Abstract
Increased hepatic lipid content and decreased insulin sensitivity have critical roles in the development of cardiometabolic diseases. Therefore, our objective was to investigate the dose-response effects of consuming high fructose corn syrup (HFCS)-sweetened beverages for two weeks on hepatic lipid content and insulin sensitivity in young (18–40 years) adults (BMI 18–35 kg/m2). In a parallel, double-blinded study, participants consumed three beverages/day providing 0% (aspartame: n = 23), 10% (n = 18), 17.5% (n = 16), or 25% (n = 28) daily energy requirements from HFCS. Magnetic resonance imaging for hepatic lipid content and oral glucose tolerance tests (OGTT) were conducted during 3.5-day inpatient visits at baseline and again at the end of a 15-day intervention. During the 12 intervening outpatient days participants consumed their usual diets with their assigned beverages. Significant linear dose-response effects were observed for increases of hepatic lipid content (p = 0.015) and glucose and insulin AUCs during OGTT (both p = 0.0004), and for decreases in the Matsuda (p = 0.0087) and Predicted M (p = 0.0027) indices of insulin sensitivity. These dose-response effects strengthen the mechanistic evidence implicating consumption of HFCS-sweetened beverages as a contributor to the metabolic dysregulation that increases risk for nonalcoholic fatty liver disease and type 2 diabetes.
1. Introduction
Approximately 1 in 10 or over 500 million adults worldwide have diabetes with individuals with type 2 diabetes (T2D) representing over 90% of cases [1,2]. Diabetes is one of the largest global health challenges in the 21st century with the number of cases estimated to rise to 700 million by 2045 [1]. The incidence of nonalcoholic fatty liver disease (NAFLD) among adults is also rising [3] with the global prevalence estimated at 25% [4] and U.S. prevalence at 37% [5]. There is a well-documented link between T2D and NAFLD with both being associated with insulin resistance and an increased risk of metabolic syndrome and cardiovascular disease (CVD) [6,7]. T2D has been described as the strongest predictor of NAFLD and driver of disease progression [8]. Likewise, it has been reported that NAFLD is one of the strongest clinical risk factors for T2D [9]. While insulin resistance is involved in this bidirectional relationship, it is unclear whether hepatic insulin resistance develops first and primes the liver for subsequent fat accumulation or if hepatic insulin resistance is primarily a consequence of increased fat accumulation in the liver [10].
Industrialization has brought pronounced changes in human behavior and lifestyle that have contributed to escalating rates of both T2D and NAFLD [11,12]. These changes include the marked increases in the availability of added sugars and low fiber processed foods [13,14]. Data from the National Health and Nutrition Examination Survey showed that processed foods comprised 65% of energy intake and contributed 91% of the energy intake from added sugars [15,16]. Sugar-sweetened beverages (-SBs), primarily sweetened with high fructose corn syrup (HFCS), are a major contributor of added sugars in the U.S. diet, with youth and adults consuming over 140 kcal per day on average [17,18].
Evidence from observational studies have consistently demonstrated positive associations between increased consumption of sugar-SBs and insulin resistance [19], T2D, [20] and NAFLD [21]. Evidence from clinical dietary intervention studies have shown that increased and sustained (+7 days) exposure to sugar-SBs increases liver lipid content [22,23,24,25] and decreases insulin sensitivity [22,24,26,27]. Clinical dietary intervention studies designed to decrease consumption of sugar-SB [28], HFCS [29], or free/added sugars [30,31,32] have led to reductions in hepatic lipid content.
Both HFCS and sucrose contain fructose and glucose; however, clinical [26] and animal studies [33,34] have demonstrated that it is the unique metabolism of fructose that is the key mediator of the relationship between sugar-SB exposure and metabolic dysregulation. Unlike glucose, the initial phosphorylation of fructose is catalyzed by fructokinase. Fructokinase operates independently of hepatic energy status, resulting in unregulated fructose uptake in both the fasted and fed state. Therefore, as much as 85% of the fructose in a sugar-SB will be taken up during its first pass through the liver [35,36]. The resulting fructose overload in the liver increases uric acid and lactate production. Fructose and/or its metabolites induce expression of sterol regulatory element binding protein 1c (SREBP-1c) and carbohydrate response element binding protein (ChREBP) and lead to increased de novo lipogenesis (DNL) [37,38] and apolipoprotein CIII (apoCIII) production [38,39], and decreased fat oxidation [10,40]. Studies in animal models and in vitro studies suggest hepatic fructose overload has direct effects on endoplasmic reticulum (ER) stress and inflammation that may lead to insulin resistance [10]. However, it has also been suggested that the increased hepatic lipid supply resulting from upregulated DNL and inhibited fat oxidation leads to increased levels of lipids such as diacylglycerol (DAG) that decrease hepatic insulin sensitivity by interfering with insulin receptor activation [41]. The increased hepatic lipid supply may also promote increased production and secretion of very low-density lipoprotein (VLDL) contributing to increased levels of circulating triglyceride (TG) and cholesterol [42]. Hepatic insulin resistance further promotes VLDL production and secretion by increasing apolipoprotein B (apoB) [43] and apoCIII [44] availability/expression. The ability of the insulin resistant liver to stimulate glycogen synthesis and suppress glycogenolysis is impaired [45], which then increases circulating glucose and compensatory insulin secretion. The compensatory hyperinsulinemia [46] and/or the chronic oversupply of substrate [45] continue to drive DNL.
Sustained increases of circulating TG may contribute to muscle lipid accumulation leading to impaired muscle insulin action and lowered whole body insulin sensitivity [47]. Fructose consumption may also impair glucose transport into muscle and adipose tissue [48,49], which could be a direct effect of fructose or one mediated by high levels of lactic acid decreasing expression of GLUT4 [50,51]. Uric acid may further contribute to the metabolic dysregulation by activation of fructokinase and induction of mitochondrial oxidative stress [52,53,54,55].
Our previously published paper, reporting dose-dependent increases in uric acid, TG, apoCIII, low density lipoprotein cholesterol (LDL-C), and apoB in young adults consuming 0, 10, 17.5, or 25% Ereq HFCS for two weeks [56], provides mechanistic support for HFCS as a mediator of the metabolic dysregulation described above. However, dose-dependent effects of HFCS on critical nodes in the progression of metabolic disease, specifically increased hepatic lipid content and decreased insulin sensitivity, have not been documented. Therefore, we hypothesized that two weeks of consumption of HFCS-SB at 0, 10, 17.5, or 25% Ereq would result in dose-dependent increases of hepatic lipid content and dose-dependent decreases in insulin sensitivity in the same young men and women [56]. In addition, to determine if our results supported the mechanisms outlined above, we conducted preliminary mediation analyses. We specifically sought to determine if the changes in hepatic lipid content potentially mediate the effects of HFCS dose on insulin sensitivity and on circulating TG concentrations. We also sought to determine if outcomes of other metabolic pathways directly affected by hepatic fructose overload, specifically increased uric acid and lactate concentrations, were potential mediators of hepatic lipid accumulation or decreased insulin sensitivity.
2. Materials and Methods
2.1. Participants
The current paper reports the results from 85 participants consuming three sweetened beverages per day containing either 0% (n = 23), 10% (n = 18), 17.5% (n = 16), or 25% (n = 28) of energy requirements (Ereq) from HFCS. The participants are a subgroup from an NIH-funded investigation that included 187 participants allocated to eight experimental groups. The results demonstrating the dose-response effects of HFCS-SB on circulating lipid/lipoprotein and uric acid concentrations from these 85 participants have been previously published [56]. Additionally, hepatic lipid content and insulin sensitivity results from the 28 participants who consumed 25% Ereq-HFCS and the 23 participants who consumed 0% Ereq-aspartame were recently reported [22]. Relevant results from this publication [22] are reported here in order to assess the dose-response effects of consuming 0, 10, 17.5, or 25% HFCS-SB on hepatic lipid content and insulin sensitivity.
This study was conducted in accordance with an experimental protocol that was approved by the University of California, Davis Institutional Review Board and is registered with Clinical Trials.gov (accessed on 7 November 2008): NCT0110392. Participants were recruited through a Craigslist.com listing and local flyer advertisements. Eligibility was assessed through telephone and in-person interviews followed by a complete blood count and serum biochemistry panel. All participants provided written informed consent. Inclusion criteria included males and females aged 18–40 years with a body mass index (BMI) range of 18–35 kg/m2 and a self-reported stable body weight of at least six months. Exclusion criteria included: diabetes (fasting glucose > 125 mg/dL), evidence of renal or hepatic disease according to aspartate aminotransferase (AST) and alanine aminotransferase (ALT) 1.5 normal limits ratio, fasting plasma TG > 400 mg/dL, hypertension (>140/90 mmHg), hemoglobin < 8.5 g/dL, and surgery for weight loss. Individuals were also excluded if they smoked, habitually ingested more than two alcoholic beverages/day, exercised more than 3.5 h/week at an intensity greater than walking, or used thyroid, lipid-lowering, glucose-lowering, anti-hypertensive, anti-depressant, or weight loss medications. By design, experimental groups were not randomized, but instead matched for sex, BMI, and circulating fasting TG, LDL-C, high-density lipoprotein cholesterol (HDL) and insulin concentrations measured in fasted serum collected during the in-person interviews. Five weeks before the start of study, enrolled participants were asked to limit their consumption of sugar-containing beverages to no more than one 8-oz serving of 100% fruit juice per day and to stop the use of any vitamin, mineral, dietary, or herbal supplements. Participant enrollment is detailed in a previous publication with the same experimental groups [56].
2.2. Dietary Protocol
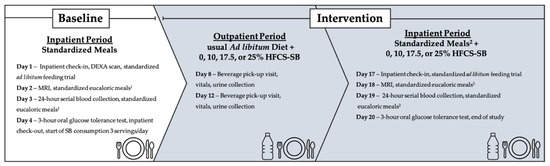
As previously described [22], this was a double-blinded, parallel-arm diet intervention study with three phases (Figure 1). The first phase included a 3.5-day inpatient baseline period at the University of California Davis Clinical and Translational Science Center’s Clinical Research Center (CCRC), where participants consumed a standardized low sugar diet and participated in experimental procedures. The second phase consisted of a 12-day outpatient intervention period where participants consumed their designated SBs providing 0% (aspartame-sweetened) or 10%, 17.5%, or 25% Ereq-HFCS along with their usual ad libitum diets. Phase three included a 3.5-day inpatient intervention period at the CCRC, where participants consumed a standardized diet that included their designated SBs and repeated all experimental procedures. Participants were instructed to maintain their usual physical activity level throughout the study. Fasted body weight was measured during baseline (Day 1) and intervention (Day 17) during inpatient testing periods at the CCRC with standardized attire.
Figure 1.
Study design and experimental testing and collection days. DEXA, dual energy X-ray absorptiometry; MRI, magnetic resonance imaging; SB, sweetened beverage; HFCS, high fructose corn syrup. 1 Fifty-five percent complex carbohydrate (<2%added sugar), 35% fat, 15% protein. 2 55–30% complex carbohydrate (depending on assigned SB, <2%added sugar), 35% fat, 15% protein. % = % of energy requirement.
