
Ellagic Acid Alleviates Diquat-Induced Jejunum Oxidative Stress in C57BL/6 Mice through Activating Nrf2 Mediated Signaling Pathway


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
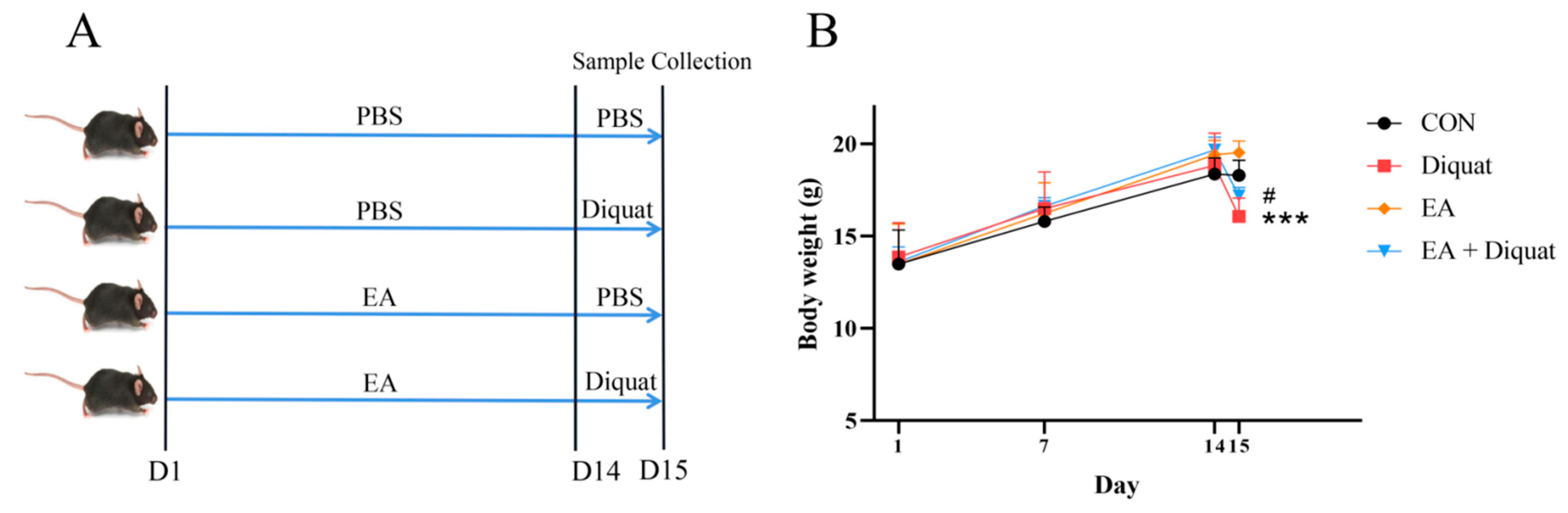
2.2. Animals and Treatment
2.3. Detection of Serum Indexes
2.4. Detection of ROS Level in Jejunum
2.5. Quantification of Intestinal Antioxidant Gene Expressions of Mice
2.6. Hematoxylin and Eosin Staining
2.7. Immunohistochemistry (IHC)
2.8. Statistics
3. Results
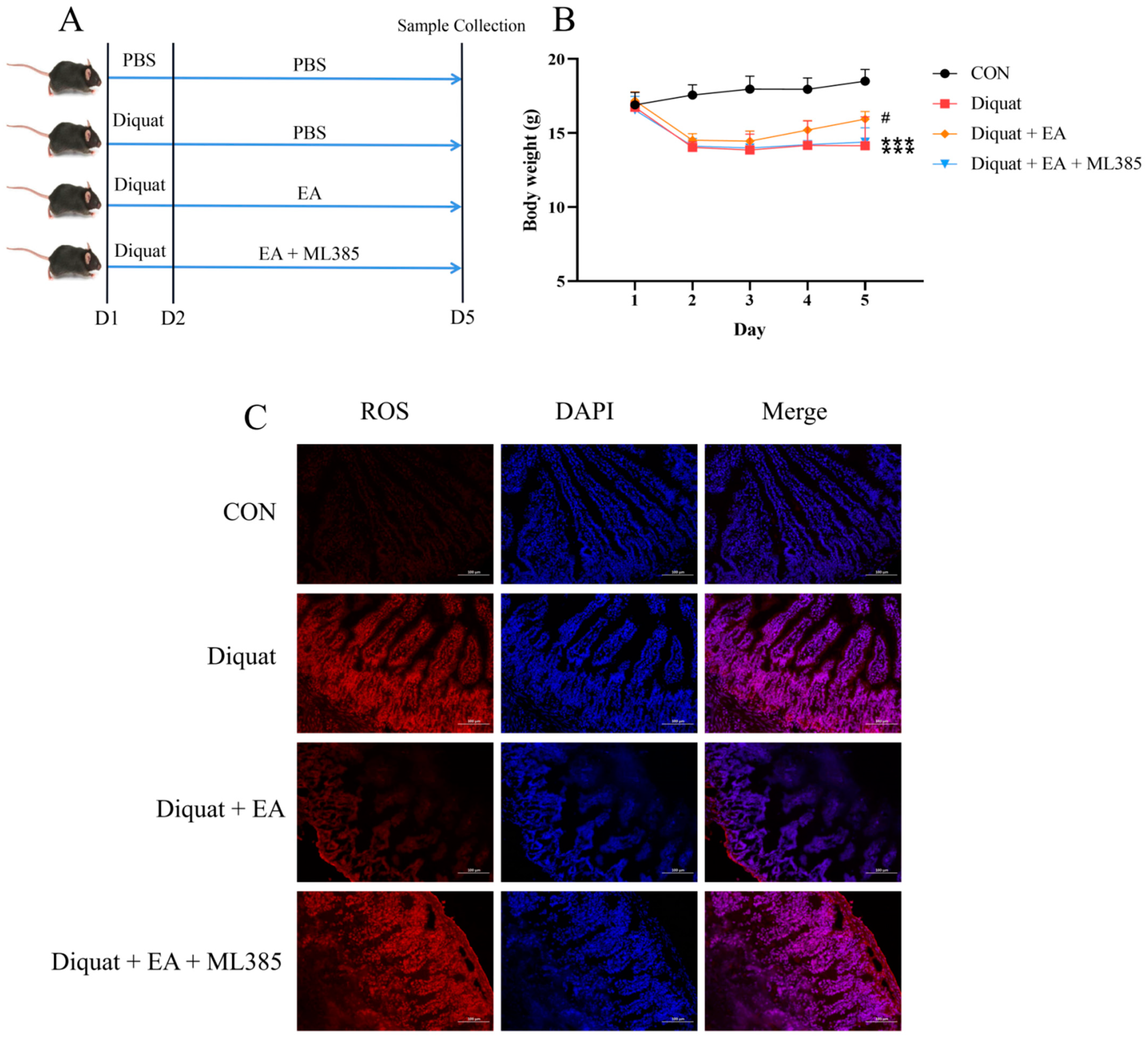
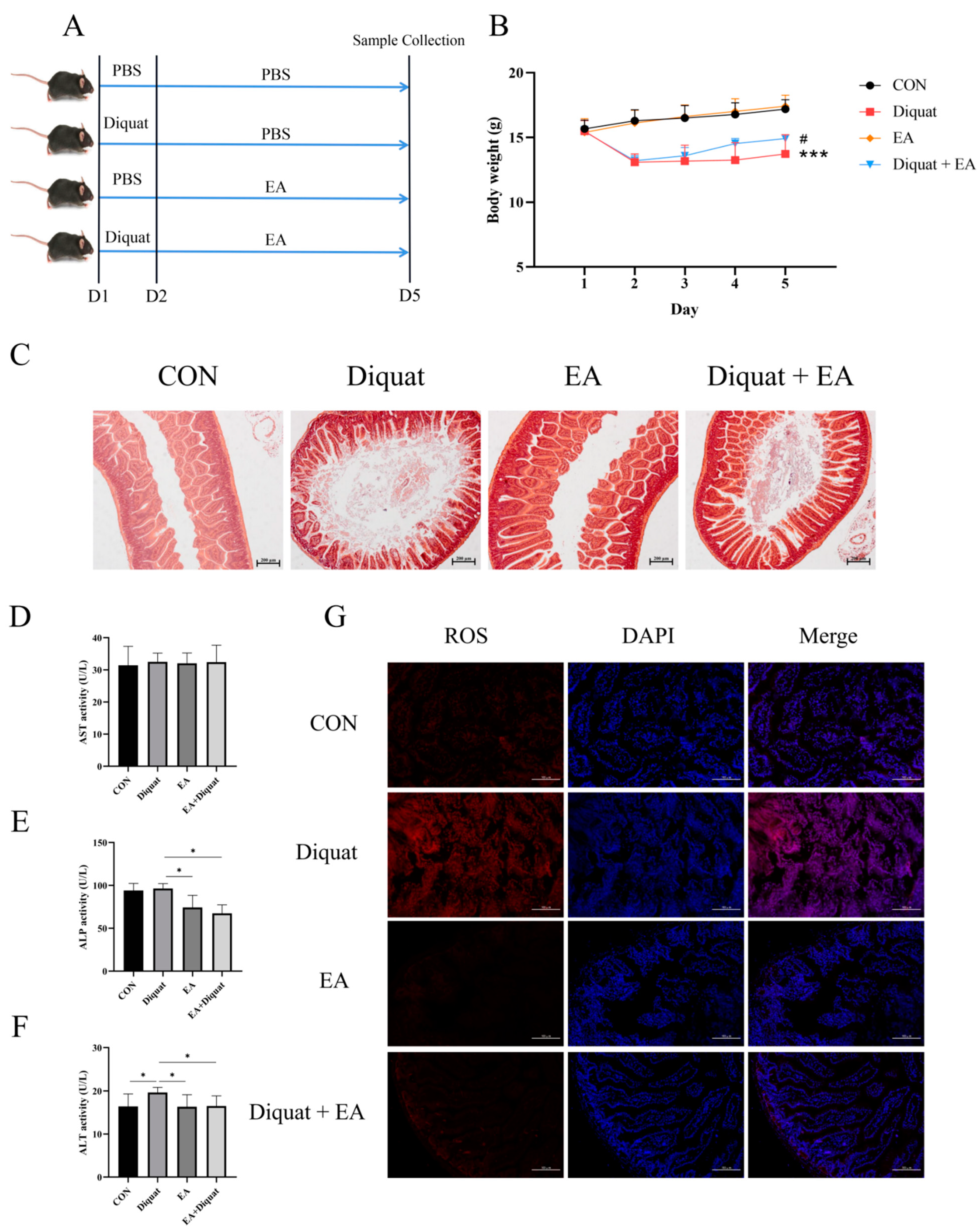
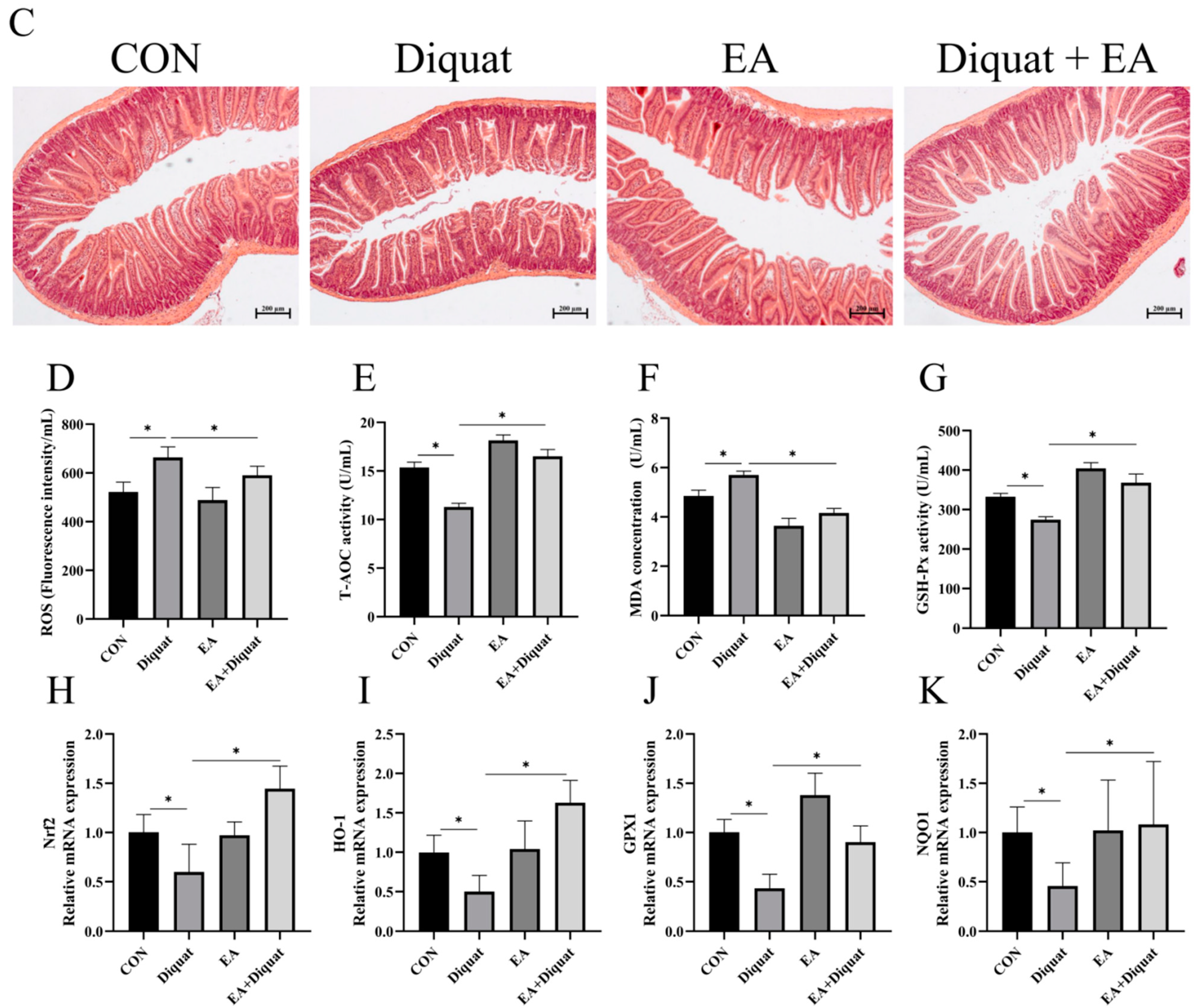
3.1. Therapeutic Ellagic Acid Treatment Reduced Diquat-Induced Weight Loss and Oxidative Damage
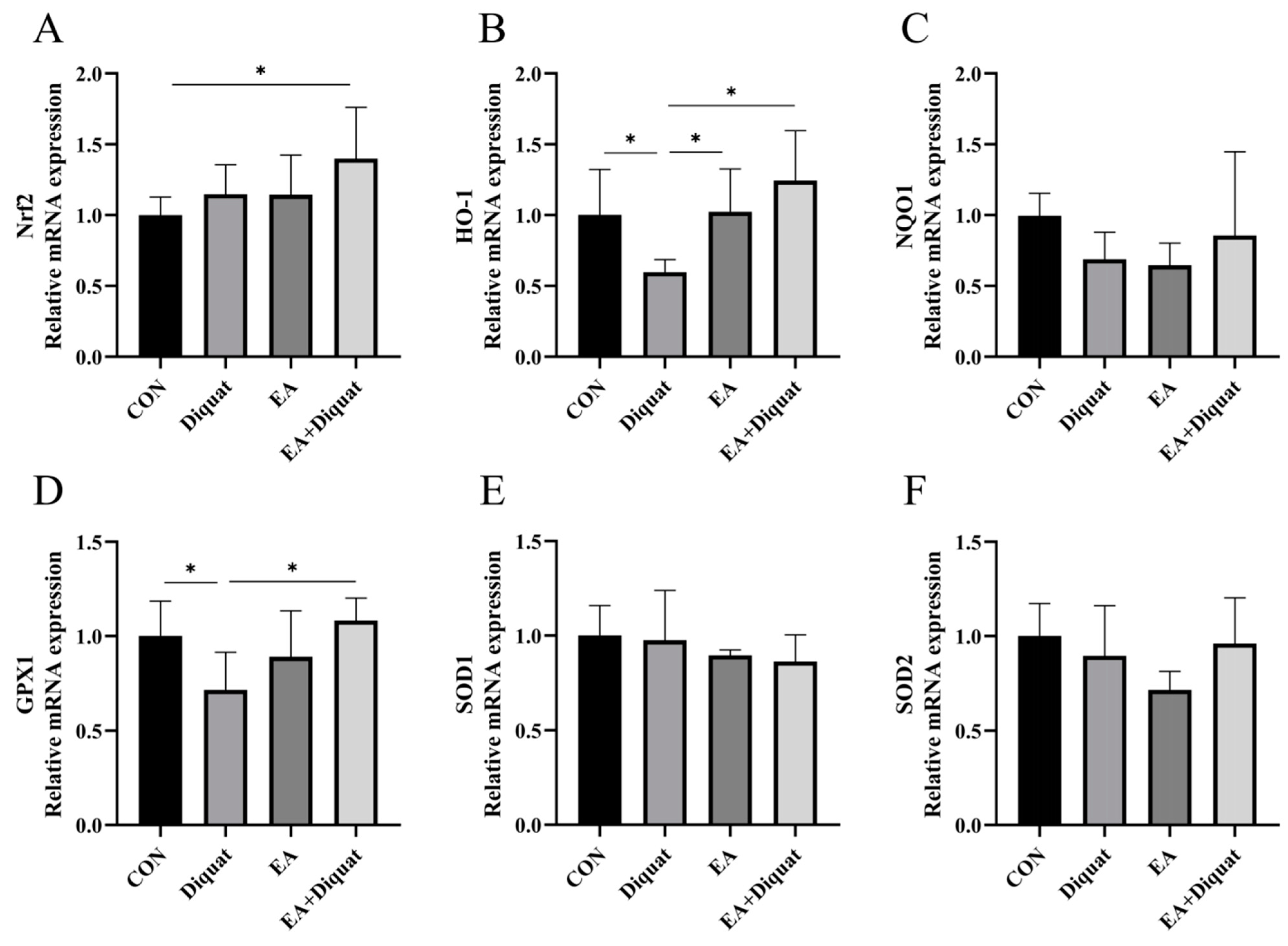
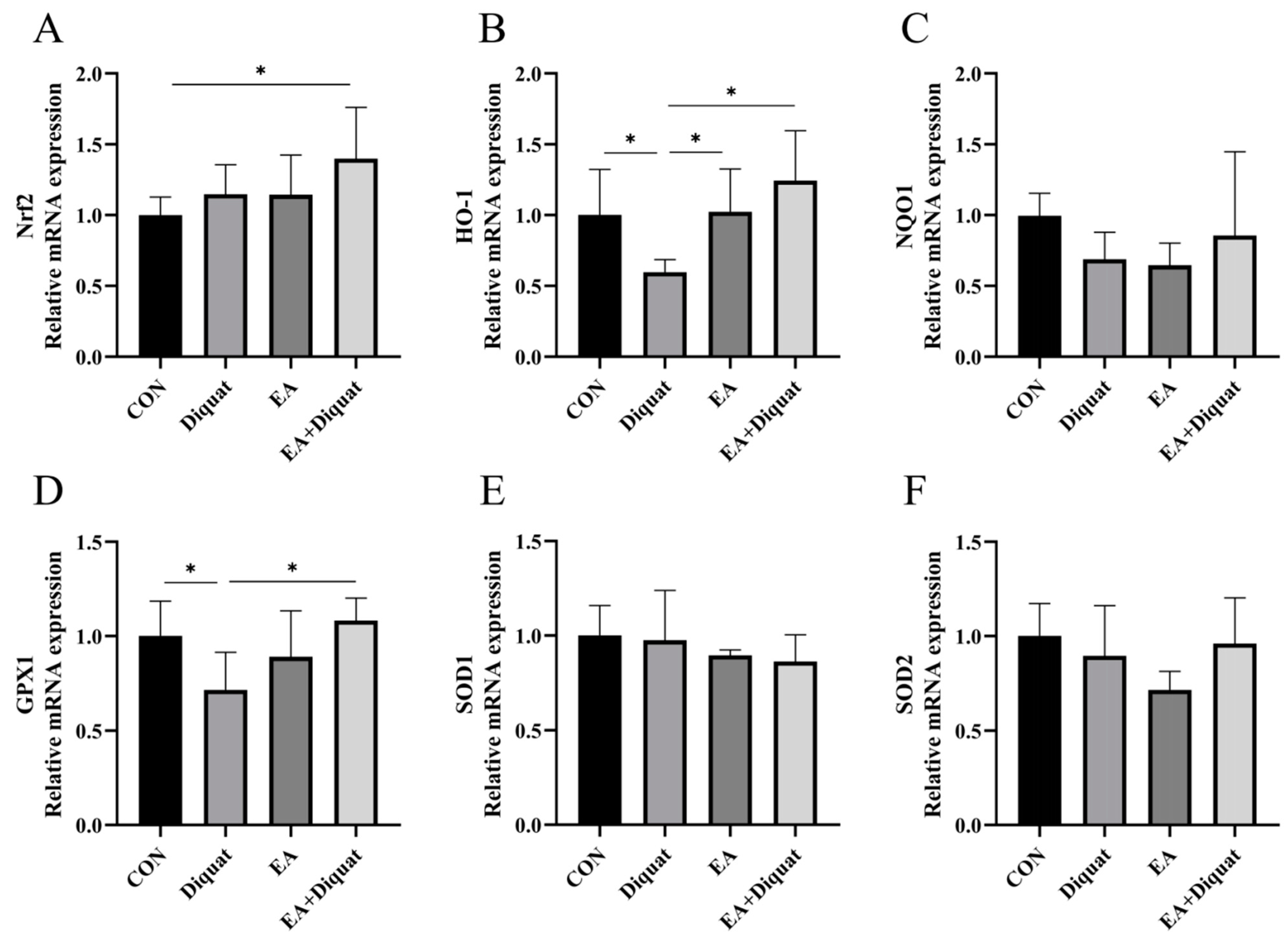
3.2. Ellagic Acid Upregulated Oxidative Stress Related Gene mRNA Expression in the Jejunum When Mice Challenged with Diquat
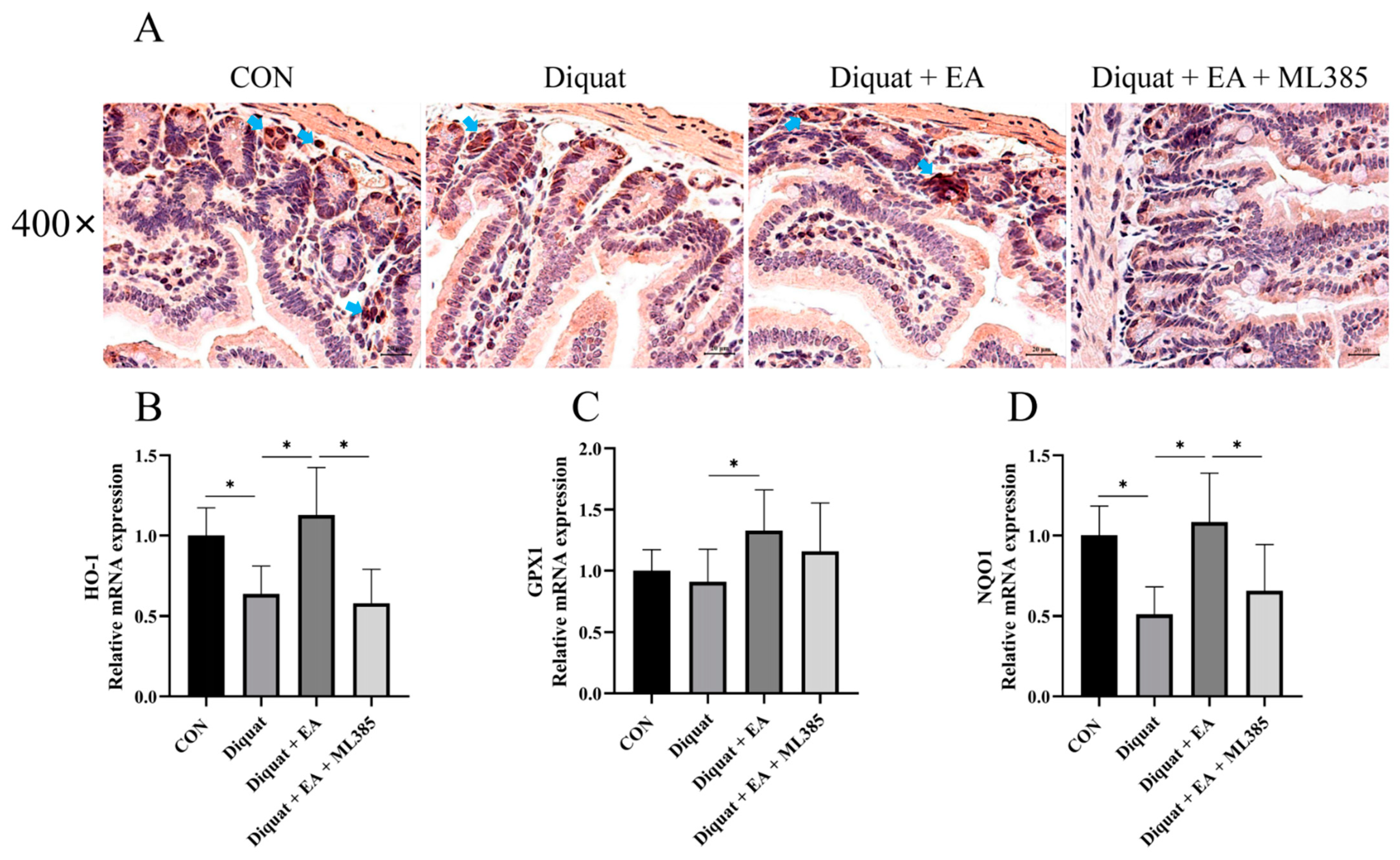
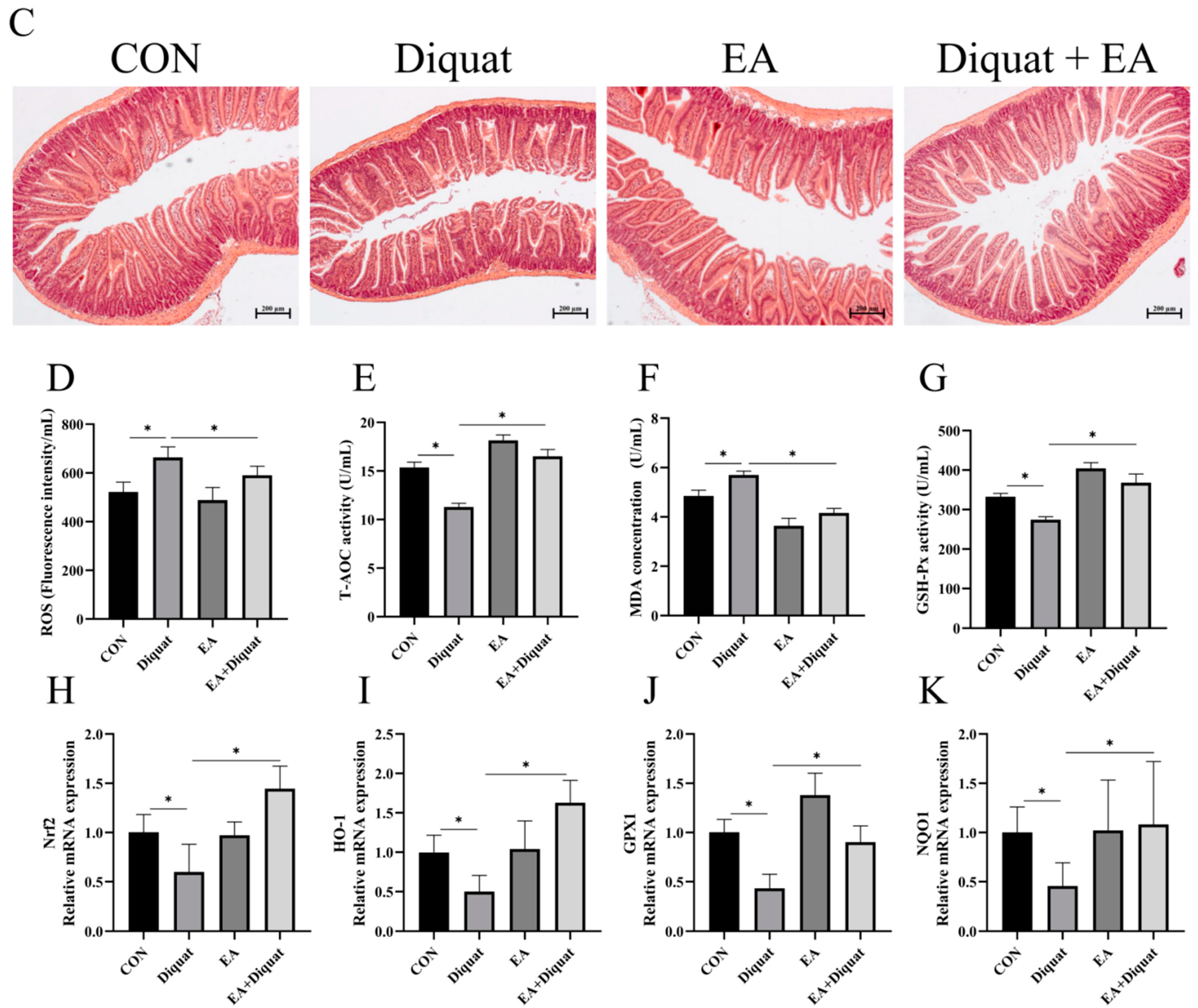
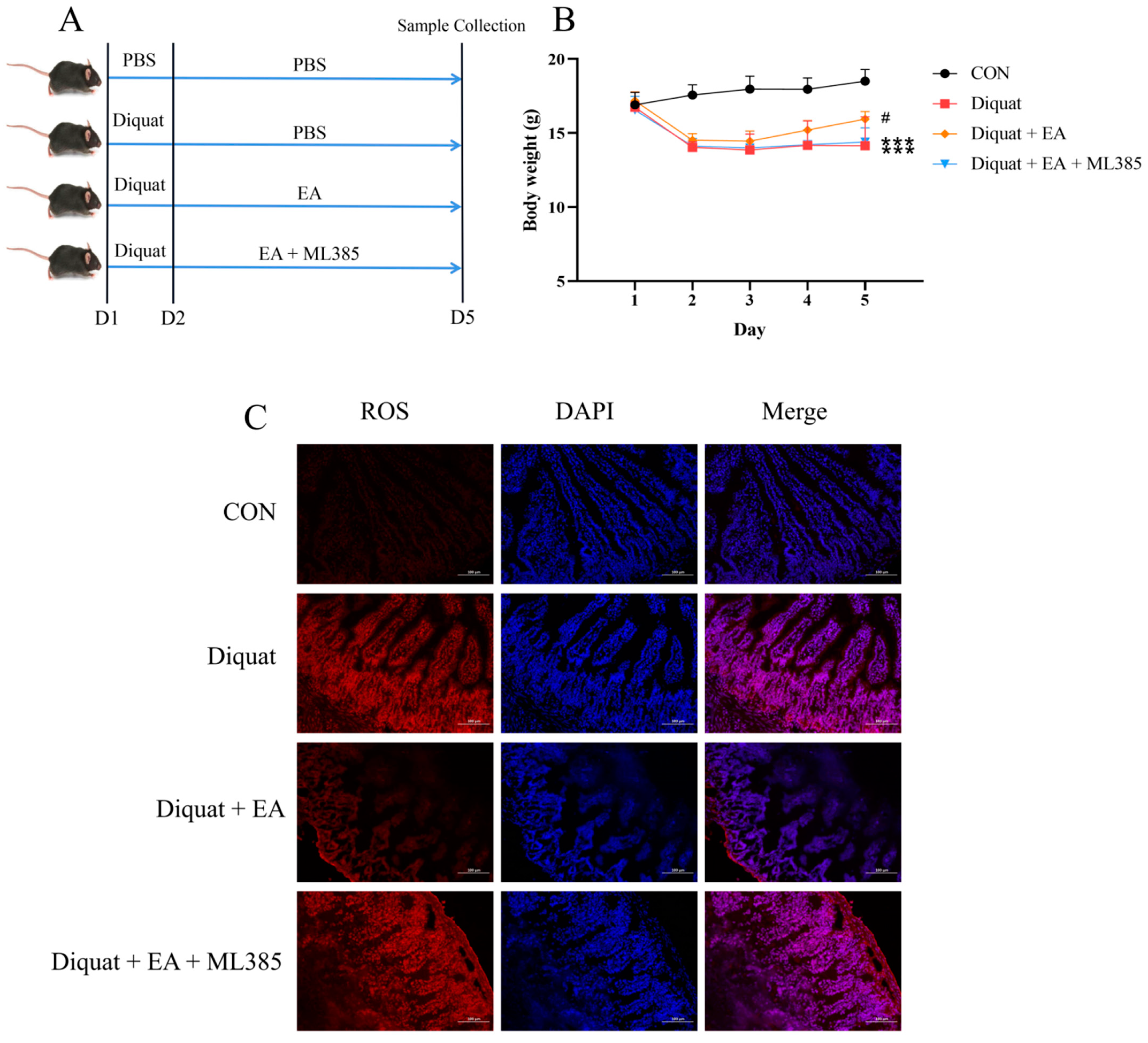
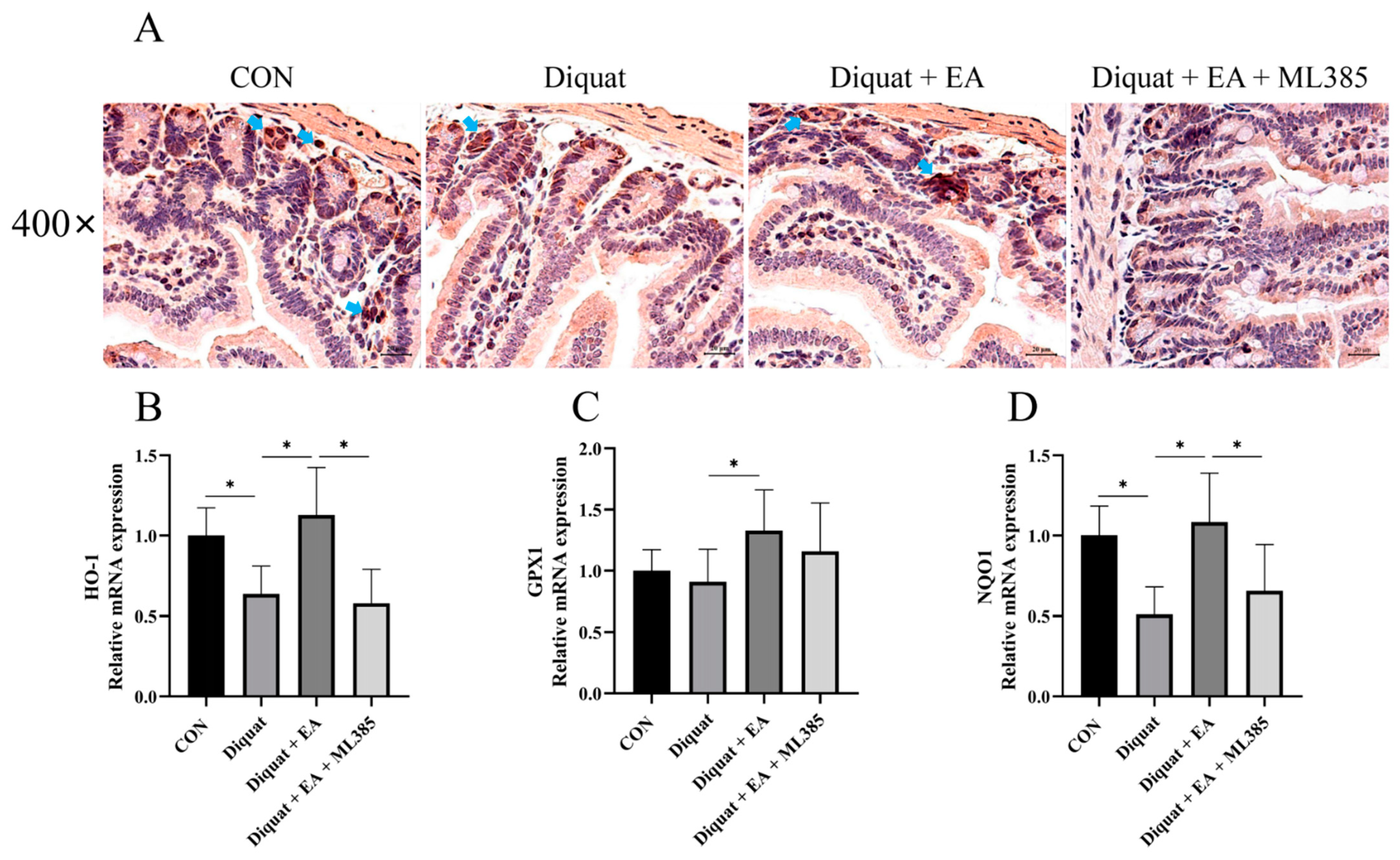
3.3. Specific Nrf2 Inhibitor (ML385) Inhibited the Therapeutic Effects of Ellagic Acid on Weight Loss and Jejunum Oxidative Stress Caused by Diquat
3.4. Ellagic Acid Alleviated Diquat-Induced Jejunal Oxidative Stress through Nrf2 Mediated Signaling Pathway
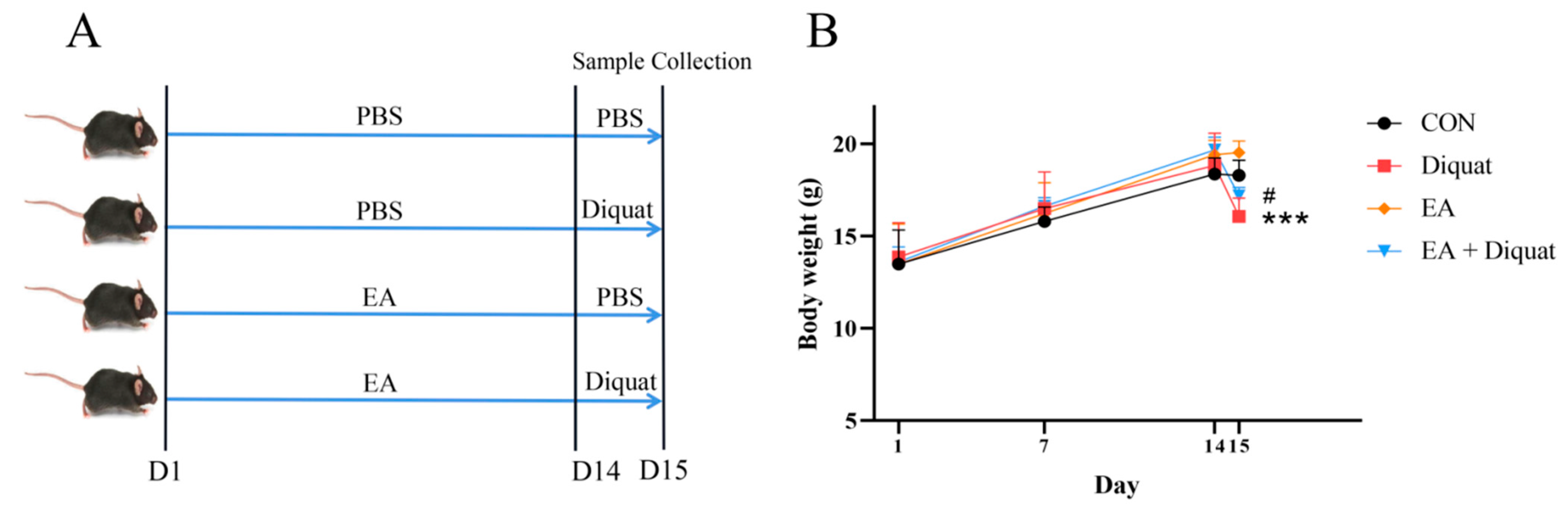
