
Short-Term Very High Carbohydrate Diet and Gut-Training Have Minor Effects on Gastrointestinal Status and Performance in Highly Trained Endurance Athletes

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
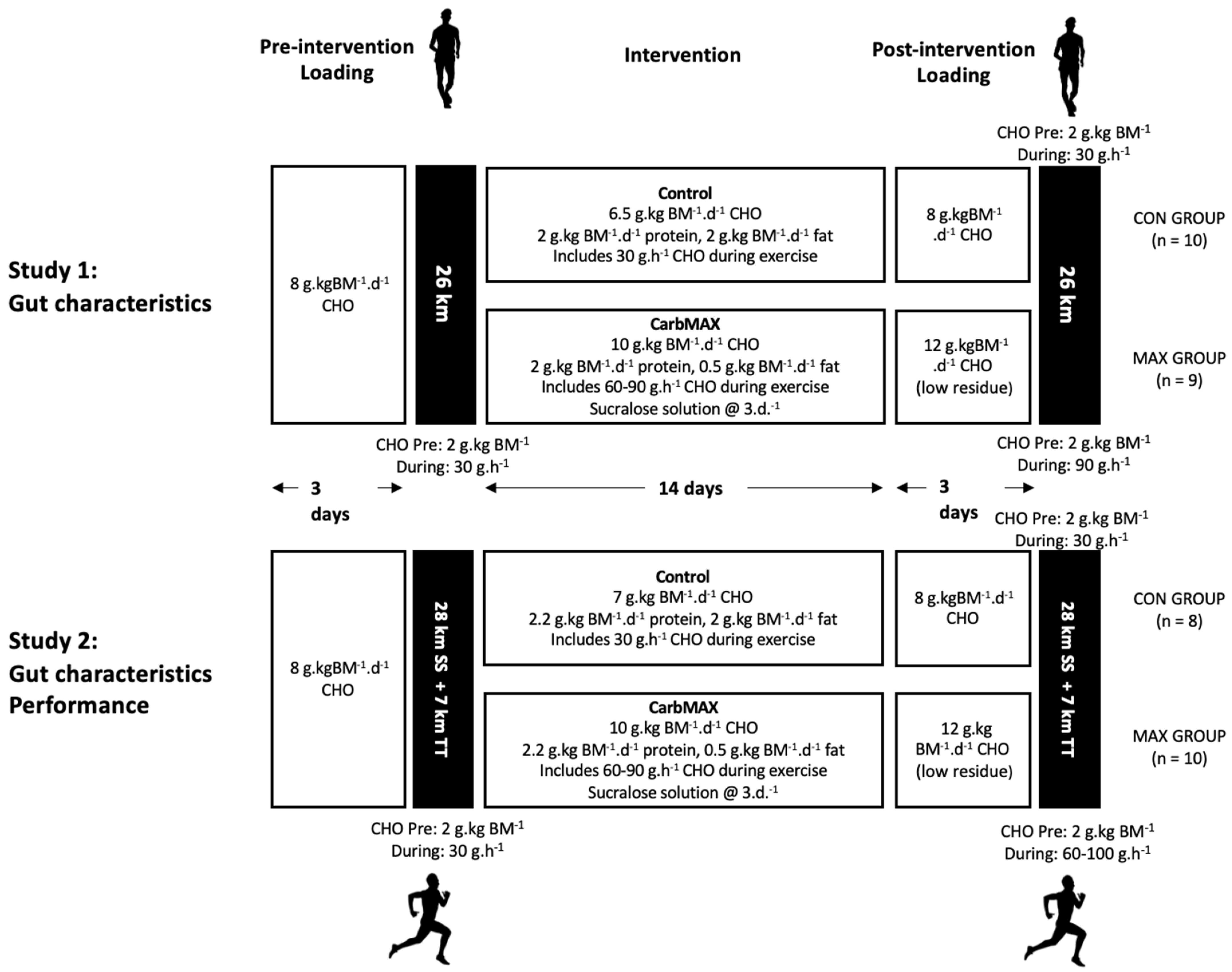
2.1. Study Overview and Participants
2.2. Overview of Dietary Standardisation
2.3. Study 1: Race Walking
2.3.1. Overview of Study
2.3.2. Overview of Diet and Training Intervention
2.3.3. 26 km Race Walking Protocol and Collection of Gastrointestinal Characteristics
2.4. Study 2: Distance Running
2.4.1. Overview of Study
2.4.2. Body Composition
2.4.3. VO2max Testing
2.4.4. Dietary Interventions
2.4.5. Running Performance
2.5. Blood Collection and Analysis
2.6. Gastrointestinal Symptoms (GIS) and Breath H2
2.7. Statistical Analysis
3. Results
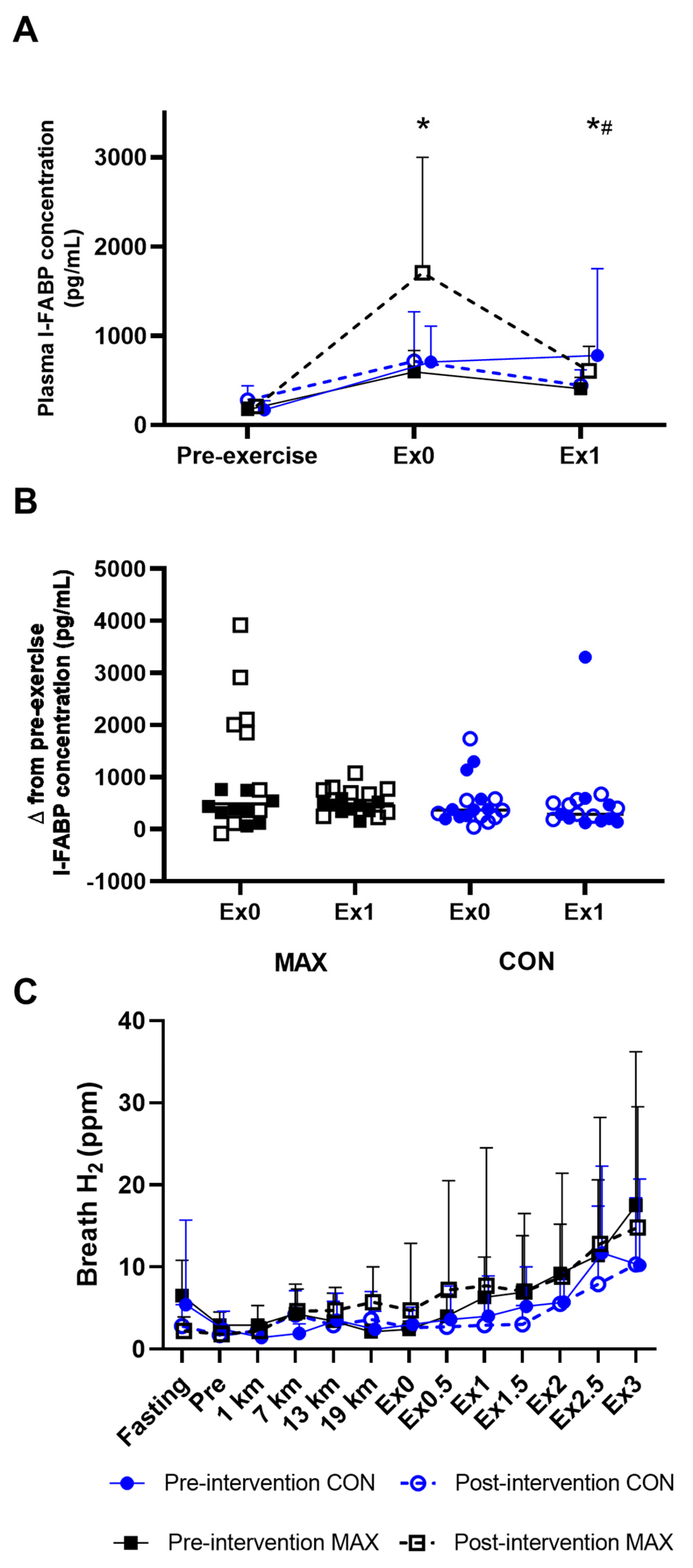
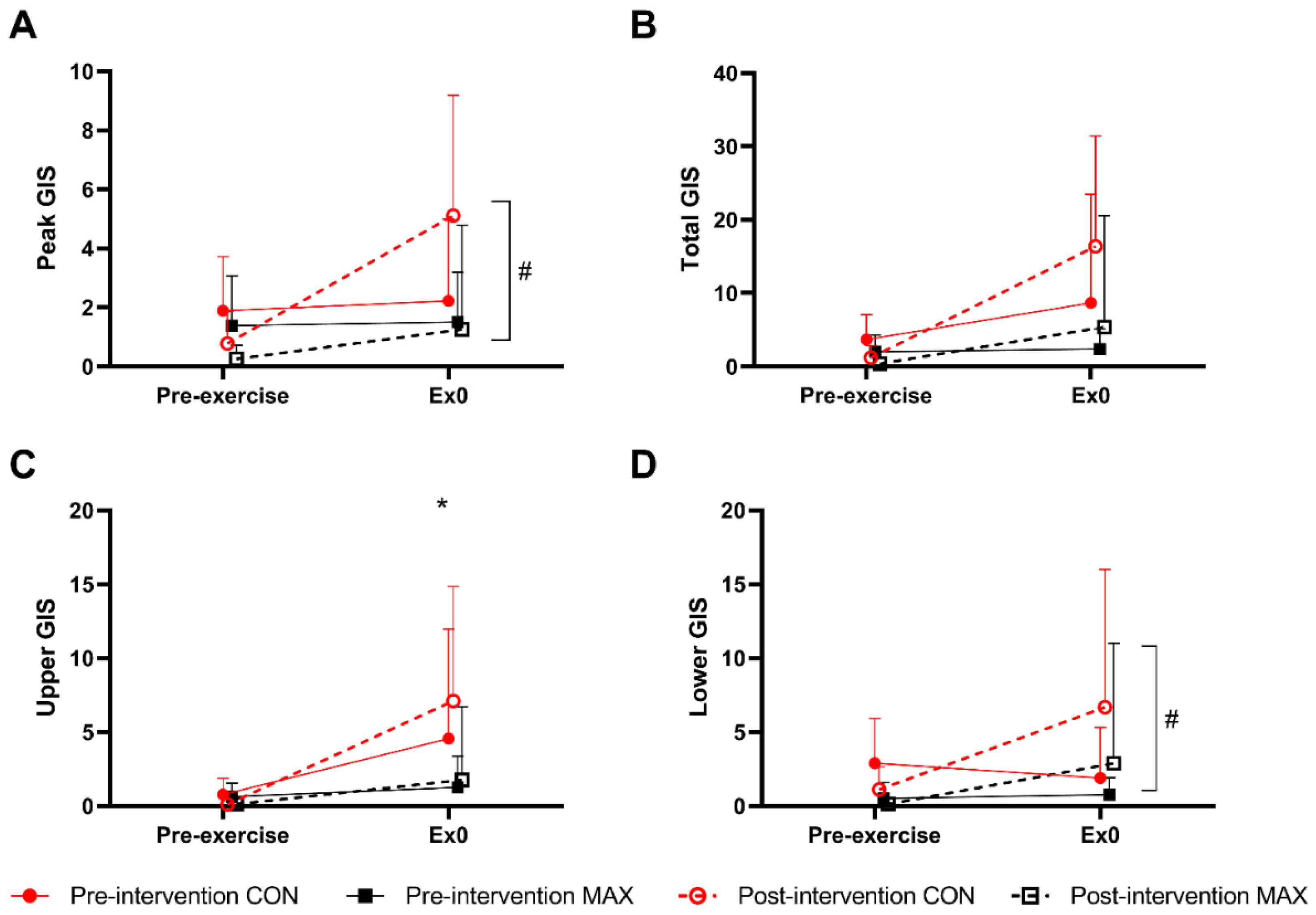
3.1. Study 1—Race Walking
3.1.1. Compliance to Dietary Interventions and Exercise Task
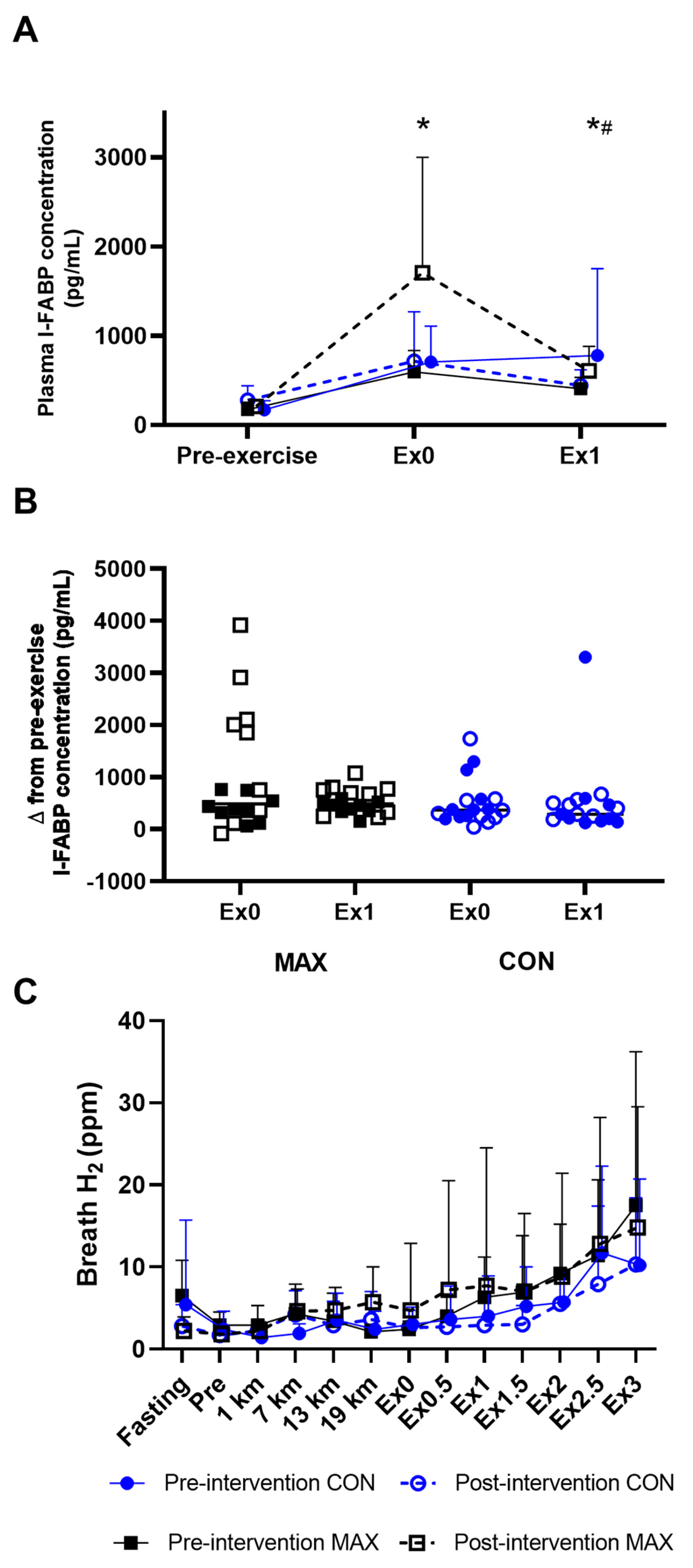
3.1.2. Gastrointestinal Integrity and Function
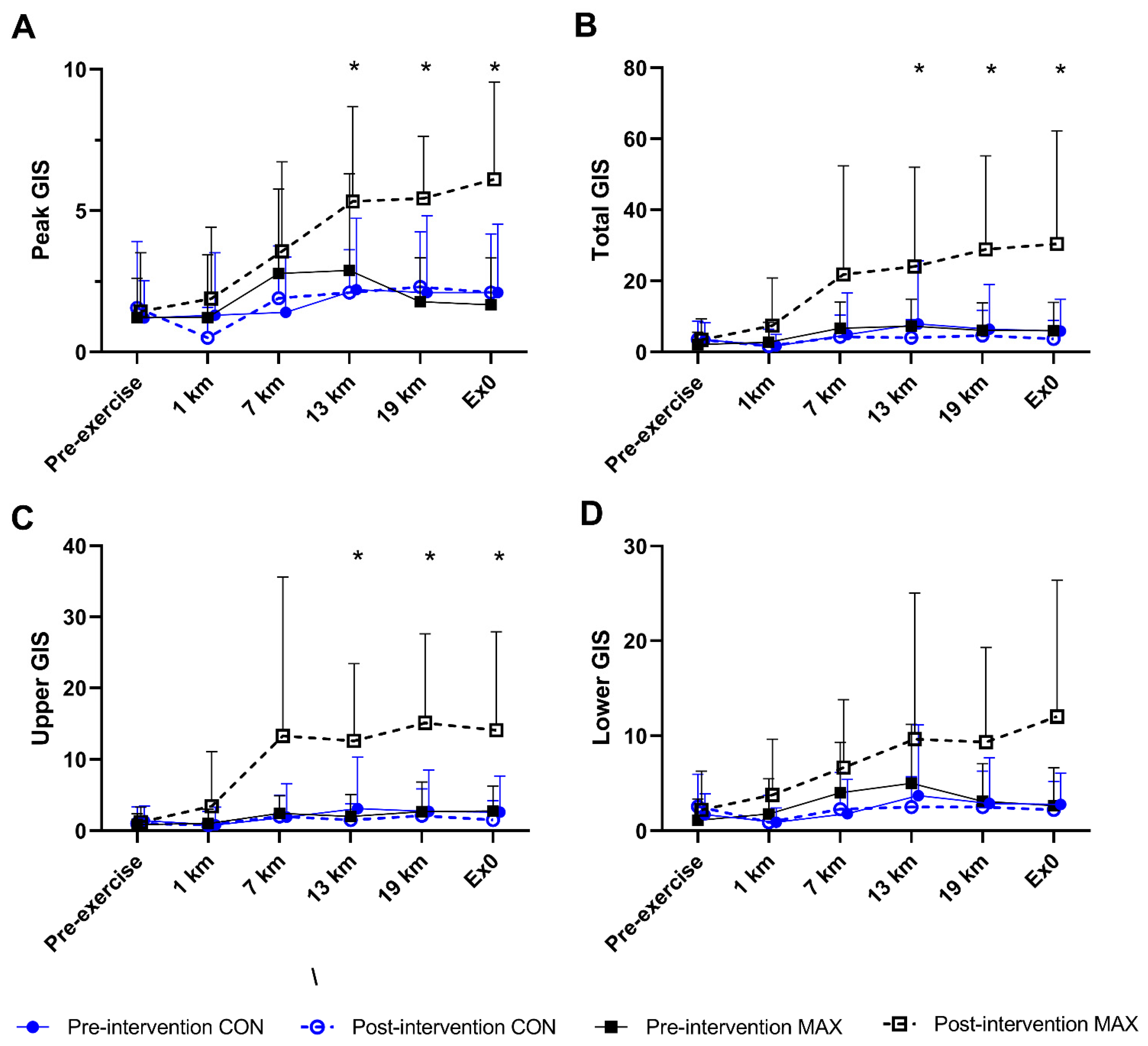
3.1.3. Gastrointestinal Symptoms (GIS)
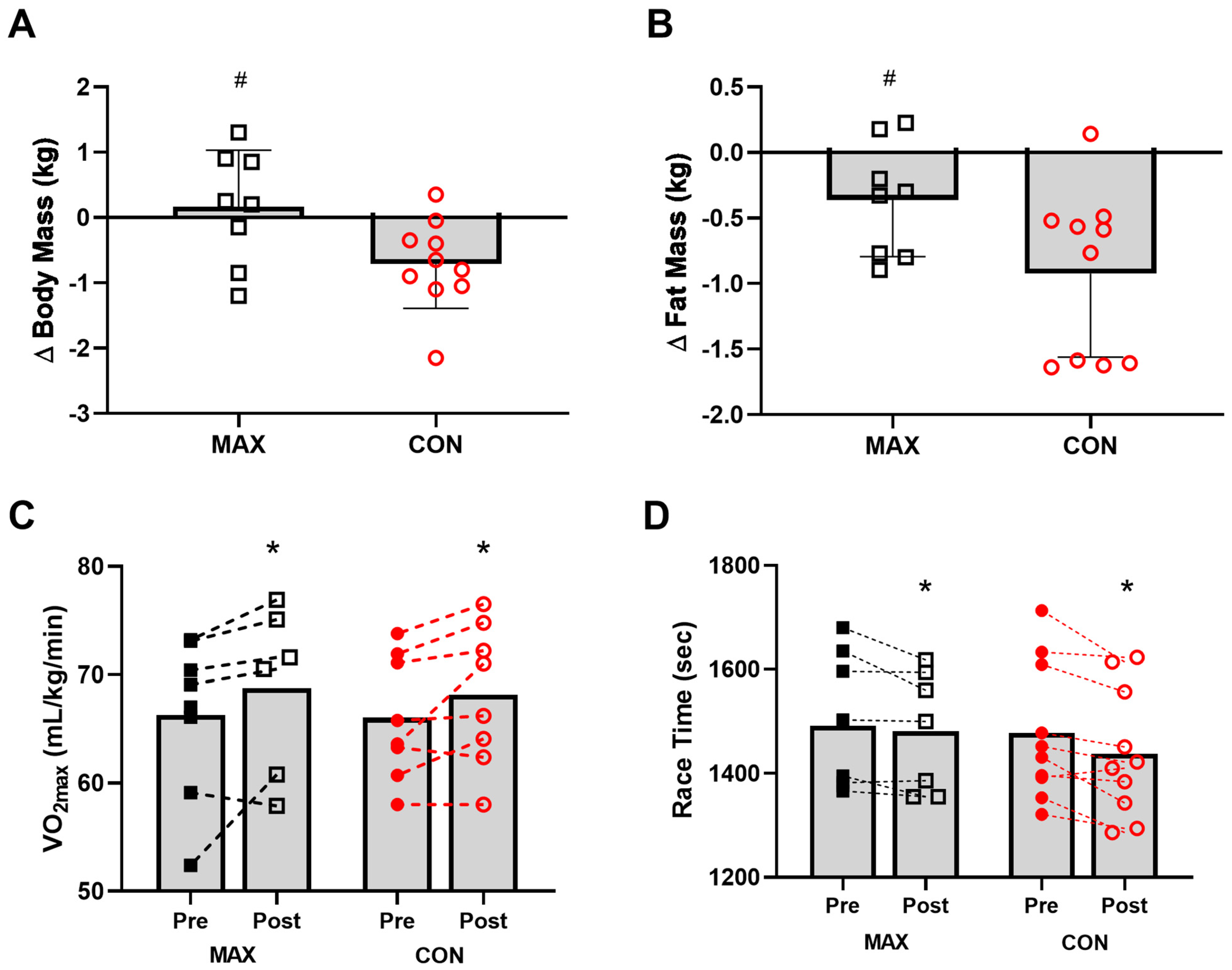
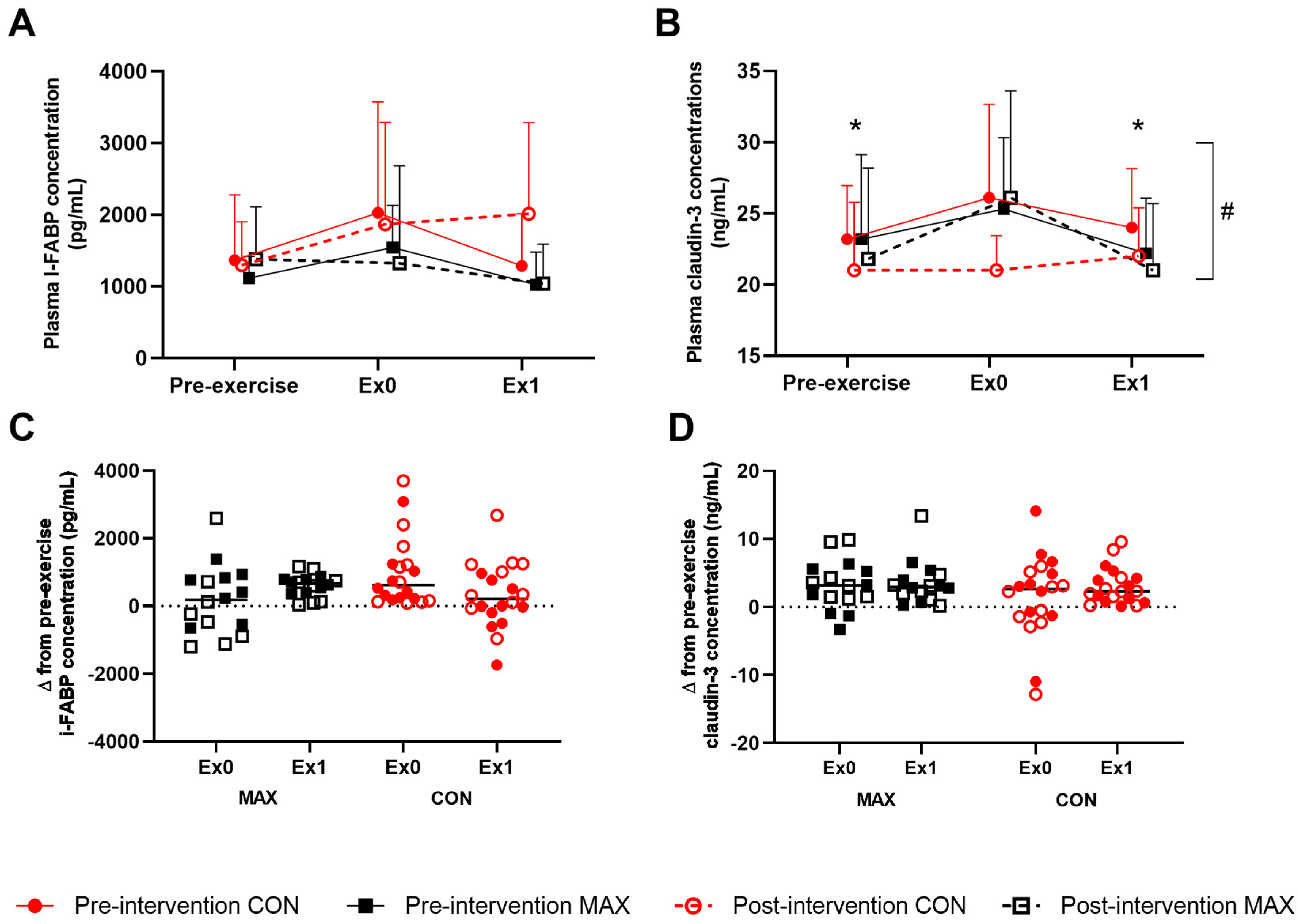
3.2. Study 2—Marathon Running
3.2.1. Dietary Intakes and Compliance with Interventions
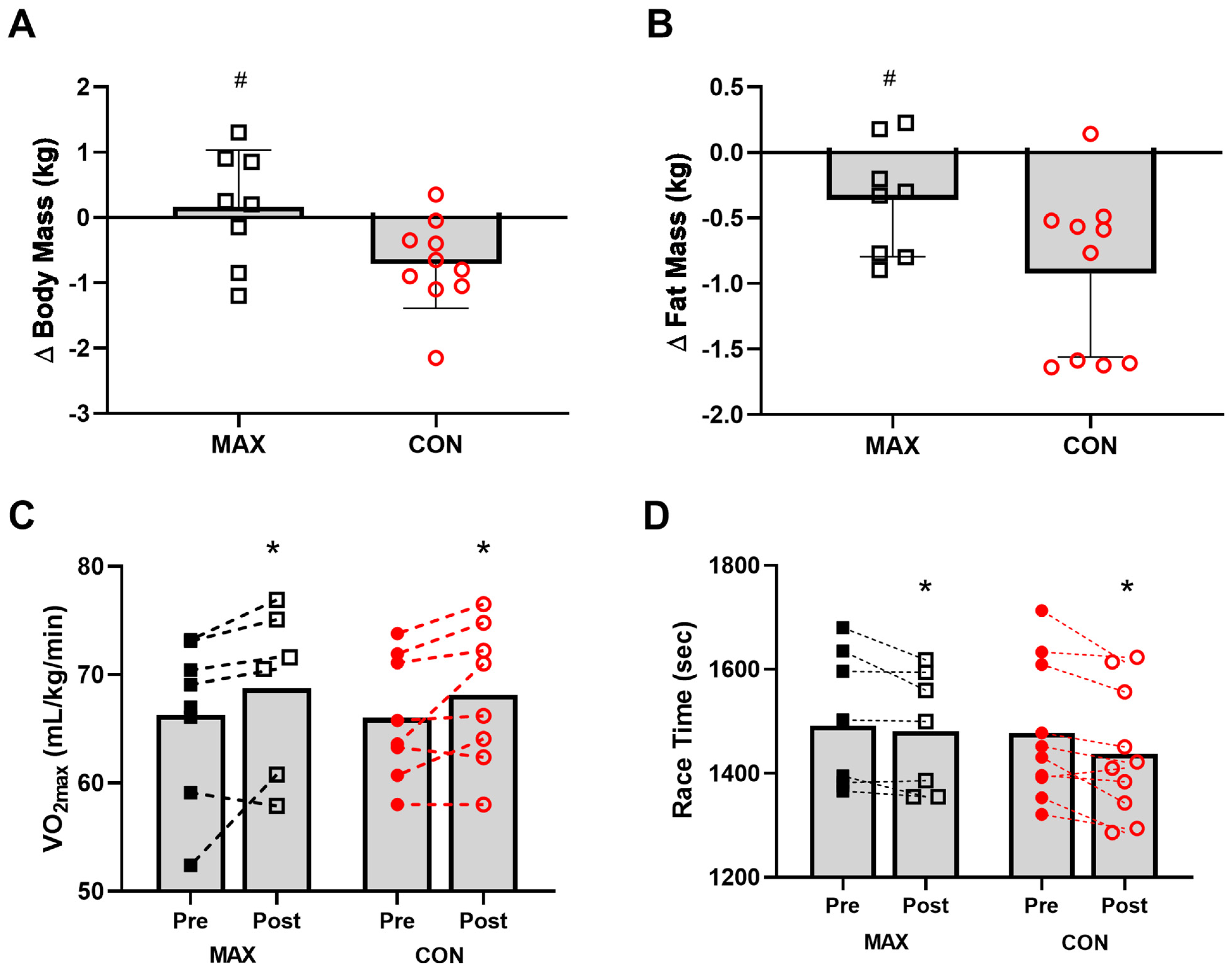
3.2.2. Body Composition
3.2.3. VO2max and Performance
3.2.4. Gastrointestinal Integrity and Function
3.2.5. Gastrointestinal Symptoms (GIS)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, A.M.; Kirby, B.S.; Clark, I.E.; Rice, H.M.; Fulkerson, E.; Wylie, L.J.; Wilkerson, D.P.; Vanhatalo, A.; Wilkins, B.W. Physiological demands of running at 2-hour marathon race pace. J. Appl. Physiol. 2021, 130, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M.; Jeukendrup, A.E.; Jones, A.M.; Mooses, M. Contemporary Nutrition Strategies to Optimize Performance in Distance Runners and Race Walkers. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 117–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, L.; Hawley, J.A. Effects of short-term fat adaptation on metabolism and performance of prolonged exercise. Med. Sci. Sports Exerc. 2002, 34, 1492–1498. [Google Scholar] [CrossRef]
- Volek, J.S.; Freidenreich, D.J.; Saenz, C.; Kunces, L.J.; Creighton, B.C.; Bartley, J.M.; Davitt, P.M.; Munoz, C.X.; Anderson, J.M.; Maresh, C.M.; et al. Metabolic characteristics of keto-adapted ultra-endurance runners. Metabolism 2016, 65, 100–110. [Google Scholar] [CrossRef] [Green Version]