2.2.1. Inpatient Meals
Identical low sugar ad libitum meals were provided to all participants on Days 1 (baseline) and 17 (intervention) [57]. During Days 2 and 3 of the baseline inpatient period, energy-balanced meals consisting of 55% Ereq as predominantly low fiber complex carbohydrate (i.e., white bread, rice, and pasta), 30% fat, and 15% protein were provided. The meals provided on Days 18 and 19 of the intervention inpatient period were matched as closely as possible to baseline meals, including for fiber content, except for the isocaloric replacement of refined complex carbohydrate with the assigned HFCS-SB. The timing of inpatient meals and the energy distribution were: Breakfast 09:00-h (25% Ereq); Lunch 13:00-h (35% Ereq); Dinner 18:00-h (40% Ereq). Daily Ereq was calculated using the Mifflin equation with a 1.3 adjustment for physical activity on the days of the 24-h serial blood collections, and a 1.5 adjustment for the other days [58]. Participant-specific meal formulations were generated and provided to the kitchen staff by the Study Supervisor, who did not interact with participants. Meals were served by the Study Coordinator and CCRC nurses who were blinded to group assignment.
2.2.2. Study Beverages and Outpatient Meals
Beverages were prepared by a designated staff member, who did not interact with participants, at the UC Davis, Department of Nutrition Ragle Clinical Research Center. All HFCS-containing beverages were flavored with an unsweetened powdered drink mix (Kool-aid®, Kraft Inc., Chicago, IL, USA) and sweetened with HFCS-55 (Isosweet 5500, 55% fructose, 45% glucose, Skidmore Sales and Distributing, Cincinnati, OH, USA), and 0% HFCS-containing beverages were prepared using a fruit-flavored aspartame drink mix (Market Pantry®, Target Brands Inc, Minneapolis, MN, USA). The aspartame drink mix was also used to increase the sweetness of the 10% Ereq-HFCS beverages. Study participants were blinded to their beverage assignment and voluntary feedback indicated that they were unable to distinguish between beverages containing aspartame or HFCS. Beverage amounts (grams) were standardized among the four groups and based on individual Ereq. During the 12-day outpatient intervention period, participants were instructed to drink one serving of study beverage with each meal of their usual diet and to not consume other sweetened beverages, including 100% fruit juice and fruit drinks. To monitor compliance, the study beverages contained a biomarker (0.015 mg riboflavin/mL; mean intake 16 mg riboflavin/day) that was measured fluorometrically in urine samples collected at the CCRC and during beverage-pickup appointments. Participants were informed of the biomarker in the study beverages but not its identity. Fasting urinary riboflavin concentrations were assessed in samples collected on Days 8, 12, and 17–20. Concentrations did not differ among groups, suggesting comparable compliance among all groups. Concentrations during or following outpatient beverage consumption (Days 8, 12, 17) were not different from those during monitored beverage consumption at the CCRC (Days 18–20), suggesting comparable compliance during the intervention outpatient and inpatient periods [56,59].
2.3. Hepatic Lipid Imaging
As previously described [22], on Day 2 (baseline) and Day 17 (intervention) of inpatient periods, magnetic resonance imaging (MRI) for hepatic lipid content was performed. Participants were scanned using a confounder-corrected chemical shift-encoded hepatic fat quantification MRI technique on a 1.5-Tesla system (General Electric Healthcare, Milwaukee, WI, USA, 1.5 T HDtx system, with an 8-channel body coil). To estimate the liver proton density fat fraction (PDFF), a quantitative image biomarker of liver fat content [60], axial 2D, T1-independent, T2*-corrected, 6-echo gradient-recalled-echo images were acquired (repetition time [TR] = 125 ms, echo time [TE] = 2.3, 4.6, 6.9, 9.2, 11.5, 13.8 ms, 8 mm slice thickness, 256 × 192 matrix size).
Hepatic Lipid Content Quantification
Participants with missing or incomplete scans were not included in the analysis of hepatic lipid content, leaving a total of 75 participants out of 85. Of the 10 sets of missing scans (aspartame-SB = 3; 10% HFCS-SB = 2; 17.5% HFCS-SB = 0; 25%HFCS-SB = 5), one was due to participant discomfort with the procedure (aspartame-SB), one was due to corrupt file format (aspartame-SB), and nine due to scanner unavailability due to malfunction or scheduling issues. Thus, the sample sizes available for analysis of hepatic lipid content were: aspartame-SB = 20; 10% HFCS-SB = 16; 17.5% HFCS-SB = 16; 25% HFCS-SB = 23. As previously described [22], MRI-PDFF was quantified using OsiriX software (OsiriX MD versions 10 & 11; Pixmeo, Geneva, Switzerland) and the LIPO-Quant (Liver Imaging of Phase-interference related signal Oscillation and Quantification) algorithm [61]. This algorithm assumes exponential decay and incorporates a multi-peak spectral model [62,63]. The technologists performing the scans and the image analyst were blinded to participant group and data.
2.4. Insulin Sensitivity
Three-hour Oral Glucose Tolerance Tests (OGTT) were conducted following a 14-h overnight fast at the CCRC on Day 4 (baseline) and Day 20 (intervention). A fasting blood sample was collected at 8:00-h through an intravenous catheter. Blood samples were collected 30, 60, 90, 120, and 180 min following a 75 g oral glucose load. One participant in the 17.5% HFCS group left the study on Day 19 due to a family emergency, thus did not participate in the intervention OGTT. Plasma samples were analyzed for glucose concentrations using a YSI glucose analyzer (YSI Inc., Yellow Springs, OH, USA) and for insulin with radioimmunoassay (Millipore Inc., St. Charles, MO, HI-14K). Two-hour OGTT data were used to derive two separate indices of insulin sensitivity: 2-h Matsuda Insulin Sensitivity Index (ISI) [64] and Predicted M ISI [65]. The Predicted M ISI utilizes the oral glucose insulin sensitivity index [66] and includes an adjustment for BMI [65]. Due to missing OGTT samples, Predicted M could not be calculated for six subjects (aspartame-SB = 1; 10% HFCS-SB = 2; 17.5% HFCS-SB = 2, 25% HFCS-SB = 1). Three-hour OGTT data were used to calculate total area under the curve (AUC) for glucose and insulin by the trapezoidal method. Fasting plasma glucose and insulin concentrations from the OGTT were used to calculate homeostasis model assessment—insulin resistance (HOMA-IR) [67].
2.5. Twenty-Four Hour Serial Blood Collection
Twenty-four-hour serial blood samples were collected via an intravenous catheter on Day 3 (baseline) and Day 19 (intervention). Three fasted samples were collected at 8:00-h, followed by 29 postprandial samples collected at 30- to 60-min intervals until 8:00-h the following morning. An extra 6 mL of blood were collected in the morning (8:00-, 8:30-, 9:00-h) and at late evening time points (22:00-, 23:00-, 24:00-h). The additional plasma from the fasting morning samples and the late-evening samples were pooled separately and aliquoted into 24 identical samples. The timing of the postprandial pool collection was based on the late-evening postprandial TG peak concentrations observed in our previous study [26]. The plasma concentrations of TG, uric acid, glucose, insulin, and lactate were analyzed at all time points and calculated for total 24-h AUC using the trapezoidal method. Lactate, glucose, and insulin amplitudes (AMP) were calculated as the difference between the post-meal peak concentration minus the pre-meal nadir for each of the three meals, then averaged. ApoCIII concentrations were measured during fasting and late-evening postprandial periods. TG, uric acid, and apoCIII concentrations were measured with a Polychem Chemistry Analyzer (PolyMedCo Inc., Cortlandt Manor, NY, USA) with reagents from MedTest DX (HORIBA, Ltd., Kyoto, Japan). Lactate was measured concurrently with glucose with a YSI Analyzer (YSI Inc., Yellow Springs, OH, USA). All assays were conducted by technicians who were blinded to beverage group assignment. The intra- and interassay coefficient of variation for the assays were as follows: glucose 3.6%, 4.5%; insulin: 6.5%, 7.6%; TG: 2.2%, 7.2%; uric acid: 1.9%, 14.5%; apoCIII: 0.9%, 5.5%.
2.6. Statistical Analyses
Data were log transformed when the baseline or absolute change values were not normally distributed (Shapiro–Wilk). The absolute change (∆) at two weeks of intervention compared with baseline for each outcome was tested for a dose-response trend in a general linear model (SAS 9.3; SAS, Cary, NC, USA), adjusted for sex, metabolic syndrome risk factor (MSRF), and outcome at baseline, using HFCS dose (0, 10, 17.5, 25) as a continuous variable. Departures from linearity were tested with polynomial terms using the same model. MSRF scores were assessed by assigning a value of 1 to measured clinical outcomes outside of established cutoffs [68] (waist circumference 120 cm for men, 88 cm for women, and 90 cm for Asian men and 80 cm for Asian women [69]; blood pressure 130 mmHg for systolic or 85 mmHg diastolic; fasting plasma glucose 100 mg/dl; serum TG 150 mg/dl; serum HDL 40 for men and 50 for women) and 0 if not, then taking the sum. To test for differences between and within groups, the ∆ or %∆ of each outcome was also analyzed in a general linear model (SAS 9.3); adjusted for sex, MSRF, and outcome at baseline; with HFCS-group (1, 2, 3, 4) as a categorical variable. Significant changes from baseline concentrations were identified as least squares means (LS means) of ∆ different from zero, and significant differences between groups were identified by Tukey’s multiple comparisons test. Statistical significance was considered at p < 0.05. Mediation analyses that included adjustment for sex and MSRF were conducted between the absolute change of correlated outcomes that were both significantly affected by HFCS dose [70,71]. These analyses only included subjects without missing data points, leaving a total of 71 participants (aspartame = 20; 10% HFCS-SB = 15; 17.5% HFCS-SB = 14; 25% HFCS-SB = 22). The percent attenuation due to mediation was calculated as: (1—(R of HFCS dose effect in model that controls for mediator/R of HFCS dose effect in model that does not control for mediator)) × 100. A multivariate analysis of the Δhepatic lipid content that included HFCS dose, sex, MSRF, and all mediators/partial mediators was conducted. Data are reported as mean ± standard deviation (SD) in Table 1, while all other data are means ± standard error of the mean (SEM).
Table 1.
Participants’ characteristics at baseline.
3. Results
As previously reported [56] and shown in Table 1, there were no significant differences between the four experimental groups in baseline anthropomorphic or metabolic parameters. The absolute values for each outcome at baseline and intervention by HFCS dose group and the p-values for the effects of HFCS dose, sex, and MSRF are presented in Table 2. Table 2 includes body weight, which as previously reported, was significantly affected by HFCS dose, but was not significantly different between groups [56]. The absolute ∆ in hepatic lipid content is presented in Figure 2, and the %∆ in the insulin sensitivity indices (Matsuda, Predicted M ISI, 3-h OGTT AUC for glucose and insulin) are presented in Figure 3 and Figure 4, with notations depicting significant differences between and within groups.
