
Development of the Gastrointestinal Tract in Newborns as a Challenge for an Appropriate Nutrition: A Narrative Review

 , , ,
, , , 
{kind=link}
Abstract
1. Introduction
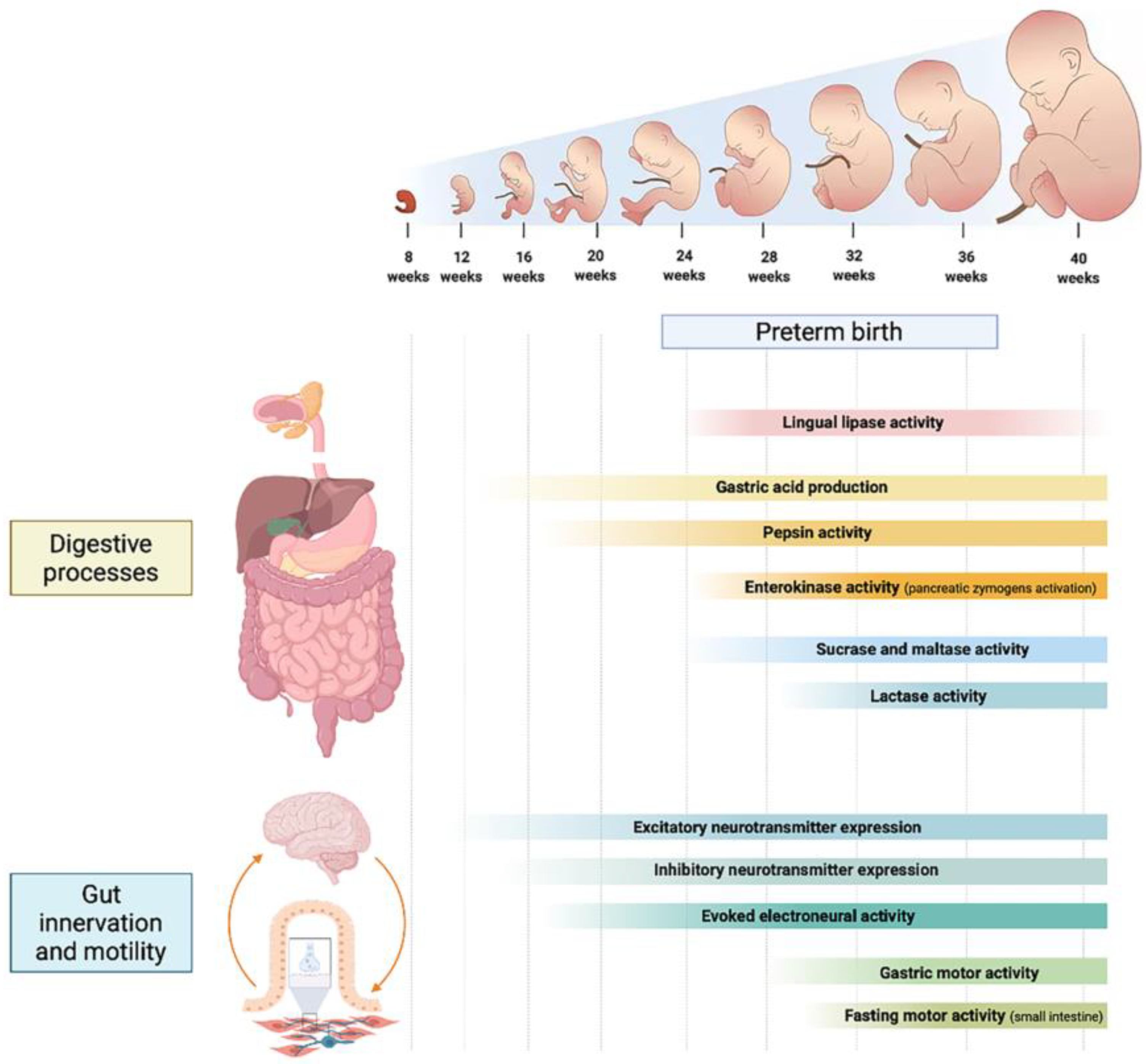
2. Development of Digestion and Absorption in the Neonate
2.1. Proteins
2.2. Carbohydrates
2.3. Lipids
3. The Development of Intestinal Motility
3.1. Anatomy and Physiology of Gastrointestinal Motility
3.2. The Enteric Nervous System
3.3. The Microbiota-Gut–Brain Axis
4. Development of the Gut Microbiome
4.1. Gut Microbiome in Term Infants
4.2. Gut Microbiome in Preterm Infants
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Harding, J.E.; Cormack, B.E.; Alexander, T.; Alsweiler, J.M.; Bloomfield, F.H. Advances in nutrition of the newborn infant. Lancet 2017, 389, 1660–1668. [Google Scholar] [CrossRef]
- Bourlieu, C.; Ménard, O.; Bouzerzour, K.; Mandalari, G.; Macierzanka, A.; Mackie, A.R.; Dupont, D. Specificity of Infant Digestive Conditions: Some Clues for Developing Relevant In Vitro Models. Crit. Rev. Food Sci. Nutr. 2014, 54, 1427–1457. [Google Scholar] [CrossRef] [PubMed]
- Poquet, L.; Wooster, T.J. Infant digestion physiology and the relevance of in vitro biochemical models to test infant formula lipid digestion. Mol. Nutr. Food Res. 2016, 60, 1876–1895. [Google Scholar] [CrossRef] [PubMed]
- Mooij, M.G.; de Koning, B.A.; Huijsman, M.L.; de Wildt, S.N. Ontogeny of oral drug absorption processes in children. Expert Opin. Drug Metab. Toxicol. 2012, 8, 1293–1303. [Google Scholar] [CrossRef]
- Lin, H.C. Small intestinal bacterial overgrowth: A framework for understanding irritable bowel syndrome. JAMA 2004, 292, 852–858. [Google Scholar] [CrossRef]
- Lucchini, R.; Bizzarri, B.; Giampietro, S.; De Curtis, M. Feeding intolerance in preterm infants. How to understand the warning signs. J. Matern.-Fetal Neonatal Med. 2011, 24 (Suppl. 1), 72–74. [Google Scholar] [CrossRef]
- Neu, J. Gastrointestinal development and meeting the nutritional needs of premature infants. Am. J. Clin. Nutr. 2007, 85, 629S–634S. [Google Scholar] [CrossRef]
- Lebenthal, A.; Lebenthal, E. The Ontogeny of the Small Intestinal Epithelium. J. Parenter. Enter. Nutr. 1999, 23, S3–S6. [Google Scholar] [CrossRef]
- Berseth, C.L. Gastrointestinal Motility in the Neonate. Clin. Perinatol. 1996, 23, 179–190. [Google Scholar] [CrossRef]
- Indrio, F.; Riezzo, G.; Cavallo, L.; Di Mauro, A.; Francavilla, R. Physiological basis of food intolerance in VLBW. J. Matern.-Fetal Neonatal Med. 2011, 24, 64–66. [Google Scholar] [CrossRef]
- Gray, H.; Warwick, R.; Williams, P.L. Gray’s Anatomy, 36th ed.; Saunders: Philadelphia, PA, USA, 1980; p. XVII. 1578p. [Google Scholar]
- Weaver, L.T.; Austin, S.; Cole, T.J. Small intestinal length: A factor essential for gut adaptation. Gut 1991, 32, 1321–1323. [Google Scholar] [CrossRef]
- Mezoff, E.A.; Shroyer, N.F. Anatomy and physiology of the small and large intestines. In Pediatric Gastrointestinal and Liver Disease, 5th ed.; Wyllie, R., Hyams, J.S., Kay, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Montgomery, R.K.; Mulberg, A.E.; Grand, R.J. Development of the human gastrointestinal tract: Twenty years of progress. Gastroenterology 1999, 116, 702–731. [Google Scholar] [CrossRef]