3.5. Prophylactic Ellagic Acid Treatment Ameliorated Diquat-Induced Jejunal Oxidative Damage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shen, Q.; Zhang, B.; Xu, R.; Wang, Y.; Ding, X.; Li, P. Antioxidant activity in vitro of the selenium-contained protein from the Se-enriched Bifidobacterium animalis. Anaerobe 2010, 16, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakash, V.; Marshall, J.R. Selenium and other anti-oxidants for chemoprevention of gastrointestinal cancers. Best Pract. Res. Clin. Gastroenterol. 2011, 25, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Fu, Y.; Wang, F.; Sinclair, A.; Li, D. Protective effects of a lipid extract from Hard-Shelled Mussel (Mytilus coruscus) on intestinal integrity after lipopolysaccharide challenge in mice. Nutrients 2018, 10, 860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ames, B.N. Dietary Carcinogens and Anticarcinogens. Science 1983, 221, 1256–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbari, P.; Braber, S.; Varasteh, S.; Alizadeh, A.; Garssen, J.; Fink-Gremmels, J. The intestinal barrier as an emerging target in the toxicological assessment of mycotoxins. Arch. Toxicol. 2017, 91, 1007–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, G.L.; Petschow, B.W.; Shaw, A.L.; Weaver, E. Gut barrier dysfunction and microbial translocation in cancer cachexia: A new therapeutic target. Curr. Opin. Support. Palliat. Care 2013, 7, 361–367. [Google Scholar] [CrossRef] [Green Version]