Table 2.
Outcomes at baseline and intervention.
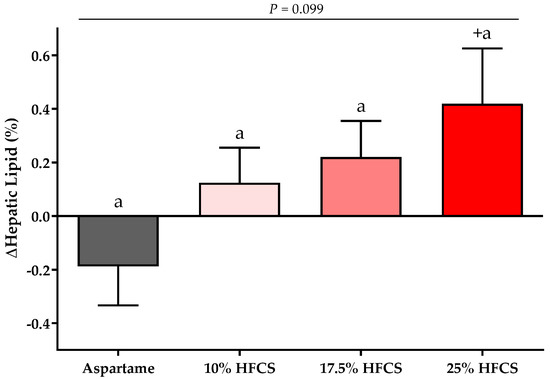
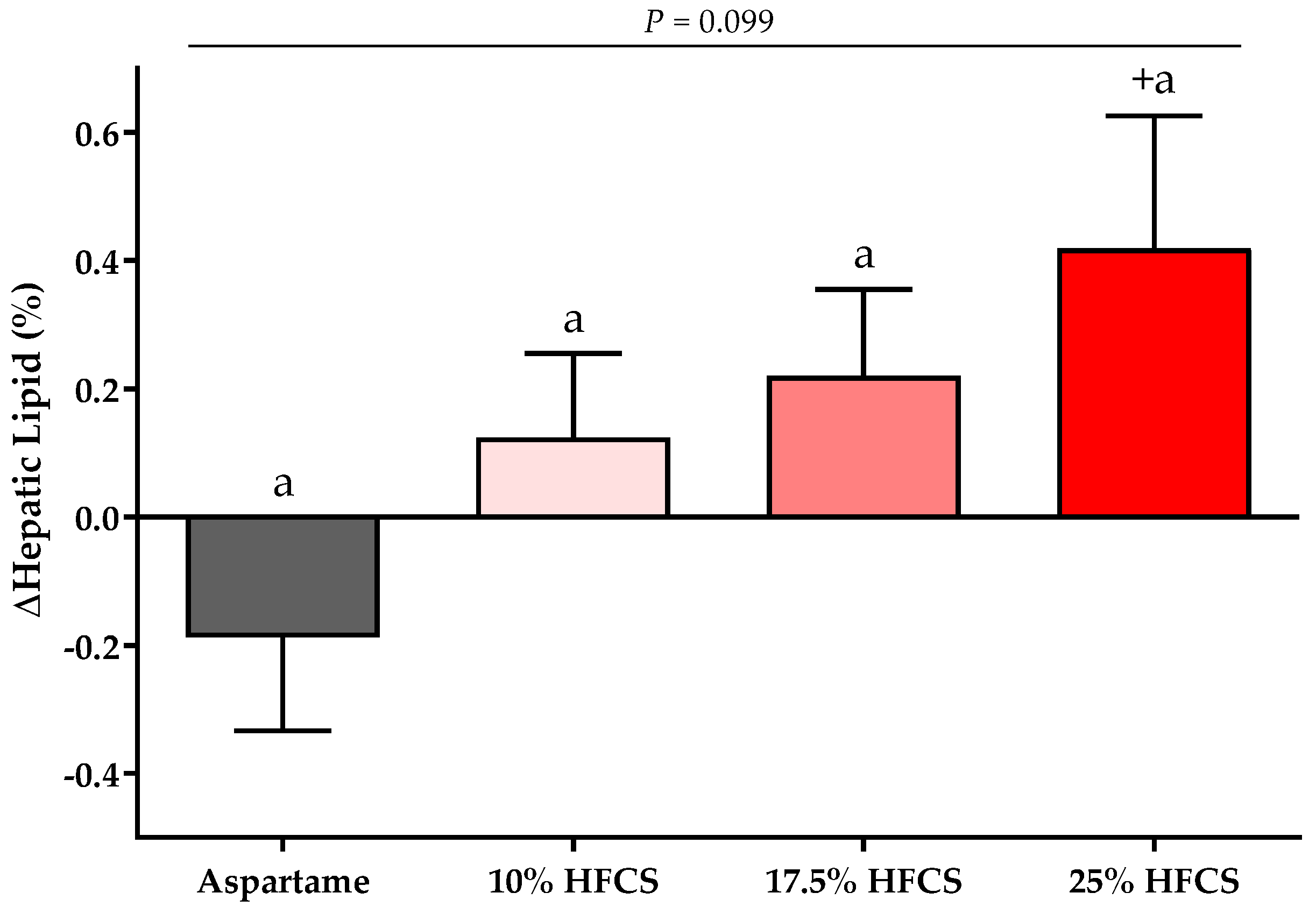
Figure 2.
Changes in hepatic lipid content: Mean ± SEM of the absolute change (intervention − baseline) of hepatic lipid content in participants consuming 0 (n = 20), 10 (n = 16), 17.5 (n = 16), or 25% (n = 23) HFCS-sweetened beverages for two weeks. p = the effect of HFCS group, two-factor (HFCS group, sex) ANCOVA with adjustment for MSRF and outcome at baseline; + p < 0.05, LS mean different from zero.
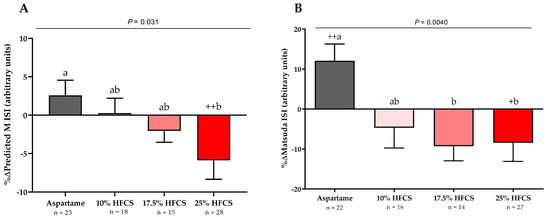
Figure 3.
Changes in Predicted M ISI and Matsuda ISI: Mean ± SEM of the % changes in (A) Predicted M Insulin Sensitivity Index (ISI) in participants consuming 0 (n = 22), 10 (n = 16), 17.5 (n = 14), or 25% (n = 27) and (B) Matsuda ISI in participants consuming 0 (n = 23), 10 (n = 18), 17.5 (n = 15), or 25% (n = 28) HFCS-sweetened beverages for two weeks. p = effect of HFCS group, two-factor (HFCS group, sex) ANCOVA with adjustment for MSRF; + p < 0.5, ++ p < 0.01, LS mean different from zero; a different from b, Tukey’s.
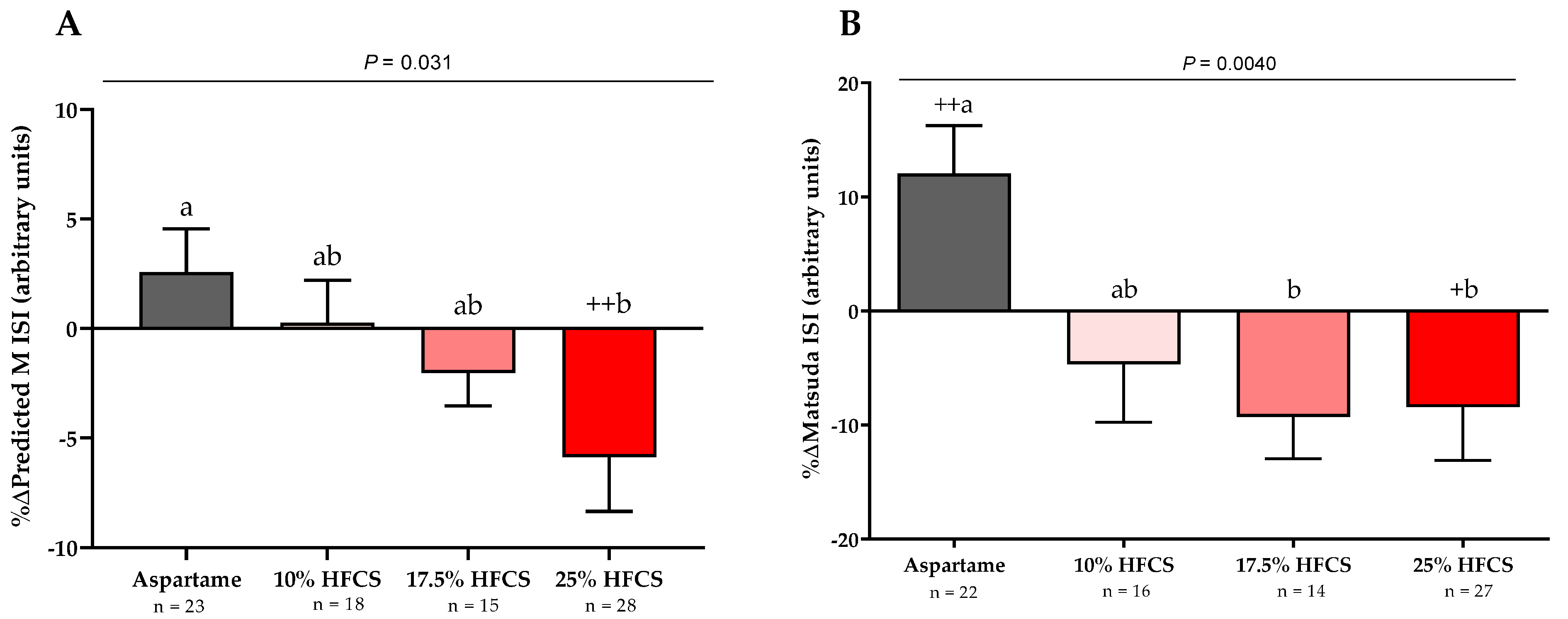
Figure 4.
Percent changes in 3-h glucose and insulin AUC: Mean ± SEM of the % change in glucose (A) and insulin (B) AUC during OGTT in participants consuming 0 (n = 23), 10 (n = 18), 17.5 (n = 15), or 25% (n = 28) HFCS-sweetened beverages for the two-week intervention. p = effect of HFCS group, two-factor (HFCS group, sex) ANCOVA with adjustment for MSRF; +p < 0.05, ++ p < 0.01, LS mean different from zero; a different from b, Tukey’s.
3.1. Hepatic Lipid Content (MRI-PDFF)
There was a significant linear dose-response effect on hepatic lipid content (p = 0.016). MSRF significantly affected hepatic lipid content (p = 0.033) (Table 2), with participants with a higher MSRF score exhibiting greater increases. As shown in Figure 2, hepatic lipid content was significantly increased in participants consuming 25% HFCS-SB compared with baseline (p = 0.031).
