
Anti-Inflammatory and Analgesic Effects of Schisandra chinensis Leaf Extracts and Monosodium Iodoacetate-Induced Osteoarthritis in Rats and Acetic Acid-Induced Writhing in Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of SCLE
2.2. Chemical Profiling of SCLE
2.3. Cell Culture
2.4. Cytotoxicity Assay
2.5. Determination of Nitric Oxide (NO), Prostaglandin E2 (PGE2), and Inflammatory Cytokines Production in the Cells
2.6. Acetic Acid-Induced Writhing Response in Mice
2.7. Development of MIA-Induced OA Rat Model
2.8. Hind Paw Weight-Bearing Distribution Measurements (Joint Pain Assessment)
2.9. Serum Analysis
2.10. Real-Time PCR Analysis
2.11. Histopathology Analysis
2.12. Statistical Analysis
3. Results
3.1. Chemical Profiling Analysis of SCLE
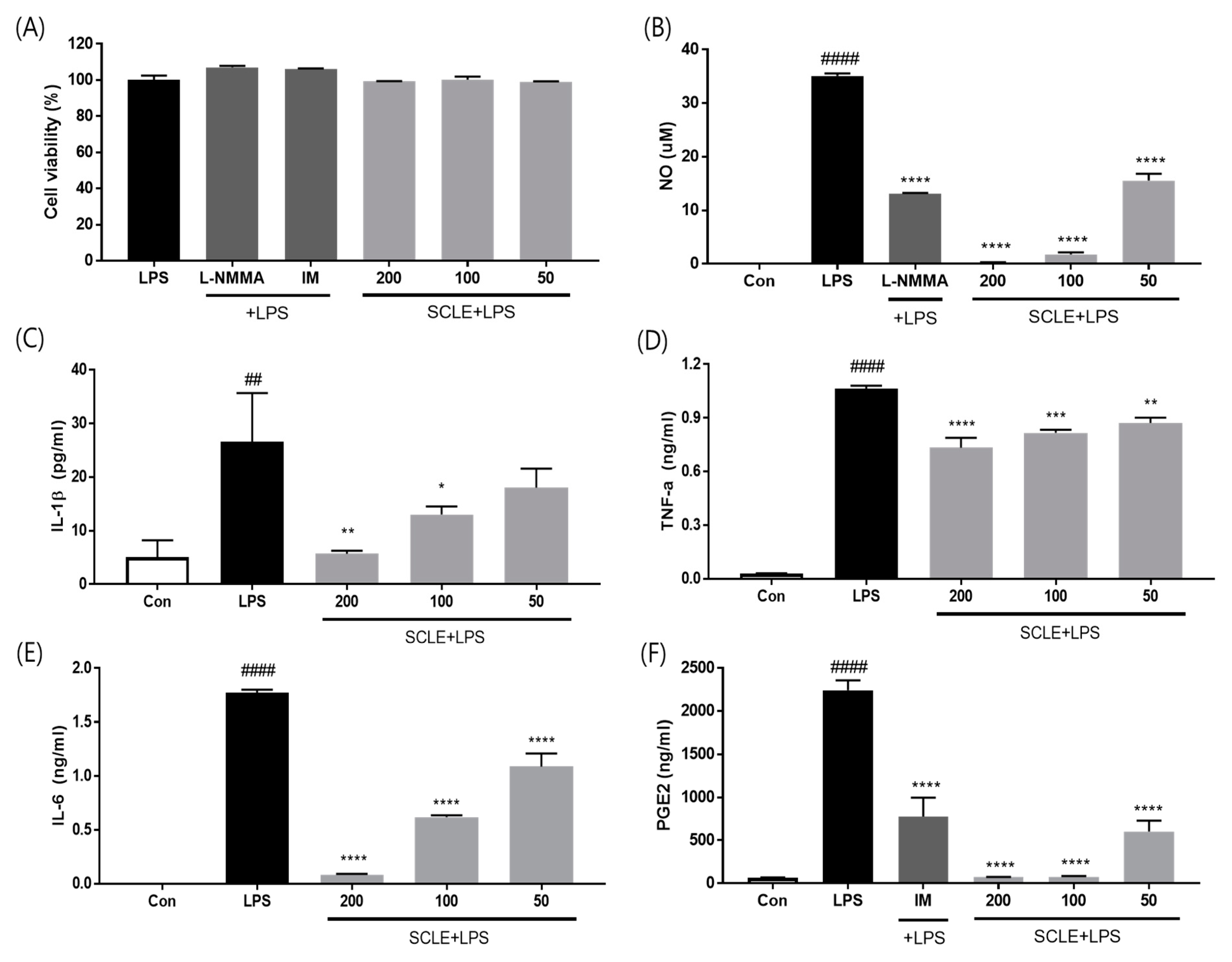
3.2. Effect of SCLE on LPS-Induced NO, IL-1β, TNF-α, IL-6, and PGE2 Production In Vitro
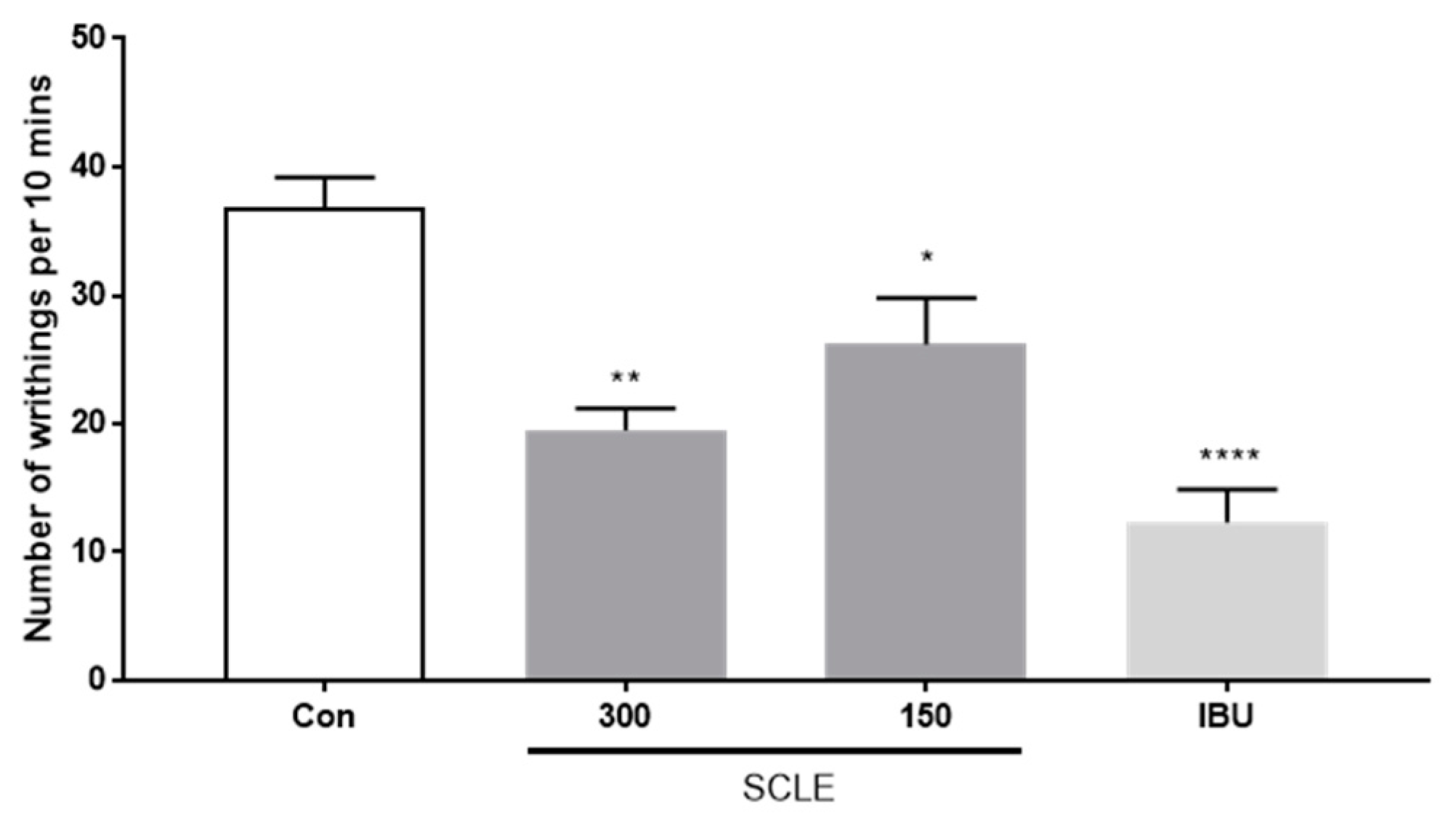
3.3. Anti-Analgesic Effect of SCLE on the Acetic Acid-Induced Writhing Response in Mice
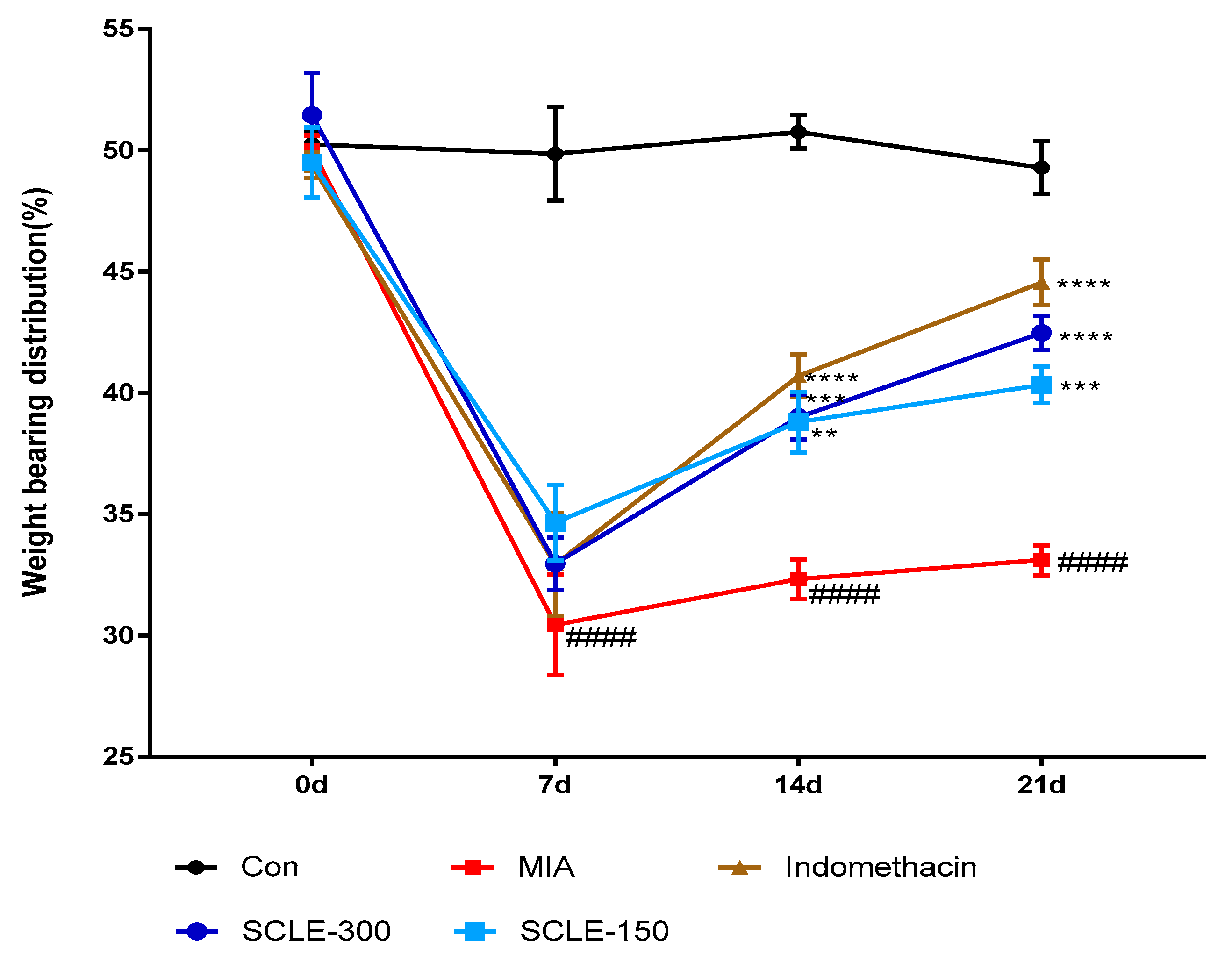
3.4. Effect of SCLE on Joint Pain in MIA-Induced Osteoarthritis in Rats
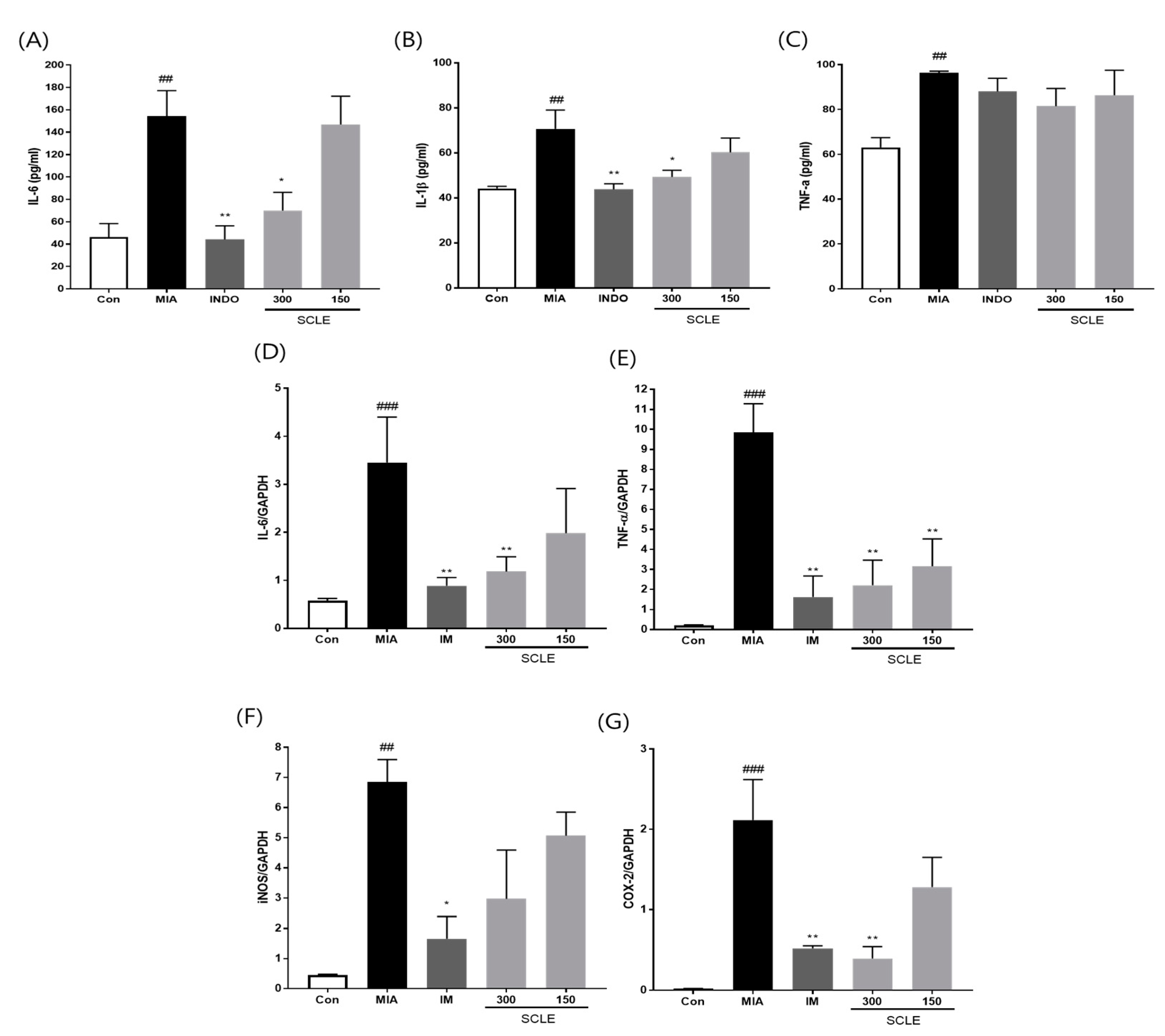
3.5. Effect of SCLE on Inflammatory Mediators and Cytokines in MIA-Induced Rats
3.6. Effect of SCLE on Matrix Degradation and Bone Metabolism in MIA-Induced Rats
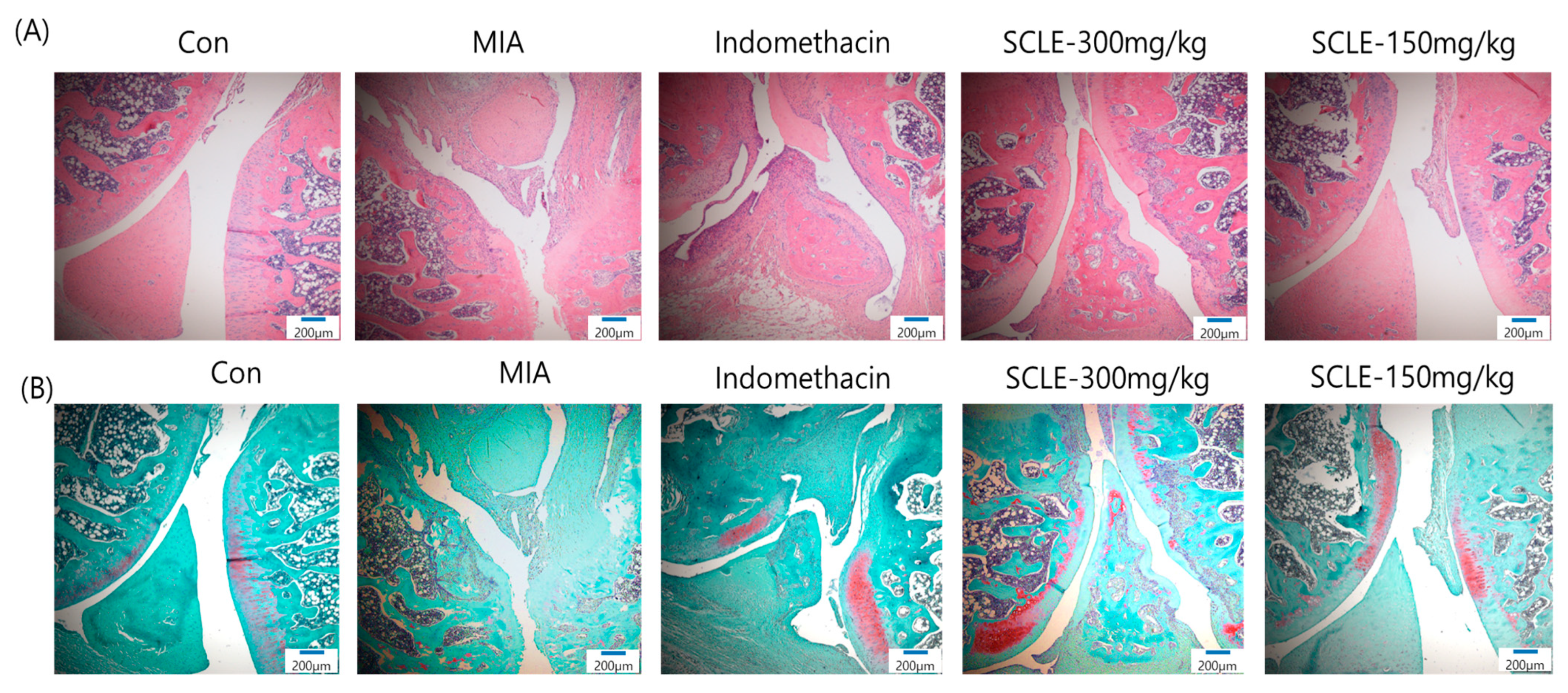
3.7. Effect of SCLE on Articular Cartilage Damage in MIA-Induced Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Robinson, W.H.; Lepus, C.M.; Wang, Q.; Raghu, H.; Mao, R.; Lindstrom, T.M.; Sokolove, J. Low-grade inflammation as a key mediator of the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 580–592. [Google Scholar] [PubMed]
