
Sulfur Amino Acid Supplementation Abrogates Protective Effects of Caloric Restriction for Enhancing Bone Marrow Regrowth Following Ionizing Radiation

 , , ,
, , ,  and
and 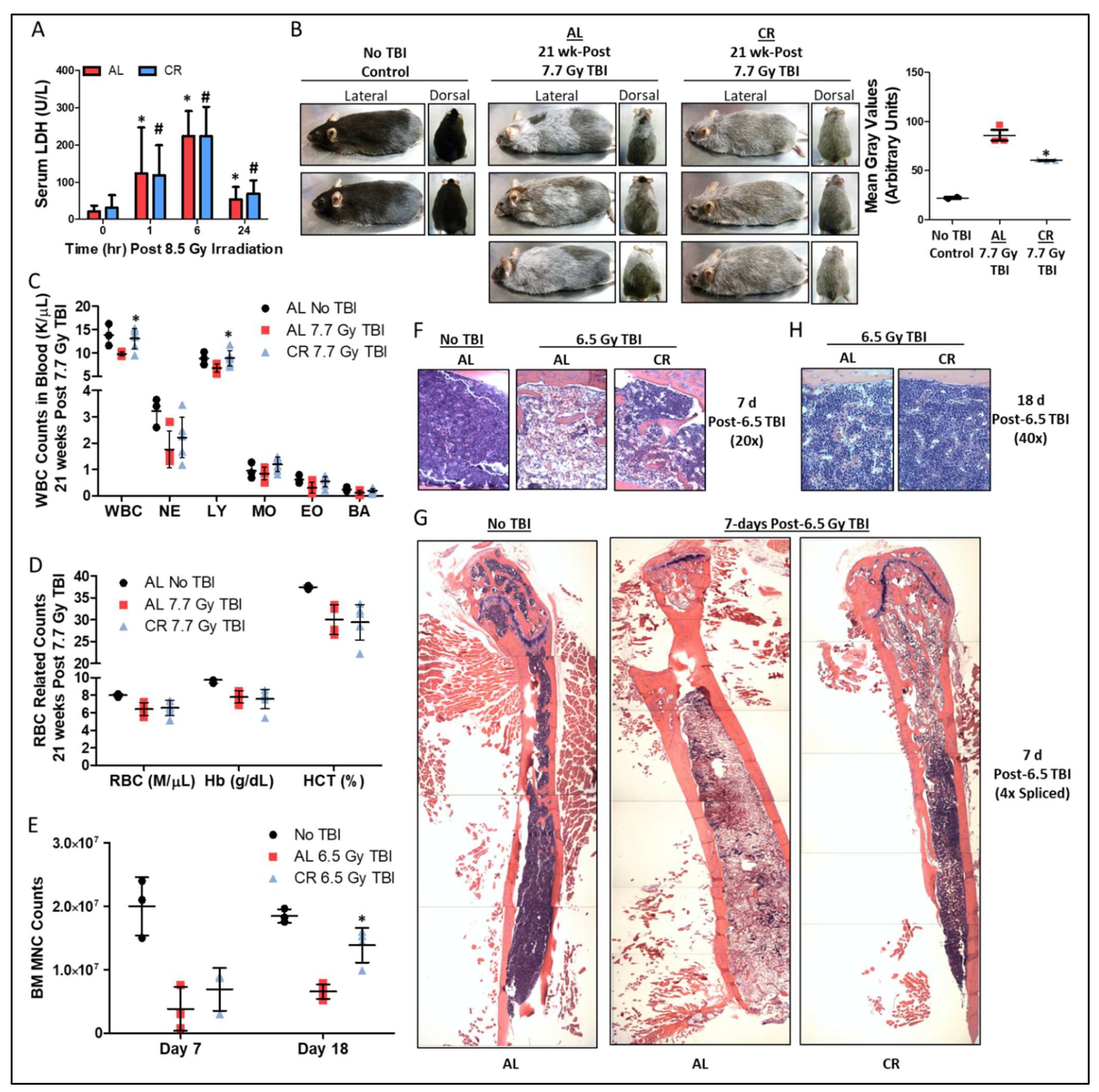{kind=link}
{kind=link}
{kind=link}
{kind=link}
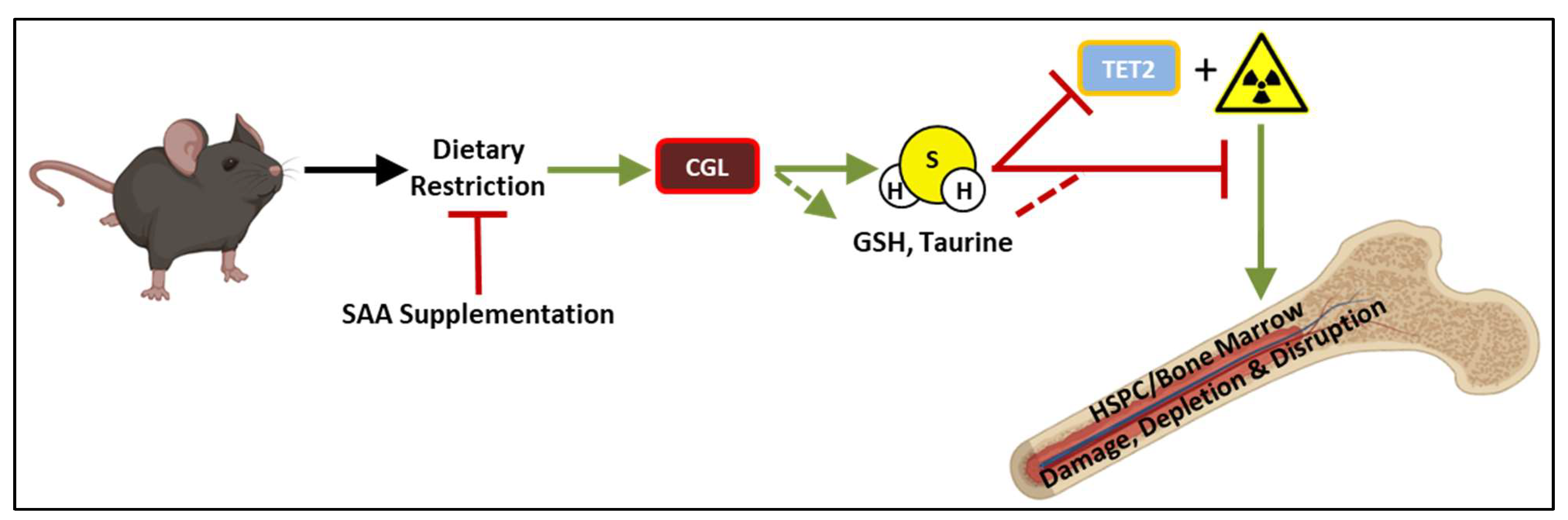{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Dietary Preconditioning
2.3. Bone Marrow Isolation and Counting of Bone Marrow Mononuclear Cells
2.4. Animal and Bone Marrow Irradiation
2.5. Detection and Analysis of Tissue Damage and Physiological Status
2.6. TET2 Activity Assays
2.7. H2S Production Assay
2.8. Data Quantification and Statistical Analysis
3. Results
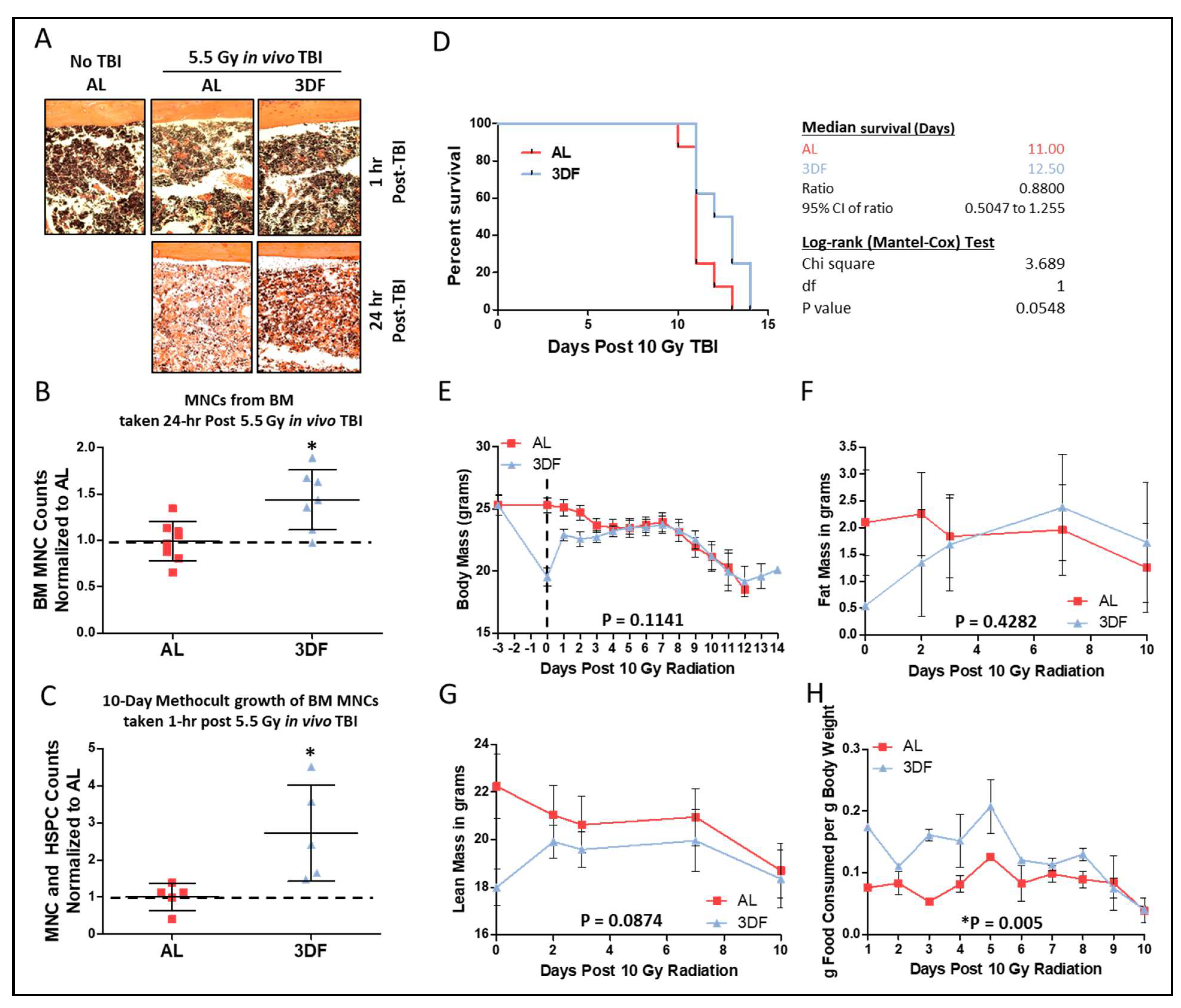
3.1. Short-Term CR Protects against Sub-Lethal γ-Irradiation Induced Damage In Vivo

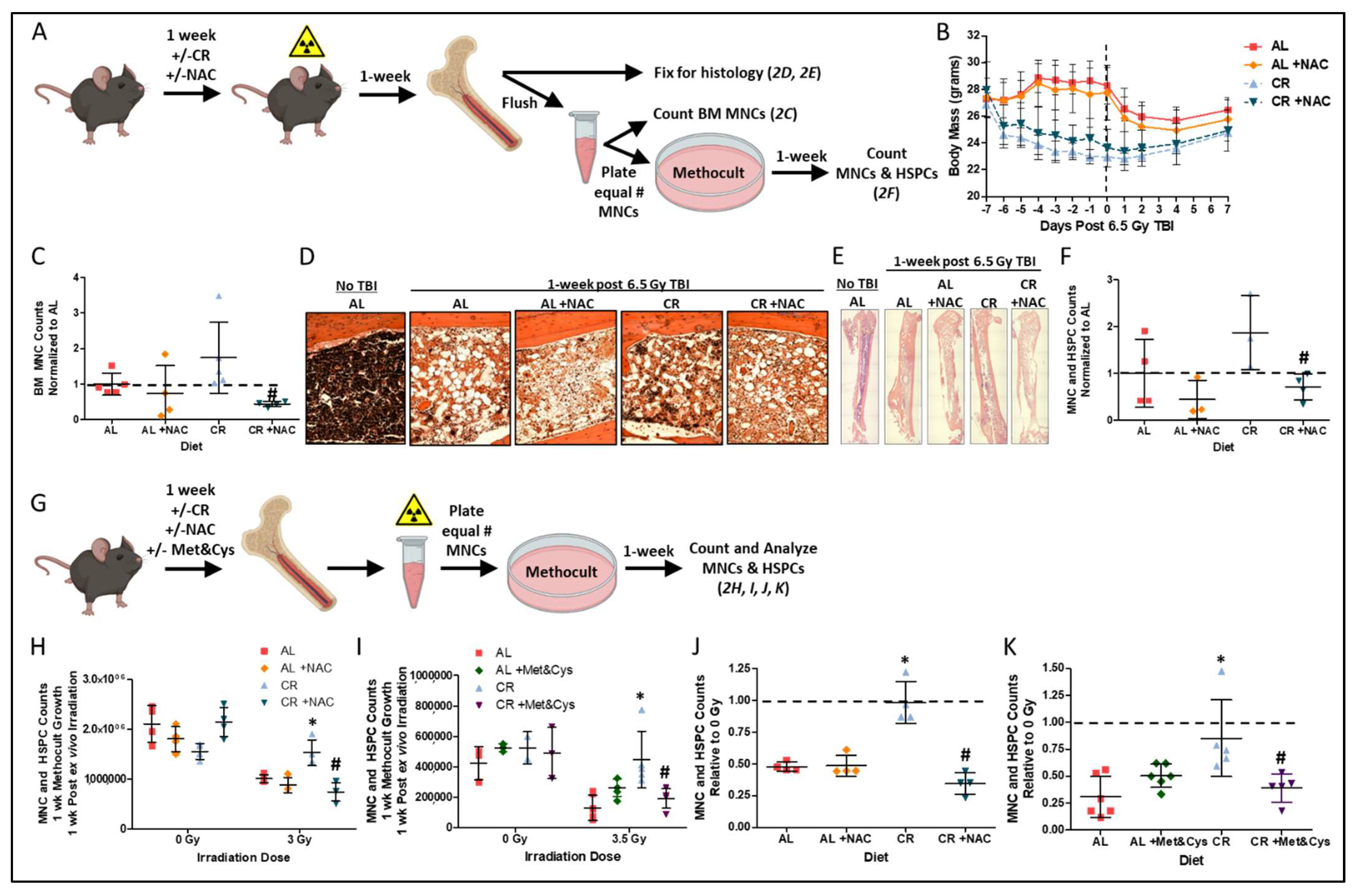
3.2. Sulfur Amino Acid Supplementation Suppresses the Radio-Protective Benefits of CR

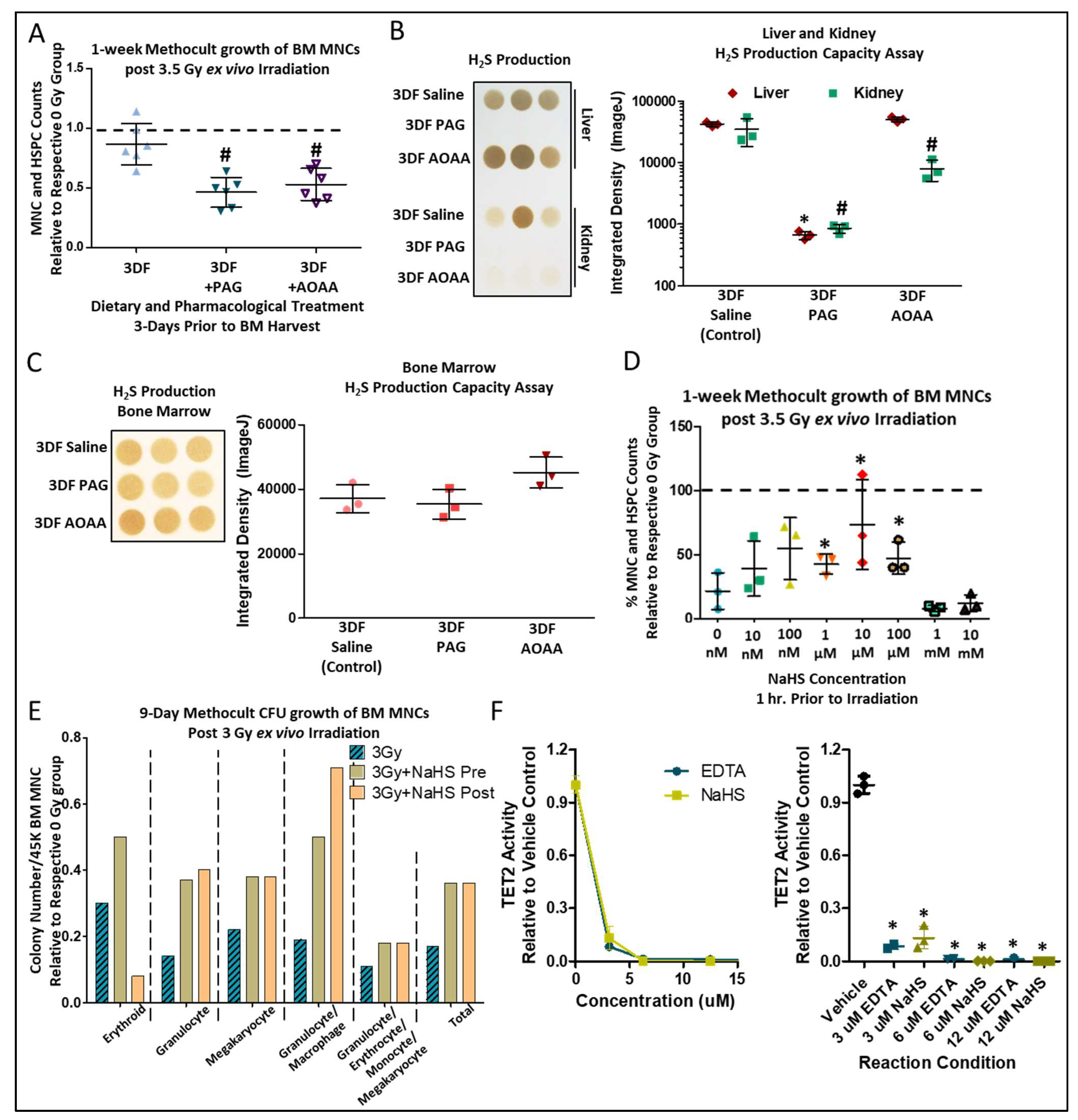
3.3. CGL Activity is Necessary, While H2S is Sufficient, for Short-Term CR-Mediated BM Radioprotection


4. Discussion

Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Mauch, P.; Constine, L.; Greenberger, J.; Knospe, W.; Sullivan, J.; Liesveld, J.L.; Deeg, H.J. Hematopoietic stem cell compartment: Acute and late effects of radiation therapy and chemotherapy. Int. J. Radiat. Oncol. Biol. Phys. 1995, 31, 1319–1339. [Google Scholar] [CrossRef]
- Lutgens, L.C.; Blijlevens, N.M.; Deutz, N.E.; Donnelly, J.P.; Lambin, P.; de Pauw, B.E. Monitoring myeloablative therapy-induced small bowel toxicity by serum citrulline concentration: A comparison with sugar permeability tests. Cancer 2005, 103, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Probin, V.; Zhou, D. Cancer therapy-induced residual bone marrow injury-Mechanisms of induction and implication for therapy. Curr. Cancer Ther. Rev. 2006, 2, 271–279. [Google Scholar] [CrossRef]
- O’Brien, M.M.; Donaldson, S.S.; Balise, R.R.; Whittemore, A.S.; Link, M.P. Second malignant neoplasms in survivors of pediatric Hodgkin’s lymphoma treated with low-dose radiation and chemotherapy. J. Clin. Oncol. 2010, 28, 1232–1239. [Google Scholar] [CrossRef]
- Costa, S.; Reagan, M.R. Therapeutic Irradiation: Consequences for Bone and Bone Marrow Adipose Tissue. Front. Endocrinol. 2019, 10, 587. [Google Scholar] [CrossRef]
- Curi, M.M.; Cardoso, C.L.; de Lima, H.G.; Kowalski, L.P.; Martins, M.D. Histopathologic and Histomorphometric Analysis of Irradiation Injury in Bone and the Surrounding Soft Tissues of the Jaws. J. Oral. Maxillofac. Surg. 2016, 74, 190–199. [Google Scholar] [CrossRef]
- Green, D.E.; Rubin, C.T. Consequences of irradiation on bone and marrow phenotypes, and its relation to disruption of hematopoietic precursors. Bone 2014, 63, 87–94. [Google Scholar] [CrossRef]
- Mendelson, A.; Frenette, P.S. Hematopoietic stem cell niche maintenance during homeostasis and regeneration. Nat. Med. 2014, 20, 833–846. [Google Scholar] [CrossRef]
- Fontana, L.; Partridge, L. Promoting health and longevity through diet: From model organisms to humans. Cell 2015, 161, 106–118. [Google Scholar] [CrossRef]
