterior cingulate cortex (BA-24) region (Total Lipids) | 5.38 ± 2.15 | 5.90 ±1.29 | ||
| Fraser et al. [92] | UK | Alzheimer’s | 114; 58 | 45; 45 | Mid-frontal cortex (BA6) Superior temporal cortex (BA22) Angular gyrus (parietal cortex) (BA39) | 15.16 ± 2.22 14.51 ± 2.42 15.37 ± 1.95 | 1.16 ± 0.37 1.02 ± 0.38 1.07 ± 0.35 | 15.42 ± 1.51 14.98 ± 1.53 15.72 ± 2.05 | 1.12 ± 0.5 1.08 ± 0.41 1.01 ± 0.43 |
| Fabelo et al. [75] | Spain | Parkinson’s disease, early stage PD, incidental PD | 8, 8; 11 | 7, 15; 7 | Frontal cortex GM, isolated lipid rafts (Total Lipids) | PD: 1.71 ± 0.22 iPD: 1.41 ± 0.41 | 0.09 ± 0.04 0.08 ± 0.04 | 6.87 ± 0.34 | 0.46 ± 0.06 |
| Hamazaki et al. [93] | US | Schizophrenia, Bipolar Disorder, and Major Depressive Disorder | 15, 15, 15; 15 | 34, 33, 27; 24 | Amygdala (Phospholipids) | Sc:9.97 ± 1.07, BD:10.03 ± 1.28, MD:9.26 ± 1.32 | Sc:0.03 ± 0.02, BD:0.03 ± 0.02, MD:0.03 ± 0.02 | 9.71 ± 1.13 | 0.02 ± 0.01 |
| Taha et al. [94] | US, Harvard Br Tissue | Schizophrenia | 10; 10 | 22; 21 | Prefrontal cortex (Total Lipids) | 12.2 ± 1.4 | 1.4 ± 0.5 | 13.5 ± 0.9 | 1.5 ± 0.5 |
| Hamazaki et al. [95] | US | Schizophrenia, Bipolar Disorder, and Major Depressive Disorder | 15, 15, 15; 15 | 34, 33, 27; 24 | Entorhinal cortex tissue (Phospholipids) | Sc:11.2 ± 1.5, BD:11.8 ± 0.9, MD:11.1 ± 1.4 | 11.9 ± 0.9 | ||
| McNamara et al. [96] | US, Stanley FNC | Bipolar Disorder, Major Depressive Disorder, Schizophrenia | 15, 15, 15; 15 | 33, 28, 34; 24 | Superior temporal gyrus (Total Lipids) | BD:12.3 ± 1.8; MDD:10.4 ± 2.8; Sc:9.7 ± 3.2 | BD:1.8 ± 0.5; MDD:1.5 ± 0.5; Sc:1.3 ± 0.5 | 9.4 ± 2.9 | 1.5 ± 0.5 |
| Hamazaki et al. [97] | Japan | Schizophrenia, Bipolar Disorder, and Major Depressive Disorder | 15, 15, 15; 15 | 43, 36, 42; 43 | Frontal cortex (BA8) (Phospholipids) | Sc:14.97 (13.98, 15.63), BD:14.92 (14.15, 15.25), MDD:15.04 (13.82, 15.55) | 15.90 (14.27, 16.27) | ||
| Hamazaki et al. [98] | Japan | Schizophrenia | 95; 93 | 42; 42 | Frontal cortex (BA8), (Phospholipids) | 13.81 (12.39, 14.51) | NR | PL:14.12 (13.08, 14.75) | NR |
| Hamazaki et al. [99] | Japan | Schizophrenia, Bipolar Disorder, and Major Depressive Disorder | 15, 15, 15; 15 | 43, 36, 42; 43 | Corpus callosum (PL) | Sc:1.96 (1.84, 2.18), BD:2.02 (1.83, 2.18), MD:1.95 (1.91,1.98) | NR | PL:1.98 (1.83, 2.09) | NR |
| Ghosh et al. [100] | US, Stanley FNC | Schizophrenia, Bipolar disorder | 34, 35; 35 | 38, 31; 29 | Prefrontal white matter (PE fraction) | BD:3.02 ± 0.7Sc:3.12 ± 0.86 | BD:1.07 ± 0. 31SSc:1.09 ± 0.36 | 3.24 ± 0.79 | 1.30 ± 0.34 |
| Beasley et al. [101] | US, Stanley FNC | Schizophrenia, Bipolar disorder | 34, 35; 35 | 31, 38; 29 | Prefrontal cortex grey matter (PE fraction) | Sc:15.1 ± 0.7 BD:15.7 ± 0.6 | Sc:2.1 ± 0.1 BD:2.0 ± 0.1 | 15.2 ± 0.6 | 2.5 ± 0.2 |
References
- Ardesch, D.J.; Scholtens, L.H.; van den Heuvel, M.P. The human connectome from an evolutionary perspective. Prog. Brain Res. 2019, 250, 129–151. [Google Scholar] [CrossRef] [PubMed]
- Bethlehem, R.A.I.; Seidlitz, J.; White, S.R.; Vogel, J.W.; Anderson, K.M.; Adamson, C.; Adler, S.; Alexopoulos, G.S.; Anagnostou, E.; Areces-Gonzalez, A.; et al. Brain charts for the human lifespan. Nature 2022, 604, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Lacombe, R.J.S.; Smith, M.E.; Perlman, K.; Turecki, G.; Mechawar, N.; Bazinet, R.P. Quantitative and carbon isotope ratio analysis of fatty acids isolated from human brain hemispheres. J. Neurochem. 2023, 164, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, A.J. Docosahexaenoic acid and the brain- what is its role? Asia Pac. J. Clin. Nutr. 2019, 28, 675–688. [Google Scholar] [CrossRef]
- von Schacky, C. Importance of EPA and DHA Blood Levels in Brain Structure and Function. Nutrients 2021, 13, 1074. [Google Scholar] [CrossRef] [PubMed]
- Ind Anal Report 2020–2028; Fortune Business Insights. Available online: https://www.fortunebusinessinsights.com/industry-reports/omega-3-fatty-acids-market-100248 (accessed on 26 December 2022).