- Grynpas, M.D.; Alpert, B.; Katz, I.; Lieberman, I.; Pritzker, K.P. Subchondral bone in osteoarthritis. Calcif. Tissue Int. 1991, 49, 20–26. [Google Scholar] [PubMed]
- Philp, A.M.; Davis, E.T.; Jones, S.W. Developing anti-inflammatory therapeutics for patients with osteoarthritis. Rheumatology 2017, 56, 869–881. [Google Scholar] [PubMed] [Green Version]
- Kapoor, M.; Martel-Pelletier, J.; Lajeunesse, D.; Pelletier, J.P.; Fahmi, H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat. Rev. Rheumatol. 2011, 7, 33–42. [Google Scholar] [PubMed]
- Buckwalter, J.A.; Brown, T.D. Joint injury, repair, and remodeling: Roles in post-traumatic osteoarthritis. Clin. Orthop. Relat. Res. 2004, 423, 7–16. [Google Scholar]
- Hunter, D.J. Pharmacologic therapy for osteoarthritis—The era of disease modification. Nat. Rev. Rheumatol. 2011, 7, 13–22. [Google Scholar] [PubMed]
- Che Ahmad Tantowi, N.A.; Lau, S.F.; Mohamed, S. Ficus deltoidea Prevented Bone Loss in Preclinical Osteoporosis/Osteoarthritis Model by Suppressing Inflammation. Calcif. Tissue Int. 2018, 103, 388–399. [Google Scholar] [PubMed]
- Sharma, V.K.; Mamontov, E.; Tyagi, M. Effects of NSAIDs on the nanoscopic dynamics of lipid membrane. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183100. [Google Scholar] [PubMed]
- Kang, Y.H.; Lee, H.J.; Lee, C.J.; Park, J.S. Natural Products as Sources of Novel Drug Candidates for the Pharmacological Management of Osteoarthritis: A Narrative Review. Biomol. Ther. 2019, 27, 503–513. [Google Scholar]
- Castrogiovanni, P.; Trovato, F.M.; Loreto, C.; Nsir, H.; Szychlinska, M.A.; Musumeci, G. Nutraceutical Supplements in the Management and Prevention of Osteoarthritis. Int. J. Mol. Sci. 2016, 17, 2042. [Google Scholar]
