
Olive Oil Improves While Trans Fatty Acids Further Aggravate the Hypomethylation of LINE-1 Retrotransposon DNA in an Environmental Carcinogen Model

 , ,
, ,  ,
,  , and
, and
Abstract
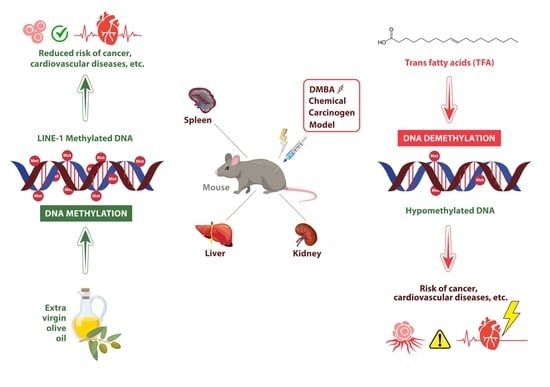
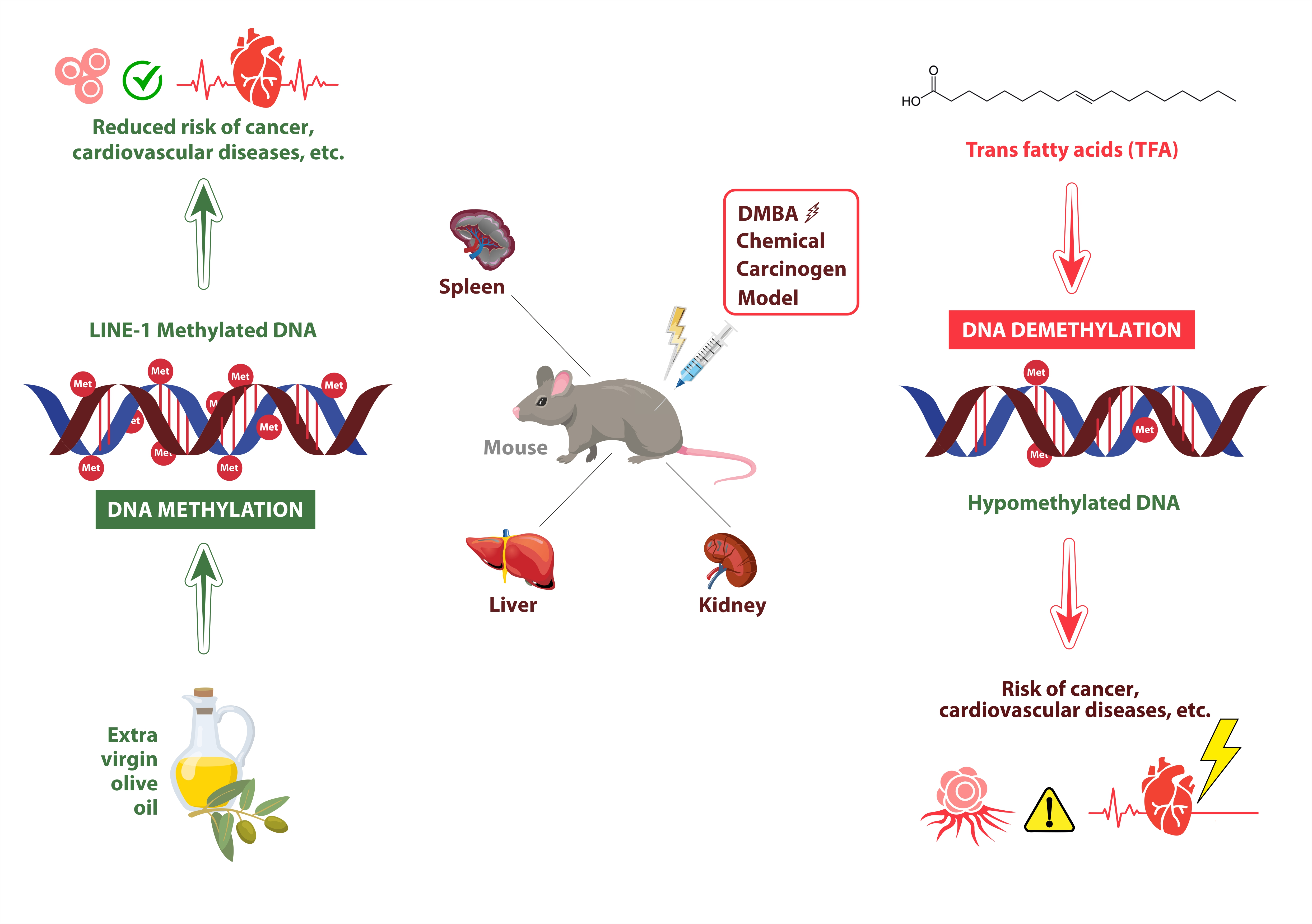
1. Introduction
1.1. Effects of Trans-Fatty Acids
1.2. Effects of Olive Oil
1.3. The Effect of DMBA
1.4. DNA Methylation
1.4.1. DNA Methylation and Malignant Tumors
1.4.2. DNA Methylation and Aging
1.5. Objective
2. Materials and Methods
2.1. Isolation of DNA
2.2. LINE-1 DNA Methylation
2.3. Calculation and Statistical Analysis
3. Results
4. Discussion
4.1. Effect of ROS on the L1-RTP DNA Methylation and Aging
4.2. Effect of DMBA on the L1-RTP DNA Methylation and Aging
4.3. Effect of TFA on the L1-RTP DNA Methylation Pattern
4.4. Protective Effect of OO
4.4.1. The Effect of Fat-Soluble Substances of OO on the LINE-1 DNA Methylation Pattern
4.4.2. Water-Soluble Substances of Olive Oil
4.5. L1-RTP DNA Methylation Patterns
4.5.1. L1-RTP DNA Methylation Pattern in the Liver and Spleen
4.5.2. L1-RTP DNA Methylation Pattern in the Kidneys
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DMBA Control | DMBA + EVOO | DMBA + TFA | |
|---|---|---|---|
| mean LINE-1 methylation | 86.1% | 94.5% | 75.5% |
| distribution | 6.3% | 6.2% | 6.9% |
| p-value | 0.0180 | 0.2852 | 0.0007 |
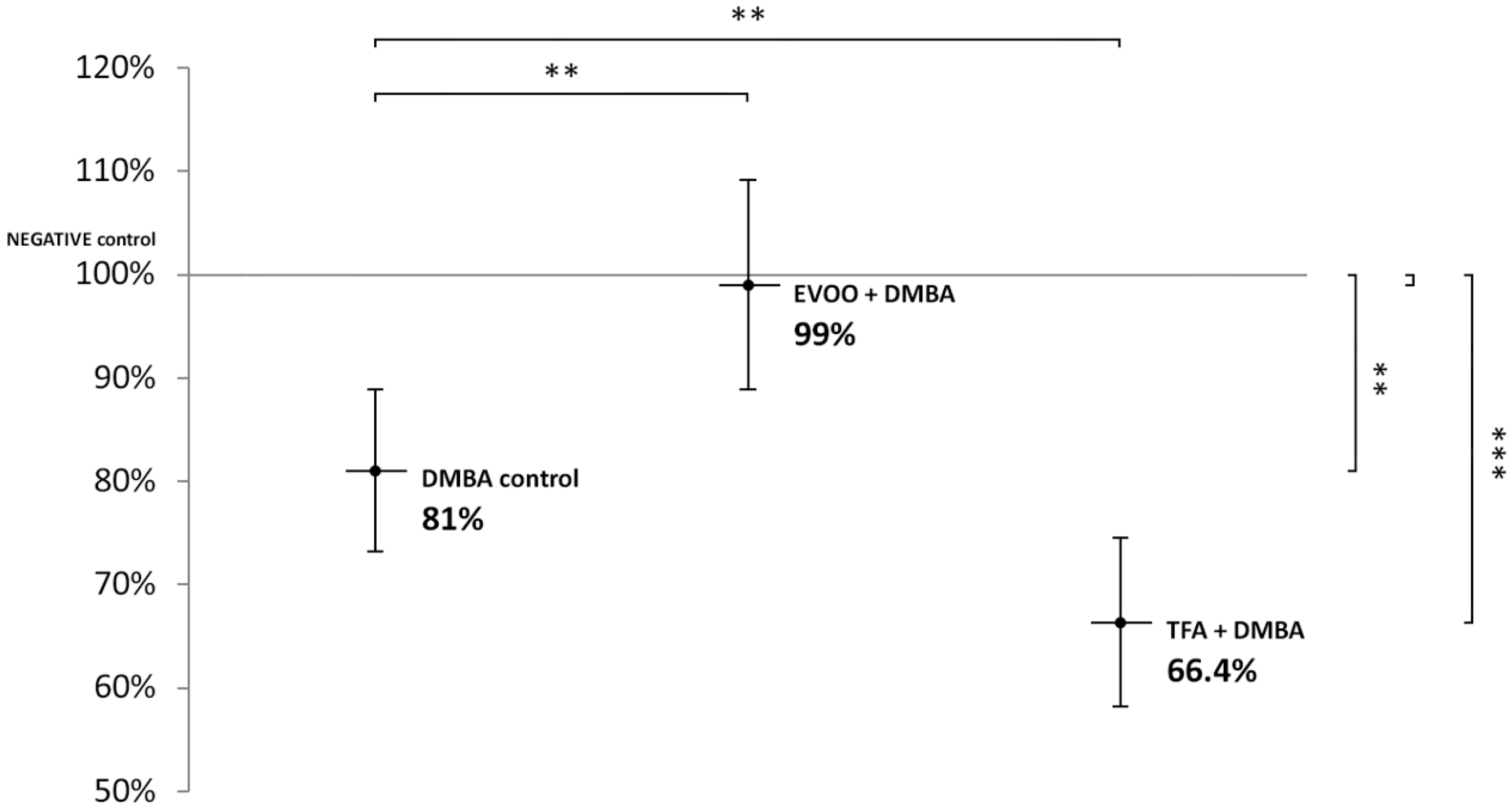
| DMBA Control | DMBA + EVOO | DMBA + TFA | |
|---|---|---|---|
| mean LINE-1 methylation | 81.0% | 99.0% | 66.4% |
| distribution | 7.9% | 10.1% | 8.2% |
| p-value | 0.0042 | 0.8635 | 0.0001 |
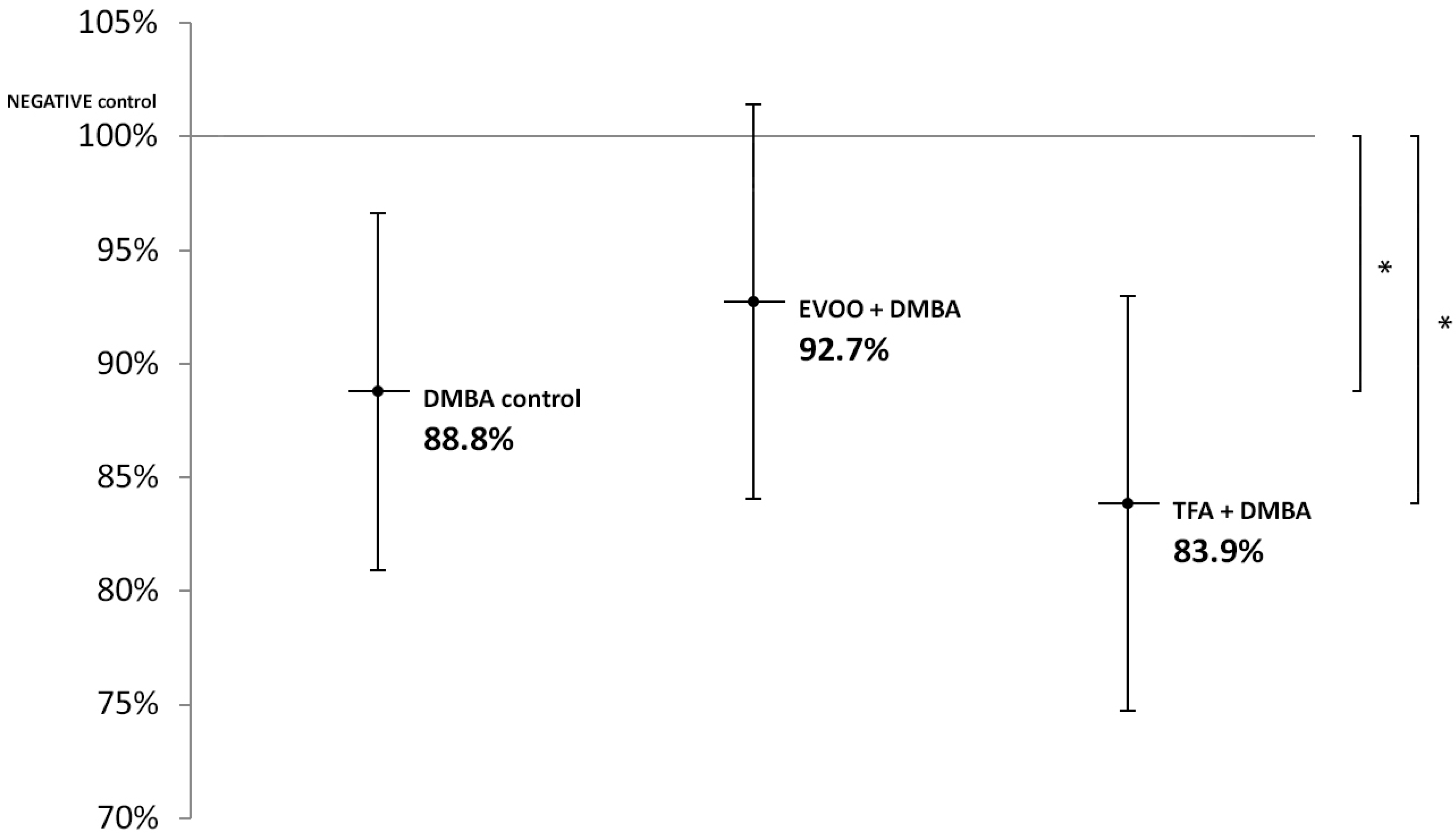
| DMBA Control | DMBA + Olive Oil | DMBA + TFA | |
|---|---|---|---|
| mean LINE-1 methylation | 88.8% | 92.7% | 83.9% |
| distribution | 7.9% | 8.7% | 9.1% |
| p-value | 0.0444 | 0.1861 | 0.0117 |
References
- Ehrlich, M. DNA methylation in cancer: Too much, but also too little. Oncogene 2002, 21, 5400–5413. [Google Scholar] [CrossRef] [PubMed]
- Li, C. Global surveillance of trans-fatty acids. Prev. Chronic Dis. 2019, 16, 190121. [Google Scholar] [CrossRef] [PubMed]
- Craig-Schmidt, M.C. World-wide consumption of trans fatty acids. Atheroscler. Suppl. 2006, 7, 1–4. [Google Scholar] [CrossRef]
- Mondal, P.; Natesh, J.; Penta, D.; Meeran, S.M. Progress and promises of epigenetic drugs and epigenetic diets in cancer prevention and therapy: A clinical update. Semin. Cancer Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Bhardwaj, A.; Aggarwal, R.S.; Seeram, N.P.; Shishodia, S.; Takada, Y. Role of resveratrol in prevention and therapy of cancer: Preclinical and clinical studies. Anticancer Res. 2004, 24, 2783–2840. [Google Scholar]