3.2. Insulin Sensitivity
The 3-h OGTT glucose and insulin responses at baseline and at the end of intervention are presented in Figure 5 and Figure 6. There were significant linear dose-response effects on the absolute changes in Matsuda ISI, Predicted M ISI, and on OGTT glucose and insulin excursions assessed as 3-h total AUC (Table 2). When analyzed as %Δ, the Predicted M ISI was significantly decreased in participants consuming 25% HFCS-SB compared with baseline (−5.9 ± 2.5%, p = 0.0036) and compared with participants consuming aspartame-SB (p = 0.022) (Figure 3A). Compared with baseline, the Matsuda ISI was significantly decreased in participants consuming 25% HFCS-SB (−8.4 ± 4.7%, p = 0.044) and significantly increased in participants consuming aspartame-SB (12.0 ± 4.2%, p = 0.0082; p = 0.0058, aspartame-SB vs. 25% HFCS-SB; p = 0.018, aspartame-SB vs. 17% HFCS-SB) (Figure 3B). OGTT glucose 3-h AUC (Figure 4A and Figure 5) increased significantly in participants consuming 17.5% (7.1 ± 2.6%, p = 0.023) and 25% HFCS-SB (6.1 ± 2.4%, p = 0.0062) compared with baseline and compared with participants consuming aspartame-SB (p = 0.031; p = 0.014, respectively). The OGTT insulin 3-h AUC (Figure 4B and Figure 6) was significantly increased in participants consuming 25% HFCS-SB compared with baseline (21.3 ± 5.9%, p = 0.0014), and compared with participants consuming aspartame-SB (p = 0.022). OGTT insulin 3-h AUC was also increased in participants consuming 10% HFCS-SB compared with those consuming aspartame-SB (p = 0.039). Fasting glucose and insulin concentrations (data not shown) and HOMA-IR (Table 2) were not significantly affected by HFCS dose or group.
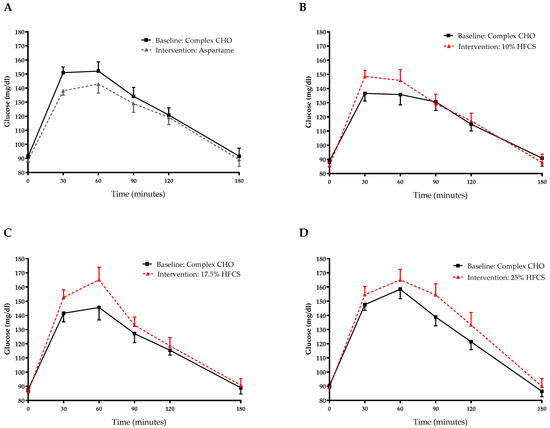
Figure 5.
Plasma glucose excursions during 3-h OGTT: Glucose concentrations during OGTT at baseline and after consuming 0 (n = 23) (A), 10 (n = 18) (B), 17.5 (n = 15) (C), or 25% (n = 28) (D) HFCS-sweetened beverages for the two-week intervention. CHO, carbohydrate; HFCS, high-fructose corn syrup.
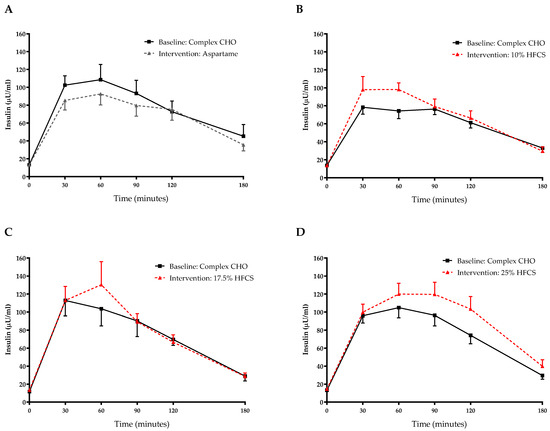
Figure 6.
Plasma insulin excursions during 3-h OGTT: Insulin concentrations during OGTT at baseline and after consuming 0 (n = 23) (A), 10 (n = 18) (B), 17.5 (n = 15) (C), or 25% (n = 28) (D) HFCS-sweetened beverages for the two-week intervention.
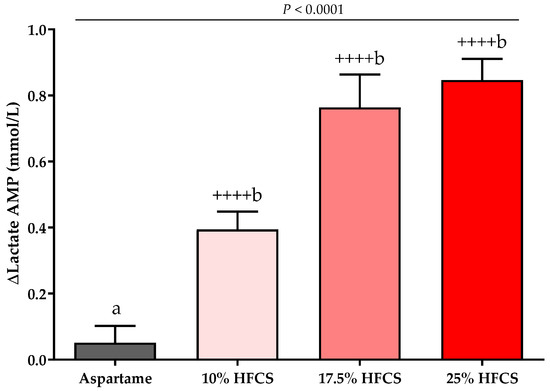
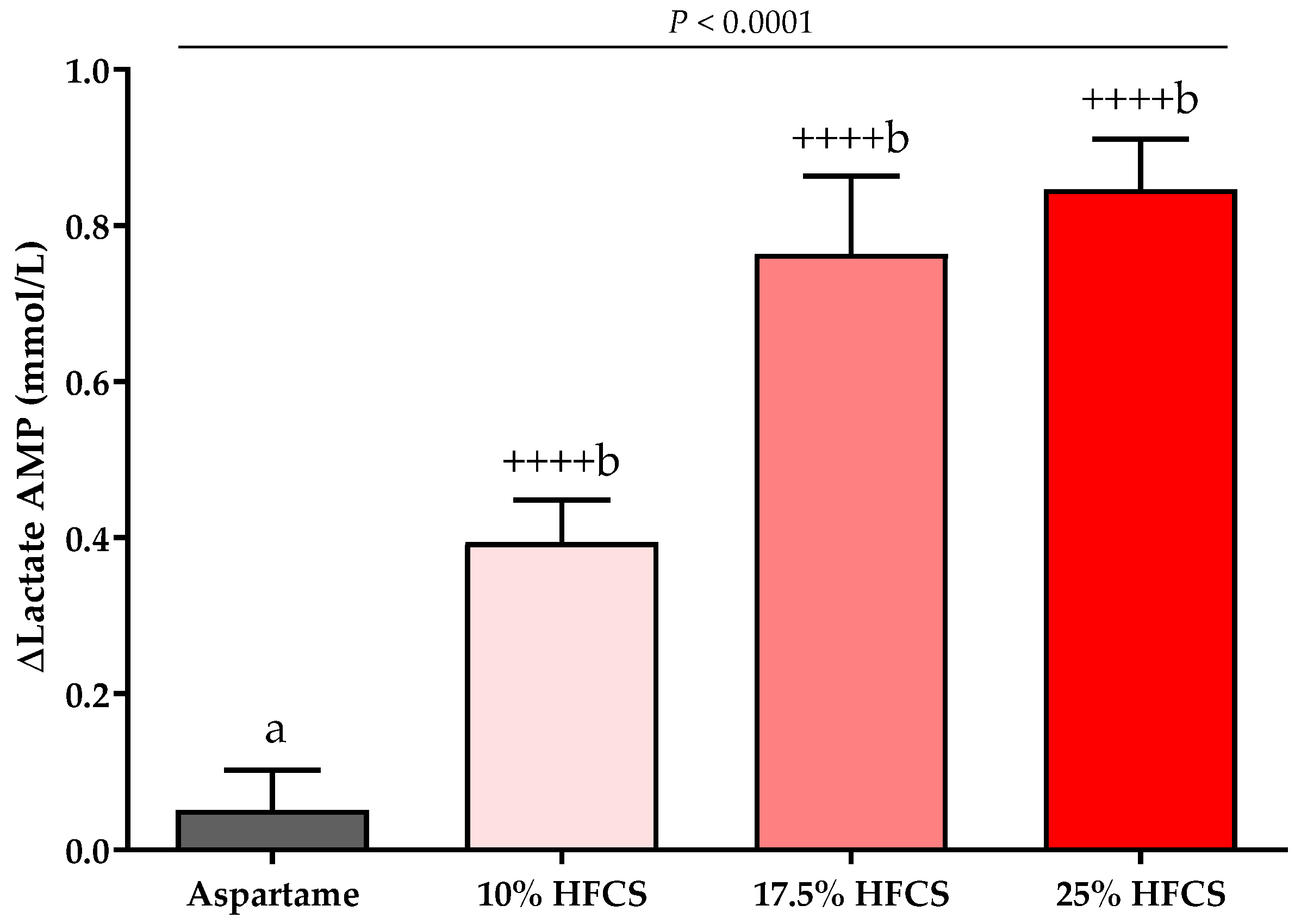
3.3. Post-Meal Lactate, Glucose and Insulin Concentrations
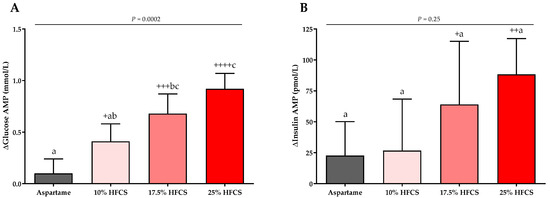
The baseline and intervention 24-h lactate, glucose, and insulin profiles are shown in the Supplement (Figures S1–S3). The profiles were quantified by both mean post-meal AMPs (Table 2, Figure 7 and Figure 8) and 24-h AUC (Supplement Table S1). Post-meal lactate AMPs were significantly affected by HFCS-dose (p < 0.0001, Table 2) and significantly increased in participants consuming 10, 17.5, or 25% HFCS-SB compared with baseline (all p < 0.0001) and compared with the aspartame-SB group (p = 0.0060, p = 0.0035, p < 0.0001, respectively) (Figure 7). The 24-h lactate AUCs were also significantly increased in participants consuming 17.5 or 25% HFCS-SB compared with those consuming aspartame-SB (Table S1). Post-meal glucose AMPs were significantly affected by HFCS-dose (p > 0.0001, Table 2), and significantly increased in participants consuming 10, 17.5, or 25% HFCS-SB compared with baseline (p = 0.014, p = 0.0002, p < 0.0001, respectively), and significantly increased in participants consuming 17.5 or 25% HFCS-SB compared with the aspartame-SB group (p = 0.044 and p < 0.0001, respectively) (Figure 8A). However, the post-meal glucose peaks following consumption of HFCS-SB were rapidly cleared such that the 24-h glucose AUCs remained unchanged in all groups (Supplement Table S1). As shown in Figure 8B, post-meal insulin AMPs were increased in participants consuming 17.5 or 25% HFCS-SB compared with baseline (p = 0.044 and p = 0.0029, respectively); however, there was no effect of HFCS-dose (p = 0.095, Table 2) or HFCS group (p = 0.25, Figure 8B).
Figure 7.
Changes in lactate amplitudes (AMP): Mean ± SEM of the absolute change (intervention − baseline) in lactate amplitudes in participants consuming 0 (n = 23), 10 (n = 18), 17.5 (n = 15), or 25% (n = 28) HFCS-sweetened beverages for the two-week intervention. p = the effect of SB group, two-factor (HFCS group, sex) ANCOVA with adjustment for MSRF and outcome at baseline; ++++ p < 0.0001, LS mean different from zero; a different from b, Tukey’s.
Figure 8.
Changes in glucose and insulin amplitudes (AMP): Mean ± SEM of the absolute changes (intervention − baseline) in glucose (A) and insulin AMPs (B) in participants consuming 0 (n = 23), 10 (n = 18), 17.5 (n = 15), or 25% (n = 28) HFCS-sweetened beverages for the two-week intervention. p = effect of HFCS group, two-factor (HFCS group, sex) ANCOVA with adjustment for MSRF and outcome at baseline; + p < 0.05, ++ p < 0.01, +++ p < 0.001, ++++ p < 0.0001, LS mean different from zero; a different from b, Tukey’s.