- Szopa, A.; Ekiert, R.; Ekiert, H. Current knowledge of Schisandra chinensis (Turcz.) Baill. (Chinese magnolia vine) as a medicinal plant species: A review on the bioactive components, pharmacological properties, analytical and biotechnological studies. Phytochem. Rev. 2017, 16, 195–218. [Google Scholar] [PubMed] [Green Version]
- Choi, H.; Seo, E.; Yeon, M.; Kim, M.S.; Hur, H.J.; Oh, B.C.; Jun, H.S. Anti-Aging Effects of Schisandrae chinensis Fructus Extract: Improvement of Insulin Sensitivity and Muscle Function in Aged Mice. Evid.-Based Complement. Altern. Med. 2019, 2019, 5642149. [Google Scholar]
- Lee, T.H.; Jung, C.H.; Lee, D.H. Neuroprotective effects of Schisandrin B against transient focal cerebral ischemia in Sprague-Dawley rats. Food Chem. Toxicol. 2012, 50, 4239–4245. [Google Scholar]
- Wat, E.; Ng, C.F.; Wong, E.C.; Koon, C.M.; Lau, C.P.; Cheung, D.W.; Fung, K.P.; Lau, C.B.; Leung, P.C. The hepatoprotective effect of the combination use of Fructus Schisandrae with statin—A preclinical evaluation. J. Ethnopharmacol. 2016, 178, 104–114. [Google Scholar] [PubMed]
- Kang, Y.S.; Han, M.H.; Hong, S.H.; Park, C.; Hwang, H.J.; Kim, B.W.; Kyoung, K.H.; Choi, Y.W.; Kim, C.M.; Choi, Y.H. Anti-inflammatory Effects of Schisandra chinensis (Turcz.) Baill. Fruit Through the Inactivation of Nuclear Factor-κB and Mitogen-activated Protein Kinases Signaling Pathways in Lipopolysaccharide-stimulated Murine Macrophages. J. Cancer Prev. 2014, 19, 279–287. [Google Scholar]
- Kang, J.S.; Han, M.H.; Kim, G.Y.; Kim, C.M.; Kim, B.W.; Hwang, H.J.; Hyun, Y. Nrf2-mediated HO-1 induction contributes to antioxidant capacity of a Schisandrae Fructus ethanol extract in C2C12 myoblasts. Nutrients 2014, 6, 5667–5678. [Google Scholar] [PubMed] [Green Version]