- Neu, J. Digestive-absorption functions in fetuses, infants and children and the developing microbiome of the fetus and newborn. In Fetal and Neonatal Physiology, 5th ed.; Polin, R.A., Abman, S.H., Rowtich, D.H., Benitz, W.E., Fox, W.W., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 1, pp. 897–908. [Google Scholar]
- Neu, J.; Douglas-Escobar, M.; Fucile, S. Gastrointestinal development: Implications for infant feeding. In Nutrition in Pediatrics, 5th ed.; Duggan, C., Watkins, J.B., Koletzko, B., Walker, W.A., Eds.; People’s Medical Publishing House: Beijing, China, 2016; Volume 1, pp. 387–398. [Google Scholar]
- Kelly, E.J.; Brownlee, K.G.; Newell, S.J. Gastric secretory function in the developing human stomach. Early Hum. Dev. 1992, 31, 163–166. [Google Scholar] [CrossRef]
- Kelly, E.J.; Lagopoulos, M.; Primrose, J.N. Immunocytochemical localisation of parietal cells and G cells in the developing human stomach. Gut 1993, 34, 1057–1059. [Google Scholar] [CrossRef] [PubMed]
- Hyman, P.E.; Clarke, D.D.; Everett, S.L.; Sonne, B.; Stewart, D.; Harada, T.; Walsh, J.H.; Taylor, I.L. Gastric acid secretory function in preterm infants. J. Pediatr. 1985, 106, 467–471. [Google Scholar] [CrossRef]
- Werner, B. Peptic and tryptic capacity of the digestive glands in newborns: A comparison between premature and full-term infants. Acta Paediatr. Jpn. 1948, 35 (Suppl. 5), 1. [Google Scholar]
- Antonowicz, I.; Lebenthal, E. Developmental Pattern of Small Intestinal Enterokinase and Disaccharidase Activities in the Human Fetus. Gastroenterology 1977, 72, 1299–1303. [Google Scholar] [CrossRef]
- Demers-Mathieu, V.; Underwood, M.A.; Borghese, R.; Dallas, D.C. Premature infants have lower gastric digestion capacity for human milk proteins than term infants. J. Pediatr. Gastroenterol. Nutr. 2018, 66, 816–821. [Google Scholar] [CrossRef]
- Mihatsch, W.A.; Franz, A.R.; Hogel, J.; Pohlandt, F. Hydrolyzed protein accelerates feeding advancement in very low birthweight infants. Pediatrics 2002, 110, 1199–1203. [Google Scholar] [CrossRef]
- Murray, R.D.; Kerzner, B.; Sloan, H.R.; McClung, H.J.; Gilbert, M.; Ailabouni, A. The Contribution of Salivary Amylase to Glucose Polymer Hydrolysis in Premature Infants. Pediatr. Res. 1986, 20, 186–191. [Google Scholar] [CrossRef]
- Davis, M.M.; Hodes, M.E.; Munsick, R.A.; Ulbright, T.M.; Goldstein, D.J. Pancreatic Amylase Expression in Human Pancreatic Development. Hybridoma 1986, 5, 137–145. [Google Scholar] [CrossRef] [PubMed]
- McClean, P.; Weaver, L.T. Ontogeny of human pancreatic exocrine function. Arch. Dis. Child. 1993, 68, 62–65. [Google Scholar] [CrossRef] [PubMed]
- Leonel, A.J.; Alvarez-Leite, J.I. Butyrate: Implications for intestinal function. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; de Roos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Schulman, R.J.; Schanler, R.J.; Lau, C.; Heitkemper, M.; Ou, C.N.; Smith, E.O. Early feeding, feeding tolerance, and lactase activity in preterm infants. J. Pediatr. 1998, 133, 645–649. [Google Scholar] [CrossRef]
- Boyle, J.T.; Celano, P.; Koldovský, O. Demonstration of a difference in expression of maximal lactase and sucrase activity along the villus in the adult rat jejunum. Gastroenterology 1980, 79, 503–507. [Google Scholar] [CrossRef]
- Neu, J.; Li, N. The Neonatal Gastrointestinal Tract: Developmental Anatomy, Physiology, and Clinical Implications. Neoreviews 2003, 4, e7–e13. [Google Scholar] [CrossRef]
