
Linseed, Baru, and Coconut Oils: NMR-Based Metabolomics, Leukocyte Infiltration Potential In Vivo, and Their Oil Characterization. Are There Still Controversies?

 ,
,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Vegetable Oils
2.1.1. Raw Material
Fatty Acid Profiles and Indices of Nutritional Quality
Carotenoid Levels
Quality and Identity Analyses of Oils
Optical Characterization
Oxidative Stability
Thermogravimetric Analysis (TGA)
2.1.2. In Vivo Analyses
Animal and Experimental Design
Morphometric and Histological Parameters
Serum Analyses
NMR-Based Metabolomic Analysis
Leukocyte Recruitment Induced by Carrageenan (Cg)
2.2. Statistical Analyses
3. Results
3.1. Fatty Acid Profiles and Nutritional Quality of Oils
3.2. Analytical Oils Approach
3.2.1. Quality and Oils Stabilities
3.2.2. Optical Characterization
3.2.3. Thermogravimetric Analyses
3.3. Experimental Study
3.3.1. Morphometric Parameters
3.3.2. Effects of Oils on Liver Tissue
3.3.3. Biochemical Profiles
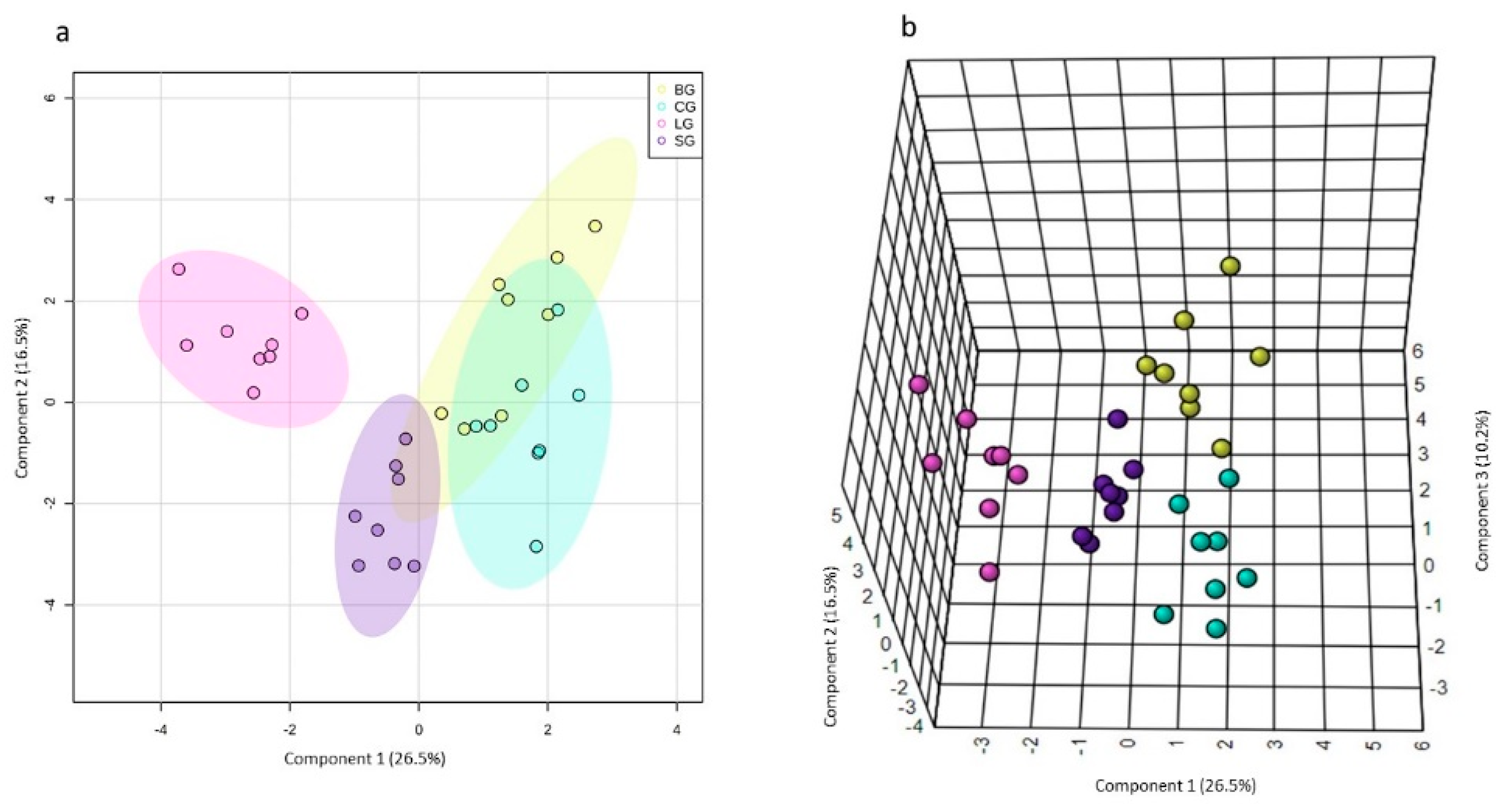
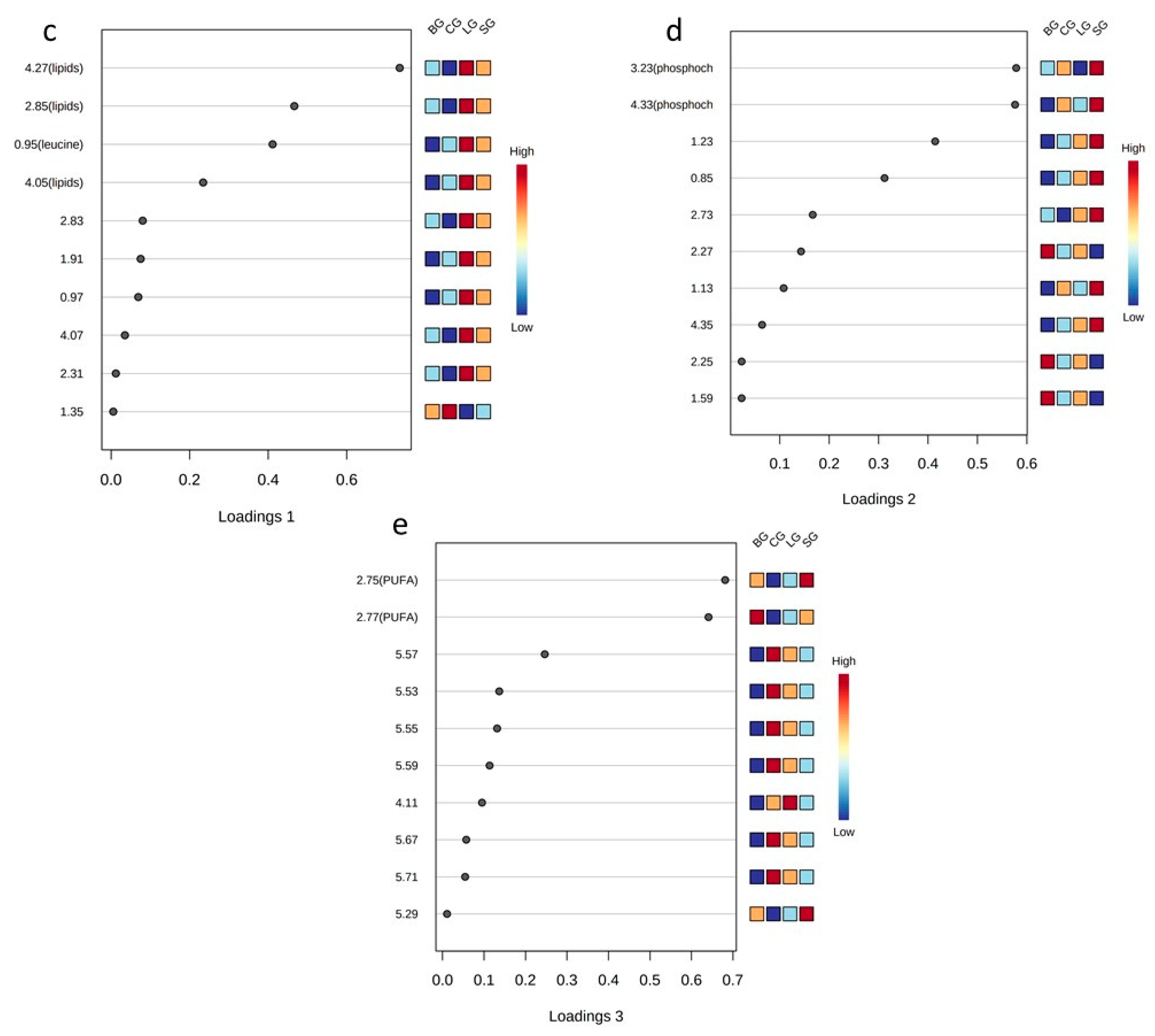
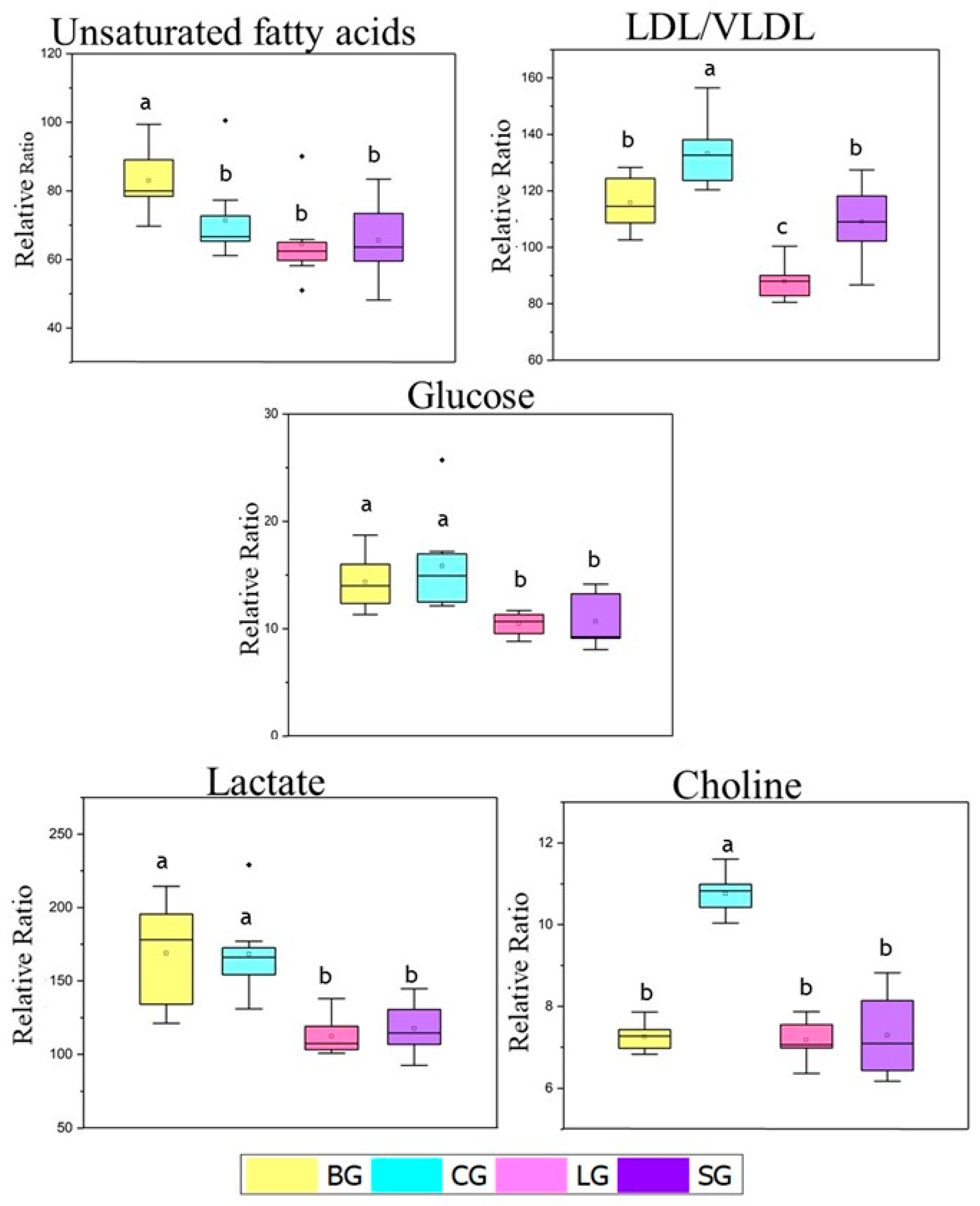
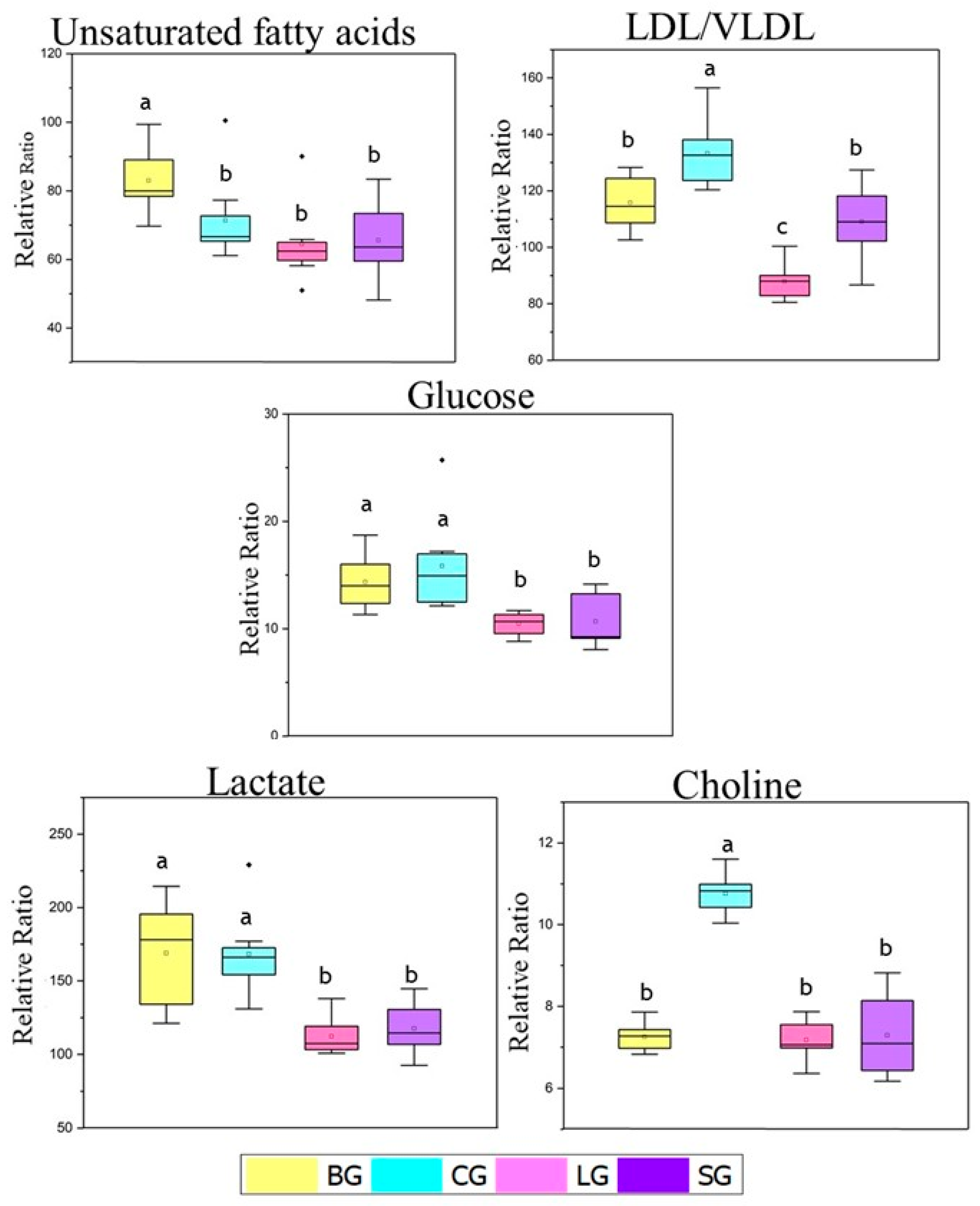
3.3.4. NMR-Based Metabolomics of the Serum Samples
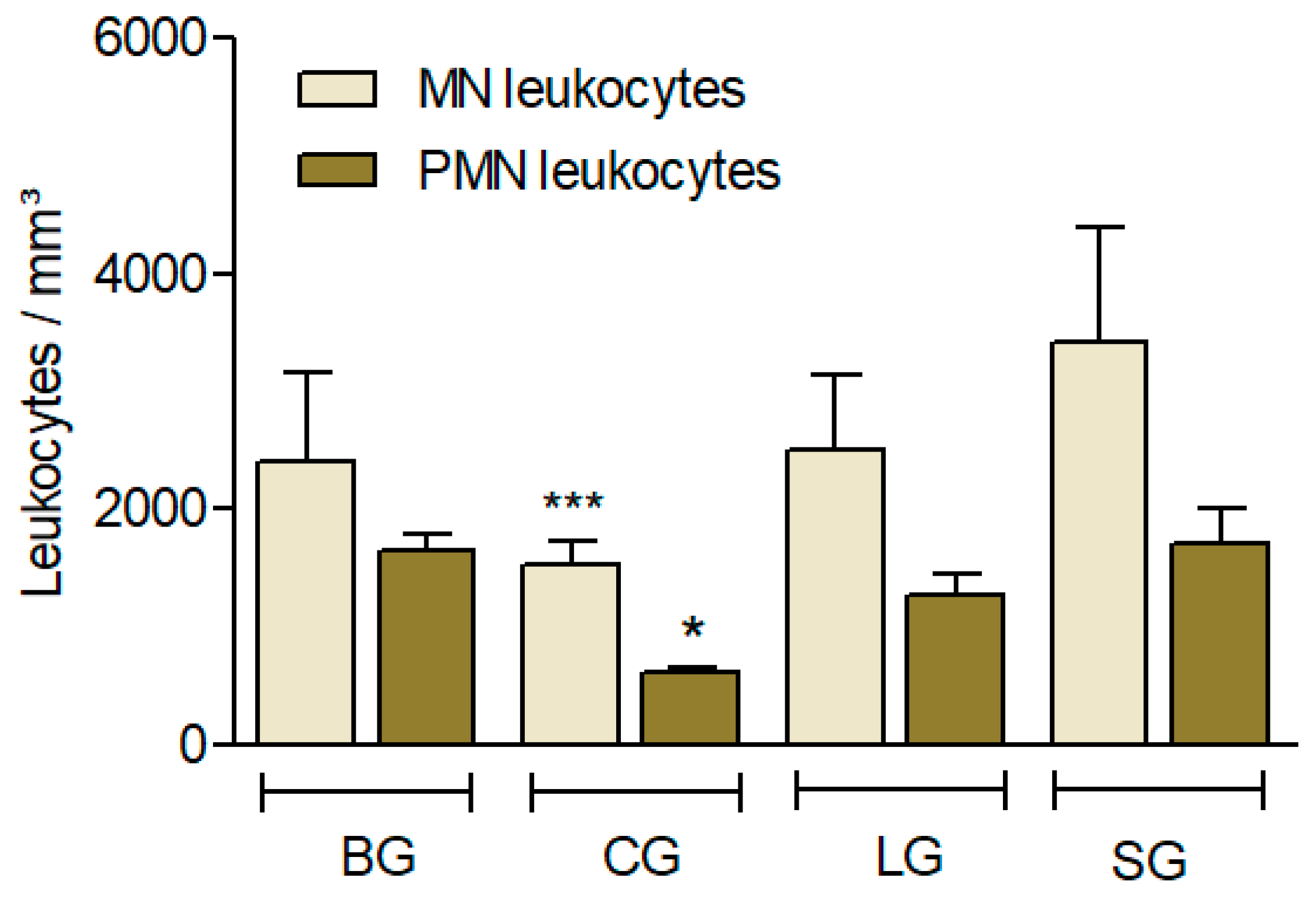
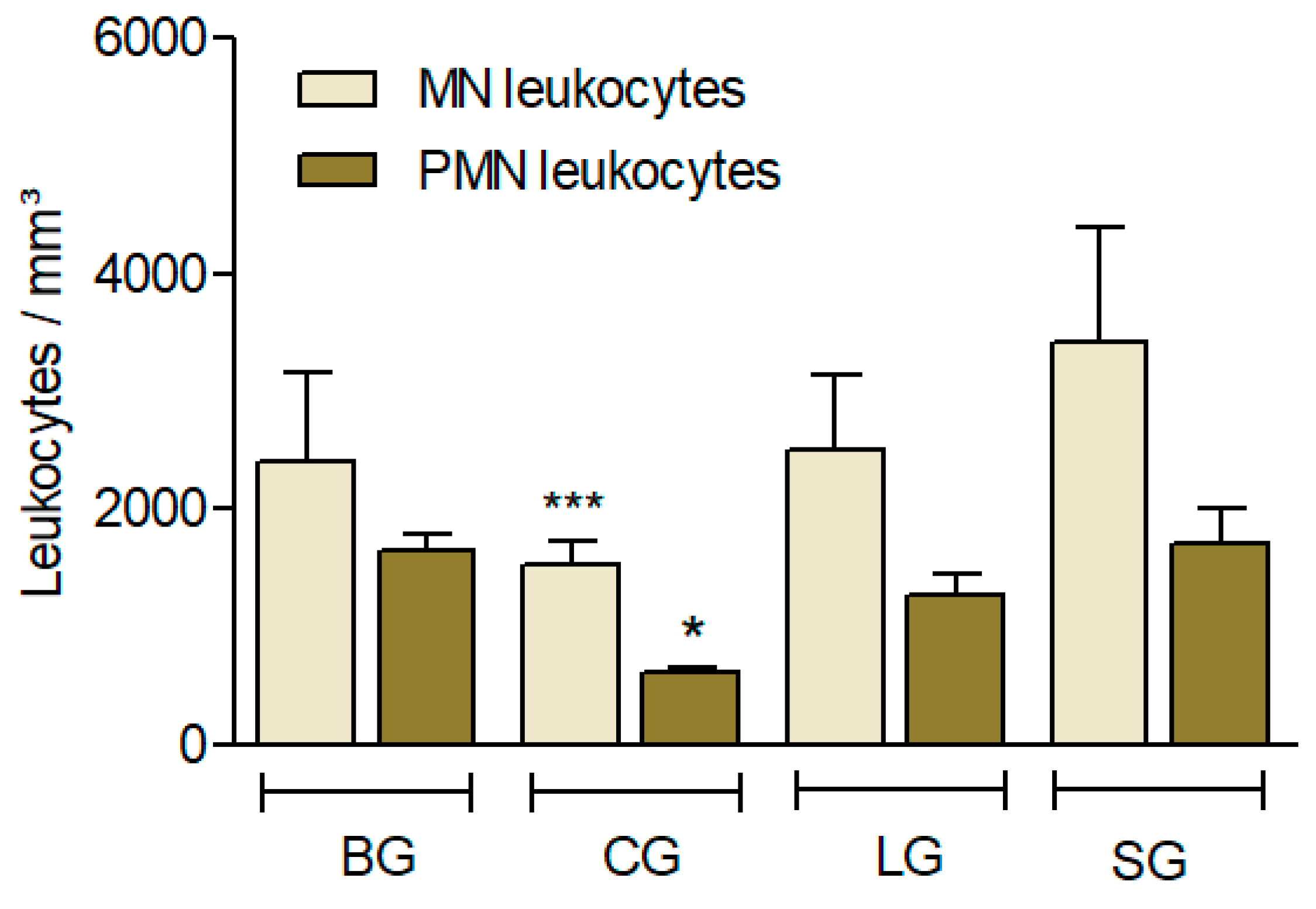
3.3.5. Leukocyte Recruitment Induced by Carrageenan
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Lenighan, Y.M.; McNulty, B.A.; Roche, H.M. Dietary Fat Composition: Replacement of Saturated Fatty Acids with PUFA as a Public Health Strategy, with an Emphasis on α-Linolenic Acid. Proc. Nutr. Soc. 2019, 78, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Garaulet, M.; Hernandez-Morante, J.J.; Lujan, J.; Tebar, F.J.; Zamora, S. Relationship between Fat Cell Size and Number and Fatty Acid Composition in Adipose Tissue from Different Fat Depots in Overweight/Obese Humans. Int. J. Obes. 2006, 30, 899–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, M.M. Subcutaneous and Visceral Adipose Tissue: Structural and Functional Differences. Obes. Rev. 2010, 11, 11–18. [Google Scholar] [CrossRef]