- Vandal, M.; Alata, W.; Tremblay, C.; Rioux-Perreault, C.; Salem, N., Jr.; Calon, F.; Plourde, M. Reduction in DHA transport to the brain of mice expressing human APOE4 compared to APOE2. J. Neurochem. 2014, 129, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, R.; Yang, T.; Xu, N.; Chen, J.; Gao, Y.; Stetler, R.A. Fatty acid transporting proteins: Roles in brain development, aging, and stroke. Prostaglandins Leukot Essent Fat. Acids 2018, 136, 35–45. [Google Scholar] [CrossRef]
- Fernandez, R.F.; Pereyra, A.S.; Diaz, V.; Wilson, E.S.; Litwa, K.A.; Martínez-Gardeazabal, J.; Jackson, S.N.; Brenna, J.T.; Hermann, B.P.; Eells, J.B.; et al. Acyl-CoA synthetase 6 is required for brain docosahexaenoic acid retention and neuroprotection during aging. JCI Insight 2021, 6, e144351. [Google Scholar] [CrossRef]
- McNamara, R.K. Long-chain omega-3 fatty acid deficiency in mood disorders: Rationale for treatment and prevention. Curr. Drug Discov. Technol. 2013, 10, 233–244. [Google Scholar] [CrossRef]
- Martinat, M.; Rossitto, M.; Di Miceli, M.; Layé, S. Perinatal Dietary Polyunsaturated Fatty Acids in Brain Development, Role in Neurodevelopmental Disorders. Nutrients 2021, 13, 1185. [Google Scholar] [CrossRef]
- Schaeffer, L.; Gohlke, H.; Müller, M.; Heid, I.M.; Palmer, L.J.; Kompauer, I.; Demmelmair, H.; Illig, T.; Koletzko, B.; Heinrich, J. Common genetic variants of the FADS1 FADS2 gene cluster and their reconstructed haplotypes are associated with the fatty acid composition in phospholipids. Hum. Mol. Genet. 2006, 15, 1745–1756. [Google Scholar] [CrossRef] [PubMed]
- Kothapalli, K.S.; Ye, K.; Gadgil, M.S.; Carlson, S.E.; O’Brien, K.O.; Zhang, J.Y.; Park, H.G.; Ojukwu, K.; Zou, J.; Hyon, S.S.; et al. Positive Selection on a Regulatory Insertion-Deletion Polymorphism in FADS2 Influences Apparent Endogenous Synthesis of Arachidonic Acid. Mol. Biol. Evol. 2016, 33, 1726–1739. [Google Scholar] [CrossRef] [PubMed]
- Stark, K.D.; Beblo, S.; Murthy, M.; Whitty, J.E.; Buda-Abela, M.; Janisse, J.; Rockett, H.; Martier, S.S.; Sokol, R.J.; Hannigan, J.H.; et al. Alcohol consumption in pregnant, black women is associated with decreased plasma and erythrocyte docosahexaenoic acid. Alcohol. Clin. Exp. Res. 2005, 29, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Osman, R.H.; Liu, L.; Xia, L.; Zhao, X.; Wang, Q.; Sun, X.; Zhang, Y.; Yang, B.; Zheng, Y.; Gong, D.; et al. Fads1 and 2 are promoted to meet instant need for long-chain polyunsaturated fatty acids in goose fatty liver. Mol. Cell Biochem. 2016, 418, 103–117. [Google Scholar] [CrossRef] [PubMed]
- DeMar, J.C., Jr.; Ma, K.; Bell, J.M.; Rapoport, S.I. Half-lives of docosahexaenoic acid in rat brain phospholipids are prolonged by 15 weeks of nutritional deprivation of n-3 polyunsaturated fatty acids. J. Neurochem. 2004, 91, 1125–1137. [Google Scholar] [CrossRef]
- Abedin, L.; Lien, E.L.; Vingrys, A.J.; Sinclair, A.J. The effects of dietary α-linolenic acid compared with docosahexaenoic acid on brain, retina, liver, and heart in the guinea pig. Lipids 1999, 34, 475–482. [Google Scholar] [CrossRef]
- Crawford, M.A.; Schmidt, W.F.; Broadhurst, C.L.; Wang, Y. Lipids in the origin of intracellular detail and speciation in the Cambrian epoch and the significance of the last double bond of docosahexaenoic acid in cell signaling. Prostaglandins Leukot Essent Fat. Acids 2021, 166, 102230. [Google Scholar] [CrossRef]
- Bourre, J.-M.; Francois, M.; Youyou, A.; Dumont, O.; Piciotti, M.; Pascal, G.; Durand, G. The Effects of Dietary α-Linolenic Acid on the Composition of Nerve Membranes, Enzymatic Activity, Amplitude of Electrophysiological Parameters, Resistance to Poisons and Performance of Learning Tasks in Rats. J. Nutr. 1989, 119, 1880–1892. [Google Scholar] [CrossRef]
- Moriguchi, T.; Loewke, J.; Garrison, M.; Catalan, J.N.; Salem, N., Jr. Reversal of docosahexaenoic acid deficiency in the rat brain, retina, liver, and serum. J. Lipid Res. 2001, 42, 419–427. [Google Scholar] [CrossRef]
- Mitchell, D.C.; Niu, S.L.; Litman, B.J. Quantifying the differential effects of DHA and DPA on the early events in visual signal transduction. Chem. Phys. Lipids 2012, 165, 393–400. [Google Scholar] [CrossRef]