- Jeong, J.W.; Kim, J.W.; Ku, S.K.; Kim, S.G.; Kim, K.Y.; Kim, G.Y.; Hwang, H.J.; Kim, B.W.; Chung, H.Y.; Kim, C.M.; et al. Essential oils purified from Schisandrae semen inhibits tumor necrosis factor-α-induced matrix metalloproteinase-9 activation and migration of human aortic smooth muscle cells. BMC Complement. Altern. Med. 2015, 15, 7. [Google Scholar]
- Jeong, J.W.; Kim, J.; Choi, E.O.; Kwon, D.H.; Kong, G.M.; Choi, I.W.; Kim, B.H.; Kim, G.Y.; Lee, K.W.; Kim, K.Y.; et al. Schisandrae Fructus ethanol extract ameliorates inflammatory responses and articular cartilage damage in monosodium iodoacetate-induced osteoarthritis in rats. EXCLI J. 2017, 16, 265–277. [Google Scholar] [PubMed]
- Liu, G.Z.; Liu, Y.; Sun, Y.P.; Li, X.M.; Xu, Z.P.; Jiang, P.; Rong, X.H.; Yang, B.Y.; Kuang, H.X. Lignans and Terpenoids from the Leaves of Schisandra chinensis. Chem. Biodivers. 2020, 17, e2000035. [Google Scholar] [PubMed]
- Lee, Y.M.; Son, E.; Kim, S.H.; Kim, D.S. Effect of Alpinia oxyphylla extract in vitro and in a monosodium iodoacetate-induced osteoarthritis rat model. Phytomedicine 2019, 65, 153095. [Google Scholar]
- Whittle, B.A. The Use of Changes in Capillary Permeability in Mice to Distinguish between Narcotic and Nonnarcotic Alalgesics. Br. J. Pharmacol. Chemother. 1964, 22, 246–253. [Google Scholar]
- Guzman, R.E.; Evans, M.G.; Bove, S.; Morenko, B.; Kilgore, K. Mono-iodoacetate-induced histologic changes in subchondral bone and articular cartilage of rat femorotibial joints: An animal model of osteoarthritis. Toxicol. Pathol. 2003, 31, 619–624. [Google Scholar] [PubMed]