- Speakman, J.R.; Mitchell, S.E. Caloric restriction. Mol. Aspects Med. 2011, 32, 159–221. [Google Scholar] [CrossRef]
- Robertson, L.T.; Mitchell, J.R. Benefits of short-term dietary restriction in mammals. Exp. Gerontol. 2013, 48, 1043–1048. [Google Scholar] [CrossRef] [PubMed]
- Trocha, K.M.; Kip, P.; Tao, M.; MacArthur, M.R.; Trevino-Villarreal, J.H.; Longchamp, A.; Toussaint, W.; Lambrecht, B.N.; de Vries, M.R.; Quax, P.H.A.; et al. Short-term preoperative protein restriction attenuates vein graft disease via induction of cystathionine gamma-lyase. Cardiovasc. Res. 2020, 116, 416–428. [Google Scholar] [CrossRef] [PubMed]
- Trevino-Villarreal, J.H.; Reynolds, J.S.; Bartelt, A.; Langston, P.K.; MacArthur, M.R.; Arduini, A.; Tosti, V.; Veronese, N.; Bertozzi, B.; Brace, L.E.; et al. Dietary protein restriction reduces circulating VLDL triglyceride levels via CREBH-APOA5-dependent and -independent mechanisms. JCI Insight 2018, 3, e99470. [Google Scholar] [CrossRef] [PubMed]
- Hastreiter, A.A.; Galvao Dos Santos, G.; Cavalcante Santos, E.W.; Makiyama, E.N.; Borelli, P.; Fock, R.A. Protein malnutrition impairs bone marrow endothelial cells affecting hematopoiesis. Clin. Nutr. 2020, 39, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Astle, C.M.; Harrison, D.E. Hematopoietic senescence is postponed and hematopoietic stem cell function is enhanced by dietary restriction. Exp. Hematol. 2003, 31, 1097–1103. [Google Scholar] [CrossRef]
- Tang, D.; Tao, S.; Chen, Z.; Koliesnik, I.O.; Calmes, P.G.; Hoerr, V.; Han, B.; Gebert, N.; Zornig, M.; Loffler, B.; et al. Dietary restriction improves repopulation but impairs lymphoid differentiation capacity of hematopoietic stem cells in early aging. J. Exp. Med. 2016, 213, 535–553. [Google Scholar] [CrossRef]
- Brandhorst, S.; Choi, I.Y.; Wei, M.; Cheng, C.W.; Sedrakyan, S.; Navarrete, G.; Dubeau, L.; Yap, L.P.; Park, R.; Vinciguerra, M.; et al. A Periodic Diet that Mimics Fasting Promotes Multi-System Regeneration, Enhanced Cognitive Performance, and Healthspan. Cell Metab. 2015, 22, 86–99. [Google Scholar] [CrossRef]
- Cheng, C.W.; Adams, G.B.; Perin, L.; Wei, M.; Zhou, X.; Lam, B.S.; Da Sacco, S.; Mirisola, M.; Quinn, D.I.; Dorff, T.B.; et al. Prolonged fasting reduces IGF-1/PKA to promote hematopoietic-stem-cell-based regeneration and reverse immunosuppression. Cell Stem Cell 2014, 14, 810–823. [Google Scholar] [CrossRef]
- Kozubík, A.; Pospísil, M.; Hosek, B. Stimulatory effect of intermittent feeding on hemopoietic recovery in sublethally gamma-irradiated mice. Acta Radiol. Oncol. 1985, 24, 199–204. [Google Scholar] [CrossRef]
- Kozubík, A.; Pospísil, M. Intermittent feeding as a factor enhancing hemopoietic stem cell proliferation and spleen colony formation in irradiated mice. Acta Radiol. Oncol. 1985, 24, 357–361. [Google Scholar] [CrossRef]
- Kozubík, A.; Pospísil, M. Adaptation to intermittent fasting as a factor modifying the radiation resistance of mice. Experientia 1982, 38, 958–959. [Google Scholar] [CrossRef] [PubMed]