- Nguyen, L.N.; Ma, D.; Shui, G.; Wong, P.; Cazenave-Gassiot, A.; Zhang, X.; Wenk, M.R.; Goh, E.L.K.; Silver, D.L. Mfsd2a is a transporter for the essential omega-3 fatty acid docosahexaenoic acid. Nature 2014, 509, 503–506. [Google Scholar] [CrossRef] [PubMed]
- Gibson, R.A.; Neumann, M.A.; Lien, E.L.; Boyd, K.A.; Tu, W.C. Docosahexaenoic acid synthesis from alpha-linolenic acid is inhibited by diets high in polyunsaturated fatty acids. Prostaglandins Leukot Essent Fat. Acids 2013, 88, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Brainard, J.S.; Jimoh, O.F.; Deane, K.H.O.; Biswas, P.; Donaldson, D.; Maas, K.; Abdelhamid, A.S.; Hooper, L.; PUFAH group. Omega-3, Omega-6, and Polyunsaturated Fat for Cognition: Systematic Review and Meta-analysis of Randomized Trials. J. Am. Med. Dir. Assoc. 2020, 21, 1439–1450.e21. [Google Scholar] [CrossRef] [PubMed]
- Gould, J.F.; Makrides, M.; Gibson, R.A.; Sullivan, T.R.; McPhee, A.J.; Anderson, P.J.; Best, K.P.; Sharp, M.; Cheong, J.L.Y.; Opie, G.F.; et al. Neonatal Docosahexaenoic Acid in Preterm Infants and Intelligence at 5 Years. N. Engl. J. Med. 2022, 387, 1579–1588. [Google Scholar] [CrossRef]
- Appleton, K.M.; Voyias, P.D.; Sallis, H.M.; Dawson, S.; Ness, A.R.; Churchill, R.; Perry, R. Omega-3 fatty acids for depression in adults. Cochrane Database Syst. Rev. 2021, 11, CD004692. [Google Scholar] [CrossRef]
- Farquharson, J.; Jamieson, E.C.; Abbasi, K.A.; Patrick, W.J.; Logan, R.W.; Cockburn, F. Effect of diet on the fatty acid composition of the major phospholipids of infant cerebral cortex. Arch. Dis. Child. 1995, 72, 198–203. [Google Scholar] [CrossRef]
- Farquharson, J.; Cockburn, F.; Patrick, W.A.; Jamieson, E.C.; Logan, R.W. Effect of diet on infant subcutaneous tissue triglyceride fatty acids. Arch. Dis. Child. 1993, 69, 589–593. [Google Scholar] [CrossRef][Green Version]
- Makrides, M.; Neumann, M.A.; Byard, R.W.; Simmer, K.; Gibson, R.A. Fatty acid composition of brain, retina, and erythrocytes in breast- and formula-fed infants. Am. J. Clin. Nutr. 1994, 60, 189–194. [Google Scholar] [CrossRef]
- Bush, R.A.; Remé, C.E.; Malnoë, A. Light damage in the rat retina: The effect of dietary deprivation of N-3 fatty acids on acute structural alterations. Exp. Eye Res. 1991, 53, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Diau, G.Y.; Hsieh, A.T.; Sarkadi-Nagy, E.A.; Wijendran, V.; Nathanielsz, P.W.; Brenna, J.T. The influence of long chain polyunsaturate supplementation on docosahexaenoic acid and arachidonic acid in baboon neonate central nervous system. BMC Med. 2005, 3, 11. [Google Scholar] [CrossRef]
- Niu, S.L.; Mitchell, D.C.; Lim, S.Y.; Wen, Z.M.; Kim, H.Y.; Salem NJr Litman, B.J. Reduced G protein-coupled signaling efficiency in retinal rod outer segments in response to n-3 fatty acid deficiency. J. Biol. Chem. 2004, 279, 31098–31104. [Google Scholar] [CrossRef] [PubMed]
- Weisinger, H.S.; Vingrys, A.J.; Sinclair, A.J. Effect of dietary n-3 deficiency on the electroretinogram in the guinea pig. Ann. Nutr Metab. 1996, 40, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Weisinger, H.S.; Armitage, J.A.; Jeffrey, B.G.; Mitchell, D.C.; Moriguchi, T.; Sinclair, A.J.; Weisinger, R.S.; Salem, N., Jr. Retinal sensitivity loss in third-generation n-3 PUFA-deficient rats. Lipids 2002, 37, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, I.; Hussein, N.; Baumann, M.H.; Di Martino, C.; Salem, N., Jr. An n-3 fatty acid deficiency impairs rat spatial learning in the Barnes maze. Behav. Neurosci. 2009, 123, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Francès, H.; Monier, C.; Bourre, J.M. Effects of dietary alpha-linolenic acid deficiency on neuromuscular and cognitive functions in mice. Life Sci 1995, 57, 1935–1947. [Google Scholar] [CrossRef]