- Shulman, R.J. Effect of enteral administration of insulin on intestinal development and feeding tolerance in preterm infants: A pilot study. Arch. Dis. Child. Fetal Neonatal Ed. 2002, 86, F131–F133. [Google Scholar] [CrossRef]
- Klenoff-Brumberg, H.L.; Genen, L.H. High versus low medium chain triglyceride content of formula for promoting short term growth of preterm neonates. Cochrane Database Syst. Rev. 2003, 1, CD002777. [Google Scholar]
- Freed, L.M.; York, C.M.; Homosh, P.; Mehta, N.R.; Hamosh, M. Bile salt-stimulated lipase of human milk: Characteristics of the enzyme in the milk of mothers of premature and full-term infants. J. Pediatr. Gastroenterol. Nutr. 1987, 6, 598–604. [Google Scholar] [CrossRef]
- Hamosh, M.; Scanlon, J.W.; Ganot, D.; Likel, M.; Scanlon, K.B.; Hamosh, P. Fat digestion in the newborn. Characterization of lipase in gastric aspirates of premature and term infants. J. Clin. Investig. 1981, 67, 838–846. [Google Scholar] [CrossRef] [PubMed]
- Zoppi, G.; Andreotti, G.; Pajno-Ferrara, F.; Njai, D.M.; Gaburro, D. Exocrine Pancreas Function in Premature and Full Term Neonates. Pediatr. Res. 1972, 6, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Balistreri, W.F. Immaturity of Hepatic Excretory Function and the Ontogeny of Bile Acid Metabolism. J. Pediatr. Gastroenterol. Nutr. 1983, 2 (Suppl. 1), 207–214. [Google Scholar] [CrossRef] [PubMed]
- Boehm, G.; Braun, W.; Moro, G.; Minoli, I. Bile acid concentrations in serum and duodenal aspirates of healthy preterm infants: Effects of gestational and postnatal age. Biol. Neonate 1997, 71, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Georgieff, M.K.; Innis, S.M. Controversial Nutrients That Potentially Affect Preterm Neurodevelopment: Essential Fatty Acids and Iron. Pediatr. Res. 2005, 57 Pt 2, 99R–103R. [Google Scholar] [CrossRef] [PubMed]
- Lapillonne, A.; Jensen, C.L. Reevaluation of the DHA requirement for the premature infant. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Huizinga, J.D.; Lammers, W.J.E.P. Gut peristalsis is governed by a multitude of cooperating mechanisms. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1–G8. [Google Scholar] [CrossRef]
- Sanders, K.M.; Ward, S.M.; Koh, S.D. Interstitial Cells: Regulators of Smooth Muscle Function. Physiol. Rev. 2014, 94, 859–907. [Google Scholar] [CrossRef]
- Commare, C.E.; Tapped, K.A. Development of the infant intestine: Implication for nutrition support. Nutr. Clin. Pract. 2007, 22, 159–173. [Google Scholar] [CrossRef]
- Neal-Kluever, A.; Fisher, J.; Grylack, L. Physiology of the Neonatal Gastrointestinal System Relevant to the Disposition of Orally Administered Medications. Drug Metab. Dispos. 2019, 47, 296–313. [Google Scholar] [CrossRef]
- Newell, S.J. Gastrointestinal function and its ontogeny: How should we feed the preterm infant? Semin. Neonatol. 1996, 1, 59–66. [Google Scholar] [CrossRef]
- Bisset, W.M.; Watt, J.B.; Rivers, R.P.; Milla, P.J. Ontogeny of fasting small intestinal motor activity in the human infant. Gut 1988, 29, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Riezzo, G.; Indrio, F.; Montagna, O.; Tripaldi, C.; Laforgia, N.; Chiloiro, M.; Mautone, A. Gastric electrical activity and gastric emptying in term and preterm newborns. Neurogastroenterol. Motil. 2000, 12, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Hasler, W.L. Motility of the small intestine and colon. In Textbook of Gastroenterology; Yamada, T., Ed.; Wiley-Blackwell: Philadelphia, PA, USA, 2009; pp. 231–263. [Google Scholar]