- Budán, F.; Szabó, I.; Varjas, T.; Nowrasteh, G.; Dávid, T.; Gergely, P.; Varga, Z.; Molnár, K.; Kádár, B.; Orsós, Z. Mixtures of Uncaria and Tabebuia extracts are potentially chemopreventive in CBA/Ca mice: A long-term experiment. Phytother. Res. 2011, 25, 493–500. [Google Scholar] [CrossRef]
- Cui, Y.; Morgenstern, H.; Greenland, S.; Tashkin, D.P.; Mao, J.T.; Cai, L.; Cozen, W.; Mack, T.M.; Lu, Q.Y.; Zhang, Z.F. Dietary flavonoid intake and lung cancer—A population-based case-control study. Cancer 2008, 112, 2241–2248. [Google Scholar] [CrossRef]
- Narayanan, B.A. Chemopreventive agents alters global gene expression pattern: Predicting their mode of action and targets. Curr. Cancer Drug Targets 2006, 6, 711–727. [Google Scholar] [CrossRef]
- Varjas, T.; Nowrasteh, G.; Budán, F.; Nadasi, E.; Horváth, G.; Makai, S.; Gracza, T.; Cseh, J.; Ember, I. Chemopreventive effect of Panax ginseng. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Product Deriv. 2009, 23, 1399–1403. [Google Scholar]
- Varjas, T.; Nowrasteh, G.; Budán, F.; Horváth, G.; Cseh, J.; Gyöngyi, Z.; Makai, S.; Ember, I. The effect of fenugreek on the gene expression of arachidonic acid metabolizing enzymes. Phytother. Res. 2011, 25, 221–227. [Google Scholar] [CrossRef]
- Fernández del Río, L.; Gutiérrez-Casado, E.; Varela-López, A.; Villalba, J.M. Olive oil and the hallmarks of aging. Molecules 2016, 21, 163. [Google Scholar] [CrossRef] [PubMed]
- Giovannelli, L. Beneficial effects of olive oil phenols on the aging process: Experimental evidence and possible mechanisms of action. Nutr. Aging 2012, 1, 207–223. [Google Scholar] [CrossRef]
- Andreadou, I.; Iliodromitis, E.K.; Mikros, E.; Constantinou, M.; Agalias, A.; Magiatis, P.; Skaltsounis, A.L.; Kamber, E.; Tsantili-Kakoulidou, A.; Kremastinos, D.T. The olive constituent oleuropein exhibits anti-ischemic, antioxidative, and hypolipidemic effects in anesthetized rabbits. J. Nutr. 2006, 136, 2213–2219. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.J.; Goodman, S.J.; Kobor, M.S. DNA methylation and healthy human aging. Aging Cell 2015, 14, 924–932. [Google Scholar] [CrossRef]
- Abd El-Aal, Y.A.; Abdel-Fattah, D.M.; Ahmed, K.E.-D. Some biochemical studies on trans fatty acid-containing diet. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 1753–1757. [Google Scholar] [CrossRef]
- Zhuang, P.; Zhang, Y.; He, W.; Chen, X.; Chen, J.; He, L.; Mao, L.; Wu, F.; Jiao, J. Dietary fats in relation to total and cause-specific mortality in a prospective cohort of 521 120 individuals with 16 years of follow-up. Circ. Res. 2019, 124, 757–768. [Google Scholar] [CrossRef]