- Qiao, L.; Dou, X.; Yan, S.; Zhang, B.; Xu, C. Biogenic selenium nanoparticles synthesized by Lactobacillus casei ATCC 393 alleviate diquat-induced intestinal barrier dysfunction in C57BL/6 mice through their antioxidant activity. Food Funct. 2020, 11, 3020–3031. [Google Scholar] [CrossRef]
- Everson, C.A.; Henchen, C.J.; Szabo, A.; Hogg, N. Cell injury and repair resulting from sleep loss and sleep recovery in laboratory rats. Sleep 2014, 37, 1929–1940. [Google Scholar] [CrossRef] [Green Version]
- Kaspar, J.W.; Niture, S.K.; Jaiswal, A.K. Nrf2:INrf2 (KEAP1) signaling in oxidative stress. Free. Radic. Biol. Med. 2009, 47, 1304–1309. [Google Scholar] [CrossRef] [Green Version]
- Milković, L.; Tomljanović, M.; Čipak Gašparović, A.; Novak Kujundžić, R.; Šimunić, D.; Konjevoda, P.; Mojzeš, A.; Đaković, N.; Žarković, N.; Gall Trošelj, K. Nutritional stress in head and neck cancer originating cell lines: The sensitivity of the NRF2-NQO1 axis. Cells 2019, 8, 1001. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Shen, T.; Yang, H.; Gu, W. Ruthenium complexes induce HepG2 human hepatocellular carcinoma cell apoptosis and inhibit cell migration and invasion through regulation of the Nrf2 pathway. Int. J. Mol. Sci. 2016, 17, 775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceci, C.; Lacal, P.M.; Tentori, L.; De Martino, M.G.; Miano, R.; Graziani, G. Experimental evidence of the antitumor, antimetastatic and antiangiogenic activity of ellagic acid. Nutrients 2018, 10, 1756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konstantinidi, M.; Koutelidakis, A.E. Functional foods and