3.4. Mediation Analysis
3.4.1. Hepatic Lipid Content
The only index of ISI that was correlated with Δhepatic lipid content was the ΔOGTT insulin AUC (Table 3). As presented in Table 4, the mediation analysis suggested that ΔOGTT insulin AUC was a stronger partial mediator of Δhepatic lipid content than vice versa. The ΔOGTT insulin response attenuated the effect of HFCS dose on hepatic lipid content by 35% (from R = 0.26 to R = 0.17), while the effect of HFCS dose on ΔOGTT insulin AUC was attenuated by 15% and remained significant (p = 0.0068) when Δhepatic lipid content was controlled as the mediating variable.
Table 3.
The relationship between the absolute change of outcomes 1 significantly affected by dose 2.
Table 4.
Potential or partial mediator(s) of the effects of HFCS dose 1,2.
When ΔapoCIII was controlled as the mediating variable, the effect of HFCS dose on Δhepatic lipid content was attenuated by 81%. As shown in Table 4, Δ24-h uric acid AUC, Δpost-meal glucose AMP, and Δbody weight attenuated the effects of HFCS dose on hepatic lipid content by 67, 40, and 33%, respectively. The effects of the partial mediators (OGTT insulin AUC, 24-h uric acid AUC, post-meal glucose AMP, and Δbody weight) on hepatic lipid content were independent of each other and of apoCIII. A model that included HFCS dose, MSRF, gender, and all the mediators accounted for 44% of the variation, with apoCIII explaining 16.6%, MSRF explaining 6.6%, and OGTT insulin AUC, Δbody weight, post-meal glucose AMP, and 24-h uric acid AUC and HFCS dose contributing 3.7, 2.7, 2.6, 1.2, and 0%, respectively.
There was no evidence that hepatic lipid content mediated the effects of HFCS dose on the changes in circulating TG. Rather, when Δ24-h TG AUC and postprandial TG were controlled as mediators, the effect of HFCS dose on hepatic lipid content was attenuated by 53% and 64%, respectively. However, when Δpostprandial apoCIII was included as a mediating variable along with Δ24-h TG AUC or postprandial TG, apoCIII remained a significant mediator of the effects of HFCS dose on hepatic lipid content and 24-h TG AUC and postprandial TG had no significant effects.
3.4.2. Indices of Insulin Sensitivity
ΔPostprandial apoCIII was a partial mediator of ΔPredicted M ISI, attenuating the effect of HFCS dose by 48% (Table 4); however, it was not correlated with the other indices of insulin sensitivity. In contrast, the Δpost-meal lactate AMP was correlated with the changes in the Matsuda ISI and insulin and glucose OGTT AUC, but not Predicted M ISI. The Δpost-meal lactate AMP attenuated the effects of HFCS dose on ΔMatsuda ISI by 71% and partially attenuated the effect of HFCS dose on Δinsulin and glucose OGTT by 38% and 36% AUC, respectively.
4. Discussion
Insulin resistance is considered a central [72] or pivotal pathogenic component [73] of T2D and metabolic syndrome. NAFLD, diagnosed as hepatic lipid content exceeding 5% [74], has been described as the liver’s manifestation of metabolic syndrome [75,76]. This study provides novel results supporting our hypothesis that consumption of HFCS-SB at 10, 17.5, or 25% of Ereq increases hepatic lipid content and decreases insulin sensitivity in a linear dose-dependent manner. Demonstration of a dose-response relationship, also termed biological gradient, is considered strong evidence for a causal relationship between the exposure and the outcome [77,78]. The current results complement our previous reported results from the same participants showing dose-dependent increases in uric acid, triglyceride, apoCIII, LDL-C, and apoB [56].
A linear dose-response relationship between sugar-SB consumption and risk of NAFLD has been documented in observational studies [21]. A meta-analysis of twelve studies (case-control, cross-sectional or cohort studies), which included a total of 35,705 participants, showed consumptions of low doses (<1 cup/week), middle doses (1–6 cups/week), and high doses (≥7 cups/week) of sugar-SB increased the relative risk (RR) of NAFLD by 14%, 26%, and 53%, respectively (p = 0.01, p < 0.00001, p = 0.03, respectively) [21]. Similarly, a meta-analysis of nineteen prospective studies representing over a million participants showed a linear dose-response relationship between sugar-SB consumption and risk of T2D. The RR of T2D was 1.19 (95% CI 1.13–1.25) for each 250 mL/day of sugar-SB consumed [20]. Another recent meta-analysis reported linear dose-response relationships between sugar-SB and CVD incidence (RR 1.08, 95% CI 1.02 to 1.14 for each 250 mL/day) and risk of coronary heart disease (RR 1.15, 95% CI 1.09 to 1.22 for each 250 mL/day) [79]. Thus, the evidence from the observational studies [20,21,79] complements our current and previous results [56] and results from previous dietary studies [22,23,24,25,26,27,28,29,30,31,32,80], and together they provide a strong scientific foundation [81] to support the conclusion that excess sugar-SB consumption increases the risk of T2D, NAFLD, CVD, and metabolic syndrome.
This conclusion is further strengthened by the results from the first randomized controlled phase 2 clinical trial testing the administration of a fructokinase inhibitor in participants with NAFLD [82]. Relative to baseline, the fructokinase inhibitor (PF-06835919) led to a 26.5% reduction in liver fat, an 11.5% reduction in fasting uric acid, and a trend for improvements of other cardiometabolic parameters, including insulin resistance and inflammation [82]. This study documents the mechanistic role of fructokinase in mediating the unregulated hepatic fructose uptake and overload that leads to metabolic dysregulation. It is also worth noting that the favorable effects of the fructokinase inhibitor were accompanied by a small (<1 kg), but significant, increase of body weight [82]. This is in contrast to dietary intervention trials in which increases or decreases in fructose/sugar intake often induce parallel changes in risk factors and body weight, thus making it difficult to differentiate direct metabolic effects of sugar from those that are mediated by changes in body weight. However, in this study, preventing hepatic fructose overload via inhibition of fructokinase led to decreased liver lipid content and favorable metabolic effects that are clearly not confounded by favorable effects on body weight. This evidence [82] refutes the contention that dietary sugars are purely a highly palatable source of energy that have no unique or detrimental impact relative to any other source of calories [83].
We conducted simple mediation analyses to statistically test whether our data are indicative that the effects of HFCS dose on insulin sensitivity and circulating TG are mediated by the changes in hepatic lipid content. The results of these analyses did not support such a relationship with either outcome. Instead, with respect to insulin sensitivity, the increase in OGTT insulin AUC partially mediated the effects of HFCS dose on hepatic lipid content. This direction of partial mediation is plausible in that the hyperinsulinemia, which compensates for insulin resistance, activates hepatic lipid synthesis by increasing the activity of lipogenic enzymes [46,84]. Additionally, our data do not support previous work suggesting that hepatic lipid content regulates VLDL and TG production and secretion [42]. While the changes in hepatic lipid content were positively associated with the changes in circulating TG (24-h AUC and postprandial), there was no evidence that hepatic lipid content mediated the effects of HFCS dose on these outcomes.
The mediation analyses suggested that, in addition to OGTT insulin AUC, other potential mediators or partial mediators are implicated in the effects of HFCS dose on hepatic lipid content. ApoCIII was the strongest mediator of the change of hepatic lipid, attenuating the effect of HFCS dose by 81%. This relationship could be mediated directly, as it has been shown that an apoCIII gain-of-function variant enhances DNL and hepatic triglyceride-rich lipoprotein production [85], and overexpression of apoCIII increases hepatic lipid content in mice consuming low fat diets [86]. However, it is also possible that this relationship reflects the known effects of fructose (and glucose to a lesser extent) to upregulate both SREBP-1c and ChREBP [37,38]. These transcription factors increase both DNL and apoCIII production [37,38,39], thus, it may be enhanced lipid synthesis via DNL that mediates the effects of HFCS on hepatic lipid content. However, since DNL was not measured in the majority of the participants who participated in this study, we were unable to assess whether DNL mediated the effects of HFCS dose on hepatic lipid. The effect of HFCS dose on hepatic lipid content were also partially mediated by the changes in 24-h uric acid concentrations, post-meal glucose AMPs, and body weight. Experimental evidence suggests that uric acid can contribute to hepatic lipid deposition through induction of mitochondrial oxidative stress and increased expression of fructokinase [53,54,55], whereas evidence to suggest increased post-meal glucose AMPs promote accumulation of liver fat is lacking.
While it has been suggested that sustained increases in circulating TG may increase muscle lipid accumulation leading to impaired muscle insulin action and lowered whole body insulin sensitivity [47], we observed no associations between the indices of insulin sensitivity and circulating TG. Instead, the mediation analysis showed that post-meal lactate AMP attenuated the effects of HFCS on Matsuda ISI by 71%. Studies in rats in which lactate infusion inhibited glucose transport in skeletal muscle [51] and suppressed glycolysis and impaired insulin signaling [87] suggest potential mechanisms. Postprandial apoCIII attenuated the effect of HFCS dose on Predict ISI by 48%, possibly by inducing inflammation and inducing ER stress in muscle [88,89].
This is the first study to demonstrate that young men and women consuming HFCS-SB at 10, 17.5, or 25% of Ereq for two weeks exhibited dose-dependent increases in hepatic lipid content and dose-dependent decreases in insulin sensitivity. The only other published study that has examined the dose-dependent effects of sugar-SB consumption on hepatic lipid content and insulin sensitivity was funded by the Corn Refiners Association and generated null findings for both outcomes [90,91]. Reasons for the discrepant results have been previously discussed, both specifically [22,56,92] and generally [93].
The 2015–2020 Dietary Guidelines for America (DGA) recommends less than 10% of daily calories be consumed as added sugar [94]. The USDA 2019 loss-adjusted per capita caloric sweetener consumption is 344 kcal/day [13]. This suggests that the great majority of Americans are exceeding the Dietary Guideline’s recommendation for added sugars [94]. However, 344 kcal/day may underestimate the actual per capita intake because it is calculated based on the assumption that 34% of the caloric sweetener to which consumers are exposed is lost to spoilage, cooking loss, or plate waste. There is no question that it is difficult to accurately estimate food losses after the point of purchase. However, when comparing the spoilage potential, shelf life, and consumer preference of high sugar foods to that of fresh fruit, fresh vegetable, meat, and dairy products, it would seem intuitive that consumer losses of caloric sweeteners would be relatively lower. However, to obtain the USDA per capita consumption estimates, these are the corrections that are applied to various food groups to account for foods not consumed after the point of purchase; caloric sweeteners: 34%; fresh fruits and vegetables: 27%; dairy products: 23%; beef, pork, and lamb: 24%; fats and oils: 21%; grain products: 20% [13].