- Burke, L.M.; Ross, M.L.; Garvican-Lewis, L.A.; Welvaert, M.; Heikura, I.A.; Forbes, S.G.; Mirtschin, J.G.; Cato, L.E.; Strobel, N.; Sharma, A.P.; et al. Low carbohydrate, high fat diet impairs exercise economy and negates the performance benefit from intensified training in elite race walkers. J. Physiol. 2017, 595, 2785–2807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, L.M.; Whitfield, J.; Heikura, I.A.; Ross, M.L.R.; Tee, N.; Forbes, S.F.; Hall, R.; McKay, A.K.A.; Wallett, A.M.; Sharma, A.P. Adaptation to a low carbohydrate high fat diet is rapid but impairs endurance exercise metabolism and performance despite enhanced glycogen availability. J. Physiol. 2021, 599, 771–790. [Google Scholar] [CrossRef]
- Burke, L.M.; Sharma, A.P.; Heikura, I.A.; Forbes, S.F.; Holloway, M.; McKay, A.K.A.; Bone, J.L.; Leckey, J.J.; Welvaert, M.; Ross, M.L. Crisis of confidence averted: Impairment of exercise economy and performance in elite race walkers by ketogenic low carbohydrate, high fat (LCHF) diet is reproducible. PLoS ONE 2020, 15, e0234027. [Google Scholar] [CrossRef]
- Burke, L.M. Ketogenic low-CHO, high-fat diet: The future of elite endurance sport? J. Physiol. 2020. [Google Scholar] [CrossRef]
- Burke, L.M.; Hall, R.; Heikura, I.A.; Ross, M.L.; Tee, N.; Kent, G.L.; Whitfield, J.; Forbes, S.F.; Sharma, A.P.; Jones, A.M.; et al. Neither Beetroot Juice Supplementation nor Increased Carbohydrate Oxidation Enhance Economy of Prolonged Exercise in Elite Race Walkers. Nutrients 2021, 13, 2767. [Google Scholar] [CrossRef]
- Stellingwerff, T.; Cox, G.R. Systematic review: Carbohydrate supplementation on exercise performance or capacity of varying durations. Appl. Physiol. Nutr. Metab. 2014, 39, 998–1011. [Google Scholar] [CrossRef]
- Rauch, C.; Gaskell, S.K.; McCubbin, A.; Costa, R.J.S. Feeding tolerance, glucose availability, whole-body total carbohydrate and fat oxidation in male endurance and ultra-endurance runners in response to prolonged exercise consuming a habitual mixed macronutrient diet and carbohydrate feeding during exercise. Front. Physiol. 2022, 12, 773054. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, M.; Spriet, L.L. Skeletal muscle energy metabolism during exercise. Nat. Metab. 