
FADS1 and FADS2 Gene Polymorphisms Modulate the Relationship of Omega-3 and Omega-6 Fatty Acid Plasma Concentrations in Gestational Weight Gain: A NISAMI Cohort Study

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
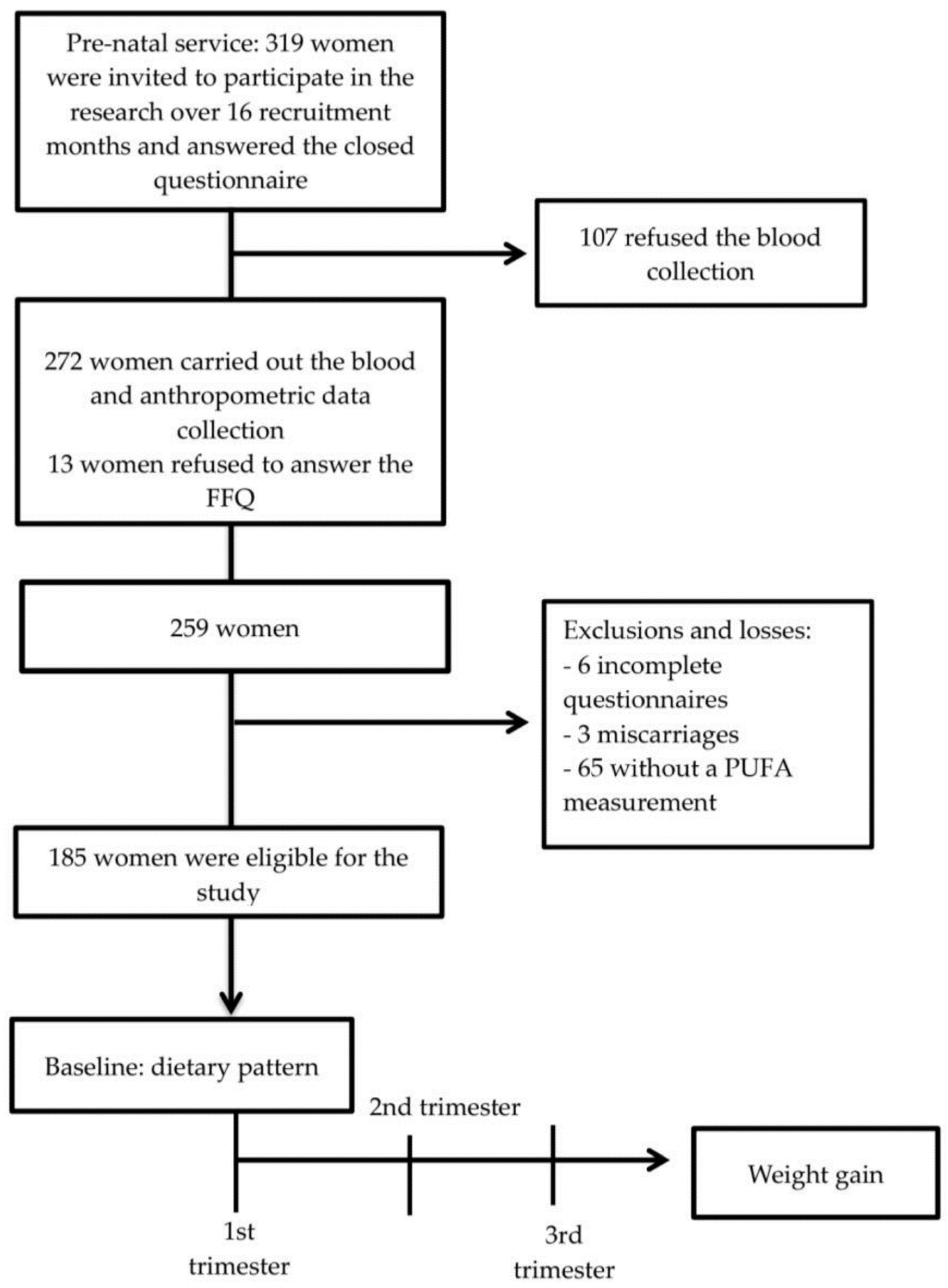
2.1. Study Design and Population
2.2. Sample Size
2.3. Exclusion and Inclusion Criteria
2.4. Data Collection
2.5. Plasma Fatty Acid Composition
2.6. Genetic Analyses
2.7. Anthropometric Measurements
2.8. Statistical Analysis
3. Results
3.1. Description of Participants
3.2. Main Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Mannan, M.; Doi, S.A.R.; Mamun, A.A. Association between weight gain during pregnancy and postpartum weight retention and obesity: A bias-adjusted meta-analysis. Nutr. Rev. 2013, 71, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Martinez, M.P.; Chow, T.; Xiang, A.H. BMI growth trajectory from ages 2 to 6 years and its association with maternal obesity, diabetes during pregnancy, gestational weight gain, and breastfeeding. Pediatr. Obes. 2019, 15, e12579. [Google Scholar] [CrossRef] [PubMed]
- Marchi, J.; Berg, M.; Dencker, A.; Olander, E.K.; Begley, C. Risks associated with obesity in pregnancy, for the mother and baby: A systematic review of reviews. Obes. Rev. 2015, 16, 621–638. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, R.F.; Abell, S.K.; Ranasinha, S.; Misso, M.; Boyle, J.A.; Black, M.H.; Li, N.; Hu, G.; Corrado, F.; Rode, L.; et al. Association of Gestational Weight Gain With Maternal and Infant Outcomes: A Systematic Review and Meta-analysis. JAMA 2017, 317, 2207–2225. [Google Scholar] [CrossRef]
- Groth, S.W.; LaLonde, A.; Wu, T.; Fernandez, I.D. Obesity candidate genes, gestational weight gain, and body weight changes in pregnant women. Nutrition 2018, 48, 61–66. [Google Scholar] [CrossRef]
- Poston, L.; Caleyachetty, R.; Cnattingius, S.; Corvalan, C.; Uauy, R.; Herring, S.; Gillman, M.W. Preconceptional and maternal obesity: Epidemiology and health consequences. Lancet Diabetes Endocrinol. 2016, 4, 1025–1036. [Google Scholar] [CrossRef]
- Keijer, J.; Hoevenaars, F.P.M.; Nieuwenhuizen, A.; van Schothorst, E.M. Nutrigenomics of body weight regulation: A rationale for careful dissection of individual contributors. Nutrients 2014, 6, 4531–4551. [Google Scholar] [CrossRef] [Green Version]
- Ordovas, J.M.; Mooser, V. Nutrigenomics and nutrigenetics. Curr. Opin. Lipidol. 2004, 15, 101–108. [Google Scholar] [CrossRef]
- Lattka, E.; Illig, T.; Koletzko, B.; Heinrich, J. Genetic variants of the FADS1 FADS2 gene cluster as related to essential fatty acid metabolism. Curr. Opin. Lipidol. 2010, 21, 64–69. [Google Scholar] [CrossRef]