While the average consumption of sugar-SBs in the U.S. has decreased over the past ten years, consumption amounts vary greatly throughout the population. Between 2011 and 2014, it was found that more than two thirds of youths and one half of adults consume at least one sugar-SB on a given day, and at least 10% of youths and adults consumed at least two sugar-SB on a given day [17,18]. With a disproportionate number of heavy consumers being racial minorities from disadvantaged socioeconomic backgrounds, our results not only support dietary recommendations, but guide public health policies directed at decreasing health disparities [95].
Study Strengths and Limitations
The use of an advanced MRI technique, providing a non-invasive and standardized quantitative biomarker suitable as a substitute for liver biopsy, for the quantification of hepatic lipid content is considered a study strength [96]. The presence of a biomarker in the study beverages as an objective measure of compliance is another strength of the study. Participants were informed of the biomarker and this awareness possibly contributed to the high degree of compliance evidenced by the consistent dose-response trends in this report and our previous publication [56]. In addition, conducting the baseline and intervention experimental procedures while subjects resided at the CCRC for 3.5 days and consumed standardized diets minimized the variability in the study results that can occur in outpatient settings due to non-compliance and differences in diet and physical activity.
A limitation of the study is that it was not randomized, which could have potentially introduced bias in the allocation of participants to the experimental groups. During the 12-day outpatient period, participants consumed the study beverages with ad libitum diets, therefore, the total amount of sugar consumed during this period cannot be accurately quantified. While the duration of the two-week intervention is relatively short, it demonstrates just how quickly excess sugar consumption can increase risk factors for cardiometabolic disease. A limitation of the mediation analyses is the smaller group sample sizes due to exclusion of participants with missing outcomes.
5. Conclusions
This study demonstrates that two weeks of consumption of HFCS-sweetened beverages providing 10, 17.5, or 25% Ereq results in dose-dependent increases in hepatic lipid content and dose-dependent decreases in insulin sensitivity in young men and women. These results contribute to the strong body of epidemiological, dietary intervention and mechanistic evidence that increased consumption of sugar-SB heightens risk for NAFLD and T2D.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/nu14081648/s1, Table S1: Twenty-four-hour glucose, insulin and lactate AUCs, Figure S1: Twenty-four hour plasma lactate concentrations, Figure S2: Twenty-four hour plasma glucose concentrations, Figure S3: Twenty-four hour plasma insulin concentration.
Author Contributions
K.L.S., N.L.K., P.J.H. were responsible for the conceptualization and experimental design; M.V.N., V.L., D.M.S., B.H., C.A.P., A.A.B., A.J.C., Y.A., J.P.M., M.I.G., C.B.S., V.M., K.L.S. were responsible for study investigation; data curation was carried out by V.L., M.V.N., C.L.C., Y.B., B.H., G.P., A.T., D.M.S., K.L.S.; D.M.S., K.L.S. prepared the original draft. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the NIH/National Heart, Lung and Blood Institute 1R01 HL09133 and 1R01 HL107256; National Center for Research Resources, a component of the NIH, and NIH Roadmap for Medical Research UL1 RR024146; a Building Interdisciplinary Research Careers in Women’s Health award (K12 HD051958), funded by the National Institute of Child Health and Human Development, Office of Research on Women’s Health, Office of Dietary Supplements, and the National Institute of Aging (to K.L.S); and intramural USDA-ARS CRIS 5306-51530-022-00D (to N.L.K). D.M.S was supported by diversity supplement from the NIH/National Heart, Lung and Blood Institute: RO1-HL-121324 and RO1-HL-137716. B.H. was supported by a research fellowship from the German Research Foundation (Deutsche Forschungsgemeinschaft) HI 2113/1-1.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Institutional Review Board (or Ethics Committee) of the University of California, Davis (protocol code 200715772-1, approved on 7 November 2008).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Restrictions apply to the availability of some or all data generated or analyzed during this study to preserve subject confidentially. The corresponding authors will on request detail the restrictions and any conditions under which access to some data may be provided.
Acknowledgments
We thank James Graham (UC Davis) for excellent technical support, Janet Peerson (UC Davis) for expert guidance on the statistical analyses and our UC Davis undergraduate student interns and employees for assistance with meal and blood processing. Additionally, we thank the nursing staff at the CCRC for their dedicated nursing support.
Conflicts of Interest
V.M. serves in the Advisory board of Alexion Pharmaceuticals. M.I.G. serves as Scientific Advisor to Yumi infant food and receives royalties for Sugarproof published by Penguin Random House. C.S. reports grants from GE, Siemens, Philips, Bayer, Foundation of NIH, Gilead, and Pfizer (grant is to UW-Madison; UCSD is a subcontract to UW-Madison); personal consultation fees from Blade, Boehringer, and Epigenomics; consultation under the auspices of the University to AMRA, BMS, Exact Sciences, GE Digital, IBM-Watson, and Pfizer; lab service agreements from Enanta, Gilead, ICON, Intercept, Nusirt, Shire, Synageva, Takeda; royalties from Wolters Kluwer for educational material outside the submitted work; honoraria to the institution from Medscape for educational material outside the submitted work; ownership of stock options in Livivos; unpaid position in advisory board to Quantix Bio. All other authors have no conflict of interest to declare.
References
- Interntional Diabetes Federation. IDF Diabetes Atlas: 10th Edition. Available online: http://www.diabetesatlas.org (accessed on 15 November 2021).
- Zimmet, P.; Alberti, K.G.; Shaw, J. Global and societal implications of the diabetes epidemic. Nature 2001, 414, 782–787. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zou, B.; Yeo, Y.H.; Feng, Y.; Xie, X.; Lee, D.H.; Fujii, H.; Wu, Y.; Kam, L.Y.; Ji, F.; et al. Prevalence, incidence, and outcome of non-alcoholic fatty liver disease in Asia, 1999–2019: A systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 2019, 4, 389–398. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciardullo, S.; Perseghin, G. Prevalence of NAFLD, MAFLD and associated advanced fibrosis in the contemporary United States population. Liver Int. 2021, 41, 1290–1293. [Google Scholar] [CrossRef]
- Caussy, C.; Aubin, A.; Loomba, R. The relationship between type 2 diabetes, NAFLD, and cardiovascular risk. Curr. Diabetes Rep. 2021, 21, 15. [Google Scholar] [CrossRef]
- Byrne, C.D.; Targher, G. NAFLD: A multisystem disease. J. Hepatol. 2015, 62, S47–S64. [Google Scholar] [CrossRef] [Green Version]
- Younossi, Z.M.; Golabi, P.; de Avila, L.; Paik, J.M.; Srishord, M.; Fukui, N.; Qiu, Y.; Burns, L.; Afendy, A.; Nader, F. The global epidemiology of NAFLD and NASH in patients with type 2 diabetes: A systematic review and meta-analysis. J. Hepatol. 2019, 71, 793–801. [Google Scholar] [CrossRef]
- Anstee, Q.M.; Targher, G.; Day, C.P. Progression of NAFLD to diabetes mellitus, cardiovascular disease or cirrhosis. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 330–344. [Google Scholar] [CrossRef]
- Softic, S.; Stanhope, K.L.; Boucher, J.; Divanovic, S.; Lanaspa, M.A.; Johnson, R.J.; Kahn, C.R. Fructose and hepatic insulin resistance. Crit. Rev. Clin. Lab. Sci. 2020, 57, 308–322. [Google Scholar] [CrossRef]
- Zheng, Y.; Ley, S.H.; Hu, F.B. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol. 2018, 14, 88–98. [Google Scholar] [CrossRef]
- Loomba, R.; Sanyal, A.J. The global NAFLD epidemic. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 686–690. [Google Scholar] [CrossRef] [PubMed]
- The USDA's Economic Research Service (ERS). Loss-Adjusted Food Availability Documentation. Available online: https://www.ers.usda.gov/data-products/food-availability-per-capita-data-system/loss-adjusted-food-availability-documentation/ (accessed on 20 January 2022).
- Popkin, B.M.; Adair, L.S.; Ng, S.W. Global nutrition transition and the pandemic of obesity in developing countries. Nutr. Rev. 2012, 70, 3–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez Steele, E.; Baraldi, L.G.; Louzada, M.L.; Moubarac, J.C.; Mozaffarian, D.; Monteiro, C.A. Ultra-processed foods and added sugars in the US diet: Evidence from a nationally representative cross-sectional study. BMJ Open 2016, 6, e009892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neri, D.; Martinez-Steele, E.; Monteiro, C.A.; Levy, R.B. Consumption of ultra-processed foods and its association with added sugar content in the diets of US children, NHANES 2009–2014. Pediatr. Obes. 2019, 14, e12563. [Google Scholar] [CrossRef] [PubMed]
- Rosinger, A.; Herrick, K.; Gahche, J.; Park, S. Sugar-Sweetened Beverage Consumption among U.S. Adults, 2011–2014; NCHS data brief, no 270; National Center for Health Statistics: Hyattsville, MD, USA, 2017.
- Rosinger, A.; Herrick, K.; Gahche, J.; Park, S. Sugar-Sweetened Beverage Consumption among U.S. Youth, 2011–2014; NCHS data brief, no 271; National Center for Health Statistics: Hyattsville, MD, USA, 2017.