- Anjom-Shoae, J.; Sadeghi, O.; Larijani, B.; Esmaillzadeh, A. Dietary intake and serum levels of trans fatty acids and risk of breast cancer: A systematic review and dose-response meta-analysis of prospective studies. Clin. Nutr. 2020, 39, 755–764. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Katan, M.B.; Ascherio, A.; Stampfer, M.J.; Willett, W.C. Trans fatty acids and cardiovascular disease. N. Eng. J. Med. 2006, 354, 1601–1613. [Google Scholar] [CrossRef] [PubMed]
- Alfin-Slater, R.B.; Wells, A.F.; Aftergood, L.; Deuel, H.J., Jr. Nutritive value and safety of hydrogenated vegetable fats as evaluated by long-term feeding experiments with rats. J. Nutr. 1957, 63, 241–261. [Google Scholar] [CrossRef]
- Katabathina, V.S.; Vikram, R.; Nagar, A.M.; Tamboli, P.; Menias, C.O.; Prasad, S.R. Mesenchymal neoplasms of the kidney in adults: Imaging spectrum with radiologic-pathologic correlation. Radiographics 2010, 30, 1525–1540. [Google Scholar] [CrossRef]
- Beltrán, G.; Del Rio, C.; Sánchez, S.; Martínez, L. Influence of harvest date and crop yield on the fatty acid composition of virgin olive oils from cv. Picual. J. Agric. Food Chem. 2004, 52, 3434–3440. [Google Scholar] [CrossRef] [PubMed]
- Lipworth, L.; Martínez, M.a.E.; Angell, J.; Hsieh, C.-C.; Trichopoulos, D. Olive oil and human cancer: An assessment of the evidence. Prev. Med. 1997, 26, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Owen, R.; Giacosa, A.; Hull, W.; Haubner, R.; Spiegelhalder, B.; Bartsch, H. The antioxidant/anticancer potential of phenolic compounds isolated from olive oil. Eur. J. Cancer 2000, 36, 1235–1247. [Google Scholar] [CrossRef]
- Pelucchi, C.; Bosetti, C.; Negri, E.; Lipworth, L.; La Vecchia, C. Olive oil and cancer risk: An update of epidemiological findings through 2010. Curr. Pharm. Des. 2011, 17, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Fortes, C.; Forastiere, F.; Farchi, S.; Mallone, S.; Trequattrinni, T.; Anatra, F.; Schmid, G.; Perucci, C.A. The protective effect of the Mediterranean diet on lung cancer. Nutr. Cancer 2003, 46, 30–37. [Google Scholar] [CrossRef]
- Bosetti, C.; La Vecchia, C.; Talamini, R.; Negri, E.; Levi, F.; Dal Maso, L.; Franceschi, S. Food groups and laryngeal cancer risk: A case-control study from Italy and Switzerland. Int. J. Cancer 2002, 100, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Brinkman, M.T.; Buntinx, F.; Kellen, E.; Van Dongen, M.C.; Dagnelie, P.C.; Muls, E.; Zeegers, M.P. Consumption of animal products, olive oil and dietary fat and results from the Belgian case–control study on bladder cancer risk. Eur. J. Cancer 2011, 47, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Anisimov, V.N. Carcinogenesis and aging 20 years after: Escaping horizon. Mech. Ageing Dev. 2009, 130, 105–121. [Google Scholar] [CrossRef]