bioactive compounds: A review of its possible role on weight management and obesity’s metabolic consequences. Medicines 2019, 6, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, X.; Jian, T.; Wu, Y.; Zuo, Y.; Li, J.; Lv, H.; Ma, L.; Ren, B.; Zhao, L.; Li, W.; et al. Ellagic acid amelio-rates oxidative stress and insulin resistance in high glucose-treated HepG2 cells via miR-223/KEAP1-Nrf2 pathway. Biomed. Pharmacother. 2019, 110, 85–94. [Google Scholar] [CrossRef]
- Singh, R.; Chandrashekharappa, S.; Bodduluri, S.R.; Baby, B.V.; Hegde, B.; Kotla, N.G.; Hiwale, A.A.; Saiyed, T.; Patel, P.; Vijay-Kumar, M.; et al. Enhancement of the gut barrier integrity by a microbial metabolite through the Nrf2 pathway. Nat. Commun. 2019, 10, 89. [Google Scholar] [CrossRef] [Green Version]
- Zheng, D.; Liu, Z.; Zhou, Y.; Hou, N.; Yan, W.; Qin, Y.; Ye, Q.; Cheng, X.; Xiao, Q.; Bao, Y.; et al. Urolithin B, a gut microbiota metabolite, protects against myocardial ischemia/reperfusion injury via p62/KEAP1/Nrf2 signaling pathway. Pharmacol. Res. 2020, 153, 104655. [Google Scholar] [CrossRef]
- Farooqui, Z.; Mohammad, R.S.; Lokhandwala, M.F.; Banday, A.A. Nrf2 inhibition induces oxidative stress, renal inflammation and hypertension in mice. Clin. Exp. Hypertens. 2021, 43, 175–180. [Google Scholar] [CrossRef]
- Wang, R.; Luo, Y.; Lu, Y.; Wang, D.; Wang, T.; Pu, W.; Wang, Y. Maggot extracts alleviate inflammation and oxidative stress in acute experimental colitis via the activation of Nrf2. Oxid. Med. Cell. Longev. 2019, 2019, 4703253. [Google Scholar] [CrossRef] [Green Version]