2020, 2, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.J.S.; Knechtle, B.; Tarnopolsky, M.; Hoffman, M.D. Nutrition for Ultramarathon Running: Trail, Track, and Road. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 130–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, A.J.; O’Hara, J.P.; Morrison, D.J.; Preston, T.; King, R. Carbohydrate dose influences liver and muscle glycogen oxidation and performance during prolonged exercise. Physiol. Rep. 2018, 6, e13555. [Google Scholar] [CrossRef] [Green Version]
- Cox, G.R.; Clark, S.A.; Cox, A.J.; Halson, S.L.; Hargreaves, M.; Hawley, J.A.; Jeacocke, N.; Snow, R.J.; Yeo, W.K.; Burke, L.M. Daily training with high carbohydrate availability increases exogenous carbohydrate oxidation during endurance cycling. J. Appl. Physiol. 2010, 109, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Costa, R.J.S.; Miall, A.; Khoo, A.; Rauch, C.; Snipe, R.; Camoes-Costa, V.; Gibson, P. Gut-training: The impact of two weeks repetitive gut-challenge during exercise on gastrointestinal status, glucose availability, fuel kinetics, and running performance. Appl. Physiol. Nutr. Metab. 2017, 42, 547–557. [Google Scholar] [CrossRef] [Green Version]
- Noakes, T.D.; Rehrer, N.J.; Maughan, R.J. The importance of volume in regulating gastric emptying. Med. Sci. Sports Exerc. 1991, 23, 307–313. [Google Scholar] [CrossRef]
- Lambert, G.P.; Lang, J.; Bull, A.; Eckerson, J.; Lanspa, S.; O’Brien, J. Fluid tolerance while running: Effect of repeated trials. Int. J. Sports Med. 2008, 29, 878–882. [Google Scholar] [CrossRef]
- Gaskell, S.K.; Rauch, C.E.; Parr, A.; Costa, R.J.S. Diurnal versus Nocturnal Exercise-Effect on the Gastrointestinal Tract. Med. Sci. Sports Exerc. 2021, 53, 1056–1067. [Google Scholar] [CrossRef]
- Miall, A.; Khoo, A.; Rauch, C.; Snipe, R.M.J.; Camoes-Costa, V.L.; Gibson, P.R.; Costa, R.J.S. Two weeks of repetitive gut-challenge reduce exercise-associated gastrointestinal symptoms and malabsorption. Scand. J. Med. Sci. Sports 2017, 28, 630–640. [Google Scholar] [CrossRef]
- Kishi, K.; Takase, S.; Goda, T. Enhancement of sucrase-isomaltase gene expression induced by luminally administered fructose in rat jejunum. J. Nutr. Biochem. 1999, 10, 8–12. [Google Scholar] [CrossRef]