- Yang, L.G.; Song, Z.X.; Yin, H.; Wang, Y.Y.; Shu, G.F.; Lu, H.X.; Wang, S.K.; Sun, G.J. Low N-6/n-3 PUFA Ratio Improves Lipid Metabolism, Inflammation, Oxidative Stress and Endothelial Function in Rats Using Plant Oils as n-3 Fatty Acid Source. Lipids 2016, 51, 49–59. [Google Scholar] [CrossRef]
- Graham, C.B.; Philip, C.C. Conversion of Alpha-Linolenic Acid to Longer-Chain Polyunsaturated Fatty Acids in Human Adults. Available online: https://pubmed.ncbi.nlm.nih.gov/16188209/ (accessed on 25 July 2020).
- USDA Oilseeds: World Markets and Trade|USDA Foreign Agricultural Service. Available online: https://www.ers.usda.gov/data-products/oil-crops-yearbook/oil-crops-yearbook/#World%20Supply%20and%20Use%20of%20Oilseeds%20and%20Oilseed%20Products (accessed on 26 July 2020).
- Reis, M.Á.; Novaes, R.D.; Baggio, S.R.; Viana, A.L.M.; Salles, B.C.C.; Duarte, S.M.D.S.; Rodrigues, M.R.; Paula, F.B.D.A. Hepatoprotective and Antioxidant Activities of Oil from Baru Almonds (Dipteryx Alata Vog.) in a Preclinical Model of Lipotoxicity and Dyslipidemia. Evid.-Based Complement. Altern. Med. 2018, 2018, 8376081. [Google Scholar] [CrossRef] [Green Version]
- El Riachy, M.; Hamade, A.; Ayoub, R.; Dandachi, F.; Chalak, L. Oil Content, Fatty Acid and Phenolic Profiles of Some Olive Varieties Growing in Lebanon. Front. Nutr. 2019, 6, 94. [Google Scholar] [CrossRef] [PubMed]
- Terés, S.; Barceló-Coblijn, G.; Benet, M.; Álvarez, R.; Bressani, R.; Halver, J.E.; Escribá, P.V. Oleic Acid Content Is Responsible for the Reduction in Blood Pressure Induced by Olive Oil. Proc. Natl. Acad. Sci. USA 2008, 105, 13811–13816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babu, A.S.; Veluswamy, S.K.; Arena, R.; Guazzi, M.; Lavie, C.J. Virgin Coconut Oil and Its Potential Cardioprotective Effects. Postgrad. Med. 2014, 126, 76–83. [Google Scholar] [CrossRef]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary Heart Disease: Seven Dietary Factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B.; Kimura, M. HarvestPlus Handbook for Carotenoid Analysis; International Food Policy Research Institute (IFPRI) International Center for Tropical Agriculture (CIAT): Washington, DC, USA, 2004; 58p. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, 21st ed.; Latimer, J.D.G.W., Ed.; AOAC International: Rockville, MD, USA, 2019; Volume 3, ISBN 978-0-935584-89-9. [Google Scholar]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C. AIN-93 Purified Diets for Laboratory Rodents: Final Report of the American Institute of Nutrition Ad Hoc Writing Committee on the Reformulation of the AIN-76A Rodent Diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Taylor, B.A.; Phillips, S.J. Detection of Obesity QTLs on Mouse Chromosomes 1 and 7 by Selective DNA Pooling. Genomics 1996, 34, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Hagen, J.H.; Hagen, P.B. An Enzymic Method for the Estimation of Glycerol in Blood and Its Use to Determine the Effect of Noradrenaline on the Concentration of Glycerol in Blood. Can. J. Biochem. Physiol. 1962, 40, 1129–1139. [Google Scholar] [CrossRef] [PubMed]
- Carey, R.N.; Feldbruegge, D.; Westgard, J.O. Evaluation of the Adaptation of the Glucose Oxidase/Peroxidase-3-Methyl-2-Benzothiazolinone Hydrazone-N, N-Dimethylaniline Procedure to the Technicon SMA 12/60, and Comparison with Other Automated Methods for Glucose. Clin. Chem. 1974, 20, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Flegg, H.M. Ames Award Lecture 1972. An Investigation of the Determination of Serum Cholesterol by an Enzymatic Method. Ann. Clin. Biochem. 1973, 10, 79–84. [Google Scholar] [CrossRef]
- de Souza, G.E.P.; Ferreira, S.H. Blockade by Antimacrophage Serum of the Migration of PMN Neutrophils into the Inflamed Peritoneal Cavity. Agents Actions 1985, 17, 97–103. [Google Scholar] [CrossRef]
- O’Brien, R.D. Fats and Oils: Formulating and Processing for Applications, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2008; ISBN 978-1-4200-6167-3. [Google Scholar]
- SECTION 2. Codex Standards for Fats and Oils from Vegetable Sources. Available online: http://www.fao.org/3/y2774e/y2774e04.htm (accessed on 19 August 2020).
- Houshia, O.J.; Zaid, O.; Shqair, H.; Zaid, M.; Fashafsheh, N.; Bzoor, R. Effect of Olive Oil Adulteration on Peroxide Value, Delta-K and on the Acidity Nabali-Baladi Olive Oil Quality. Adv. Life Sci. 2014, 4, 235–244. [Google Scholar]