- Gross, L.; Dreyfuss, Y. Reduction in the incidence of radiation-induced tumors in rats after restriction of food intake. Proc. Natl. Acad. Sci. USA 1984, 81, 7596–7598. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Inoue, T.; Nojima, K.; Hirabayashi, Y.; Sado, T. Calorie restriction reduces the incidence of myeloid leukemia induced by a single whole-body radiation in C3H/He mice. Proc. Natl. Acad. Sci. USA 1997, 94, 2615–2619. [Google Scholar] [CrossRef] [PubMed]
- Harputlugil, E.; Hine, C.; Vargas, D.; Robertson, L.; Manning, B.D.; Mitchell, J.R. The TSC Complex Is Required for the Benefits of Dietary Protein Restriction on Stress Resistance In Vivo. Cell Rep. 2014, 8, 1160–1170. [Google Scholar] [CrossRef]
- Robertson, L.T.; Trevino-Villarreal, J.H.; Mejia, P.; Grondin, Y.; Harputlugil, E.; Hine, C.; Vargas, D.; Zheng, H.; Ozaki, C.K.; Kristal, B.S.; et al. Protein and Calorie Restriction Contribute Additively to Protection from Renal Ischemia Reperfusion Injury Partly via Leptin Reduction in Male Mice. J. Nutr. 2015, 145, 1717–1727. [Google Scholar] [CrossRef]
- Hine, C.; Zhu, Y.; Hollenberg, A.N.; Mitchell, J.R. Dietary and Endocrine Regulation of Endogenous Hydrogen Sulfide Production: Implications for Longevity. Antioxid Redox Signal. 2018, 28, 1483–1502. [Google Scholar] [CrossRef]
- Kabil, H.; Kabil, O.; Banerjee, R.; Harshman, L.G.; Pletcher, S.D. Increased transsulfuration mediates longevity and dietary restriction in Drosophila. Proc. Natl. Acad. Sci. USA 2011, 108, 16831–16836. [Google Scholar] [CrossRef]
- Longchamp, A.; Mirabella, T.; Arduini, A.; MacArthur, M.R.; Das, A.; Trevino-Villarreal, J.H.; Hine, C.; Ben-Sahra, I.; Knudsen, N.H.; Brace, L.E.; et al. Amino Acid Restriction Triggers Angiogenesis via GCN2/ATF4 Regulation of VEGF and H2S Production. Cell 2018, 173, 117–129.e114. [Google Scholar] [CrossRef]
- Kabil, O.; Vitvitsky, V.; Xie, P.; Banerjee, R. The quantitative significance of the transsulfuration enzymes for H2S production in murine tissues. Antioxid Redox Signal. 2011, 15, 363–372. [Google Scholar] [CrossRef]
- Hine, C.; Harputlugil, E.; Zhang, Y.; Ruckenstuhl, C.; Lee, B.C.; Brace, L.; Longchamp, A.; Treviño-Villarreal, J.H.; Mejia, P.; Ozaki, C.K.; et al. Endogenous hydrogen sulfide production is essential for dietary restriction benefits. Cell 2015, 160, 132–144. [Google Scholar] [CrossRef]
- Guan, Y.; Hasipek, M.; Jiang, D.; Tiwari, A.D.; Grabowski, D.R.; Pagliuca, S.; Kongkiatkamon, S.; Patel, B.; Singh, S.; Parker, Y.; et al. Eltrombopag inhibits TET dioxygenase to contribute to hematopoietic stem cell expansion in aplastic anemia. J. Clin. Investig. 2022, 132, e2382. [Google Scholar] [CrossRef] [PubMed]