- Lamptey, M.S.; Walker, B.L. A possible essential role for dietary linolenic acid in the development of the young rat. J. Nutr. 1976, 106, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.Y.; Hoshiba, J.; Moriguchi, T.; Salem, N., Jr. N-3 fatty acid deficiency induced by a modified artificial rearing method leads to poorer performance in spatial learning tasks. Pediatr. Res. 2005, 58, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Moriguchi, T.; Saleml, N., Jr. Recovery of brain docosahexaenoate leads to recovery of spatial task performance. J. Neurochem. 2003, 87, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, R.S.; Luxwolda, M.F.; Offringa, P.J.; Boersma, E.R.; Dijck-Brouwer, D.A.; Muskiet, F.A. Gestational age dependent changes of the fetal brain, liver and adipose tissue fatty acid compositions in a population with high fish intakes. Prostaglandins Leukot Essent Fat. Acids. 2012, 86, 189–199. [Google Scholar] [CrossRef]
- Kuipers, R.S.; Luxwolda, M.F.; Dijck-Brouwer, D.A.; Eaton, S.B.; Crawford, M.A.; Cordain, L.; Muskiet, F.A. Estimated macronutrient and fatty acid intakes from an East African Paleolithic diet. Br. J. Nutr. 2010, 104, 1666–1687. [Google Scholar] [CrossRef]
- Mann, N.; Pirotta, Y.; O’Connell, S.; Li, D.; Kelly, F.; Sinclair, A. Fatty acid composition of habitual omnivore and vegetarian diets. Lipids 2006, 41, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.M.; Sanders, T.A.; Kendall, A.C.; Nicolaou, A.; Gray, R.; Al-Khatib, H.; Hall, W.L. A comparison of heart rate variability, n-3 PUFA status and lipid mediator profile in age- and BMI-matched middle-aged vegans and omnivores. Br. J. Nutr. 2017, 117, 669–685. [Google Scholar] [CrossRef] [PubMed]
- Agren, J.J.; Törmälä, M.L.; Nenonen, M.T.; Hänninen, O.O. Fatty acid composition of erythrocyte, platelet, and serum lipids in strict vegans. Lipids 1995, 30, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Chamorro, R.; Gonzalez, M.F.; Aliaga, R.; Gengler, V.; Balladares, C.; Barrera, C.; Bascuñan, K.A.; Bazinet, R.P.; Valenzuela, R. Diet, Plasma, Erythrocytes, and Spermatozoa Fatty Acid Composition Changes in Young Vegan Men. Lipids 2020, 55, 639–648. [Google Scholar] [CrossRef]
- Burns-Whitmore, B.; Froyen, E.; Heskey, C.; Parker, T.; San Pablo, G. Alpha-Linolenic and Linoleic Fatty Acids in the Vegan Diet: Do They Require Dietary Reference Intake/Adequate Intake Special Consideration? Nutrients 2019, 11, 2365. [Google Scholar] [CrossRef]
- Harika, R.K.; Eilander, A.; Alssema, M.; Osendarp, S.J.; Zock, P.L. Intake of fatty acids in general populations worldwide does not meet dietary recommendations to prevent coronary heart disease: A systematic review of data from 40 countries. Ann. Nutr Metab 2013, 63, 229–238. [Google Scholar] [CrossRef]
- Neufingerl, N.; Eilander, A. Nutrient Intake and Status in Adults Consuming Plant-Based Diets Compared to Meat-Eaters: A Systematic Review. Nutrients 2021, 14, 29. [Google Scholar] [CrossRef]
- Miyake, Y.; Tanaka, K.; Okubo, H.; Sasaki, S.; Arakawa, M. Fish and fat intake and prevalence of depressive symptoms during pregnancy in Japan: Baseline data from the Kyushu Okinawa Maternal and Child Health Study. J. Psychiatr. Res. 2013, 47, 572–578. [Google Scholar] [CrossRef]
- Lakshmipriya, N.; Gayathri, R.; Praseena, K.; Vijayalakshmi, P.; Geetha, G.; Sudha, V.; Krishnaswamy, K.; Anjana, R.M.; Henry, J.; Mohan, V. Type of vegetable oils used in cooking and risk of metabolic syndrome among Asian Indians. Int. J. Food Sci. Nutr. 2013, 64, 131–139. [Google Scholar] [CrossRef]
- Hibbeln, J.R.; Nieminen, L.R.; Blasbalg, T.L.; Riggs, J.A.; Lands, W.E. Healthy intakes of n-3 and n-6 fatty acids: Estimations considering worldwide diversity. Am. J. Clin. Nutr. 2006, 83 (Suppl. S6), 1483S–1493S. [Google Scholar] [CrossRef] [PubMed]
- Sanders, T.A.; Ellis, F.R.; Dickerson, J.W. Studies of vegans: The fatty acid composition of plasma choline phosphoglycerides, erythrocytes, adipose tissue, and breast milk, and some indicators of susceptibility to ischemic heart disease in vegans and omnivore controls. Am. J. Clin. Nutr. 1978, 31, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Parasannanavar, D.; Gaddam, I.; Bukya, T.; Ibrahim, S.A.; Reddy, K.S.; Banjara, S.K.; Salvadi, B.P.P.; Kumar, B.N.; Rao, S.F.; Geddam, J.J.B.; et al. Omega-3 polyunsaturated fatty acid intake and plasma fatty acids of school going Indian children-a cross-sectional study. Prostaglandins Leukot Essent Fat. Acids. 2021, 170, 102294. [Google Scholar] [CrossRef] [PubMed]