- Margolskee, R.F.; Dyer, J.; Kokrashvili, Z.; Salmon, K.S.; Ilegems, E.; Daly, K.; Maillet, E.L.; Ninomiya, Y.; Mosinger, B.; Shirazi-Beechey, S.P. T1R3 and gustducin in gut sense sugars to regulate expression of Na+-glucose cotransporter 1. Proc. Natl. Acad. Sci. USA 2007, 104, 15075–15080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirazi-Beechey, S.P.; Moran, A.W.; Batchelor, D.J.; Daly, K.; Al-Rammahi, M. Glucose sensing and signalling; regulation of intestinal glucose transport. Proc. Nutr. Soc. 2011, 70, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeukendrup, A.E. Training the Gut for Athletes. Sports Med. 2017, 47, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Costa, R.J.; Snipe, R.; Camoes-Costa, V.; Scheer, V.; Murray, A. The Impact of Gastrointestinal Symptoms and Dermatological Injuries on Nutritional Intake and Hydration Status During Ultramarathon Events. Sports Med. Open 2016, 2, 16. [Google Scholar] [CrossRef] [Green Version]
- Gaskell, S.K.; Rauch, C.E.; Costa, R.J.S. Gastrointestinal Assessment and Therapeutic Intervention for the Management of Exercise-Associated Gastrointestinal Symptoms: A Case Series Translational and Professional Practice Approach. Front. Physiol. 2021, 12, 719142. [Google Scholar] [CrossRef]
- Thomas, D.T.; Erdman, K.A.; Burke, L.M. American College of Sports Medicine Joint Position Statement. Nutrition and Athletic Performance. Med. Sci. Sports Exerc. 2016, 48, 543–568. [Google Scholar] [CrossRef]
- Wardenaar, F.; Brinkmans, N.; Ceelen, I.; Van Rooij, B.; Mensink, M.; Witkamp, R.; De Vries, J. Macronutrient Intakes in 553 Dutch Elite and Sub-Elite Endurance, Team, and Strength Athletes: Does Intake Differ between Sport Disciplines? Nutrients 2017, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- McKay, A.K.A.; Stellingwerff, T.; Smith, E.S.; Martin, D.T.; Mujika, I.; Goosey-Tolfrey, V.L.; Sheppard, J.; Burke, L.M. Defining Training and Performance Caliber: A Participant Classification Framework. Int. J. Sports Physiol. Perform. 2022, 17, 317–331. [Google Scholar] [CrossRef]
- Lis, D.; Ahuja, K.D.; Stellingwerff, T.; Kitic, C.M.; Fell, J. Case Study: Utilizing a Low FODMAP Diet to Combat Exercise-Induced Gastrointestinal Symptoms. Int. J. Sport Nutr. Exerc. Metab. 2016, 26, 481–487. [Google Scholar] [CrossRef]