- Hine, C.; Mitchell, J.R. Endpoint or Kinetic Measurement of Hydrogen Sulfide Production Capacity in Tissue Extracts. Bio-protocol 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Levin, S.G.; Young, R.W.; Stohler, R.L. Estimation of median human lethal radiation dose computed from data on occupants of reinforced concrete structures in Nagasaki, Japan. Health Phys. 1992, 63, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.H.; Park, J.H.; Jeong, E.S.; Choi, S.Y.; Ham, S.H.; Park, J.I.; Jeon, H.Y.; Kim, J.Y.; Yoo, R.J.; Lee, Y.J.; et al. Establishment of a mouse model of 70% lethal dose by total-body irradiation. Lab. Anim. Res. 2016, 32, 116–121. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yoshida, S.; Yamahara, K.; Kume, S.; Koya, D.; Yasuda-Yamahara, M.; Takeda, N.; Osawa, N.; Chin-Kanasaki, M.; Adachi, Y.; Nagao, K.; et al. Role of dietary amino acid balance in diet restriction-mediated lifespan extension, renoprotection, and muscle weakness in aged mice. Aging Cell 2018, 17, e12796. [Google Scholar] [CrossRef]
- Kaufman, D.S.; Hanson, E.T.; Lewis, R.L.; Auerbach, R.; Thomson, J.A. Hematopoietic colony-forming cells derived from human embryonic stem cells. Proc. Natl. Acad. Sci. USA 2001, 98, 10716–10721. [Google Scholar] [CrossRef] [PubMed]
- Hine, C.; Kim, H.J.; Zhu, Y.; Harputlugil, E.; Longchamp, A.; Matos, M.S.; Ramadoss, P.; Bauerle, K.; Brace, L.; Asara, J.M.; et al. Hypothalamic-Pituitary Axis Regulates Hydrogen Sulfide Production. Cell Metab. 2017, 25, 1320–1333.e1325. [Google Scholar] [CrossRef]
- Bithi, N.; Link, C.; Henderson, Y.O.; Kim, S.; Yang, J.; Li, L.; Wang, R.; Willard, B.; Hine, C. Dietary restriction transforms the mammalian protein persulfidome in a tissue-specific and cystathionine gamma-lyase-dependent manner. Nat. Commun. 2021, 12, 1745. [Google Scholar] [CrossRef]
- Sun, X.; Wang, W.; Dai, J.; Jin, S.; Huang, J.; Guo, C.; Wang, C.; Pang, L.; Wang, Y. A Long-Term and Slow-Releasing Hydrogen Sulfide Donor Protects against Myocardial Ischemia/Reperfusion Injury. Sci. Rep. 2017, 7, 3541. [Google Scholar] [CrossRef]
- Sun, X.; Wang, W.; Dai, J.; Huang, J.; Shi, M.; Chu, X.; Wang, F.; Guo, C.; Wang, C.; Pang, L.; et al. Donor heart preservation with a novel long-term and slow-releasing hydrogen sulfide system. Nitric Oxide 2018, 81, 1–10. [Google Scholar] [CrossRef]
- Polhemus, D.J.; Kondo, K.; Bhushan, S.; Bir, S.C.; Kevil, C.G.; Murohara, T.; Lefer, D.J.; Calvert, J.W. Hydrogen sulfide attenuates cardiac dysfunction after heart failure via induction of angiogenesis. Circ. Heart Fail. 2013, 6, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- Lv, S.; Wang, Z.; Wang, J.; Wang, H. Exogenous Hydrogen Sulfide Plays an Important Role Through Regulating Autophagy in Ischemia/Reperfusion Injury. Front Mol. Biosci. 2021, 8, 681676. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.D.; Zhang, A.J.; Xu, J.J.; Chen, Y.; Zhu, Y.C. H2S protects against fatal myelosuppression by promoting the generation of megakaryocytes/platelets. J. Hematol. Oncol. 2016, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xie, Y.; Xu, Y.; Pan, Y.; Shao, C. Hydrogen sulfide contributes to hypoxia-induced radioresistance on hepatoma cells. J. Radiat. Res. 2011, 52, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.R.; Verweij, M.; Brand, K.; van de Ven, M.; Goemaere, N.; van den Engel, S.; Chu, T.; Forrer, F.; Muller, C.; de Jong, M.; et al. Short-term dietary restriction and fasting precondition against ischemia reperfusion injury in mice. Aging Cell. 2010, 9, 40–53. [Google Scholar] [CrossRef]