- Sanders, T.A.; Reddy, S. The influence of a vegetarian diet on the fatty acid composition of human milk and the essential fatty acid status of the infant. J. Pediatr. 1992, 120 Pt 2, S71–S77. [Google Scholar] [CrossRef] [PubMed]
- Mani, I.; Dwarkanath, P.; Thomas, T.; Thomas, A.; Kurpad, A.V. Maternal fat and fatty acid intake and birth outcomes in a South Indian population. Int. J. Epidemiol. 2016, 45, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Sinclair, A.J. Increased alpha-linolenic acid intake increases tissue alpha-linolenic acid content and apparent oxidation with little effect on tissue docosahexaenoic acid in the guinea pig. Lipids 2000, 35, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, R.S.; Luxwolda, M.F.; Offringa, P.J.; Boersma, E.R.; Dijck-Brouwer, D.A.; Muskiet, F.A. Fetal intrauterine whole body linoleic, arachidonic and docosahexaenoic acid contents and accretion rates. Prostaglandins Leukot Essent Fat. Acids. 2012, 86, 13–20. [Google Scholar] [CrossRef]
- Hodson, L.; Skeaff, C.M.; Fielding, B.A. Fatty acid composition of adipose tissue and blood in humans and its use as a biomarker of dietary intake. Prog Lipid Res. 2008, 47, 348–380. [Google Scholar] [CrossRef]
- Miles, F.L.; Lloren, J.I.C.; Haddad, E.; Jaceldo-Siegl, K.; Knutsen, S.; Sabate, J.; Fraser, G.E. Plasma, Urine, and Adipose Tissue Biomarkers of Dietary Intake Differ Between Vegetarian and Non-Vegetarian Diet Groups in the Adventist Health Study-2. J. Nutr. 2019, 149, 667–675. [Google Scholar] [CrossRef]
- Halliwell, K.J.; Fielding, B.A.; Samra, J.S.; Humphreys, S.M.; Frayn, K.N. Release of individual fatty acids from human adipose tissue in vivo after an overnight fast. J. Lipid Res. 1996, 37, 1842–1848. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Liu, X.; Zhou, B.; Jiang, A.C.; Chai, L. An updated review of worldwide levels of docosahexaenoic and arachidonic acid in human breast milk by region. Public Health Nutr. 2016, 19, 2675–2687. [Google Scholar] [CrossRef]
- Sanders, T.A. Essential fatty acid requirements of vegetarians in pregnancy, lactation, and infancy. Am. J. Clin. Nutr. 1999, 70 (Suppl. S3), 555S–559S. [Google Scholar] [CrossRef] [PubMed]
- Perrin, M.T.; Pawlak, R.; Dean, L.L.; Christis, A.; Friend, L. A cross-sectional study of fatty acids and brain-derived neurotrophic factor (BDNF) in human milk from lactating women following vegan, vegetarian, and omnivore diets. Eur. J. Nutr. 2019, 58, 2401–2410. [Google Scholar] [CrossRef] [PubMed]
- Nyuar, K.B.; Min, Y.; Ghebremeskel, K.; Khalil, A.K.; Elbashir, M.I.; Cawford, M.A. Milk of northern Sudanese mothers whose traditional diet is high in carbohydrate contains low docosahexaenoic acid. Acta Paediatr. 2010, 99, 1824–1827. [Google Scholar] [CrossRef] [PubMed]
- Nyuar, K.B.; Min, Y.; Dawood, M.; Abukashawa, S.; Daak, A.; Ghebremeskel, K. Regular consumption of Nile river fish could ameliorate the low milk DHA of Southern Sudanese women living in Khartoum City area. Prostaglandins Leukot Essent Fat. Acids. 2013, 89, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Adam, I.; Babiker, S.; Mohmmed, A.A.; Salih, M.M.; Prins, M.H.; Zaki, Z.M. Low body mass index, anaemia and poor perinatal outcome in a rural hospital in eastern Sudan. J. Trop. Pediatr. 2008, 54, 202–204. [Google Scholar] [CrossRef]
- Carver, J.D.; Benford, V.J.; Han, B.; Cantor, A.B. The relationship between age and the fatty acid composition of cerebral cortex and erythrocytes in human subjects. Brain Res. Bull. 2001, 56, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.; Mougan, I. Fatty acid composition of human brain phospholipids during normal development. J. Neurochem 1998, 71, 2525–2533. [Google Scholar] [CrossRef] [PubMed]
- Fraser, T.; Tayler, H.; Love, S. Low-temperature improved-throughput method for analysis of brain fatty acids and assessment of their post-mortem stability. J. Neurosci. Methods. 2008, 169, 135–140. [Google Scholar] [CrossRef]