- De Oliveira, I.P.; Correa, W.A.; Neves, P.V.; Silva, P.V.B.; Lescano, C.H.; Michels, F.S.; Passos, W.E.; Muzzi, R.M.; Oliveira, S.L.; Caires, A.R.L. Optical Analysis of the Oils Obtained from Acrocomia Aculeata (Jacq.) Lodd: Mapping Absorption-Emission Profiles in an Induced Oxidation Process. Photonics 2017, 4, 3. [Google Scholar] [CrossRef]
- Magalhães, K.F.; Caires, A.R.L.; Silva, M.S.; Alcantara, G.B.; Oliveira, S.L. Endogenous Fluorescence of Biodiesel and Products Thereof: Investigation of the Molecules Responsible for This Effect. Fuel 2014, 119, 120–128. [Google Scholar] [CrossRef]
- Figueiredo, P.S.; Candido, C.J.; Jaques, J.A.; Nunes, Â.A.; Caires, A.R.; Michels, F.S.; Almeida, J.A.; Filiú, W.F.; Hiane, P.A.; Nascimento, V.A.; et al. Oxidative Stability of Sesame and Flaxseed Oils and Their Effects on Morphometric and Biochemical Parameters in an Animal Model. J. Sci. Food Agric. 2017, 97, 3359–3364. [Google Scholar] [CrossRef]
- Melo, E.; Michels, F.; Arakaki, D.; Lima, N.; Gonçalves, D.; Cavalheiro, L.; Oliveira, L.; Caires, A.; Hiane, P.; Nascimento, V. First Study on the Oxidative Stability and Elemental Analysis of Babassu (Attalea Speciosa) Edible Oil Produced in Brazil Using a Domestic Extraction Machine. Molecules 2019, 24, 4235. [Google Scholar] [CrossRef] [Green Version]
- Garcia, C.C.; Franco, P.I.B.M.; Zuppa, T.O.; Antoniosi Filho, N.R.; Leles, M.I.G. Thermal Stability Studies of Some Cerrado Plant Oils. J. Therm. Anal. Calorim. 2007, 87, 645–648. [Google Scholar] [CrossRef]
- Gerbaix, M.; Metz, L.; Ringot, E.; Courteix, D. Visceral Fat Mass Determination in Rodent: Validation of Dual-Energy x-Ray Absorptiometry and Anthropometric Techniques in Fat and Lean Rats. Lipids Health Dis. 2010, 9, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degirolamo, C.; Shelness, G.S.; Rudel, L.L. LDL Cholesteryl Oleate as a Predictor for Atherosclerosis: Evidence from Human and Animal Studies on Dietary Fat. J. Lipid Res. 2009, 50, S434–S439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanai, H.; Katsuyama, H.; Hamasaki, H.; Abe, S.; Tada, N.; Sako, A. Effects of Dietary Fat Intake on HDL Metabolism. J. Clin. Med. Res. 2015, 7, 145–149. [Google Scholar] [CrossRef] [Green Version]