- Ma, J.; Jacques, P.F.; Meigs, J.B.; Fox, C.S.; Rogers, G.T.; Smith, C.E.; Hruby, A.; Saltzman, E.; McKeown, N.M. Sugar-sweetened beverage but not diet soda consumption is positively associated with progression of insulin resistance and prediabetes. J. Nutr. 2016, 146, 2544–2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, P.; Li, Q.; Zhao, Y.; Chen, Q.; Sun, X.; Liu, Y.; Li, H.; Wang, T.; Chen, X.; Zhou, Q.; et al. Sugar and artificially sweetened beverages and risk of obesity, type 2 diabetes mellitus, hypertension, and all-cause mortality: A dose-response meta-analysis of prospective cohort studies. Eur. J. Epidemiol. 2020, 35, 655–671. [Google Scholar] [CrossRef]
- Chen, H.; Wang, J.; Li, Z.; Lam, C.W.K.; Xiao, Y.; Wu, Q.; Zhang, W. Consumption of sugar-sweetened beverages has a dose-dependent effect on the risk of non-alcoholic fatty liver disease: An updated systematic review and dose-response meta-analysis. Int. J. Environ. Res. Public Health 2019, 16, 2192. [Google Scholar] [CrossRef] [Green Version]
- Sigala, D.M.; Hieronimus, B.; Medici, V.; Lee, V.; Nunez, M.V.; Bremer, A.A.; Cox, C.L.; Price, C.A.; Benyam, Y.; Chaudhari, A.J.; et al. Consuming sucrose- or HFCS-sweetened beverages increases hepatic lipid and decreases insulin sensitivity in adults. J. Clin. Endocrinol. Metab. 2021, 106, 3248–3264. [Google Scholar] [CrossRef]
- Maersk, M.; Belza, A.; Stodkilde-Jorgensen, H.; Ringgaard, S.; Chabanova, E.; Thomsen, H.; Pedersen, S.B.; Astrup, A.; Richelsen, B. Sucrose-sweetened beverages increase fat storage in the liver, muscle, and visceral fat depot: A 6-mo randomized intervention study. Am. J. Clin. Nutr. 2012, 95, 283–289. [Google Scholar] [CrossRef]
- Schwarz, J.M.; Noworolski, S.M.; Wen, M.J.; Dyachenko, A.; Prior, J.L.; Weinberg, M.E.; Herraiz, L.A.; Tai, V.W.; Bergeron, N.; Bersot, T.P.; et al. Effect of a high-fructose weight-maintaining diet on lipogenesis and liver fat. J. Clin. Endocrinol. Metab. 2015, 100, 2434–2442. [Google Scholar] [CrossRef]
- Taskinen, M.R.; Soderlund, S.; Bogl, L.H.; Hakkarainen, A.; Matikainen, N.; Pietilainen, K.H.; Rasanen, S.; Lundbom, N.; Bjornson, E.; Eliasson, B.; et al. Adverse effects of fructose on cardiometabolic risk factors and hepatic lipid metabolism in subjects with abdominal obesity. J. Intern. Med. 2017, 282, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Stanhope, K.L.; Schwarz, J.M.; Keim, N.L.; Griffen, S.C.; Bremer, A.A.; Graham, J.L.; Hatcher, B.; Cox, C.L.; Dyachenko, A.; Zhang, W.; et al. Consuming fructose-sweetened, not glucose-sweetened, beverages increases visceral adiposity and lipids and decreases insulin sensitivity in overweight/obese humans. J. Clin. Investig. 2009, 119, 1322–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aeberli, I.; Hochuli, M.; Gerber, P.A.; Sze, L.; Murer, S.B.; Tappy, L.; Spinas, G.A.; Berneis, K. Moderate amounts of fructose consumption impair insulin sensitivity in healthy young men: A randomized controlled trial. Diabetes Care 2013, 36, 150–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, V.; Despland, C.; Brandejsky, V.; Kreis, R.; Schneiter, P.; Chiolero, A.; Boesch, C.; Tappy, L. Sugar- and artificially sweetened beverages and intrahepatic fat: A randomized controlled trial. Obesity 2015, 23, 2335–2339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarra-Reynoso, L.D.R.; Lopez-Lemus, H.L.; Garay-Sevilla, M.E.; Malacara, J.M. Effect of restriction of foods with high fructose corn syrup content on metabolic indices and fatty liver in obese children. Obes. Facts 2017, 10, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Schwimmer, J.B.; Ugalde-Nicalo, P.; Welsh, J.A.; Angeles, J.E.; Cordero, M.; Harlow, K.E.; Alazraki, A.; Durelle, J.; Knight-Scott, J.; Newton, K.P.; et al. Effect of a low free sugar diet vs. usual diet on nonalcoholic fatty liver disease in adolescent boys: A randomized clinical trial. JAMA 2019, 321, 256–265. [Google Scholar] [CrossRef] [Green Version]
- Simons, N.; Veeraiah, P.; Simons, P.; Schaper, N.C.; Kooi, M.E.; Schrauwen-Hinderling, V.B.; Feskens, E.J.M.; van der Ploeg, E.; Van den Eynde, M.D.G.; Schalkwijk, C.G.; et al. Effects of fructose restriction on liver steatosis (FRUITLESS); a double-blind randomized controlled trial. Am. J. Clin. Nutr. 2021, 113, 391–400. [Google Scholar] [CrossRef]
- Schwarz, J.M.; Noworolski, S.M.; Erkin-Cakmak, A.; Korn, N.J.; Wen, M.J.; Tai, V.W.; Jones, G.M.; Palii, S.P.; Velasco-Alin, M.; Pan, K.; et al. Effects of dietary fructose restriction on liver fat, de novo lipogenesis, and insulin kinetics in children with obesity. Gastroenterology 2017, 153, 743–752. [Google Scholar] [CrossRef] [Green Version]
- Ishimoto, T.; Lanaspa, M.A.; Le, M.T.; Garcia, G.E.; Diggle, C.P.; Maclean, P.S.; Jackman, M.R.; Asipu, A.; Roncal-Jimenez, C.A.; Kosugi, T.; et al. Opposing effects of fructokinase C and A isoforms on fructose-induced metabolic syndrome in mice. Proc. Natl. Acad. Sci. USA 2012, 109, 4320–4325. [Google Scholar] [CrossRef] [Green Version]
- Softic, S.; Gupta, M.K.; Wang, G.X.; Fujisaka, S.; O’Neill, B.T.; Rao, T.N.; Willoughby, J.; Harbison, C.; Fitzgerald, K.; Ilkayeva, O.; et al. Divergent effects of glucose and fructose on hepatic lipogenesis and insulin signaling. J. Clin. Investig. 2017, 127, 4059–4074. [Google Scholar] [CrossRef] [Green Version]
- Francey, C.; Cros, J.; Rosset, R.; Creze, C.; Rey, V.; Stefanoni, N.; Schneiter, P.; Tappy, L.; Seyssel, K. The extra-splanchnic fructose escape after ingestion of a fructose-glucose drink: An exploratory study in healthy humans using a dual fructose isotope method. Clin. Nutr. ESPEN 2019, 29, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Havel, P.J. Dietary fructose: Implications for dysregulation of energy homeostasis and lipid/carbohydrate metabolism. Nutr. Rev. 2005, 63, 133–157. [Google Scholar] [CrossRef] [PubMed]
- Softic, S.; Cohen, D.E.; Kahn, C.R. Role of dietary fructose and hepatic de novo lipogenesis in fatty liver disease. Dig. Dis. Sci. 2016, 61, 1282–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, M.A.; Birnbaum, M.J. Molecular aspects of fructose metabolism and metabolic disease. Cell Metab. 2021, 33, 2329–2354. [Google Scholar] [CrossRef]
- Hieronimus, B.; Stanhope, K.L. Dietary fructose and dyslipidemia: New mechanisms involving apolipoprotein CIII. Curr. Opin. Lipidol. 2020, 31, 20–26. [Google Scholar] [CrossRef]
- Cox, C.L.; Stanhope, K.L.; Schwarz, J.M.; Graham, J.L.; Hatcher, B.; Griffen, S.C.; Bremer, A.A.; Berglund, L.; McGahan, J.P.; Havel, P.J.; et al. Consumption of fructose-sweetened beverages for 10 weeks reduces net fat oxidation and energy expenditure in overweight/obese men and women. Eur. J. Clin. Nutr. 2012, 66, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Jornayvaz, F.R.; Shulman, G.I. Diacylglycerol activation of protein kinase Cepsilon and hepatic insulin resistance. Cell Metab. 2012, 15, 574–584. [Google Scholar] [CrossRef] [Green Version]
- Adiels, M.; Taskinen, M.R.; Packard, C.; Caslake, M.J.; Soro-Paavonen, A.; Westerbacka, J.; Vehkavaara, S.; Hakkinen, A.; Olofsson, S.O.; Yki-Jarvinen, H.; et al. Overproduction of large VLDL particles is driven by increased liver fat content in man. Diabetologia 2006, 49, 755–765. [Google Scholar] [CrossRef] [Green Version]
- Lewis, G.F.; Carpentier, A.; Adeli, K.; Giacca, A. Disordered fat storage and mobilization in the pathogenesis of insulin resistance and type 2 diabetes. Endocr. Rev. 2002, 23, 201–229. [Google Scholar] [CrossRef]
- Boren, J.; Packard, C.J.; Taskinen, M.R. The roles of ApoC-III on the metabolism of triglyceride-rich lipoproteins in humans. Front. Endocrinol. 2020, 11, 474. [Google Scholar] [CrossRef]
- Petersen, M.C.; Shulman, G.I. Mechanisms of insulin action and insulin resistance. Physiol. Rev. 2018, 98, 2133–2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, J.R.; Langlet, F.; Kido, Y.; Accili, D. Pathogenesis of selective insulin resistance in isolated hepatocytes. J. Biol. Chem. 2015, 290, 13972–13980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krssak, M.; Falk Petersen, K.; Dresner, A.; DiPietro, L.; Vogel, S.M.; Rothman, D.L.; Roden, M.; Shulman, G.I. Intramyocellular lipid concentrations are correlated with insulin sensitivity in humans: A 1H NMR spectroscopy study. Diabetologia 1999, 42, 113–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baena, M.; Sanguesa, G.; Davalos, A.; Latasa, M.J.; Sala-Vila, A.; Sanchez, R.M.; Roglans, N.; Laguna, J.C.; Alegret, M. Fructose, but not glucose, impairs insulin signaling in the three major insulin-sensitive tissues. Sci. Rep. 2016, 6, 26149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyssel, K.; Meugnier, E.; Le, K.A.; Durand, C.; Disse, E.; Blond, E.; Pays, L.; Nataf, S.; Brozek, J.; Vidal, H.; et al. Fructose overfeeding in first-degree relatives of type 2 diabetic patients impacts energy metabolism and mitochondrial functions in skeletal muscle. Mol. Nutr. Food Res. 2016, 60, 2691–2699. [Google Scholar] [CrossRef]
- Tappy, L. Fructose metabolism and noncommunicable diseases: Recent findings and new research perspectives. Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 214–222. [Google Scholar] [CrossRef]
- Lombardi, A.M.; Fabris, R.; Bassetto, F.; Serra, R.; Leturque, A.; Federspil, G.; Girard, J.; Vettor, R. Hyperlactatemia reduces muscle glucose uptake and GLUT-4 mRNA while increasing (E1alpha)PDH gene expression in rat. Am. J. Physiol. 1999, 276, E922–E929. [Google Scholar] [CrossRef]
- King, C.; Lanaspa, M.A.; Jensen, T.; Tolan, D.R.; Sanchez-Lozada, L.G.; Johnson, R.J. Uric acid as a cause of the metabolic syndrome. In Uric Acid in Chronic Kidney Disease; Treviño-Becerra, A., Iseki, K., Eds.; Contributions to Nephrology; Karger: Basel, Switzerland, 2018; Volume 192, pp. 88–102. [Google Scholar] [CrossRef]
- Lanaspa, M.A.; Sanchez-Lozada, L.G.; Cicerchi, C.; Li, N.; Roncal-Jimenez, C.A.; Ishimoto, T.; Le, M.; Garcia, G.E.; Thomas, J.B.; Rivard, C.J.; et al. Uric acid stimulates fructokinase and accelerates fructose metabolism in the development of fatty liver. PLoS ONE 2012, 7, e47948. [Google Scholar] [CrossRef] [Green Version]