- McNamara, R.K.; Jandacek, R. Investigation of postmortem brain polyunsaturated fatty acid composition in psychiatric disorders: Limitations, challenges, and future directions. J. Psychiatr. Res. 2011, 45, 44–46. [Google Scholar] [CrossRef][Green Version]
- Martinez, M.; Ballabriga, A.; Gil-Gibernau, J.J. Lipids of the developing human retina: I. Total fatty acids, plasmalogens, and fatty acid composition of ethanolamine and choline phosphoglycerides. J. Neurosci. Res. 1988, 20, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.; Conde, C.; Ballabriga, A. Some chemical aspects of human brain development. II. Phosphoglyceride Fat. Acids. Pediatr. Res. 1974, 8, 93–102. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Svennerholm, L. Distribution and fatty acid composition of phosphoglycerides in normal human brain. J. Lipid Res. 1968, 9, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Söderberg, M.; Edlund, C.; Kristensson, K.; Dallner, G. Fatty acid composition of brain phospholipids in aging and in Alzheimer's disease. Lipids 1991, 26, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Fabelo, N.; Martín, V.; Santpere, G.; Marín, R.; Torrent, L.; Ferrer, I.; Díaz, M. Severe alterations in lipid composition of frontal cortex lipid rafts from Parkinson’s disease and incidental Parkinson’s disease. Mol. Med. 2011, 17, 1107–1118. [Google Scholar] [CrossRef] [PubMed]
- Carrié, I.; Clément, M.; de Javel, D.; Francès, H.; Bourre, J.M. Phospholipid supplementation reverses behavioral and biochemical alterations induced by n-3 polyunsaturated fatty acid deficiency in mice. J. Lipid Res. 2000, 41, 473–480. [Google Scholar] [CrossRef]
- Hopiavuori, B.R.; Agbaga, M.P.; Brush, R.S.; Sullivan, M.T.; Sonntag, W.E.; Anderson, R.E. Regional changes in CNS and retinal glycerophospholipid profiles with age: A molecular blueprint. J. Lipid Res. 2017, 58, 668–680. [Google Scholar] [CrossRef]
- Phillips, G.R.; Hancock, S.E.; Jenner, A.M.; McLean, C.; Newell, K.A.; Mitchell, T.W. Phospholipid Profiles Are Selectively Altered in the Putamen and White Frontal Cortex of Huntington’s Disease. Nutrients 2022, 14, 2086. [Google Scholar] [CrossRef]
- Market Analysis Report 2020–2028, Report ID: 978-1-68038-079-8. Available online: https://www.grandviewresearch.com/industry-analysis/omega-3-market (accessed on 26 December 2022).
- Business Standard, 14 October 2022. Available online: https://www.business-standard.com/article/economy-policy/poverty-ratio-32-75-in-rural-areas-against-8-81-in-urban-niti-report-121120500971_1.html (accessed on 26 December 2022).
- UNHCR 2022. Available online: https://www.unrefugees.org/refugee-facts/statistics/ (accessed on 26 December 2022).
- Lacombe, R.J.S.; Lee, C.C.; Bazinet, R.P. Turnover of brain DHA in mice is accurately determined by tracer-free natural abundance carbon isotope ratio analysis. J. Lipid Res. 2020, 61, 116–126. [Google Scholar] [CrossRef]
- Weisinger, H.S.; Vingrys, A.J.; Bui, B.V.; Sinclair, A.J. Effects of dietary n-3 fatty acid deficiency and repletion in the guinea pig retina. Investig. Ophthalmol. Vis. Sci. 1999, 40, 327–338. [Google Scholar]
- Horrobin, D.F.; Manku, M.S.; Hillman, H.; Iain, A.; Glen, M. Fatty acid levels in the brains of schizophrenics and normal controls. Biol. Psychiatry 1991, 30, 795–805. [Google Scholar] [CrossRef]
- Skinner, E.R.; Watt, C.; Besson, J.A.; Best, P.V. Differences in the fatty acid composition of the grey and white matter of different regions of the brains of patients with Alzheimer’s disease and control subjects. Brain 1993, 116 Pt 3, 717–725. [Google Scholar] [CrossRef]
- Yao, J.K.; Leonard, S.; Reddy, R.D. Membrane phospholipid abnormalities in postmortem brains from schizophrenic patients. Schizophr. Res. 2000, 42, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Landén, M.; Davidsson, P.; Gottfries, C.G.; Månsson, J.E.; Blennow, K. Reduction of the synaptophysin level but normal levels of glycerophospholipids in the gyrus cinguli in schizophrenia. Schizophr. Res. 2002, 55, 83–88. [Google Scholar] [CrossRef]
- Lalovic, A.; Levy, E.; Canetti, L.; Sequeira, A.; Montoudis, A.; Turecki, G. Fatty acid composition in postmortem brains of people who completed suicide. J. Psychiatry Neurosci. 2007, 32, 363–370. [Google Scholar] [PubMed]