- Lottering, R.T.; Govender, M.; Peerbhay, K.; Lottering, S. Comparing Partial Least Squares (PLS) Discriminant Analysis and Sparse PLS Discriminant Analysis in Detecting and Mapping Solanum Mauritianum in Commercial Forest Plantations Using Image Texture. ISPRS J. Photogramm. Remote Sens. 2020, 159, 271–280. [Google Scholar] [CrossRef]
- Bernhard, W.; Poets, C.; Franz, A. Choline and Choline-Related Nutrients in Regular and Preterm Infant Growth. Eur. J. Nutr. 2019, 58, 931–945. [Google Scholar] [CrossRef]
- Abdelhamid, A.S.; Martin, N.; Bridges, C.; Brainard, J.S.; Wang, X.; Brown, T.J.; Hanson, S.; Jimoh, O.F.; Ajabnoor, S.M.; Deane, K.H.; et al. Polyunsaturated Fatty Acids for the Primary and Secondary Prevention of Cardiovascular Disease. Cochrane Database Syst. Rev. 2018, 7, CD012345. [Google Scholar] [CrossRef]
- Dayrit, F.M. Lauric Acid Is a Medium-Chain Fatty Acid, Coconut Oil Is a Medium-Chain Triglyceride. Philipp. J. Sci. 2014, 143, 157–166. [Google Scholar]
- Deol, P.; Evans, J.R.; Dhahbi, J.; Chellappa, K.; Han, D.S.; Spindler, S.; Sladek, F.M. Soybean Oil Is More Obesogenic and Diabetogenic than Coconut Oil and Fructose in Mouse: Potential Role for the Liver. PLoS ONE 2015, 10, e0132672. [Google Scholar] [CrossRef] [Green Version]
- Tsartsou, E.; Proutsos, N.; Castanas, E.; Kampa, M. Network Meta-Analysis of Metabolic Effects of Olive-Oil in Humans Shows the Importance of Olive Oil Consumption With Moderate Polyphenol Levels as Part of the Mediterranean Diet. Front. Nutr. 2019, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Silva, É.; Spontoni, B.; Santo, E.; Paniago, G.; Portugal, L.; Dos, D.; Freitas, S.; Alcantara, G.; Filiú, W.; Freitas, K.; et al. Metabolic evaluation of the effects of a hyperlipid diet for obesity induction and standard normolipid diet (ain 93) consumption in wistar rats. Int. J. Dev. Res. 2017, 7, 11061. [Google Scholar]