- Gillingham, L.G.; Harding, S.V.; Rideout, T.C.; Yurkova, N.; Cunnane, S.C.; Eck, P.K.; Jones, P.J.H. Dietary oils and FADS1-FADS2 genetic variants modulate [13C]alpha-linolenic acid metabolism and plasma fatty acid composition. Am. J. Clin. Nutr. 2013, 97, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Steer, C.D.; Lattka, E.; Koletzko, B.; Golding, J.; Hibbeln, J.R. Maternal fatty acids in pregnancy, FADS polymorphisms, and child intelligence quotient at 8 y of age. Am. J. Clin. Nutr. 2013, 98, 1575–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Garza Puentes, A.; Montes Goyanes, R.; Chisaguano Tonato, A.M.; Torres-Espínola, F.J.; Arias García, M.; de Almeida, L.; Bonilla Aguirre, M.; Guerendiain, M.; Castellote Bargalló, A.I.; Segura Moreno, M.; et al. Association of maternal weight with FADS and ELOVL genetic variants and fatty acid levels—The PREOBE follow-up. PLoS ONE 2017, 12, e0179135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koletzko, B.; Lattka, E.; Zeilinger, S.; Illig, T.; Steer, C. Genetic variants of the fatty acid desaturase gene cluster predict amounts of red blood cell docosahexaenoic and other polyunsaturated fatty acids in pregnant women: Findings from the Avon Longitudinal Study of Parents and Children. Am. J. Clin. Nutr. 2011, 93, 211–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, L.; Innis, S.M. Genetic variants of the FADS1 FADS2 gene cluster are associated with altered (n-6) and (n-3) essential fatty acids in plasma and erythrocyte phospholipids in women during pregnancy and in breast milk during lactation. J. Nutr. 2008, 138, 2222–2228. [Google Scholar] [CrossRef] [Green Version]
- Mathias, R.A.; Vergara, C.; Gao, L.; Rafaels, N.; Hand, T.; Campbell, M.; Bickel, C.; Ivester, P.; Sergeant, S.; Barnes, K.C.; et al. FADS genetic variants and omega-6 polyunsaturated fatty acid metabolism in a homogeneous island population. J. Lipid Res. 2010, 51, 2766–2774. [Google Scholar] [CrossRef] [Green Version]
- Molto-Puigmarti, C.; Plat, J.; Mensink, R.P.; Muller, A.; Jansen, E.; Zeegers, M.P.; Thijs, C. FADS1 FADS2 gene variants modify the association between fish intake and the docosahexaenoic acid proportions in human milk. Am. J. Clin. Nutr. 2010, 91, 1368–1376. [Google Scholar] [CrossRef] [Green Version]
- Santana, J.M.; Alves de Oliveira Queiroz, V.; Monteiro Brito, S.; Barbosa Dos Santos, D.; Marlucia Oliveira Assis, A. Food Consumption Patterns During Pregnancy: A Longitudinal Study in a Region of the North East of Brazil. Nutr. Hosp. 2015, 32, 130–138. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Hartman, L.; Lago, R.C. Rapid preparation of fatty acid methyl esters from lipids. Lab. Pract. 1973, 22, 475–476. [Google Scholar]