- Lis, D.M.; Stellingwerff, T.; Kitic, C.M.; Fell, J.W.; Ahuja, K.D.K. Low FODMAP: A Preliminary Strategy to Reduce Gastrointestinal Distress in Athletes. Med. Sci. Sports Exerc. 2018, 50, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Gaskell, S.K.; Costa, R.J.S. Applying a Low-FODMAP Dietary Intervention to a Female Ultraendurance Runner with Irritable Bowel Syndrome During a Multistage Ultramarathon. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Gaskell, S.K.; Snipe, R.M.J.; Costa, R.J.S. Test–Retest Reliability of a Modified Visual Analog Scale Assessment Tool for Determining Incidence and Severity of Gastrointestinal Symptoms in Response to Exercise Stress. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Bate, J.P.; Irving, P.M.; Barrett, J.S.; Gibson, P.R. Benefits of breath hydrogen testing after lactulose administration in analysing carbohydrate malabsorption. Eur. J. Gastroenterol. Hepatol. 2010, 22, 318–326. [Google Scholar] [CrossRef]
- Hanley, B. An analysis of pacing profiles of world-class racewalkers. Int. J. Sports Physiol. Perform. 2013, 8, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Nana, A.; Slater, G.J.; Hopkins, W.G.; Halson, S.L.; Martin, D.T.; West, N.P.; Burke, L.M. Importance of Standardized DXA Protocol for Assessing Physique Changes in Athletes. Int. J. Sport Nutr. Exerc. Metab. 2016, 26, 259–267. [Google Scholar] [CrossRef]
- Robertson, E.Y.; Saunders, P.U.; Pyne, D.B.; Gore, C.J.; Anson, J.M. Effectiveness of intermittent training in hypoxia combined with live high/train low. Eur. J. Appl. Physiol. 2010, 110, 379–387. [Google Scholar] [CrossRef]
- King, A.J.; Rowe, J.T.; Burke, L.M. Carbohydrate Hydrogel Products Do Not Improve Performance or Gastrointestinal Distress During Moderate-Intensity Endurance Exercise. Int. J. Sport Nutr. Exerc. Metab. 2020, 30, 305–314. [Google Scholar] [CrossRef]
- Borg, G.A. Psychophysical bases of perceived exertion. Med. Sci. Sports Exerc. 1982, 14, 377–381. [Google Scholar] [CrossRef]
- Pfeiffer, B.; Stellingwerff, T.; Hodgson, A.B.; Randell, R.; Pöttgen, K.; Res, P.; Jeukendrup, A.E. Nutritional Intake and Gastrointestinal Problems during Competitive Endurance Events. Med. Sci. Sports Exerc. 2012, 44, 344–351. [Google Scholar] [CrossRef] [Green Version]
- Costa, R.J.S.; Gaskell, S.K.; McCubbin, A.J.; Snipe, R.M.J. Exertional-heat stress-associated gastrointestinal perturbations during Olympic sports: Management strategies for athletes preparing and competing in the 2020 Tokyo Olympic Games. Temperature 2019, 7, 58–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, R.J.S.; Snipe, R.M.J.; Kitic, C.M.; Gibson, P.R. Systematic review: Exercise-induced gastrointestinal syndrome-implications for health and intestinal disease. Aliment. Pharmacol. Ther. 2017, 46, 246–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, K.A.; Pugh, J.N.; Duca, F.A.; Close, G.L.; Ormsbee, M.J. Gastrointestinal pathophysiology during endurance exercise: Endocrine, microbiome, and nutritional influences. Eur. J. Appl. Physiol. 2021, 121, 2657–2674. [Google Scholar] [CrossRef] [PubMed]