- Chen, J.D.; McCallum, R.W. Clinical applications of electrogastrography. Am. J. Gastroenterol. 1993, 88, 1324–1336. [Google Scholar] [PubMed]
- Sarna, S. In vivo myoelectrical activity: Methods, analysis, and interpretation. In Handbook of Physiology. The Gastrointestinal System; Schultz, S., Wood, J.D., Eds.; Waverly Press: Baltimore, MD, USA, 1988; pp. 817–863. [Google Scholar]
- Berseth, C. Gestational evolution of small intestine motility in preterm and term infants. J. Pediatr. 1989, 115, 646–651. [Google Scholar] [CrossRef]
- Ittmann, P.I.; Amarnath, R.; Berseth, C.L. Maturation of antroduodenal motor activity in preterm and term infants. Am. J. Dig. Dis. 1992, 37, 14–19. [Google Scholar] [CrossRef]
- McCann, C.; Alves, M.M.; Brosens, E.; Natarajan, D.; Perin, S.; Chapman, C.; Hofstra, R.M.; Burns, A.J.; Thapar, N. Neuronal Development and Onset of Electrical Activity in the Human Enteric Nervous System. Gastroenterology 2019, 156, 1483–1495. [Google Scholar] [CrossRef]
- Fu, M.; Tam, P.K.; Sham, M.H.; Lui, V.C.H. Embryonic development of the ganglion plexuses and the concentric layer structure of human gut: A topographical study. Anat. Embryol. 2004, 208, 33–41. [Google Scholar] [CrossRef]
- Luo, S.; Zhu, H.; Zhang, J.; Wan, D. The Pivotal Role of Microbiota in Modulating the Neuronal–Glial–Epithelial Unit. Infect. Drug Resist. 2021, 14, 5613–5628. [Google Scholar] [CrossRef]
- Heiss, C.N.; Olofsson, L.E. The role of the gut microbiota in development, function and disorders of the central nervous system and the enteric nervous system. J. Neuroendocrinol. 2019, 31, e12684. [Google Scholar] [CrossRef]
- Wallace, A.S.; Burns, A.J. Development of the enteric nervous system, smooth muscle and interstitial cells of Cajal in the human gastrointestinal tract. Cell Tissue Res. 2005, 319, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Burns, A.J.; Douarin, N.M. The sacral neural crest contributes neurons and glia to the post-umbilical gut: Spatio temporal analysis of the development of the enteric nervous system. Development 1998, 125, 4335–4347. [Google Scholar] [CrossRef] [PubMed]
- Kapur, R.; Yost, C.; Palmiter, R. A transgenic model for studying development of the enteric nervous system in normal and aganglionic mice. Development 1992, 116, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Young, H.M.; Hearn, C.J.; Ciampoli, D.; Southwell, B.R.; Brunet, J.F.; Newgreen, D.F. A Single Rostrocaudal Colonization of the Rodent Intestine by Enteric Neuron Precursors Is Revealed by the Expression of Phox2b, Ret, and p75 and by Explants Grown under the Kidney Capsule or in Organ Culture. Dev. Biol. 1998, 202, 67–84. [Google Scholar] [CrossRef]
- Huang, T.; Hou, Y.; Wang, X.; Wang, L.; Yi, C.; Wang, C.; Sun, X.; Tam, P.K.H.; Ngai, S.M.; Sham, M.H.; et al. Direct Interaction of Sox10 with Cadherin-19 Mediates Early Sacral Neural Crest Cell Migration: Implications for Enteric Nervous System Development Defects. Gastroenterology 2022, 162, 179–192.e11. [Google Scholar] [CrossRef]