- Asimakopoulou, A.; Panopoulos, P.; Chasapis, C.T.; Coletta, C.; Zhou, Z.; Cirino, G.; Giannis, A.; Szabo, C.; Spyroulias, G.A.; Papapetropoulos, A. Selectivity of commonly used pharmacological inhibitors for cystathionine beta synthase (CBS) and cystathionine gamma lyase (CSE). Br. J. Pharmacol. 2013, 169, 922–932. [Google Scholar] [CrossRef]
- Yang, J.; Minkler, P.; Grove, D.; Wang, R.; Willard, B.; Dweik, R.; Hine, C. Non-enzymatic hydrogen sulfide production from cysteine in blood is catalyzed by iron and vitamin B6. Commun. Biol. 2019, 2, 194. [Google Scholar] [CrossRef]
- Henderson, Y.O.; Bithi, N.; Link, C.; Yang, J.; Schugar, R.; Llarena, N.; Brown, J.M.; Hine, C. Late-life intermittent fasting decreases aging-related frailty and increases renal hydrogen sulfide production in a sexually dimorphic manner. Geroscience 2021, 43, 1527–1554. [Google Scholar] [CrossRef]
- Yang, J.; Link, C.; Henderson, Y.O.; Bithi, N.; Hine, C. Peripubertal Bisphenol A Exposure Imparts Detrimental Age-Related Changes in Body Composition, Cognition, and Hydrogen Sulfide Production Capacities. Antioxid Redox Signal 2021. [Google Scholar] [CrossRef]
- Mitchell, J.B.; Biaglow, J.E.; Russo, A. Role of glutathione and other endogenous thiols in radiation protection. Pharmacol. Ther. 1988, 39, 269–274. [Google Scholar] [CrossRef]
- Smith, T.A.; Kirkpatrick, D.R.; Smith, S.; Smith, T.K.; Pearson, T.; Kailasam, A.; Herrmann, K.Z.; Schubert, J.; Agrawal, D.K. Radioprotective agents to prevent cellular damage due to ionizing radiation. J. Transl. Med. 2017, 15, 232. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Kato, T.; Isogai, T.; Gu, Y.; Ma, N. Protective Effects of Taurine on the Radiation Exposure Induced Cellular Damages in the Mouse Intestine. Adv. Exp. Med. Biol. 2019, 1155, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Olson, K.R. Is hydrogen sulfide a circulating "gasotransmitter" in vertebrate blood? Biochim. Biophys. Acta 2009, 1787, 856–863. [Google Scholar] [CrossRef]
- Mauro, C.R.; Tao, M.; Yu, P.; Treviño-Villerreal, J.H.; Longchamp, A.; Kristal, B.S.; Ozaki, C.K.; Mitchell, J.R. Preoperative dietary restriction reduces intimal hyperplasia and protects from ischemia-reperfusion injury. J. Vasc. Surg. 2014, 63, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Koch, C.J.; Evans, S.M. Cysteine concentrations in rodent tumors: Unexpectedly high values may cause therapy resistance. Int. J. Cancer 1996, 67, 661–667. [Google Scholar] [CrossRef]
- Reisz, J.A.; Bansal, N.; Qian, J.; Zhao, W.; Furdui, C.M. Effects of ionizing radiation on biological molecules--mechanisms of damage and emerging methods of detection. Antioxid Redox Signal. 2014, 21, 260–292. [Google Scholar] [CrossRef]
- Calvert, J.W.; Coetzee, W.A.; Lefer, D.J. Novel insights into hydrogen sulfide--mediated cytoprotection. Antioxid Redox Signal. 2010, 12, 1203–1217. [Google Scholar] [CrossRef]