- Snipe, R.M.J.; Khoo, A.; Kitic, C.M.; Gibson, P.R.; Costa, R.J.S. The Impact of Mild Heat Stress During Prolonged Running on Gastrointestinal Integrity, Gastrointestinal Symptoms, Systemic Endotoxin and Cytokine Profiles. Int. J. Sports Med. 2018, 39, 255–263. [Google Scholar] [CrossRef]
- Snipe, R.M.J.; Costa, R.J.S. Does the temperature of water ingested during exertional-heat stress influence gastrointestinal injury, symptoms, and systemic inflammatory profile? J. Sci. Med. Sport 2018, 21, 771–776. [Google Scholar] [CrossRef]
- Gaskell, S.K.; Costa, R.J.; Lis, D. Exercise-induced gastrointestinal syndrome, gastrointestinal disorders, food intolerance and allergies. In Clinical Sports Nutrition, 6th ed.; Burke, L.M., Deakin, V., Minehan, M., Eds.; McGraw Hill: Sydney, Australia, 2021; pp. 551–579. [Google Scholar]
- Smith, J.W.; Pascoe, D.D.; Passe, D.H.; Ruby, B.C.; Stewart, L.K.; Baker, L.B.; Zachwieja, J.J. Curvilinear dose-response relationship of carbohydrate (0-120 g.h(-1)) and performance. Med. Sci. Sports Exerc. 2013, 45, 336–341. [Google Scholar] [CrossRef] [Green Version]
- Costa, R.J.S.; Camoes-Costa, V.; Snipe, R.M.J.; Dixon, D.; Russo, I.; Huschtscha, Z. Impact of exercise-induced hypohydration on gastrointestinal integrity, function, symptoms, and systemic endotoxin and inflammatory profile. J. Appl. Physiol. 1985 2019, 126, 1281–1291. [Google Scholar] [CrossRef]
- Gaskell, S.K.; Taylor, B.; Muir, J.; Costa, R.J.S. Impact of 24-h high and low fermentable oligo-, di-, monosaccharide, and polyol diets on markers of exercise-induced gastrointestinal syndrome in response to exertional heat stress. Appl. Physiol. Nutr. Metab. 2020, 45, 569–580. [Google Scholar] [CrossRef]
- Snipe, R.M.J.; Khoo, A.; Kitic, C.M.; Gibson, P.R.; Costa, R.J.S. The impact of exertional-heat stress on gastrointestinal integrity, gastrointestinal symptoms, systemic endotoxin and cytokine profile. Eur. J. Appl. Physiol. 2018, 118, 389–400. [Google Scholar] [CrossRef]
- Maurten: Technology. Available online: https://www.youtube.com/watch?v=m-am-MuBdP4 (accessed on 8 March 2022).