- Nishiyama, C.; Uesaka, T.; Manabe, T.; Yonekura, Y.; Nagasawa, T.; Newgreen, D.F.; Young, H.M.; Enomoto, H. Trans-mesenteric neural crest cells are the principal source of the colonic enteric nervous system. Nat. Neurosci. 2012, 15, 1211–1218. [Google Scholar] [CrossRef]
- Lasrado, R.; Boesmans, W.; Kleinjung, J.; Pin, C.; Bell, D.; Bhaw, L.; McCallum, S.; Zong, H.; Luo, L.; Clevers, H.; et al. Lineage-dependent spatial and functional organization of the mammalian enteric nervous system. Science 2017, 356, 722–726. [Google Scholar] [CrossRef]
- Young, H.M.; Jones, B.R.; McKeown, S.J. The Projections of Early Enteric Neurons Are Influenced by the Direction of Neural Crest Cell Migration. J. Neurosci. 2002, 22, 6005–6018. [Google Scholar] [CrossRef]
- Ward, S.M.; Burns, A.J.; Torihashi, S.; Sanders, K.M. Mutation of the proto-oncogene c-kit blocks development of interstitial cells and electrical rhythmicity in murine intestine. J. Physiol. 1994, 480 Pt 1, 91–97. [Google Scholar] [CrossRef]
- Hulzinga, J.D.; Thuneberg, L.; Klüppel, M.; Malysz, J.; Mikkelsen, H.B.; Bernstein, A. W/kit gene required for interstitial cells of Cajal and for intestinal pacemaker activity. Nature 1995, 373, 347–349. [Google Scholar] [CrossRef]
- Klüppel, M.; Huizinga, J.D.; Malysz, J.; Bernstein, A. Developmental origin and kit-dependent development of the interstitial cells of cajal in the mammalian small intestine. Dev. Dyn. 1998, 211, 60–71. [Google Scholar] [CrossRef]
- Torihashi, S.; Ward, S.M.; Sanders, K.M. Development of c-Kitpositive cells and the onset of electrical rhythmicity in murine small intestine. Gastroenterology 1997, 112, 144–155. [Google Scholar] [CrossRef]
- Carmona, R.; Cano, E.; Mattiotti, A.; Gaztambide, J.; Muñoz-Chápuli, R. Cells Derived from the Coelomic Epithelium Contribute to Multiple Gastrointestinal Tissues in Mouse Embryos. PLoS ONE 2013, 8, e55890. [Google Scholar] [CrossRef] [PubMed]
- Jena, A.; Montoya, C.A.; Mullaney, J.A.; Dilger, R.N.; Young, W.; McNabb, W.C.; Roy, N.C. Gut-Brain Axis in the Early Postnatal Years of Life: A Developmental Perspective. Front. Integr. Neurosci. 2020, 14, 44. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The microbiota-gut-brain axis. Physiol Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef] [PubMed]
- Brett, B.E.; de Weerth, C. The microbiota–gut–brain axis: A promising avenue to foster healthy developmental outcomes. Dev. Psychobiol. 2019, 61, 772–782. [Google Scholar] [CrossRef]
- Collins, S.M.; Surette, M.; Bercik, P. The interplay between the intestinal microbiota and the brain. Nat. Rev. Microbiol. 2012, 10, 735–742. [Google Scholar] [CrossRef]
- Deoni, S.; Dean, D.; Joelson, S.; O’Regan, J.; Schneider, N. Early nutrition influences developmental myelination and cognition in infants and young children. NeuroImage 2018, 178, 649–659. [Google Scholar] [CrossRef]
- Erny, D.; Dokalis, N.; Mezö, C.; Castoldi, A.; Mossad, O.I.; Staszewski, O.; Frosch, M.; Villa, M.; Fuchs, V.; Mayer, A.; et al. Microbiota-derived acetate enables the metabolic fitness of the brain innate immune system duringhealth and disease. Cell Metab. 2021, 33, 2260–2276.e7. [Google Scholar] [CrossRef]
- Tosoni, G.; Conti, M.; Heijtz, R.D. Bacterial peptidoglycans as novel signaling molecules from microbiota to brain. Curr. Opin. Pharmacol. 2019, 48, 107–113. [Google Scholar] [CrossRef]