- Carvalho, G.Q.; Pereira-Santos, M.; Marcon, L.D.; Louro, I.D.; Peluzio, M.C.G.; Santos, D.B. Maternal polymorphisms in the FADS1 and FADS2 genes modify the association between PUFA ingestion and plasma concentrations of omega-3 polyunsaturated fatty acids. Prostaglandins Leukot. Essent. Fat. Acids 2019, 150, 38–46. [Google Scholar] [CrossRef]
- Lohman, T.G.; Roche, A.F.; Martorell, R. Anthropometric standardization reference manual. In Human Kinetics Books; Human Kinetics: Champaign, IL, USA, 1988; Volume 177. [Google Scholar] [CrossRef]
- Atalah, S.E.; Castillo, L.C.; Castro, S.R.; Aldea, P.A. Propuesta de un nuevo estándar de evaluación nutricional en embarazadas. Rev. Med. Chil. 1997, 125, 1429–1436. [Google Scholar] [PubMed]
- Institute of Medicine. Weight Gain during Pregnancy: Reexamining the Guidelines; Rasmussen, K.M., Yaktine, A.L., Eds.; National A: Washington, DC, USA, 2009; ISBN 9780309131131. [Google Scholar]
- Amorim, L.D.A.F.; Fiaccone, R.L.; Santos, C.A.S.T.; dos Santos, T.N.; de Moraes, L.T.L.P.; Oliveira, N.F.; Barbosa, S.O.; dos Santos, D.N.; dos Santos, L.M.; Matos, S.M.A.; et al. Structural equation modeling in epidemiology. Cad. Saude Publica 2010, 26, 2251–2262. [Google Scholar] [CrossRef] [PubMed]
- Kline, R.B. Principles and Practice of Structural Equation Modeling; Guilford Publications: New York, NY, USA, 2011; Volume 77, ISBN 1606238760. [Google Scholar]
- Tai, C.C.; Ding, S.T. N-3 polyunsaturated fatty acids regulate lipid metabolism through several inflammation mediators: Mechanisms and implications for obesity prevention. J. Nutr. Biochem. 2010, 21, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Feskens, E.J.; Dolle, M.E.; Imholz, S.; Verschuren, W.M.; Muller, M.; Boer, J.M. Dietary n-3 and n-6 polyunsaturated fatty acid intake interacts with FADS1 genetic variation to affect total and HDL-cholesterol concentrations in the Doetinchem Cohort Study. Am. J. Clin. Nutr. 2010, 92, 258–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinelli, N.; Girelli, D.; Malerba, G.; Guarini, P.; Illig, T.; Trabetti, E.; Sandri, M.; Friso, S.; Pizzolo, F.; Schaeffer, L.; et al. FADS genotypes and desaturase activity estimated by the ratio of arachidonic acid to linoleic acid are associated with inflammation and coronary artery disease. Am. J. Clin. Nutr. 2008, 88, 941–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vimaleswaran, K.S.; Cavadino, A.; Berry, D.J.; Power, C.; Hyppönen, E.; Jorde, R.; Grimnes, G.; Dieffenbach, A.K.; Schöttker, B.; Saum, K.-U.; et al. Association of vitamin D status with arterial blood pressure and hypertension risk: A mendelian randomisation study. Lancet Diabetes Endocrinol. 2014, 2, 719–729. [Google Scholar] [CrossRef] [Green Version]
- Scholtz, S.A.; Kerling, E.H.; Shaddy, D.J.; Li, S.; Thodosoff, J.M.; Colombo, J.; Carlson, S.E. Docosahexaenoic acid (DHA) supplementation in pregnancy differentially modulates arachidonic acid and DHA status across FADS genotypes in pregnancy. Prostaglandins Leukot. Essent. Fat. Acids 2015, 94, 29–33. [Google Scholar] [CrossRef] [Green Version]