- Schenkel, L.C.; Sivanesan, S.; Zhang, J.; Wuyts, B.; Taylor, A.; Verbrugghe, A.; Bakovic, M. Choline Supplementation Restores Substrate Balance and Alleviates Complications of Pcyt2 Deficiency. J. Nutr. Biochem. 2015, 26, 1221–1234. [Google Scholar] [CrossRef]
- Zeisel, S.H.; da Costa, K.-A. Choline: An Essential Nutrient for Public Health. Nutr. Rev. 2009, 67, 615–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinegar, R.; Truax, J.F.; Selph, J.L. Some Quantitative Temporal Characteristics of Carrageenin-Induced Pleurisy in the Rat. Proc. Soc. Exp. Biol. Med. 1973, 143, 711–714. [Google Scholar] [CrossRef] [PubMed]
- Vinegar, R.; Truax, J.F.; Selph, J.L.; Voelker, F.A. Pathway of Onset, Development, and Decay of Carrageenan Pleurisy in the Rat. Fed. Proc. 1982, 41, 2588–2595. [Google Scholar]
- Rankin, J.A. Biological Mediators of Acute Inflammation. AACN Clin. Issues 2004, 15, 3–17. [Google Scholar] [CrossRef]
- Wong, C.H.Y.; Heit, B.; Kubes, P. Molecular Regulators of Leucocyte Chemotaxis during Inflammation. Cardiovasc. Res. 2010, 86, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Smart, E.J. Myristic Acid Stimulates Endothelial Nitric-Oxide Synthase in a CD36- and an AMP Kinase-Dependent Manner. J. Biol. Chem. 2013, 288, 6583. [Google Scholar] [CrossRef] [Green Version]
- Alves, N.F.B.; de Queiroz, T.M.; de Almeida Travassos, R.; Magnani, M.; de Andrade Braga, V. Acute Treatment with Lauric Acid Reduces Blood Pressure and Oxidative Stress in Spontaneously Hypertensive Rats. Basic Clin. Pharmacol. Toxicol. 2017, 120, 348–353. [Google Scholar] [CrossRef] [Green Version]
- Angeles-Agdeppa, I.; Nacis, J.S.; Capanzana, M.V.; Dayrit, F.M.; Tanda, K.V. Virgin Coconut Oil Is Effective in Lowering C-Reactive Protein Levels among Suspect and Probable Cases of COVID-19. J. Funct. Foods 2021, 83, 104557. [Google Scholar] [CrossRef]