- McDougall, J.J.; Watkins, L.; Li, Z. Vasoactive intestinal peptide (VIP) is a modulator of joint pain in a rat model of osteoarthritis. Pain 2006, 123, 98–105. [Google Scholar] [PubMed]
- Lee, Y.M.; Son, E.; Kim, S.H.; Kim, O.S.; Kim, D.S. Anti-inflammatory and anti-osteoarthritis effect of Mollugo pentaphylla extract. Pharm. Biol. 2019, 57, 74–81. [Google Scholar]
- Chen, D.; Shen, J.; Zhao, W.; Wang, T.; Han, L.; Hamilton, J.L.; Im, H.J. Osteoarthritis: Toward a comprehensive understanding of pathological mechanism. Bone Res. 2017, 5, 16044. [Google Scholar] [PubMed]
- Manicone, A.M.; McGuire, J.K. Matrix metalloproteinases as modulators of inflammation. Semin. Cell Dev. Biol. 2008, 19, 34–41. [Google Scholar] [PubMed] [Green Version]
- Jeong, J.W.; Lee, H.H.; Choi, E.O.; Lee, K.W.; Kim, K.Y.; Kim, S.G.; Hong, S.H.; Kim, G.Y.; Park, C.; Kim, H.K.; et al. Schisandrae Fructus Inhibits IL-1β-Induced Matrix Metalloproteinases and Inflammatory Mediators Production in SW1353 Human Chondrocytes by Suppressing NF-κB and MAPK Activation. Drug Dev. Res. 2015, 76, 474–483. [Google Scholar] [PubMed]
- Ashraf, S.; Radhi, M.; Gowler, P.; Burston, J.J.; Gandhi, R.D.; Thorn, G.J.; Piccinini, A.M.; Walsh, D.A.; Chapman, V.; de Moor, C.H. The polyadenylation inhibitor cordycepin reduces pain, inflammation and joint pathology in rodent models of osteoarthritis. Sci. Rep. 2019, 9, 4696. [Google Scholar] [PubMed]
- Fernandes, J.C.; Martel-Pelletier, J.; Pelletier, J.P. The role of cytokines in osteoarthritis pathophysiology. Biorheology 2002, 39, 237–246. [Google Scholar] [PubMed]
- Kumavat, R.; Kumar, V.; Malhotra, R.; Pandit, H.; Jones, E.; Ponchel, F.; Biswas, S. Biomarkers of Joint Damage in Osteoarthritis: Current Status and Future Directions. Mediat. Inflamm. 2021, 2021, 5574582. [Google Scholar]