- Layunta, E.; Buey, B.; Mesonero, J.E.; Latorre, E. Crosstalk Between Intestinal Serotonergic System and Pattern Recognition Receptors on the Microbiota-Gut-Brain Axis. Front. Endocrinol. 2021, 12, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.C.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous Bacteria from the Gut Microbiota Regulate Host Serotonin Biosynthesis. Cell 2015, 161, 264–276, Erratum in Cell 2015, 163, 258. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Bresesti, I.; Salvatore, S.; Valetti, G.; Baj, A.; Giaroni, C.; Agosti, M. The Microbiota-Gut Axis in Premature Infants: Physio-Pathological Implications. Cells 2022, 11, 379. [Google Scholar] [CrossRef] [PubMed]
- Ratsika, A.; Codagnone, M.; O’Mahony, S.; Stanton, C.; Cryan, J. Priming for Life: Early Life Nutrition and the Microbiota-Gut-Brain Axis. Nutrients 2021, 13, 423. [Google Scholar] [CrossRef]
- Codagnone, M.G.; Spichak, S.; O’Mahony, S.M.; O’Leary, O.F.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Programming Bugs: Microbiota and the Developmental Origins of Brain Health and Disease. Biol. Psychiatry 2019, 85, 150–163. [Google Scholar] [CrossRef]
- Brink, L.R.; Lönnerdal, B. Milk fat globule membrane: The role of its various components in infant health and development. J. Nutr. Biochem. 2020, 85, 108465. [Google Scholar] [CrossRef]
- Al-Khafaji, A.H.; Jepsen, S.D.; Christensen, K.R.; Vigsnæs, L.K. The potential of human milk oligosaccharides to impact the microbiota-gut-brain axis through modulation of the gut microbiota. J. Funct. Foods 2020, 74, 104176. [Google Scholar] [CrossRef]
- Miller, J.; Tonkin, E.; Damarell, R.A.; McPhee, A.J.; Suganuma, M.; Suganuma, H.; Middleton, P.F.; Makrides, M.; Collins, C.T. A Systematic Review and Meta-Analysis of Human Milk Feeding and Morbidity in Very Low Birth Weight Infants. Nutrients 2018, 10, 707. [Google Scholar] [CrossRef]
- Lu, J.; Claud, E.C. Connection between gut microbiome and brain development in preterm infants. Dev. Psychobiol. 2019, 61, 739–751. [Google Scholar] [CrossRef]
- Carlson, A.L.; Xia, K.; Azcarate-Peril, M.A.; Goldman, B.D.; Ahn, M.; Styner, M.A.; Thompson, A.L.; Geng, X.; Gilmore, J.H.; Knickmeyer, R.C. Infant Gut Microbiome Associated with Cognitive Development. Biol. Psychiatry 2018, 83, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Rozé, J.C.; Ancel, P.Y.; Marchand-Martin, L.; Rousseau, C.; Montassier, E.; Monot, C.; Le Roux, K.; Butin, M.; Resche-Rigon, M.; Aires, A.; et al. Assessment of Neonatal Intensive Care Unit Practices and Preterm Newborn Gut Microbiota and 2-Year Neurodevelopmental Outcomes. JAMA Netw. Open 2020, 3, e2018119. [Google Scholar] [CrossRef] [PubMed]
- Seki, D.; Mayer, M.; Hausmann, B.; Pjevac, P.; Giordano, V.; Goeral, J.; Unterasinger, L.; Klebermaß-Schrehof, K.; De Paepe, K.; Van de Wiele, T.; et al. Aberrant gut-microbiota-immune-brain axis development in premature neonates with brain damage. Cell Host Microbe 2021, 29, 1558–1572.e6. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Salzwedel, A.P.; Carlson, A.L.; Xia, K.; Azcarate-Peril, M.A.; Styner, M.A.; Thompson, A.L.; Geng, X.; Goldman, B.D.; Gilmore, J.H.; et al. Gut microbiome and brain functional connectivity in infants-a preliminary study focusing on the amygdala. Psychopharmacology 2019, 236, 1641–1651. [Google Scholar] [CrossRef]