- Ember, I.; Gyöngyi, Z.; Kiss, I.; Ghodratollah, N.; Arany, I. The possible relationship between onco/suppressor gene expression and carcinogen exposure in vivo: Evaluation of a potential biomarker in preventive and predictive medicine. Anticancer Res. 2002, 22, 2109–2116. [Google Scholar]
- Perjési, P.; Gyöngyi, Z.; Bayer, Z. Effect of E-2-(4’-methoxybenzylidene)-1-benzosuberone on the 7, 12-dimethylbenz [alpha] anthracene-induced onco/suppressor gene action in vivo II: A 48-hour experiment. Anticancer Res. 2000, 20, 1839–1848. [Google Scholar] [PubMed]
- Szabo, L.; Molnar, R.; Tomesz, A.; Deutsch, A.; Darago, R.; Nowrasteh, G.; Varjas, T.; Nemeth, B.; Budan, F.; Kiss, I. The effects of flavonoids, green tea polyphenols and coffee on DMBA induced LINE-1 DNA hypomethylation. PLoS ONE 2021, 16, e0250157. [Google Scholar] [CrossRef] [PubMed]
- Weidner, C.I.; Lin, Q.; Koch, C.M.; Eisele, L.; Beier, F.; Ziegler, P.; Bauerschlag, D.O.; Jöckel, K.-H.; Erbel, R.; Mühleisen, T.W. Aging of blood can be tracked by DNA methylation changes at just three CpG sites. Genome Biol. 2014, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Baba, Y.; Yagi, T.; Sawayama, H.; Hiyoshi, Y.; Ishimoto, T.; Iwatsuki, M.; Miyamoto, Y.; Yoshida, N.; Baba, H. Long interspersed element-1 methylation level as a prognostic biomarker in gastrointestinal cancers. Digestion 2018, 97, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Orozco, J.M.; Saxton, R.A.; Condon, K.J.; Liu, G.Y.; Krawczyk, P.A.; Scaria, S.M.; Harper, J.W.; Gygi, S.P.; Sabatini, D.M. SAMTOR is an S-adenosylmethionine sensor for the mTORC1 pathway. Science 2017, 358, 813–818. [Google Scholar] [CrossRef]
- Fraga, M.F.; Agrelo, R.; Esteller, M. Cross-talk between aging and cancer: The epigenetic language. Annu. N. Y. Acad. Sci. 2007, 1100, 60–74. [Google Scholar] [CrossRef] [PubMed]
- Laird, P.W. Oncogenic mechanisms mediated by DNA methylation. Mol. Med. Today 1997, 3, 223–229. [Google Scholar] [CrossRef]
- Li, M.; Liu, W.; Yuan, T.; Bai, R.; Liu, G.-H.; Zhang, W.; Qu, J. DNA methylome: Unveiling your biological age. Protein Cell 2013, 4, 723–725. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lim, U.; Song, M. DNA methylation as a biomarker of aging in epidemiologic studies. Cancer Epigenetics Precis. Med. 2018, 1856, 219–231. [Google Scholar]
- Snir, S.; vonHoldt, B.M.; Pellegrini, M. A statistical framework to identify deviation from time linearity in epigenetic aging. PLoS Comput. Biol. 2016, 12, e1005183. [Google Scholar] [CrossRef]