- Liu, S.; Deng, Z.; Chen, K.; Jian, S.; Zhou, F.; Yang, Y.; Fu, Z.; Xie, H.; Xiong, J.; Zhu, W. Cartilage tissue engineering: From proinflammatory and antiinflammatory cytokines to osteoarthritis treatments (Review). Mol. Med. Rep. 2022, 25, 99. [Google Scholar] [PubMed]
- Wojdasiewicz, P.; Poniatowski, L.A.; Szukiewicz, D. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediat. Inflamm. 2014, 2014, 561459. [Google Scholar]
- Shin, M.R.; Lee, J.A.; Kim, M.J.; Park, H.J.; Park, B.W.; Seo, S.B.; Roh, S.S. Protective Effects of Phellinus linteus Mycelium on the Development of Osteoarthritis after Monosodium Iodoacetate Injection. Evid.-Based Complement. Altern. Med. 2020, 2020, 7240858. [Google Scholar]
- Mixon, A.; Savage, A.; Bahar-Moni, A.S.; Adouni, M.; Faisal, T. An in vitro investigation to understand the synergistic role of MMPs-1 and 9 on articular cartilage biomechanical properties. Sci. Rep. 2021, 11, 14409. [Google Scholar] [PubMed]
- Han, S.; Gao, H.; Chen, S.; Wang, Q.; Li, X.; Du, L.J.; Li, J.; Luo, Y.Y.; Li, J.X.; Zhao, L.C.; et al. Procyanidin A1 Alleviates Inflammatory Response induced by LPS through NF-κB, MAPK, and Nrf2/HO-1 Pathways in RAW264.7 cells. Sci. Rep. 2019, 9, 15087. [Google Scholar] [PubMed] [Green Version]
- Pitcher, T.; Sousa-Valente, J.; Malcangio, M. The Monoiodoacetate Model of Osteoarthritis Pain in the Mouse. J. Vis. Exp. 2016, 111, 53746. [Google Scholar]
- Syx, D.; Tran, P.B.; Miller, R.E.; Malfait, A.M. Peripheral Mechanisms Contributing to Osteoarthritis Pain. Curr. Rheumatol. Rep. 2018, 20, 9. [Google Scholar]