- Zhao, K.; Ju, Y.; Li, S.; Altaany, Z.; Wang, R.; Yang, G. S-sulfhydration of MEK1 leads to PARP-1 activation and DNA damage repair. EMBO Rep. 2014, 15, 792–800. [Google Scholar] [CrossRef]
- Shuang, T.; Fu, M.; Yang, G.; Wu, L.; Wang, R. The interaction of IGF-1/IGF-1R and hydrogen sulfide on the proliferation of mouse primary vascular smooth muscle cells. Biochem. Pharmacol. 2017, 149, 143–152. [Google Scholar] [CrossRef]
- Zhang, J.; Xie, Y.; Xu, Y.; Shao, C. Suppression of endogenous hydrogen sulfide contributes to the radiation-induced bystander effects on hypoxic HepG2 cells. Radiat. Res. 2012, 178, 395–402. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, R.; Liu, X.; Zhou, Y.; Qu, C.; Kikuiri, T.; Wang, S.; Zandi, E.; Du, J.; Ambudkar, I.S.; et al. Hydrogen sulfide maintains mesenchymal stem cell function and bone homeostasis via regulation of Ca(2+) channel sulfhydration. Cell Stem Cell 2014, 15, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; MacArthur, M.R.; Trevino-Villarreal, J.H.; Kip, P.; Ozaki, C.K.; Mitchell, S.J.; Mitchell, J.R. Intracellular H2S production is an autophagy-dependent adaptive response to DNA damage. Cell Chem Biol. 2021, 28, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Glode, L.M.; Epstein, A.; Smith, C.G. Reduced gamma-cystathionase protein content in human malignant leukemia cell lines as measured by immunoassay with monoclonal antibody. Cancer Res. 1981, 41, 2249–2254. [Google Scholar] [PubMed]
- Glode, L.M.; Kriegler, M.P.; Livingston, D.M. Cysteine auxotrophy of human leukemic lymphoblasts is associated with decreased amounts of intracellular cystathionase protein. Biochemistry 1981, 20, 1306–1311. [Google Scholar] [CrossRef]
- Zivanovic, J.; Kouroussis, E.; Kohl, J.B.; Adhikari, B.; Bursac, B.; Schott-Roux, S.; Petrovic, D.; Miljkovic, J.L.; Thomas-Lopez, D.; Jung, Y.; et al. Selective Persulfide Detection Reveals Evolutionarily Conserved Antiaging Effects of S-Sulfhydration. Cell Metab. 2020, 31, 207. [Google Scholar] [CrossRef]
- Genovese, G.; Kahler, A.K.; Handsaker, R.E.; Lindberg, J.; Rose, S.A.; Bakhoum, S.F.; Chambert, K.; Mick, E.; Neale, B.M.; Fromer, M.; et al. Clonal hematopoiesis and blood-cancer risk inferred from blood DNA sequence. N. Engl. J. Med. 2014, 371, 2477–2487. [Google Scholar] [CrossRef]
- Peng, W.; Robertson, L.; Gallinetti, J.; Mejia, P.; Vose, S.; Charlip, A.; Chu, T.; Mitchell, J.R. Surgical stress resistance induced by single amino Acid deprivation requires gcn2 in mice. Sci. Transl. Med. 2012, 4, 118ra111. [Google Scholar] [CrossRef]
- Jonsson, W.O.; Margolies, N.S.; Anthony, T.G. Dietary Sulfur Amino Acid Restriction and the Integrated Stress Response: Mechanistic Insights. Nutrients 2019, 11, 1349. [Google Scholar] [CrossRef]