- Zampieri, M.; Ciccarone, F.; Calabrese, R.; Franceschi, C.; Bürkle, A.; Caiafa, P. Reconfiguration of DNA methylation in aging. Mech. Ageing Dev. 2015, 151, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Ardeljan, D.; Taylor, M.S.; Ting, D.T.; Burns, K.H. The human long interspersed element-1 retrotransposon: An emerging biomarker of neoplasia. Clin. Chem. 2017, 63, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, W.; Erichsen, L.; Ott, P.; Schulz, W.A.; Fischer, J.C.; Arauzo-Bravo, M.J.; Bendhack, M.L.; Hassan, M.; Santourlidis, S. Aging-associated distinctive DNA methylation changes of LINE-1 retrotransposons in pure cell-free DNA from human blood. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tangkijvanich, P.; Hourpai, N.; Rattanatanyong, P.; Wisedopas, N.; Mahachai, V.; Mutirangura, A. Serum LINE-1 hypomethylation as a potential prognostic marker for hepatocellular carcinoma. Clinica Chimica Acta 2007, 379, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Budán, F.; Varjas, T.; Nowrasteh, G.; Varga, Z.; Boncz, I.; Cseh, J.; Prantner, I.; Antal, T.; Pazsit, E.; GŐBEL, G. Early Modification of c-myc, Ha-ras and p53 Expressions by N-Methyl-N-nitrosourea. In Vivo 2008, 22, 793–797. [Google Scholar]
- Matheu, A.; Maraver, A.; Klatt, P.; Flores, I.; Garcia-Cao, I.; Borras, C.; Flores, J.M.; Viña, J.; Blasco, M.A.; Serrano, M. Delayed ageing through damage protection by the Arf/p53 pathway. Nature 2007, 448, 375–379. [Google Scholar] [CrossRef]
- Ono, T.; Takahashi, N.; Okada, S. Age-associated changes in DNA methylation and mRNA level of the c-myc gene in spleen and liver of mice. Mutat. Res./DNAging 1989, 219, 39–50. [Google Scholar] [CrossRef]
- Hannum, G.; Guinney, J.; Zhao, L.; Zhang, L.; Hughes, G.; Sadda, S.; Klotzle, B.; Bibikova, M.; Fan, J.-B.; Gao, Y. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol. Cell 2013, 49, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Hunt, N.J.; Kang, S.W.S.; Lockwood, G.P.; Le Couteur, D.G.; Cogger, V.C. Hallmarks of aging in the liver. Comput. Struct. Biotechnol. J. 2019, 17, 1151–1161. [Google Scholar] [CrossRef]
- Marques-Rocha, J.L.; Milagro, F.I.; Mansego, M.L.; Mourão, D.M.; Martínez, J.A.; Bressan, J. LINE-1 methylation is positively associated with healthier lifestyle but inversely related to body fat mass in healthy young individuals. Epigenetics 2016, 11, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Mackenbach, J.P.; Valverde, J.R.; Bopp, M.; Brønnum-Hansen, H.; Deboosere, P.; Kalediene, R.; Kovács, K.; Leinsalu, M.; Martikainen, P.; Menvielle, G. Determinants of inequalities in life expectancy: An international comparative study of eight risk factors. Lancet Public Health 2019, 4, e529–e537. [Google Scholar] [CrossRef]
- Ghazani, S.; Marangoni, A. Nutrition and food grains in Encyclopedia of Food Grains; Academic Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Budán, F.; Varjas, T.; Nowrasteh, G.; Prantner, I.; Varga, Z.; Ember, Á.; Cseh, J.; Gombos, K.; Pázsit, E.; Gőbel, G. Early Modification of c-myc, Ha-ras and p53 Expressions by Chemical Carcinogens (DMBA, MNU). In Vivo 2009, 23, 591–598. [Google Scholar]