- Lee, D.; Ju, M.K.; Kim, H. Commiphora Extract Mixture Ameliorates Monosodium Iodoacetate-Induced Osteoarthritis. Nutrients 2020, 12, 1477. [Google Scholar]
- Zhuang, Z.; Ye, G.; Huang, B. Kaempferol Alleviates the Interleukin-1β-Induced Inflammation in Rat Osteoarthritis Chondrocytes via Suppression of NF-κB. Med. Sci. Monit. 2017, 23, 3925–3931. [Google Scholar] [PubMed] [Green Version]
- Hu, Y.; Gui, Z.; Zhou, Y.; Xia, L.; Lin, K.; Xu, Y. Quercetin alleviates rat osteoarthritis by inhibiting inflammation and apoptosis of chondrocytes, modulating synovial macrophages polarization to M2 macrophages. Free Radic. Biol. Med. 2019, 145, 146–160. [Google Scholar]
- Tu, C.; Huang, X.; Xiao, Y.; Song, M.; Ma, Y.; Yan, J.; You, H.; Wu, H. Schisandrin A Inhibits the IL-1β-Induced Inflammation and Cartilage Degradation via Suppression of MAPK and NF-κB Signal Pathways in Rat Chondrocytes. Front. Pharmacol. 2019, 10, 41. [Google Scholar]
- Ran, J.; Ma, C.; Xu, K.; Xu, L.; He, Y.; Moqbel, S.A.A.; Hu, P.; Jiang, L.; Chen, W.; Bao, J.; et al. Schisandrin B ameliorated chondrocytes inflammation and osteoarthritis via suppression of NF-κB and MAPK signal pathways. Drug Des. Dev. Ther. 2018, 12, 1195–1204. [Google Scholar]
- Liao, S.; Zhou, K.; Li, D.; Xie, X.; Jun, F.; Wang, J. Schisantherin A suppresses interleukin-1β-induced inflammation in human chondrocytes via inhibition of NF-κB and MAPKs activation. Eur. J. Pharmacol. 2016, 780, 65–70. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | |
|---|---|---|
| IL-1β | Forward | 5′-ACAAGGCTGCCCCGACTA T-3′ |
| Reverse | 5′-CTCCTGGTATGAAGTGGCAAATC-3′ | |
| IL-6 | Forward | 5′-GCC CTT CAG GAA CAG CTA TGA-3′ |
| Reverse | 5′-TGTCAACAACATCAGTCCCAAGA-3′ | |