- Sutehall, S.; Galloway, S.D.R.; Bosch, A.; Pitsiladis, Y. Addition of an Alginate Hydrogel to a Carbohydrate Beverage Enhances Gastric Emptying. Med. Sci. Sports Exerc. 2020, 52, 1785–1792. [Google Scholar] [CrossRef]
- Rowe, J.T.; King, R.; King, A.J.; Morrison, D.J.; Preston, T.; Wilson, O.J.; O’Hara, J.P. Glucose and Fructose Hydrogel Enhances Running Performance, Exogenous Carbohydrate Oxidation, and Gastrointestinal Tolerance. Med. Sci. Sports Exerc. 2021, 54, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, S.; Ahnoff, M.; Edin, F.; Lingström, P.; Simark Mattsson, C.; Andersson-Hall, U. A Hydrogel Drink with High Fructose Content Generates Higher Exogenous Carbohydrate Oxidation and Lower Dental Biofilm pH Compared to Two Other, Commercially Available, Carbohydrate Sports Drinks. Front. Nutr. 2020, 7, 88. [Google Scholar] [CrossRef] [PubMed]
- Rehrer, N.J.; Meijer, G.A. Biomechanical vibration of the abdominal region during running and bicycling. J. Sports Med. Phys. Fit. 1991, 31, 231–234. [Google Scholar]
- Chantler, S.; Griffiths, A.; Matu, J.; Davison, G.; Jones, B.; Deighton, K. The Effects of Exercise on Indirect Markers of Gut Damage and Permeability: A Systematic Review and Meta-analysis. Sports Med. 2021, 51, 113–124. [Google Scholar] [CrossRef]
- Gasbarrini, A.; Corazza, G.R.; Gasbarrini, G.; Montalto, M.; Di Stefano, M.; Basilisco, G.; Parodi, A.; Usai-Satta, P.; Vernia, P.; Anania, C.; et al. Methodology and Indications of H2-Breath Testing in Gastrointestinal Diseases: The Rome Consensus Conference. Aliment. Pharmacol. Ther. 2009, 29, 1–49. [Google Scholar] [CrossRef]
- Russo, I.; Della Gatta, P.A.; Garnham, A.; Porter, J.; Burke, L.M.; Costa, R.J.S. Does the Nutritional Composition of Dairy Milk Based Recovery Beverages Influence Post-exercise Gastrointestinal and Immune Status, and Subsequent Markers of Recovery Optimisation in Response to High Intensity Interval Exercise? Front. Nutr. 2020, 7, 622270. [Google Scholar] [CrossRef]
- Russo, I.; Della Gatta, P.A.; Garnham, A.; Porter, J.; Burke, L.M.; Costa, R.J.S. Assessing Overall Exercise Recovery Processes Using Carbohydrate and Carbohydrate-Protein Containing Recovery Beverages. Front. Physiol. 2021, 12, 628863. [Google Scholar] [CrossRef]
- Russo, I.; Della Gatta, P.A.; Garnham, A.; Porter, J.; Burke, L.M.; Costa, R.J.S. The Effects of an Acute "Train-Low" Nutritional Protocol on Markers of Recovery Optimization in Endurance-Trained Male Athletes. Int. J. Sports Physiol. Perform. 2021, 16, 1764–1776. [Google Scholar] [CrossRef]
- Burke, L.M.; Hawley, J.A. Swifter, higher, stronger: What’s on the menu? Science 2018, 362, 781–787. [Google Scholar] [CrossRef] [Green Version]
- Snipe, R.M.J.; Khoo, A.; Kitic, C.M.; Gibson, P.R.; Costa, R.J.S. Carbohydrate and protein intake during exertional heat stress ameliorates intestinal epithelial injury and small intestine permeability. Appl. Physiol. Nutr. Metab. 2017, 42, 1283–1292. [Google Scholar] [CrossRef]
- Flood, T.R.; Montanari, S.; Wicks, M.; Blanchard, J.; Sharpe, H.; Taylor, L.; Kuennen, M.R.; Lee, B.J. Addition of pectin-alginate to a carbohydrate beverage does not maintain gastrointestinal barrier function during exercise in hot-humid conditions better than carbohydrate ingestion alone. Appl. Physiol. Nutr. Metab. 2020, 45, 1145–1155. [Google Scholar] [CrossRef] [PubMed]
- Jonvik, K.L.; Lenaerts, K.; Smeets, J.S.J.; Kolkman, J.J.; LJC, V.A.N.L.; Verdijk, L.B. Sucrose but Not Nitrate Ingestion Reduces Strenuous Cycling-induced Intestinal Injury. Med. Sci. Sports Exerc. 2019, 51, 436–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehrer, N.J.; Goes, E.; DuGardeyn, C.; Reynaert, H.; DeMeirleir, K. Effect of carbohydrate on portal vein blood flow during exercise. Int. J. Sports Med. 2005, 26, 171–176. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, E.P.; Burini, R.C. Carbohydrate-dependent, exercise-induced gastrointestinal distress. Nutrients 2014, 6, 4191–4199. [Google Scholar] [CrossRef] [Green Version]
- Zuhl, M.; Schneider, S.; Lanphere, K.; Conn, C.; Dokladny, K.; Moseley, P. Exercise regulation of intestinal tight junction proteins. Br. J. Sports Med. 2014, 48, 980–986. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Hernandez, V.; Quiros, M.; Nusrat, A. Intestinal epithelial claudins: Expression and regulation in homeostasis and inflammation. Ann. N. Y. Acad. Sci. 2017, 1397, 66–79. [Google Scholar] [CrossRef]
- Mirtschin, J.G.; Forbes, S.F.; Cato, L.E.; Heikura, I.A.; Strobel, N.; Hall, R.; Burke, L.M. Organization of Dietary Control for Nutrition-Training Intervention Involving Periodized Carbohydrate Availability and Ketogenic Low-Carbohydrate High-Fat Diet. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 480–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, L.M. Fueling strategies to optimize performance: Training high or training low? Scand. J. Med. Sci. Sports 2010, 20 (Suppl. 2), 48–58. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study 1: Race Walkers | Study 2: Marathon Runners | |||
|---|---|---|---|---|
| CON (n = 10 M) | MAX (n = 9 M) | CON (n = 7 M, 3F) | MAX (n = 5 M, 3F) | |
| Age (years) | 29.4 (4.6) | 29.7 (4.2) | 35.1 (7.2) | 30.5 (8.1) |
| Body Mass (BM) (kg) | 68.4 (9.4) | 68.7 (5.0) | 60.6 (8.5) | 57.3 (9.7) |
| VO2max (mL·kg−1·min−1) | 60.9 (5.3) | 63.1 (4.6) | 66.8 (5.8) | 65.3 (7.2) |
| Personal Best (h:min:s) | 01:24:30 * | 01:23:04 * | 02:29:00 $ | 02:31:00 $ |
| (0:04:45) | (0:01:59) | (0:06:00) | (0:08:58) | |
| Pre-Intervention Loading | Intervention | Post-Intervention Loading | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MAX | CON | MAX | CON | MAX | CON | |||||||||||||||
| Goal Intake | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | ||||||||
| Energy | kJ | 13,035 | ± | 2157 | 13,452 | ± | 1390 | 13,922 | ± | 2178 | 13,414 | ± | 1365 | 15,024 | ± | 2404 bbb | 13,757 | ± | 1596 | |
| kJ·kg−1 | 225 | ± | 5.1 | 223 | ± | 8.7 | 241 | ± | 21 | 223 | ± | 16 | 259 | ± | 11 a,bbb | 229 | ± | 30 | ||
| Carbohydrate | g | Pre: 8 g·kg−1 Intervention: MAX: 10 g·kg−1 CON: 6.5 g·kg−1 Post: MAX: 12 g·kg−1 | 465 | ± | 77 | 486 | ± | 54 | 605 | ± | 88 aa | 415 | ± | 45 | 712 | ± | 117 aa,bbb | 477 | ± | 55 |