- Igarashi, M.; Ma, K.; Gao, F.; Kim, H.W.; Greenstein, D.; Rapoport, S.I.; Rao, J.S. Brain lipid concentrations in bipolar disorder. J. Psychiatr. Res. 2010, 44, 177–182. [Google Scholar] [CrossRef] [PubMed]
- McNamara, R.K.; Jandacek, R.; Rider, T.; Tso, P.; Dwivedi, Y.; Roberts, R.C.; Conley, R.R.; Pandey, G.N. Fatty acid composition of the postmortem prefrontal cortex of adolescent male and female suicide victims. Prostaglandins Leukot Essent Fat. Acids 2009, 80, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Conklin, S.M.; Runyan, C.A.; Leonard, S.; Reddy, R.D.; Muldoon, M.F.; Yao, J.K. Age-related changes of n-3 and n-6 polyunsaturated fatty acids in the anterior cingulate cortex of individuals with major depressive disorder. Prostaglandins Leukot Essent Fat. Acids 2010, 82, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Fraser, T.; Tayler, H.; Love, S. Fatty acid composition of frontal, temporal and parietal neocortex in the normal human brain and in Alzheimer’s disease. Neurochem. Res. 2010, 35, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Hamazaki, K.; Hamazaki, T.; Inadera, H. Fatty acid composition in the postmortem amygdala of patients with schizophrenia, bipolar disorder, and major depressive disorder. J. Psychiatr. Res. 2012, 46, 1024–1028. [Google Scholar] [CrossRef]
- Taha, A.Y.; Cheon, Y.; Ma, K.; Rapoport, S.I.; Rao, J.S. Altered fatty acid concentrations in prefrontal cortex of schizophrenic patients. J. Psychiatr. Res. 2013, 47, 636–643. [Google Scholar] [CrossRef]
- Hamazaki, K.; Hamazaki, T.; Inadera, H. Abnormalities in the fatty acid composition of the postmortem entorhinal cortex of patients with schizophrenia, bipolar disorder, and major depressive disorder. Psychiatry Res. 2013, 210, 346–350. [Google Scholar] [CrossRef] [PubMed]
- McNamara, R.K.; Rider, T.; Jandacek, R.; Tso, P. Abnormal fatty acid pattern in the superior temporal gyrus distinguishes bipolar disorder from major depression and schizophrenia and resembles multiple sclerosis. Psychiatry Res. 2014, 215, 560–567. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hamazaki, K.; Maekawa, M.; Toyota, T.; Dean, B.; Hamazaki, T.; Yoshikawa, T. Fatty acid composition of the postmortem prefrontal cortex of patients with schizophrenia, bipolar disorder, and major depressive disorder. Psychiatry Res. 2015, 227, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Hamazaki, K.; Maekawa, M.; Toyota, T.; Iwayama, Y.; Dean, B.; Hamazaki, T.; Yoshikawa, T. Fatty acid composition and fatty acid binding protein expression in the postmortem frontal cortex of patients with schizophrenia: A case-control study. Schizophr. Res. 2016, 171, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Hamazaki, K.; Maekawa, M.; Toyota, T.; Dean, B.; Hamazaki, T.; Yoshikawa, T. Fatty acid composition of the postmortem corpus callosum of patients with schizophrenia, bipolar disorder, or major depressive disorder. Eur. Psychiatry 2017, 39, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Dyer, R.A.; Beasley, C.L. Evidence for altered cell membrane lipid composition in postmortem prefrontal white matter in bipolar disorder and schizophrenia. J. Psychiatr. Res. 2017, 95, 135–142. [Google Scholar] [CrossRef]
- Beasley, C.L.; Honer, W.G.; Ramos-Miguel, A.; Vila-Rodriguez, F.; Barr, A.M. Prefrontal fatty acid composition in schizophrenia and bipolar disorder: Association with reelin expression. Schizophr. Res. 2020, 215, 493–498. [Google Scholar] [CrossRef]



| Condition | Description | |
|---|---|---|
| 1 | Dietary deficiency of ALA | Dietary insufficiency of ALA which would be expected to result in decreased DHA synthesis in the liver and increased 22:5n-6 synthesis from LA, and hence less DHA and more 22:5n-6 transported to brain via plasma lipoproteins. |
| 2 | Brain DHA uptake receptors | Polymorphisms in brain fatty acid transport proteins such as fatty acid binding proteins (FABP), fatty acid transport proteins (FATP), fatty acid translocase FAT/CD36, DHA uptake receptors Mfsd2a, and APOE4, and hence less uptake of DHA into the brain from the circulation [7,8]. |