| COX-2 | Forward | 5′-CCTCGTCCAGATGCTATCTTTG-3′ |
| Reverse | 5′-GAAGGTCGTAGGTTTCCAGTATT-3′ | |
| iNOS | Forward | 5′-CTTTACGCCACTAACAGTGGCA-3′ |
| Reverse | 5′-AGTCATGCTTCCCATCGCTC-3′ | |
| MMP-2 | Forward | 5′-CACCAAGAACTTCCGACTATCC-3′ |
| Reverse | 5′-TCCAGTACCAGTGTCAGTATCA-3′ | |
| MMP-9 | Forward | 5′-CCCAACCTTTACCAGCTACTC-3′ |
| Reverse | 5′-GTCAGAACCGACCCTACAAAG-3′ | |
| GAPDH | Forward | 5′-TGGCCTCCAAGGAGTAAGAAAC-3′ |
| Reverse | 5′-CAG CAA CTG AGG GCC TCT CT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.M.; Son, E.; Kim, S.-H.; Kim, D.-S. Anti-Inflammatory and Analgesic Effects of Schisandra chinensis Leaf Extracts and Monosodium Iodoacetate-Induced Osteoarthritis in Rats and Acetic Acid-Induced Writhing in Mice. Nutrients 2022, 14, 1356. https://doi.org/10.3390/nu14071356
Lee YM, Son E, Kim S-H, Kim D-S. Anti-Inflammatory and Analgesic Effects of Schisandra chinensis Leaf Extracts and Monosodium Iodoacetate-Induced Osteoarthritis in Rats and Acetic Acid-Induced Writhing in Mice. Nutrients. 2022; 14(7):1356. https://doi.org/10.3390/nu14071356
Chicago/Turabian StyleLee, Yun Mi, Eunjung Son, Seung-Hyung Kim, and Dong-Seon Kim. 2022. "Anti-Inflammatory and Analgesic Effects of Schisandra chinensis Leaf Extracts and Monosodium Iodoacetate-Induced Osteoarthritis in Rats and Acetic Acid-Induced Writhing in Mice" Nutrients 14, no. 7: 1356. https://doi.org/10.3390/nu14071356
APA StyleLee, Y. M., Son, E., Kim, S.-H., & Kim, D.-S. (2022). Anti-Inflammatory and Analgesic Effects of Schisandra chinensis Leaf Extracts and Monosodium Iodoacetate-Induced Osteoarthritis in Rats and Acetic Acid-Induced Writhing in Mice. Nutrients, 14(7), 1356. https://doi.org/10.3390/nu14071356