- Beghetti, I.; Barone, M.; Turroni, S.; Biagi, E.; Sansavini, A.; Brigidi, P.; Corvaglia, L.; Aceti, A. Early-life gut microbiota and neurodevelopment in preterm infants: Any role for Bifidobacterium? Eur. J. Pediatr. 2021. [Google Scholar] [CrossRef]
- Niemarkt, H.J.; De Meij, T.G.; Van Ganzewinkel, C.J.; de Boer, N.K.H.; Andriessen, P.; Hütten, M.C.; Kramer, B.W. Necrotizing Enterocolitis, Gut Microbiota, and Brain Development: Role of the Brain-Gut Axis. Neonatology 2019, 115, 423–431. [Google Scholar] [CrossRef]
- Ihekweazu, F.D.; Versalovic, J. Development of the Pediatric Gut Microbiome: Impact on Health and Disease. Am. J. Med. Sci. 2018, 356, 413–423. [Google Scholar] [CrossRef]
- Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef]
- Dimitrakopoulou, E.I.; Pouliakis, A.; Falaina, V.; Xanthos, T.; Zoumpoulakis, P.; Tsiaka, T.; Sokou, S.; Iliodromiti, Z.; Boutsikou, T.; Iacovidou, N. The Metagenomic and Metabolomic Profile of the Infantile Gut: Can They Be “Predicted” by the Feed Type? Children 2022, 9, 154. [Google Scholar] [CrossRef]
- Matara, D.-I.; Pouliakis, A.; Xanthos, T.; Sokou, R.; Kafalidis, G.; Iliodromiti, Z.; Boutsikou, T.; Iacovidou, N.; Salakos, C. Microbial Translocation and Perinatal Asphyxia/Hypoxia: A Systematic Review. Diagnostics 2022, 12, 214. [Google Scholar] [CrossRef]
- Willyard, C. Could baby’s first bacteria take root before birth? Nature 2018, 553, 264–266. [Google Scholar] [CrossRef] [PubMed]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The Placenta Harbors a Unique Microbiome. Sci. Transl. Med. 2014, 6, 237ra65. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, R.; Rani, A.; Metwally, A.; McGee, H.S.; Perkins, D.L. Analysis of the microbiome: Advantages of whole genome shotgun versus 16S amplicon sequencing. Biochem. Biophys. Res. Commun. 2016, 469, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Ahearn-Ford, S.; Berrington, J.E.; Stewart, C.J. Development of the gut microbiome in early life. Exp. Physiol. 2022. [Google Scholar] [CrossRef]
- Chu, D.M.; Antony, K.M.; Ma, J.; Prince, A.L.; Showalter, L.; Moller, M.; Aagaard, K.M. The early infant gut microbiome varies in association with a maternal high-fat diet. Genome Med. 2016, 8, 1–12. [Google Scholar] [CrossRef]
- Ferretti, P.; Pasolli, E.; Tett, A.; Asnicar, F.; Gorfer, V.; Fedi, S.; Armanini, F.; Truong, D.T.; Manara, S.; Zolfo, M.; et al. Mother-to-Infant Microbial Transmission from Different Body Sites Shapes the Developing Infant Gut Microbiome. Cell Host Microbe 2018, 24, 133–145.e5. [Google Scholar] [CrossRef]
- Stearns, J.C.; Zulyniak, M.A.; de Souza, R.J.; Campbell, N.C.; Fontes, M.; Shaikh, M.; Sears, M.R.; Becker, A.B.; Mandhane, P.J.; Subbarao, P.; et al. Ethnic and diet-related differences in the healthy infant microbiome. Genome Med. 2017, 9, 1–12. [Google Scholar] [CrossRef]
- Romano-Keeler, J.; Zhang, J.; Sun, J. COVID-19 and the neonatal microbiome: Will the pandemic cost infants their microbes? Gut Microbes 2021, 13, 1–7. [Google Scholar] [CrossRef]
- La Rosa, P.S.; Warner, B.B.; Zhou, Y.; Weinstock, G.M.; Sodergren, E.; Hall-Moore, C.M.; Stevens, H.J.; Bennett, W.E.; Shaikh, N.; Linneman, L.A.; et al. Patterned progression of bacterial populations in the premature infant gut. Proc. Natl. Acad. Sci. USA 2014, 111, 12522–12527. [Google Scholar] [CrossRef]