| g·kg−1 | 8.0 | ± | 0.1 | 8.1 | ± | 0.2 | 10.5 | ± | 0.6 aa | 6.9 | ± | 0.4 | 12.3 | ± | 0.4 aa,bbb | 7.9 | ± | 0.2 | ||
| Sugars | g | 240 | ± | 43 | 254 | ± | 33 | 291 | ± | 42 aa | 207 | ± | 23 | 320 | ± | 74 a,bb | 252 | ± | 35 | |
| Starch | g | 223 | ± | 44 | 230 | ± | 27 | 313 | ± | 50 aa | 206 | ± | 25 | 388 | ± | 46 aa,bbb | 223 | ± | 29 | |
| Protein | g | 111 | ± | 21 | 112 | ± | 9.6 | 125 | ± | 28 | 124 | ± | 16 | 113 | ± | 20 | 123 | ± | 30 | |
| g·kg−1 | 1.8 g.kg−1 | 1.9 | ± | 0.2 | 1.9 | ± | 0.1 | 2.2 | ± | 0.3 | 2.1 | ± | 0.3 | 1.9 | ± | 0.2 | 2.1 | ± | 0.7 | |
| Fat | g g·kg−1 | Pre: 1.5 g·kg−1 | 90 1.5 | ± ± | 15 0.1 | 93 1.5 | ± ± | 10 0.1 | 46 0.8 | ± ± | 10 aa 0.2 aa | 117 2 | ± ± | 13 0.2 | 32 0.6 | ± ± | 7 aa,bbb 0.1 aa,bbb | 99 1.7 | ± ± | 23 0.5 |
| Intervention: | ||||||||||||||||||||
| MAX: 0.5 g·kg−1 CON: 1.8 g·kg−1 | ||||||||||||||||||||
| Post: MAX: 0.5 g·kg−1 | ||||||||||||||||||||
| Saturated | g | 31 | ± | 7 | 32 | ± | 5 | 14 | ± | 4 aa | 40 | ± | 5 | 11 | ± | 3 aa,bbb | 36 | ± | 14 | |
| Monounsat | g | 35 | ± | 6 | 36 | ± | 4 | 17 | ± | 4 aa | 50 | ± | 6 | 12 | ± | 2 aa,bbb | 37 | ± | 6 | |
| Polyunsat | g | 15 | ± | 3 | 16 | ± | 1 | 9 | ± | 2 aa | 17 | ± | 3 | 5 | ± | 1 aa,bbb | 16 | ± | 4 | |
| Cholesterol | mg | 239 | ± | 70 | 222 | ± | 36 | 272 | ± | 97 a | 379 | ± | 90 | 191 | ± | 57 a | 249 | ± | 51 | |
| Fibre | g | 47 | ± | 7 | 47 | ± | 5 | 50 | ± | 7 | 48 | ± | 4 | 29 | ± | 3 aa,bbb | 47 | ± | 5 | |
| g·kg−1 | 0.8 | ± | 0.1 | 0.8 | ± | 0.1 | 0.9 | ± | 0.1 | 0.8 | ± | 0.1 | 0.5 | ± | 0.1 aa,bbb | 0.8 | ± | 0.1 | ||
| Pre-Intervention Loading | Intervention | Post-Intervention Loading | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MAX | CON | MAX | CON | MAX | CON | |||||||||||||||
| Goal Intake | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | ||||||||
| Thiamin | mg | RDI: 1.2 mg·d−1 | 2.6 | ± | 0.5 | 2.6 | ± | 0.3 | 2.8 | ± | 1 | 2.7 | ± | 1.2 | 3.5 | 0.7 bbb | 2.9 | ± | 0.9 | |
| mg/MJ | 0.2 | ± | 0 | 0.2 | ± | 0 | 0.19 | ± | 0.05 | 0.19 | ± | 0.07 | 0.22 | ± | 0.02 bb | 0.21 | ± | 0.06 | ||
| Riboflavin | mg | RDI: 1.3 mg·d−1 | 2.6 | ± | 0.4 | 2.8 | ± | 0.3 | 3.3 | ± | 0.8 | 3.2 | ± | 0.9 | 3.6 | ± | 0.6 bb | 3.2 | ± | 0.7 |
| mg/MJ | 0.2 | ± | 0 | 0.2 | ± | 0 | 0.2 | ± | 0 | 0.2 | ± | 0.1 | 0.2 | ± | 0.0 b | 0.2 | ± | 0 | ||
| Niacin equivalents | mg | RDI: 16 mg·d−1 | 47 | ± | 8 | 48 | ± | 5 | 55 | ± | 16 | 52 | ± | 13 | 56 | ± | 10 bbb | 51 | ± | 9 |
| mg/MJ | 3.5 | ± | 0.2 | 3.4 | ± | 0.2 | 3.8 | ± | 0.6 | 3.8 | ± | 9.8 | 3.6 | ± | 0.2 | 3.6 | ± | 0.5 | ||
| Vitamin C | mg | RDI: 45 mg·d−1 | 261 | ± | 68 | 272 | ± | 58 | 266 | ± | 42 aaa | 200 | ± | 28 | 376 | ± | 173 | 280 | ± | 46 |
| mg/MJ | 20 | ± | 4 | 19 | ± | 3 | 18 | ± | 1 aa | 15 | ± | 3 | 24 | ± | 7 | 20 | ± | 2 | ||
| Dietary Folate Equivalents | µg | RDI: 400 µg.d−1 | 1497 111 | ± ± | 291 13 | 1495 107 | ± ± | 191 7 | 1310 90 | ± ± | 331 13 | 1235 89 | ± ± | 274 16 | 2091 135 | ± ± | 426 a,bb 18b | 1665 119 | ± ± | 245 b 14 b |
| µg/MJ | ||||||||||||||||||||
| Retinol equivalents | µg | RDI: 900 µg.d−1 | 1326 | ± | 210 | 1381 | ± | 229 | 1632 | ± | 211 | 1642 | ± | 248 | 1386 | ± | 315 | 1477 | ± | 169 |
| µg/MJ | 100 | ± | 17 | 100 | ± | 18 | 114 | ± | 19 | 121 | ± | 24 | 91 | ± | 22 | 106 | ± | 15 | ||
| Iron | mg | RDI: 8 mg·d−1 | 17 | ± | 3 | 18 | ± | 2 | 21 | ± | 3 | 22 | ± | 2 | 23 | ± | 4 aa,bb | 18 | ± | 2 |
| mg/MJ | 1.3 | ± | 0.1 | 1.3 | ± | 0.1 | 1.5 | ± | 0.1 | 1.6 | ± | 0.2 | 1.5 | ± | 0.1 aa,b | 1.3 | ± | 0.1 | ||
| Potassium | mg | AI: 3800 mg·d−1 | 5872 | ± | 812 | 6129 | ± | 598 | 6250 | ± | 867 | 5626 | ± | 558 | 5323 | ± | 837 a,bb | 6128 | ± | 512 |
| mg/MJ | 437 | ± | 32 | 440 | ± | 19 | 433 | ± | 27 | 407 | ± | 40 | 343 | ± | 25 aaa,bbb | 437 | ± | 25 | ||
| Magnesium | mg | RDI: 400 mg·d−1 | 623 | ± | 89 | 649 | ± | 66 | 630 | ± | 106 | 647 | ± | 58 | 406 | ± | 58 aaa,bbb | 652 | ± | 82 |
| mg/MJ | 46 | ± | 2 | 47 | ± | 2 | 44 | ± | 5 a | 47 | ± | 4 | 26 | ± | 2 aaa,bbb | 46 | ± | 4 | ||
| Calcium | mg | RDI: 1000 mg·d−1 | 1214 | ± | 201 | 1354 | ± | 257 | 1225 | ± | 162 aa | 1354 | ± | 139 | 986 | ± | 178 b | 1587 | ± | 852 |
| mg/MJ | 90 | ± | 10 | 97 | ± | 13 | 85 | ± | 8 aa | 98 | ± | 10 | 64 | ± | 10 aaa,bbb | 102 | ± | 26 | ||
| Zinc (mg) | mg | RDI: 14 mg·d−1 | 17 | ± | 3 | 18 | ± | 2 | 17 | ± | 3 | 17 | ± | 2 | 20 | ± | 3 bb | 19 | ± | 4 |
| mg/MJ | 1.2 | ± | 0.1 | 1.3 | ± | 0.1 | 1.2 | ± | 0 | 1.2 | ± | 0 | 1.3 | ± | 0.1 | 1.3 | ± | 0.1 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
King, A.J.; Etxebarria, N.; Ross, M.L.; Garvican-Lewis, L.; Heikura, I.A.; McKay, A.K.A.; Tee, N.; Forbes, S.F.; Beard, N.A.; Saunders, P.U.; et al. Short-Term Very High Carbohydrate Diet and Gut-Training Have Minor Effects on Gastrointestinal Status and Performance in Highly Trained Endurance Athletes. Nutrients 2022, 14, 1929. https://doi.org/10.3390/nu14091929
King AJ, Etxebarria N, Ross ML, Garvican-Lewis L, Heikura IA, McKay AKA, Tee N, Forbes SF, Beard NA, Saunders PU, et al. Short-Term Very High Carbohydrate Diet and Gut-Training Have Minor Effects on Gastrointestinal Status and Performance in Highly Trained Endurance Athletes. Nutrients. 2022; 14(9):1929. https://doi.org/10.3390/nu14091929
Chicago/Turabian StyleKing, Andy J., Naroa Etxebarria, Megan L. Ross, Laura Garvican-Lewis, Ida A. Heikura, Alannah K. A. McKay, Nicolin Tee, Sara F. Forbes, Nicole A. Beard, Philo U. Saunders, and et al. 2022. "Short-Term Very High Carbohydrate Diet and Gut-Training Have Minor Effects on Gastrointestinal Status and Performance in Highly Trained Endurance Athletes" Nutrients 14, no. 9: 1929. https://doi.org/10.3390/nu14091929
APA StyleKing, A. J., Etxebarria, N., Ross, M. L., Garvican-Lewis, L., Heikura, I. A., McKay, A. K. A., Tee, N., Forbes, S. F., Beard, N. A., Saunders, P. U., Sharma, A. P., Gaskell, S. K., Costa, R. J. S., & Burke, L. M. (2022). Short-Term Very High Carbohydrate Diet and Gut-Training Have Minor Effects on Gastrointestinal Status and Performance in Highly Trained Endurance Athletes. Nutrients, 14(9), 1929. https://doi.org/10.3390/nu14091929