| 3 | Brain DHA metabolism | Acyl CoA synthetase 6 controls neuronal DHA levels independent of diet by ligating CoA to free DHA, and hence polymorphisms could reduce brain DHA levels [9] |
| 4 | Brain PUFA peroxidation | Increased peroxidation of PUFA in brain, where it would be expected to result in increased indices of lipid peroxidation, resulting peroxidation/loss of DHA in the brain [10]. |
| 5 | Liver ALA metabolism | Polymorphisms in liver FADS1/2 desaturases and elongases [11,12,13] resulting in reduced DHA synthesis from ALA, and hence less DHA transported to brain via plasma lipoproteins. |
| 6 | Liver function | Liver damage, such as fatty liver induced by diet, alcohol, or liver diseases, which might impact on the capacity of the liver to synthesise DHA from ALA, and hence less DHA transported to brain via plasma lipoproteins [14,15]. |
| Fatty Acid % | Vegan Plasma PC 1 | Omni Plasma PC 1 | Vegan RBC 2 | Omni RBC 2 | Vegan RBC PS 3 | Omni RBC PS 3 | Vegan Plasma Total 4 | Omni Plasma Total 4 | School Age (M) Plasma PL 5 | School Age (F) Plasma PL 5 |
|---|---|---|---|---|---|---|---|---|---|---|
| 18:2n-6 | 33.3 | 26.0 | 14.2 | 11.6 | 3.3 | 2.2 | 33.1 | 27.1 | 27.2 | 27.5 |
| 20:4n-6 | 10.6 | 9.1 | 14.3 | 13.7 | 28.5 | 23.1 | 6.6 | 6.7 | 10.0 | 9.3 |
| 22:4n-6 | 0.6 | 0.3 | 3.0 | 2.0 | 3.6 | 2.1 | 0.2 | 0.2 | 2.7 | 2.9 |
| 22:5n-6 | 0.4 | 0.2 | 0.5 | 0.2 | 1.2 | 0.6 | 0.2 | 0.3 | 0.6 | 0.6 |
| 18:3n-3 | nr | nr | 0.1 | 0.1 | 0.0 | 0.0 | 0.7 | 0.5 | nr | nr |
| 20:5n-3 | 0.2 | 1.3 | 0.5 | 1.4 | 0.4 | 0.9 | 0.5 | 1.0 | nr | nr |
| 22:5n-3 | 0.8 | 1.1 | 2.1 | 2.5 | 3.4 | 4.0 | 0.5 | 0.6 | 0.2 | 0.2 |
| 22:6n-3 | 1.4 | 4.0 | 3.3 | 6.7 | 7.8 | 14.1 | 0.9 | 2.2 | 1.0 | 1.0 |
| 22:5n-6/DHA | 0.29 | 0.05 | 0.15 | 0.03 | 0.15 | 0.04 | 0.26 | 0.12 | 0.6 | 0.6 |
| Fatty Acid | Cord Plasma Vegetarian | Cord Plasma Omnivore | Cord Artery Vegetarian | Cord Artery Omnivore |
|---|---|---|---|---|
| 22:5n-6 | 2.34 ± 0.16 a,1 | 1.58 ± 0.13 b | 4.15 ± 0.16 a | 3.19 ± 0.15 b |
| 22:6n-3 (DHA) | 4.00 ± 0.36 a | 5.84 ± 0.31 b | 4.05 ± 0.17 a | 5.75 ± 0.19 b |
| 22:5n-6/DHA | 0.59 | 0.27 | 1.02 | 0.55 |
| Population | LA | AA | ALA | EPA | DPA n-3 | DHA | LA/ALA |
|---|---|---|---|---|---|---|---|
| Vegan n ≥ 67 | 23.3 a,2 | 0.39 a | 1.6 a | 0.02 | 0.21 a | 0.12 a | 14.6 a |
| LO-veg n ≥ 218 | 21.7 a | 0.44 a | 1.2 a | 0.02 | 0.20 a | 0.12 a | 18.1 b |
| Non-veg n ≥ 374 | 19.1 b | 0.51 b | 1.0 b | 0.03 | 0.24 b | 0.18 b | 19.1 b |
| Population | LA | AA | ALA | EPA | DHA | LA/ALA | Total n-3 |
|---|---|---|---|---|---|---|---|
| Vegans a | 23.8 | 0.32 | 1.36 | nr | 0.14 | 17.5 | 1.50 |
| Vegetarians a | 19.5 | 0.38 | 1.25 | nr | 0.30 | 15.6 | 1.55 |
| Omnivores a | 10.9 | 0.35 | 0.49 | nr | 0.37 | 22.2 | 0.86 |
| South Sudan b | 14.7 | 0.60 | 0.28 | 0.04 | 0.10 | 52.5 | 0.48 |
| North Sudan c | 12.8 | 0.48 | 0.20 | 0.05 | 0.06 | 64.0 | 0.36 |
| Vulnerable Lifespan Periods | Vulnerable Diets | Vulnerable Nutrient Situations | Vulnerable Populations |
|---|---|---|---|
| Multiple pregnancies, Postanal infants, Weanling Infants, Children, Adolescents | High LA vegetable oils, No green vegetables, No eggs, No fish/shellfish, No fresh milk and meat | High LA/ALA, Low ALA, No LC n-3 PUFA | Dryland agriculture, Deserts, Famines, floods, fires, Refugees |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinclair, A.J.; Wang, Y.; Li, D. What Is the Evidence for Dietary-Induced DHA Deficiency in Human Brains? Nutrients 2023, 15, 161. https://doi.org/10.3390/nu15010161
Sinclair AJ, Wang Y, Li D. What Is the Evidence for Dietary-Induced DHA Deficiency in Human Brains? Nutrients. 2023; 15(1):161. https://doi.org/10.3390/nu15010161
Chicago/Turabian StyleSinclair, Andrew J., Yonghua Wang, and Duo Li. 2023. "What Is the Evidence for Dietary-Induced DHA Deficiency in Human Brains?" Nutrients 15, no. 1: 161. https://doi.org/10.3390/nu15010161
APA StyleSinclair, A. J., Wang, Y., & Li, D. (2023). What Is the Evidence for Dietary-Induced DHA Deficiency in Human Brains? Nutrients, 15(1), 161. https://doi.org/10.3390/nu15010161