- Qian, X.; Wang, J.; Cai, M.; Sun, H.; Xu, H.; Wen, H.; Zhu, H. Estradiol valerate enhances cardiac function via the Nrf2 signaling pathway to protect against oxidative stress by the Nrf2 signaling pathway in an Ovariectomized Rat Model. Curr. Pharm. Des. 2021, 27, 4716–4725. [Google Scholar] [CrossRef]
- Luo, L.; Chen, Y.; Wu, D.; Shou, J.; Wang, S.; Ye, J.; Tang, X.; Wang, X. Butylated hydroxyanisole induces distinct expression patterns of Nrf2 and detoxification enzymes in the liver and small intestine of C57BL/6 mice. Toxicol. Appl. Pharmacol. 2015, 288, 339–348. [Google Scholar] [CrossRef]
- Zhang, J.; Gao, B.; Wang, J.; Ren, Q.; Chen, J.; Ma, Q.; Zhang, Z.; Xing, B. Critical role of FoxO1 in granulosa cell apoptosis caused by oxidative stress and protective effects of grape seed procyanidin B2. Oxid. Med. Cell. Longev. 2016, 2016, 6147345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Gao, B.; Wang, J.; Wang, X.; Ren, Q.; Chen, J.; Ma, Q.; Xing, B. Chronic exposure to diquat causes reproductive toxicity in female mice. PLoS ONE 2016, 11, e0147075. [Google Scholar]
- Bokov, A.; Lindsey, M.L.; Khodr, C.; Sabia, M.R.; Richardson, A. Long-lived ames dwarf mice are resistant to chemical stressors. J. Gerontol. A Biol. Sci. Med. Sci. 2009, 64, 819–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Chen, Y.; Chen, Y.; Jia, P.; Ji, S.; Xu, J.; Li, Y.; Wang, T. Comparison of the effects of resveratrol and its derivative pterostilbene on hepatic oxidative stress and mitochondrial dysfunction in piglets chal-lenged with diquat. Food Funct. 