- Muller, M.; Kersten, S. Nutrigenomics: Goals and strategies. Nat. Rev. Genet. 2003, 4, 315–322. [Google Scholar] [CrossRef]
- Ralston, J.C.; Matravadia, S.; Gaudio, N.; Holloway, G.P.; Mutch, D.M. Polyunsaturated fatty acid regulation of adipocyte FADS1 and FADS2 expression and function. Obesity 2015, 23, 725–728. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Athinarayanan, S.; Jiang, G.; Chalasani, N.; Zhang, M.; Liu, W. Fatty acid desaturase 1 gene polymorphisms control human hepatic lipid composition. Hepatology 2015, 61, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Ding, Z.; Liu, G.-L.; Li, X.; Chen, X.-Y.; Wu, Y.-X.; Cui, C.-C.; Zhang, X.; Yang, G.; Xie, L. Association of polyunsaturated fatty acids in breast milk with fatty acid desaturase gene polymorphisms among Chinese lactating mothers. Prostaglandins Leukot. Essent. Fat. Acids 2016, 109, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, J.P.; Kobe, T.; Witte, V.; Willers, J.; Gingrich, A.; Tesky, V.; Pantel, J.; Rujescu, D.; Illig, T.; Floel, A.; et al. Genetic Variants of the FADS Gene Cluster Are Associated with Erythrocyte Membrane LC PUFA Levels in Patients with Mild Cognitive Impairment. J. Nutr. Health Aging 2016, 20, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Yeates, A.J.; Love, T.M.; Engstrom, K.; Mulhern, M.S.; McSorley, E.M.; Grzesik, K.; Alhamdow, A.; Wahlberg, K.; Thurston, S.W.; Davidson, P.W.; et al. Genetic variation in FADS genes is associated with maternal long-chain PUFA status but not with cognitive development of infants in a high fish-eating observational study. Prostaglandins Leukot. Essent. Fat. Acids 2015, 102–103, 13–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Lima Pereira Coelho, N.; Cunha, D.B.; Esteves, A.P.P.; de Aquino Lacerda, E.M.; Theme Filha, M.M. Dietary patterns in pregnancy and birth weight. Rev. Saude Publica 2015, 49, 1–10. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Variables | No Excess Weight Gain | Excess Gestational Weight Gain | p-Value * | ||
|---|---|---|---|---|---|
| Mean | SD | Mean | SD | ||
| Age (years) | 26.3 | 5.8 | 27.9 | 5.8 | <0.001 |
| Maternal education (years of study) | 10.4 | 3.1 | 10.7 | 2.5 | 0.1317 |
| Family income (minimum wage) | 1302 | 803.4 | 1452.8 | 1334 | 0.0504 |
| Number of residents | 3.0 | 1.5 | 4.0 | 1.6 | 0.0020 |
| Pre-gestational BMI | 23.9 | 4.6 | 25.0 | 4.4 | 0.0025 |
| Plasma ALA (%) | 1.8 | 1.7 | 1.4 | 0.9 | 0.9958 |
| Plasma LA (%) | 13.7 | 5.8 | 12.8 | 4.8 | 0.9753 |
| Plasma EPA (%) | 2.5 | 1.9 | 2.2 | 1.8 | 0.9199 |
| Plasma ARA (%) | 1.7 | 1.3 | 1.5 | 1.2 | 0.9690 |