- Smith, B.K.; Robinson, L.E.; Nam, R.; Ma, D.W. Trans-fatty acids and cancer: A mini-review. Br. J. Nutr. 2009, 102, 1254–1266. [Google Scholar] [CrossRef]
- Furlan, D.; Trapani, D.; Berrino, E.; Debernardi, C.; Panero, M.; Libera, L.; Sahnane, N.; Riva, C.; Tibiletti, M.G.; Sessa, F. Oxidative DNA damage induces hypomethylation in a compromised base excision repair colorectal tumourigenesis. Br. J. Cancer 2017, 116, 793–801. [Google Scholar] [CrossRef]
- Storz, P. Reactive oxygen species in tumor progression. Front. Biosci. 2005, 10, 1881–1896. [Google Scholar] [CrossRef]
- Park, L.K.; Friso, S.; Choi, S.-W. Nutritional influences on epigenetics and age-related disease. Proc. Nutr. Soc. 2012, 71, 75–83. [Google Scholar] [CrossRef]
- Ponnaluri, V.C.; Estève, P.-O.; Ruse, C.I.; Pradhan, S. S-adenosylhomocysteine hydrolase participates in DNA methylation inheritance. J. Mol. Biol. 2018, 430, 2051–2065. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. and Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef]
- Karouzakis, E.; Gay, R.E.; Gay, S.; Neidhart, M. Increased recycling of polyamines is associated with global DNA hypomethylation in rheumatoid arthritis synovial fibroblasts. Arthritis Rheum. 2012, 64, 1809–1817. [Google Scholar] [CrossRef]
- Lertratanangkoon, K.; Wu, C.J.; Savaraj, N.; Thomas, M.L. Alterations of DNA methylation by glutathione depletion. Cancer Lett. 1997, 120, 149–156. [Google Scholar] [CrossRef]
- Tibbetts, A.S.; Appling, D.R. Compartmentalization of Mammalian folate-mediated one-carbon metabolism. Annu. Review Nutr. 2010, 30, 57–81. [Google Scholar] [CrossRef]
- Beetch, M.; Stefanska, B. DNA Methylation in Anti-cancer Effects of Dietary Catechols and Stilbenoids: An Overview of Underlying Mechanisms. In Handbook of Nutrition, Diet, and Epigenetics; Patel, V., Preedy, V., Eds.; Springer: Cham, Switzerland, 2019; pp. 1819–1844. [Google Scholar]
- Zhu, Y.; Carvey, P.M.; Ling, Z. Age-related changes in glutathione and glutathione-related enzymes in rat brain. Brain Res. 2006, 1090, 35–44. [Google Scholar] [CrossRef]
- Bryk, D.; Zapolska-Downar, D.; Malecki, M.; Hajdukiewicz, K.; Sitkiewicz, D. Trans fatty acids induce a proinflammatory response in endothelial cells through ROS-dependent nuclear factor-κB activation. J. Physiol. Pharmacol. 2011, 62, 229. [Google Scholar]
- RM, P.E.P.; Stein, I.; Bramovitch, R.; Amit, S.; Kasem, S.; Gutkovich-Pyest, E.; Urieli-Shoval, S.; Galun, E.; Ben-Neriah, Y. NF-kappaB functions as a tumour promoter in inflammation-associated cancer. Nature 2004, 431, 461–466. [Google Scholar]