- Jädert, C.; Petersson, J.; Massena, S.; Ahl, D.; Grapensparr, L.; Holm, L.; Lundberg, J.O.; Phillipson, M. Decreased Leukocyte Recruitment by Inorganic Nitrate and Nitrite in Microvascular Inflammation and NSAID-Induced Intestinal Injury. Free Radic. Biol. Med. 2012, 52, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Vysakh, A.; Ratheesh, M.; Rajmohanan, T.P.; Pramod, C.; Premlal, S.; Girish kumar, B.; Sibi, P.I. Polyphenolics Isolated from Virgin Coconut Oil Inhibits Adjuvant Induced Arthritis in Rats through Antioxidant and Anti-Inflammatory Action. Int. Immunopharmacol. 2014, 20, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Jantz, M.A.; Antony, V.B. Pathophysiology of the Pleura. Respiration 2008, 75, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.-L.; Zhu, C.; Zhao, Y.-P.; Chen, X.-H.; Ji, C.-B.; Zhang, C.-M.; Zhu, J.-G.; Xia, Z.-K.; Tong, M.-L.; Guo, X.-R. Mitochondrial Dysfunction Is Induced by High Levels of Glucose and Free Fatty Acids in 3T3-L1 Adipocytes. Mol. Cell. Endocrinol. 2010, 320, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.D.S.; Block, J.M. Coconut Oil: What Do We Really Know about It so Far? Food Qual. Saf. 2019, 3, 61–72. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | SG | CG | LG | BG |
|---|---|---|---|---|
| Food ingestion | 5.29 ± 0.64 | 5.13 ± 0.82 | 5.27 ± 0.77 | 5.28 ± 0.69 |
| Body Weight | 49.8 ± 7.41 | 46.72 ± 2.69 | 47.1 ± 4.39 | 44.81 ± 5.04 |
| Liver | 2.16 ± 0.54 | 1.97 ± 0.15 | 1.75 ± 0.24 | 1.83 ± 0.42 |
| Epididymal | 2.54 ± 0.41 a | 2.09 ± 0.32 a | 2.34 ± 0.52 a | 1.40 ± 0.49 b,* |
| Mesenteric | 1.97 ± 0.58 a | 1.16 ± 0.21 b,† | 1.55 ± 0.28 a | 1.08 ± 0.54 b,* |
| Retroperitoneal | 1.11 ± 0.41 a | 0.75 ± 0.15 a | 0.94 ± 0.22 a | 0.61 ± 0.19 b,† |
| Omental | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.01 ± 0.00 |
| Perirenal | 0.64 ± 0.23 | 0.32 ± 0.06 | 0.34 ± 0.12 | 0.24 ± 0.17 |
| Adiposity index (AI) | 12.53 ± 2.80 a | 9.58 ± 1.02 b,* | 11.69 ± 1.67 a | 7.32 ± 3.13 b,* |
| Parameter | SG | CG | LG | BG |
|---|---|---|---|---|
| Blood glucose | 147.44 ± 65.01 | 137.39 ± 50.62 | 155.24 ± 82.13 | 98.70 ± 28.05 |
| Total cholesterol | 200.03 ± 35.42 | 208.30 ± 23.75 | 176.99 ± 19.26 | 194.96 ± 44.82 |
| HDL-Cholesterol | 95.00 ± 28.68 a | 47.37 ± 7.47 b,† | 76.84 ± 20.06 a | 43.35 ± 12.33 b,* |
| LDL-Cholesterol | 131.63 ± 29.03 a | 109.35 ± 23.75 a | 59.29 ± 7.56 b | 103.33 ± 31.72 a |