| EPA/ALA | 1.7 | 1.4 | 1.9 | 2.22 | 0.1443 |
| ARA/LA | 0.1 | 0.1 | 0.1 | 0.09 | 0.9837 |
| Variables | Standardized Coefficients | p Value | 95% CI |
|---|---|---|---|
| ALA←SNP rs174561 | −0.25 | 0.039 | −0.48; −0.013 |
| ALA←SNP rs3834458 | 0.29 | 0.020 | 0.04; 0.52 |
| ARA←SNP rs174575 | 0.24 | <0.001 | 0.15; 0.33 |
| ARA/LA←SNP rs174575 | 0.21 | <0.001 | 0.12; 0.30 |
| EPA←SNP rs174575 | −0.12 | 0.009 | −0.22; −0.03 |
| EPA←SNP rs174561 | −0.25 | 0.036 | −0.480; −0.02 |
| EPA←SNP rs3834458 | 0.27 | 0.026 | 0.10; 0.51 |
| EPA/ALA←SNP rs174575 | −0.12 | 0.014 | −0.21; −0.10 |
| IMCPG←SNP rs174575 | 0.17 | <0.001 | 0.11; 0.26 |
| Gestational weight←SNP rs174575 | 0.11 | 0.016 | 0.10; 0.19 |
| Gestational weight←Age | 0.28 | <0.001 | 0.20; 0.35 |
| Gestational weight←Alcohol consumer | 0.14 | <0.001 | 0.07; 0.22 |
| Gestational weight←Mother’s education level | −0.10 | 0.012 | −0.18; −0.02 |
| Gestational weight←Trimester of pregnancy | 0.12 | 0.002 | 0.04; 0.19 |
| RMSEA: 0.0001 | |||
| Variables | Direct Effect | Indirect Effect | Total Effect |
|---|---|---|---|
| Gestational weight←SNP rs174575 | 0.11 | −0.01 | 0.09 |
| Gestational weight←SNP rs 174561 | −0.19 | 0.02 | −0.17 |
| Gestational weight←SNP rs 3834458 | 0.13 | −0.01 | 0.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santana, J.d.M.; Pereira, M.; Carvalho, G.Q.; Gouveia Peluzio, M.d.C.; Drumond Louro, I.; Santos, D.B.d.; Oliveira, A.M. FADS1 and FADS2 Gene Polymorphisms Modulate the Relationship of Omega-3 and Omega-6 Fatty Acid Plasma Concentrations in Gestational Weight Gain: A NISAMI Cohort Study. Nutrients 2022, 14, 1056. https://doi.org/10.3390/nu14051056
Santana JdM, Pereira M, Carvalho GQ, Gouveia Peluzio MdC, Drumond Louro I, Santos DBd, Oliveira AM. FADS1 and FADS2 Gene Polymorphisms Modulate the Relationship of Omega-3 and Omega-6 Fatty Acid Plasma Concentrations in Gestational Weight Gain: A NISAMI Cohort Study. Nutrients. 2022; 14(5):1056. https://doi.org/10.3390/nu14051056
Chicago/Turabian StyleSantana, Jerusa da Mota, Marcos Pereira, Gisele Queiroz Carvalho, Maria do Carmo Gouveia Peluzio, Iúri Drumond Louro, Djanilson Barbosa dos Santos, and Ana Marlucia Oliveira. 2022. "FADS1 and FADS2 Gene Polymorphisms Modulate the Relationship of Omega-3 and Omega-6 Fatty Acid Plasma Concentrations in Gestational Weight Gain: A NISAMI Cohort Study" Nutrients 14, no. 5: 1056. https://doi.org/10.3390/nu14051056
APA StyleSantana, J. d. M., Pereira, M., Carvalho, G. Q., Gouveia Peluzio, M. d. C., Drumond Louro, I., Santos, D. B. d., & Oliveira, A. M. (2022). FADS1 and FADS2 Gene Polymorphisms Modulate the Relationship of Omega-3 and Omega-6 Fatty Acid Plasma Concentrations in Gestational Weight Gain: A NISAMI Cohort Study. Nutrients, 14(5), 1056. https://doi.org/10.3390/nu14051056