- Jonsson, W.O.; Margolies, N.S.; Mirek, E.T.; Zhang, Q.; Linden, M.A.; Hill, C.M.; Link, C.; Bithi, N.; Zalma, B.; Levy, J.L.; et al. Physiologic Responses to Dietary Sulfur Amino Acid Restriction in Mice Are Influenced by Atf4 Status and Biological Sex. J. Nutr. 2021, 151, 785–799. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, S.; Galson, D.L.; Luo, M.; Fan, J.; Zhang, J.; Guan, Y.; Xiao, G. Activating transcription factor 4 is critical for proliferation and survival in primary bone marrow stromal cells and calvarial osteoblasts. J. Cell. Biochem. 2008, 105, 885–895. [Google Scholar] [CrossRef]
- Nakano, S.; Ishii, I.; Shinmura, K.; Tamaki, K.; Hishiki, T.; Akahoshi, N.; Ida, T.; Nakanishi, T.; Kamata, S.; Kumagai, Y.; et al. Hyperhomocysteinemia abrogates fasting-induced cardioprotection against ischemia/reperfusion by limiting bioavailability of hydrogen sulfide anions. J. Mol. Med. 2015, 93, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Elshorbagy, A.K.; Valdivia-Garcia, M.; Mattocks, D.A.; Plummer, J.D.; Orentreich, D.S.; Orentreich, N.; Refsum, H.; Perrone, C.E. Effect of taurine and N-acetylcysteine on methionine restriction-mediated adiposity resistance. Metabolism 2013, 62, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Elshorbagy, A.K.; Valdivia-Garcia, M.; Mattocks, D.A.; Plummer, J.D.; Smith, A.D.; Drevon, C.A.; Refsum, H.; Perrone, C.E. Cysteine supplementation reverses methionine restriction effects on rat adiposity: Significance of stearoyl-coenzyme A desaturase. J. Lipid. Res. 2011, 52, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Chen, J.; Zhou, J.; Martin, C.K.; Ravussin, E.; Redman, L.M. Effect of 2-year caloric restriction on organ and tissue size in nonobese 21- to 50-year-old adults in a randomized clinical trial: The CALERIE study. Am. J. Clin. Nutr. 2021, 114, 1295–1303. [Google Scholar] [CrossRef] [PubMed]
- Ravussin, E.; Redman, L.M.; Rochon, J.; Das, S.K.; Fontana, L.; Kraus, W.E.; Romashkan, S.; Williamson, D.A.; Meydani, S.N.; Villareal, D.T.; et al. A 2-Year Randomized Controlled Trial of Human Caloric Restriction: Feasibility and Effects on Predictors of Health Span and Longevity. J. Gerontol. A Biol. Sci. Med. Sci. 2015, 70, 1097–1104. [Google Scholar] [CrossRef]
- Charoensri, S.; Sothornwit, J.; Trirattanapikul, A.; Pongchaiyakul, C. Ketogenic Diet-Induced Diabetic Ketoacidosis in a Young Adult with Unrecognized Type 1 Diabetes. Case Rep. Endocrinol. 2021, 2021, 6620832. [Google Scholar] [CrossRef]
- Bianchi, V.E. Role of nutrition on anemia in elderly. Clin. Nutr. ESPEN 2016, 11, e1–e11. [Google Scholar] [CrossRef]
- Spadaro, O.; Youm, Y.; Shchukina, I.; Ryu, S.; Sidorov, S.; Ravussin, A.; Nguyen, K.; Aladyeva, E.; Predeus, A.N.; Smith, S.R.; et al. Caloric restriction in humans reveals immunometabolic regulators of health span. Science 2022, 375, 671–677. [Google Scholar] [CrossRef]
- Dong, Z.; Gao, X.; Chinchilli, V.M.; Sinha, R.; Muscat, J.; Winkels, R.M.; Richie, J.P., Jr. Association of sulfur amino acid consumption with cardiometabolic risk factors: Cross-sectional findings from NHANES III. Eclinical Med. 2020, 19, 100248. [Google Scholar] [CrossRef]