| VLDL-Cholesterol | 36.63 ± 4.41 a | 51.59 ± 14.46 b | 40.86 ± 7.80 a | 48.29 ± 16.79 a |
| Non-HDL Cholesterol | 131.63 ± 29.03 | 160.93 ± 22.05 | 100.15 ± 12.53 | 148.88 ± 41.80 |
| Triglycerides | 183.14 ± 22.06 a | 257.93 ± 72.30 b | 204.30 ± 38.99 a | 241.44 ± 83.93 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figueiredo, P.S.; Martins, T.N.; Ravaglia, L.M.; Alcantara, G.B.; Guimarães, R.d.C.A.; Freitas, K.d.C.; Nunes, Â.A.; de Oliveira, L.C.S.; Cortês, M.R.; Michels, F.S.; et al. Linseed, Baru, and Coconut Oils: NMR-Based Metabolomics, Leukocyte Infiltration Potential In Vivo, and Their Oil Characterization. Are There Still Controversies? Nutrients 2022, 14, 1161. https://doi.org/10.3390/nu14061161
Figueiredo PS, Martins TN, Ravaglia LM, Alcantara GB, Guimarães RdCA, Freitas KdC, Nunes ÂA, de Oliveira LCS, Cortês MR, Michels FS, et al. Linseed, Baru, and Coconut Oils: NMR-Based Metabolomics, Leukocyte Infiltration Potential In Vivo, and Their Oil Characterization. Are There Still Controversies? Nutrients. 2022; 14(6):1161. https://doi.org/10.3390/nu14061161
Chicago/Turabian StyleFigueiredo, Priscila Silva, Taynara Nogueira Martins, Luciana Marçal Ravaglia, Glaucia Braz Alcantara, Rita de Cássia Avellaneda Guimarães, Karine de Cássia Freitas, Ângela Alves Nunes, Lincoln Carlos Silva de Oliveira, Mário Rodrigues Cortês, Flavio Santana Michels, and et al. 2022. "Linseed, Baru, and Coconut Oils: NMR-Based Metabolomics, Leukocyte Infiltration Potential In Vivo, and Their Oil Characterization. Are There Still Controversies?" Nutrients 14, no. 6: 1161. https://doi.org/10.3390/nu14061161
APA StyleFigueiredo, P. S., Martins, T. N., Ravaglia, L. M., Alcantara, G. B., Guimarães, R. d. C. A., Freitas, K. d. C., Nunes, Â. A., de Oliveira, L. C. S., Cortês, M. R., Michels, F. S., Kadri, M. C. T., Bonfá, I. S., Filiú, W. F. d. O., Asato, M. A., de Faria, B. B., Nascimento, V. A. d., & Hiane, P. A. (2022). Linseed, Baru, and Coconut Oils: NMR-Based Metabolomics, Leukocyte Infiltration Potential In Vivo, and Their Oil Characterization. Are There Still Controversies? Nutrients, 14(6), 1161. https://doi.org/10.3390/nu14061161