- De Souza, V.R.; Cabrera, W.K.; Galvan, A.; Ribeiro, O.G.; De Franco, M.; Vorraro, F.; Starobinas, N.; Massa, S.; Dragani, T.A.; Ibanez, O.M. Aryl hydrocarbon receptor polymorphism modulates DMBA-induced inflammation and carcinogenesis in phenotypically selected mice. Int. J. Cancer 2009, 124, 1478–1482. [Google Scholar] [CrossRef]
- Molnar, R.; Szabo, L.; Tomesz, A.; Deutsch, A.; Darago, R.; Ghodratollah, N.; Varjas, T.; Nemeth, B.; Budan, F.; Kiss, I. In vivo effects of olive oil and trans-fatty acids on miR-134, miR-132, miR-124-1, miR-9-3 and mTORC1 gene expression in a DMBA-treated mouse model. PLoS ONE 2021, 16, e0246022. [Google Scholar] [CrossRef]
- Hendrayani, S.-F.; Al-Harbi, B.; Al-Ansari, M.M.; Silva, G.; Aboussekhra, A. The inflammatory/cancer-related IL-6/STAT3/NF-κB positive feedback loop includes AUF1 and maintains the active state of breast myofibroblasts. Oncotarget 2016, 7, 41974. [Google Scholar] [CrossRef]
- Kyriakis, J.M.; Avruch, J. Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol. Rev. 2001, 81, 807–869. [Google Scholar] [CrossRef]
- De Cecco, M.; Ito, T.; Petrashen, A.P.; Elias, A.E.; Skvir, N.J.; Criscione, S.W.; Caligiana, A.; Brocculi, G.; Adney, E.M.; Boeke, J.D. L1 drives IFN in senescent cells and promotes age-associated inflammation. Nature 2019, 566, 73–78. [Google Scholar] [CrossRef]
- Apte, R.N.; Dotan, S.; Elkabets, M.; White, M.R.; Reich, E.; Carmi, Y.; Song, X.; Dvozkin, T.; Krelin, Y.; Voronov, E. The involvement of IL-1 in tumorigenesis, tumor invasiveness, metastasis and tumor-host interactions. Cancer Metastasis Rev. 2006, 25, 387–408. [Google Scholar] [CrossRef]
- Stetler-Stevenson, W.G.; Yu, A.E. Proteases in invasion: Matrix metalloproteinases. Semin. Cancer Biol. 2001, 11, 143–152. [Google Scholar] [CrossRef]
- Yeh, C.-B.; Hsieh, M.-J.; Hsieh, Y.-H.; Chien, M.-H.; Chiou, H.-L.; Yang, S.-F. Antimetastatic effects of norcantharidin on hepatocellular carcinoma by transcriptional inhibition of MMP-9 through modulation of NF-kB activity. PLoS ONE 2012, 7, e31055. [Google Scholar] [CrossRef]
- Wajant, H. The Role of TNF in Cancer. Death Recept. Cogn. Ligands Cancer 2009, 49, 1–15. [Google Scholar]
- Webster, G.A.; Perkins, N.D. Transcriptional cross talk between NF-κB and p53. Mol. Cell. Biol. 1999, 19, 3485–3495. [Google Scholar] [CrossRef]
- Brasier, A.R. The nuclear factor-κB–interleukin-6 signalling pathway mediating vascular inflammation. Cardiovasc. Res. 2010, 86, 211–218. [Google Scholar] [CrossRef]
- Haque, M.W.; Bose, P.; Siddique, M.U.M.; Sunita, P.; Lapenna, A.; Pattanayak, S.P. Taxifolin binds with LXR (α & β) to attenuate DMBA-induced mammary carcinogenesis through mTOR/Maf-1/PTEN pathway. Biomed. Pharmacother. 2018, 105, 27–36. [Google Scholar]