- Lanaspa, M.A.; Sanchez-Lozada, L.G.; Choi, Y.J.; Cicerchi, C.; Kanbay, M.; Roncal-Jimenez, C.A.; Ishimoto, T.; Li, N.; Marek, G.; Duranay, M.; et al. Uric acid induces hepatic steatosis by generation of mitochondrial oxidative stress: Potential role in fructose-dependent and -independent fatty liver. J. Biol. Chem. 2012, 287, 40732–40744. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.J.; Nakagawa, T.; Sanchez-Lozada, L.G.; Shafiu, M.; Sundaram, S.; Le, M.; Ishimoto, T.; Sautin, Y.Y.; Lanaspa, M.A. Sugar, uric acid, and the etiology of diabetes and obesity. Diabetes 2013, 62, 3307–3315. [Google Scholar] [CrossRef] [Green Version]
- Stanhope, K.L.; Medici, V.; Bremer, A.A.; Lee, V.; Lam, H.D.; Nunez, M.V.; Chen, G.X.; Keim, N.L.; Havel, P.J. A dose-response study of consuming high-fructose corn syrup-sweetened beverages on lipid/lipoprotein risk factors for cardiovascular disease in young adults. Am. J. Clin. Nutr. 2015, 101, 1144–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigala, D.M.; Widaman, A.M.; Hieronimus, B.; Nunez, M.V.; Lee, V.; Benyam, Y.; Bremer, A.A.; Medici, V.; Havel, P.J.; Stanhope, K.L.; et al. Effects of consuming sugar-sweetened beverages for 2 weeks on 24-h circulating leptin profiles, ad libitum food intake and body weight in young adults. Nutrients 2020, 12, 3893. [Google Scholar] [CrossRef] [PubMed]
- Mifflin, M.D.; St Jeor, S.T.; Hill, L.A.; Scott, B.J.; Daugherty, S.A.; Koh, Y.O. A new predictive equation for resting energy expenditure in healthy individuals. Am. J. Clin. Nutr. 1990, 51, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Hieronimus, B.; Medici, V.; Bremer, A.A.; Lee, V.; Nunez, M.V.; Sigala, D.M.; Keim, N.L.; Havel, P.J.; Stanhope, K.L. Synergistic effects of fructose and glucose on lipoprotein risk factors for cardiovascular disease in young adults. Metabolism 2020, 112, 154356. [Google Scholar] [CrossRef]
- Reeder, S.B.; Sirlin, C.B. Quantification of liver fat with magnetic resonance imaging. Magn. Reson. Imaging Clin. N. Am. 2010, 18, 337–357. [Google Scholar] [CrossRef] [Green Version]
- Yokoo, T.; Shiehmorteza, M.; Bydder, M.; Hamilton, G.; Kono, Y.; Kono, A.; Lavine, J.E.; Sirlin, C.B. Spectrally-Modeled Hepatic Fat Quantification by Multi-Echo Gradient-Recalled-Echo Magnetic Resonance Imaging at 3.0T. Proc. Intl. Soc. Mag. Reson. Med. 2009, 17, 209. [Google Scholar]
- Hamilton, G.; Yokoo, T.; Bydder, M.; Cruite, I.; Schroeder, M.E.; Sirlin, C.B.; Middleton, M.S. In vivo characterization of the liver fat (1)H MR spectrum. NMR Biomed. 2011, 24, 784–790. [Google Scholar] [CrossRef] [Green Version]
- Tang, A.; Tan, J.; Sun, M.; Hamilton, G.; Bydder, M.; Wolfson, T.; Gamst, A.C.; Middleton, M.; Brunt, E.M.; Loomba, R.; et al. Nonalcoholic fatty liver disease: MR imaging of liver proton density fat fraction to assess hepatic steatosis. Radiology 2013, 267, 422–431. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, M.; DeFronzo, R.A. Insulin sensitivity indices obtained from oral glucose tolerance testing: Comparison with the euglycemic insulin clamp. Diabetes Care 1999, 22, 1462–1470. [Google Scholar] [CrossRef]
- Tura, A.; Chemello, G.; Szendroedi, J.; Gobl, C.; Faerch, K.; Vrbikova, J.; Pacini, G.; Ferrannini, E.; Roden, M. Prediction of clamp-derived insulin sensitivity from the oral glucose insulin sensitivity index. Diabetologia 2018, 61, 1135–1141. [Google Scholar] [CrossRef] [Green Version]
- Mari, A.; Pacini, G.; Murphy, E.; Ludvik, B.; Nolan, J.J. A model-based method for assessing insulin sensitivity from the oral glucose tolerance test. Diabetes Care 2001, 24, 539–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1989, 28, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grundy, S.M.; Cleeman, J.I.; Daniels, S.R.; Donato, K.A.; Eckel, R.H.; Franklin, B.A.; Gordon, D.J.; Krauss, R.M.; Savage, P.J.; Smith, S.C., Jr.; et al. Diagnosis and management of the metabolic syndrome: An American Heart Association/National Heart, Lung, and Blood Institute scientific statement. Curr. Opin. Cardiol. 2006, 21, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Grundy, S.M.; Wang, W.; Smith, S.C., Jr.; Vega, G.L.; Wu, Z.; Zeng, Z.; Wang, W.; Zhao, D. Ethnic-specific criteria for the metabolic syndrome: Evidence from China. Diabetes Care 2006, 29, 1414–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Judd, C.M.; Kenny, D.A. Process Analysis: Estimating Mediation Treatment Evaluations. Evaluation Rev. 1981, 5, 602–619. [Google Scholar] [CrossRef]
- Fairchild, A.J.; McDaniel, H.L. Best (but oft-forgotten) practices: Mediation analysis. Am. J. Clin. Nutr. 2017, 105, 1259–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, C.R. Knockout mice challenge our concepts of glucose homeostasis and the pathogenesis of diabetes. Exp. Diabesity Res. 2003, 4, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Park, S.Y.; Choi, C.S. Insulin resistance: From mechanisms to therapeutic strategies. Diabetes Metab. J. 2021. [Google Scholar] [CrossRef]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]
- Bondini, S.; Younossi, Z.M. Non-alcoholic fatty liver disease and hepatitis C infection. Minerva Gastroenterol. Dietol. 2006, 52, 135–143. [Google Scholar]
- Day, C.P. Non-alcoholic fatty liver disease: Current concepts and management strategies. Clin. Med. 2006, 6, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Boslaugh, S. Encyclopedia of Epidemiology: Dose-Response Relationship; SAGE Publications, Inc.: Thousand Oaks, CA, USA, 2008. [Google Scholar]
- Rothman, K.J.; Greenland, S. Causation and causal inference in epidemiology. Am. J. Public Health 2005, 95 (Suppl. S1), S144–S150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Zhu, Y.; Malik, V.; Li, X.; Peng, X.; Zhang, F.F.; Shan, Z.; Liu, L. Intake of sugar-sweetened and low-calorie sweetened beverages and risk of cardiovascular disease: A meta-analysis and systematic review. Adv. Nutr 2021, 12, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Black, R.N.; Spence, M.; McMahon, R.O.; Cuskelly, G.J.; Ennis, C.N.; McCance, D.R.; Young, I.S.; Bell, P.M.; Hunter, S.J. Effect of eucaloric high- and low-sucrose diets with identical macronutrient profile on insulin resistance and vascular risk: A randomized controlled trial. Diabetes 2006, 55, 3566–3572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, V.; Micha, R.; Choi, E.; Karageorgou, D.; Webb, P.; Mozaffarian, D. Evaluation of the quality of evidence of the association of foods and nutrients with cardiovascular disease and diabetes: A systematic review. JAMA Netw. Open 2022, 5, e2146705. [Google Scholar] [CrossRef]
- Kazierad, D.J.; Chidsey, K.; Somayaji, V.R.; Bergman, A.J.; Birnbaum, M.J.; Calle, R.A. Inhibition of ketohexokinase in adults with NAFLD reduces liver fat and inflammatory markers: A randomized phase 2 trial. Clin. Adv. 2021, 2, 800–813. [Google Scholar] [CrossRef]
- Kahn, R.; Sievenpiper, J.L. Dietary sugar and body weight: Have we reached a crisis in the epidemic of obesity and diabetes? We have, but the pox on sugar is overwrought and overworked. Diabetes Care 2014, 37, 957–962. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J.C.; Horton, J.D.; Hobbs, H.H. Human fatty liver disease: Old questions and new insights. Science 2011, 332, 1519–1523. [Google Scholar] [CrossRef] [Green Version]
- Ramms, B.; Gordts, P. Apolipoprotein C-III in triglyceride-rich lipoprotein metabolism. Curr. Opin. Lipidol. 2018, 29, 171–179. [Google Scholar] [CrossRef]
- Paiva, A.A.; Raposo, H.F.; Wanschel, A.C.; Nardelli, T.R.; Oliveira, H.C. Apolipoprotein CIII Overexpression-Induced Hypertriglyceridemia Increases Nonalcoholic Fatty Liver Disease in Association with Inflammation and Cell Death. Oxid. Med. Cell. Longev. 2017, 2017, 1838679. [Google Scholar] [CrossRef]
- Choi, C.S.; Kim, Y.B.; Lee, F.N.; Zabolotny, J.M.; Kahn, B.B.; Youn, J.H. Lactate induces insulin resistance in skeletal muscle by suppressing glycolysis and impairing insulin signaling. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E233–E240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botteri, G.; Montori, M.; Guma, A.; Pizarro, J.; Cedo, L.; Escola-Gil, J.C.; Li, D.; Barroso, E.; Palomer, X.; Kohan, A.B.; et al. VLDL and apolipoprotein CIII induce ER stress and inflammation and attenuate insulin signalling via Toll-like receptor 2 in mouse skeletal muscle cells. Diabetologia 2017, 60, 2262–2273. [Google Scholar] [CrossRef] [PubMed]
- Christopoulou, E.; Tsimihodimos, V.; Filippatos, T.; Elisaf, M. Apolipoprotein CIII and diabetes. Is there a link? Diabetes Metab. Res. Rev. 2019, 35, e3118. [Google Scholar] [CrossRef] [PubMed]
- Bravo, S.; Lowndes, J.; Sinnett, S.; Yu, Z.; Rippe, J. Consumption of sucrose and high-fructose corn syrup does not increase liver fat or ectopic fat deposition in muscles. Appl. Physiol. Nutr. Metab. 2013, 38, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Lowndes, J.; Rippe, J. High-fructose corn syrup and sucrose have equivalent effects on energy-regulating hormones at normal human consumption levels. Nutr. Res. 2013, 33, 1043–1052. [Google Scholar] [CrossRef] [Green Version]
- Stanhope, K.L. Sugar consumption, metabolic disease and obesity: The state of the controversy. Crit. Rev. Clin. Lab. Sci. 2016, 53, 52–67. [Google Scholar] [CrossRef]
- Nestle, M. Corporate Funding of Food and Nutrition Research: Science or Marketing? JAMA Intern. Med. 2016, 176, 13–14. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture; U.S. Department of Health and Human Services. Dietary Guidelines for Americans, 2020–2025, 9th ed.; U.S. Department of Agriculture: Washington, DC, USA, 2020.
- Han, E.; Powell, L.M. Consumption patterns of sugar-sweetened beverages in the United States. J. Acad. Nutr. Diet. 2013, 113, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Caussy, C.; Reeder, S.B.; Sirlin, C.B.; Loomba, R. Noninvasive, quantitative assessment of liver fat by MRI-PDFF as an endpoint in NASH trials. Hepatology 2018, 68, 763–772. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).