- MacArthur, M.R.; Mitchell, S.J.; Trevino-Villarreal, J.H.; Grondin, Y.; Reynolds, J.S.; Kip, P.; Jung, J.; Trocha, K.M.; Ozaki, C.K.; Mitchell, J.R. Total protein, not amino acid composition, differs in plant-based versus omnivorous dietary patterns and determines metabolic health effects in mice. Cell Metab. 2021, 33, 1808–1819.e1802. [Google Scholar] [CrossRef]
- Wiliński, B.; Wiliński, J.; Somogyi, E.; Piotrowska, J.; Opoka, W. Metformin raises hydrogen sulfide tissue concentrations in various mouse organs. Pharmacol. Rep. 2013, 65, 737–742. [Google Scholar] [CrossRef]
- Xu, G.; Wu, H.; Zhang, J.; Li, D.; Wang, Y.; Wang, Y.; Zhang, H.; Lu, L.; Li, C.; Huang, S.; et al. Metformin ameliorates ionizing irradiation-induced long-term hematopoietic stem cell injury in mice. Free Radic. Biol. Med. 2015, 87, 15–25. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F.; Barroso-Aranda, J.; Contreras, F. The low-methionine content of vegan diets may make methionine restriction feasible as a life extension strategy. Med. Hypotheses 2009, 72, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Lobb, I.; Davison, M.; Carter, D.; Liu, W.; Haig, A.; Gunaratnam, L.; Sener, A. Hydrogen Sulfide Treatment Mitigates Renal Allograft Ischemia-Reperfusion Injury during Cold Storage and Improves Early Transplant Kidney Function and Survival Following Allogeneic Renal Transplantation. J. Urol. 2015, 194, 1806–1815. [Google Scholar] [CrossRef]
- Hu, X.; Li, T.; Bi, S.; Jin, Z.; Zhou, G.; Bai, C.; Li, L.; Cui, Q.; Liu, W. Possible role of hydrogen sulfide on the preservation of donor rat hearts. Transpl. Proc. 2007, 39, 3024–3029. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hine, C.; Treviño-Villarreal, J.H.; Mejia, P.; Longchamp, A.; Brace, L.E.; Harputlugil, E.; Mitchell, S.J.; Yang, J.; Guan, Y.; Maciejewski, J.P.; et al. Sulfur Amino Acid Supplementation Abrogates Protective Effects of Caloric Restriction for Enhancing Bone Marrow Regrowth Following Ionizing Radiation. Nutrients 2022, 14, 1529. https://doi.org/10.3390/nu14071529
Hine C, Treviño-Villarreal JH, Mejia P, Longchamp A, Brace LE, Harputlugil E, Mitchell SJ, Yang J, Guan Y, Maciejewski JP, et al. Sulfur Amino Acid Supplementation Abrogates Protective Effects of Caloric Restriction for Enhancing Bone Marrow Regrowth Following Ionizing Radiation. Nutrients. 2022; 14(7):1529. https://doi.org/10.3390/nu14071529
Chicago/Turabian StyleHine, Christopher, J. Humberto Treviño-Villarreal, Pedro Mejia, Alban Longchamp, Lear E. Brace, Eylul Harputlugil, Sarah J. Mitchell, Jie Yang, Yihong Guan, Jaroslaw P. Maciejewski, and et al. 2022. "Sulfur Amino Acid Supplementation Abrogates Protective Effects of Caloric Restriction for Enhancing Bone Marrow Regrowth Following Ionizing Radiation" Nutrients 14, no. 7: 1529. https://doi.org/10.3390/nu14071529
APA StyleHine, C., Treviño-Villarreal, J. H., Mejia, P., Longchamp, A., Brace, L. E., Harputlugil, E., Mitchell, S. J., Yang, J., Guan, Y., Maciejewski, J. P., Jha, B. K., & Mitchell, J. R. (2022). Sulfur Amino Acid Supplementation Abrogates Protective Effects of Caloric Restriction for Enhancing Bone Marrow Regrowth Following Ionizing Radiation. Nutrients, 14(7), 1529. https://doi.org/10.3390/nu14071529