2020, 11, 4202–4215. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Huang, H.; Liu, S.; Zhuang, Y.; Yang, H.; Li, Y.; Chen, S.; Wang, L.; Yin, L.; Yao, Y.; et al. Tannic acid modulates intestinal barrier functions associated with intestinal morphology, antioxidative activity, and intestinal tight junction in a diquat-induced mouse model. RSC Adv. 2019, 9, 31988–31998. [Google Scholar] [CrossRef] [Green Version]
- Jiao, N.; Wu, Z.; Ji, Y.; Wang, B.; Dai, Z.; Wu, G. L-Glutamate enhances barrier and antioxidative functions in intestinal porcine epithelial cells. J. Nutr. 2015, 145, 2258–2264. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Zhu, G.; Zheng, C.; Li, J.; Sheng, S.; Li, D.; Wang, G.; Zhang, F. Ellagic acid protects dopamine neurons from rotenone-induced neurotoxicity via activation of Nrf2 signalling. J. Cell. Mol. Med. 2020, 24, 9446–9456. [Google Scholar] [CrossRef]
- Eizirik, D.L.; Flodström, M.; Karlsen, A.E.; Welsh, N. The harmony of the spheres: Inducible nitric oxide synthase and related genes in pancreatic beta cells. Diabetologia 1996, 39, 875–890. [Google Scholar] [CrossRef]
- Kongpetch, S.; Kukongviriyapan, V.; Prawan, A.; Senggunprai, L.; Kukongviriyapan, U.; Buranrat, B. Crucial role of heme oxygenase-1 on the sensitivity of cholangiocarcinoma cells to chemotherapeutic agents. PLoS ONE 2012, 7, e34994. [Google Scholar] [CrossRef] [Green Version]