- Jia, J.; Xun, P.; Wang, X.; He, K.; Tang, Q.; Zhang, T.; Wang, Y.; Tang, W.; Lu, L.; Yan, W.; et al. Impact of Postnatal Antibiotics and Parenteral Nutrition on the Gut Microbiota in Preterm Infants During Early Life. J. Parenter. Enter. Nutr. 2020, 44, 639–654. [Google Scholar] [CrossRef]
- Henderickx, J.G.E.; Zwittink, R.D.; Van Lingen, R.A.; Knol, J.; Belzer, C. The Preterm Gut Microbiota: An Inconspicuous Challenge in Nutritional Neonatal Care. Front. Cell. Infect. Microbiol. 2019, 9, 85. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Lu, L.; Sun, J.; Petrof, E.O.; Claud, E.C. Preterm infant gut microbiota affects intestinal epithelial development in a humanized microbiome gnotobiotic mouse model. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G521–G532. [Google Scholar] [CrossRef] [PubMed]
- Battersby, C.; Santhalingam, T.; Costeloe, K.; Moodi, N. Incidence of neonatal necrotising enterocolitis in high-income countries: A systematic review. Arch. Dis. Child. Fetal Neonatal Ed. 2018, 103, F182–F189. [Google Scholar] [CrossRef] [PubMed]
- Henrick, B.M.; Rodriguez, L.; Lakshmikanth, T.; Pou, C.; Henckel, E.; Arzoomand, A.; Olin, A.; Wang, J.; Mikes, J.; Tan, Z.; et al. Bifidobacteria-mediated immune system imprinting early in life. Cell 2021, 184, 3884–3898.e11. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Speer, C.P. Late-onset neonatal sepsis: Recent developments. Arch. Dis. Child. Fetal Neonatal Ed. 2015, 100, F257–F263. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.J.; Ajami, N.J.; O’Brien, J.L.; Hutchinson, D.S.; Smith, D.P.; Wong, M.C.; Ross, M.C.; Lloyd, R.E.; Doddapaneni, H.; Metcalf, G.A.; et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature 2018, 562, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Kurath-Koller, S.; Neumann, C.; Moissl-Eichinger, C.; Kraschl, R.; Kanduth, C.; Hopfer, B.; Pausan, M.-R.; Urlesberger, B.; Resch, B. Hospital Regimens Including Probiotics Guide the Individual Development of the Gut Microbiome of Very Low Birth Weight Infants in the First Two Weeks of Life. Nutrients 2020, 12, 1256. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Indrio, F.; Neu, J.; Pettoello-Mantovani, M.; Marchese, F.; Martini, S.; Salatto, A.; Aceti, A. Development of the Gastrointestinal Tract in Newborns as a Challenge for an Appropriate Nutrition: A Narrative Review. Nutrients 2022, 14, 1405. https://doi.org/10.3390/nu14071405
Indrio F, Neu J, Pettoello-Mantovani M, Marchese F, Martini S, Salatto A, Aceti A. Development of the Gastrointestinal Tract in Newborns as a Challenge for an Appropriate Nutrition: A Narrative Review. Nutrients. 2022; 14(7):1405. https://doi.org/10.3390/nu14071405
Chicago/Turabian StyleIndrio, Flavia, Josef Neu, Massimo Pettoello-Mantovani, Flavia Marchese, Silvia Martini, Alessia Salatto, and Arianna Aceti. 2022. "Development of the Gastrointestinal Tract in Newborns as a Challenge for an Appropriate Nutrition: A Narrative Review" Nutrients 14, no. 7: 1405. https://doi.org/10.3390/nu14071405
APA StyleIndrio, F., Neu, J., Pettoello-Mantovani, M., Marchese, F., Martini, S., Salatto, A., & Aceti, A. (2022). Development of the Gastrointestinal Tract in Newborns as a Challenge for an Appropriate Nutrition: A Narrative Review. Nutrients, 14(7), 1405. https://doi.org/10.3390/nu14071405