- Rubiolo, J.A.; Mithieux, G.; Vega, F.V. Resveratrol protects primary rat hepatocytes against oxidative stress damage: Activation of the Nrf2 transcription factor and augmented activities of antioxidant enzymes. Eur. J. Pharmacol. 2008, 591, 66–72. [Google Scholar] [CrossRef]
- Zhu, H.; Jia, Z.; Misra, B.R.; Zhang, L.; Cao, Z.; Yamamoto, M.; Trush, M.A.; Misra, H.P.; Li, Y. Nuclear factor E2-related factor 2-dependent myocardiac cytoprotection against oxidative and electrophilic stress. Cardiovasc. Toxicol. 2008, 8, 71–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinbrenner, H.; Sies, H. Protection against reactive oxygen species by selenoproteins. Biochim. Biophys. Acta. 2009, 1790, 1478–1485. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Li, J.; Zhang, L.; Sun, Y.; Jiang, J.; Huang, Y.; Xu, H.; Jiang, H.; Hu, R. Nrf2 protects against acute lung injury and inflammation by modulating TLR4 and Akt signaling. Free Radic. Biol. Med. 2018, 121, 78–85. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′-3′) | Primer Length (bp) | Product Length (bp) |
|---|---|---|---|
| β-actin F | GGCTGTATTCCCCTCCATCG | 20 | 154 |
| β-actin R | CCAGTTGGTAACAATGCCATGT | 22 | |
| HO-1 F | AAGCCGAGAATGCTGAGTTCA | 21 | 100 |
| HO-1 R | GCCGTGTAGATATGGTACAAGGA | 23 | |
| NQO1 F | AGGATGGGAGGTACTCGAATC | 21 | 144 |
| NQO1 R | AGGCGTCCTTCCTTATATGCTA | 22 | |
| SOD1 F | AACCAGTTGTGTTGTCAGGAC | 21 | 139 |
| SOD1 R | CCACCATGTTTCTTAGAGTGAGG | 23 | |
| SOD2 F | CAGACCTGCCTTACGACTATGG | 22 | 113 |
| SOD2 R | CTCGGTGGCGTTGAGATTGTT | 21 | |
| GPX1 F | CCACCGTGTATGCCTTCTCC | 20 | 105 |
| GPX1 R | AGAGAGACGCGACATTCTCAAT | 22 | |
| Nrf2 F | CTGAACTCCTGGACGGGACTA | 21 | 182 |
| Nrf2 R | CGGTGGGTCTCCGTAAATGG | 20 | |
| CAT F | GGAGGCGGGAACCCAATAG | 19 | 102 |
| CAT R | GTGTGCCATCTCGTCAGTGAA | 21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Wang, S.; Wu, Y.; Liu, X.; Wang, J.; Han, D. Ellagic Acid Alleviates Diquat-Induced Jejunum Oxidative Stress in C57BL/6 Mice through Activating Nrf2 Mediated Signaling Pathway. Nutrients 2022, 14, 1103. https://doi.org/10.3390/nu14051103
Zhang X, Wang S, Wu Y, Liu X, Wang J, Han D. Ellagic Acid Alleviates Diquat-Induced Jejunum Oxidative Stress in C57BL/6 Mice through Activating Nrf2 Mediated Signaling Pathway. Nutrients. 2022; 14(5):1103. https://doi.org/10.3390/nu14051103
Chicago/Turabian StyleZhang, Xiangyu, Shilan Wang, Yujun Wu, Xiaoyi Liu, Junjun Wang, and Dandan Han. 2022. "Ellagic Acid Alleviates Diquat-Induced Jejunum Oxidative Stress in C57BL/6 Mice through Activating Nrf2 Mediated Signaling Pathway" Nutrients 14, no. 5: 1103. https://doi.org/10.3390/nu14051103
APA StyleZhang, X., Wang, S., Wu, Y., Liu, X., Wang, J., & Han, D. (2022). Ellagic Acid Alleviates Diquat-Induced Jejunum Oxidative Stress in C57BL/6 Mice through Activating Nrf2 Mediated Signaling Pathway. Nutrients, 14(5), 1103. https://doi.org/10.3390/nu14051103