- Oteng, A.-B.; Kersten, S. Mechanisms of action of trans fatty acids. Adv. Nutr. 2020, 11, 697–708. [Google Scholar] [CrossRef]
- Huang, Z.; Wang, B.; Pace, R.D.; Yoon, S. Trans fat intake lowers total cholesterol and high-density lipoprotein cholesterol levels without changing insulin sensitivity index in Wistar rats. Nutr. Res. 2009, 29, 206–212. [Google Scholar] [CrossRef]
- Pervaiz, S.; Taneja, R.; Ghaffari, S. Oxidative stress regulation of stem and progenitor cells. Antioxidants Redox Signal. 2009, 11, 2777–2789. [Google Scholar] [CrossRef]
- Morrow, J.D.; Awad, J.A.; Boss, H.J.; Blair, I.A.; Roberts, L.J. Non-cyclooxygenase-derived prostanoids (F2-isoprostanes) are formed in situ on phospholipids. Proc. Natl. Acad. Sci. USA 1992, 89, 10721–10725. [Google Scholar] [CrossRef]
- Clandinin, M.; Cheema, S.; Field, C.; Garg, M.; Venkatraman, J.; Clandinin, T. Dietary fat: Exogenous determination of membrane structure and cell function. FASEB J. 1991, 5, 2761–2769. [Google Scholar] [CrossRef]
- Ma, D.W. Lipid mediators in membrane rafts are important determinants of human health and disease. Appl. Physiol. Nutr. Metab. 2007, 32, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Prior, I.A.; Hancock, J.F. Compartmentalization of Ras proteins. J. Cell Sci. 2001, 114, 1603–1608. [Google Scholar] [CrossRef] [PubMed]
- Czerska, M.; Zieliński, M.; Gromadzińska, J. Isoprostanes–A novel major group of oxidative stress markers. Int. J. Occup. Med. Environ. Health 2016, 29, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Comporti, M.; Signorini, C.; Arezzini, B.; Vecchio, D.; Monaco, B.; Gardi, C. F2-isoprostanes are not just markers of oxidative stress. Free Radic. Biol. Med. 2008, 44, 247–256. [Google Scholar] [CrossRef]
- Ward, W.F.; Qi, W.; Remmen, H.V.; Zackert, W.E.; Roberts, L.J.; Richardson, A. Effects of age and caloric restriction on lipid peroxidation: Measurement of oxidative stress by F2-isoprostane levels. J. Gerontol. Ser. A Biol. Sci. Med. Sc. 2005, 60, 847–851. [Google Scholar] [CrossRef]
- Watson, R.E.; Curtin, G.M.; Doolittle, D.J.; Goodman, J.I. Progressive alterations in global and GC-rich DNA methylation during tumorigenesis. Toxicol. Sci. 2003, 75, 289–299. [Google Scholar] [CrossRef][Green Version]
- Tiwari, B.; Jones, A.E.; Abrams, J.M. Transposons, p53 and genome security. Trends Genet. 2018, 34, 846–855. [Google Scholar] [CrossRef]
- Rufini, A.; Tucci, P.; Celardo, I.; Melino, G. Senescence and aging: The critical roles of p53. Oncogene 2013, 32, 5129–5143. [Google Scholar] [CrossRef]
- Shuang, T.; Wang, M.; Zhou, Y.; Shi, C.; Wang, D. NF-κB1, c-Rel, and ELK1 inhibit miR-134 expression leading to TAB1 upregulation in paclitaxel-resistant human ovarian cancer. Oncotarget 2017, 8, 24853. [Google Scholar] [CrossRef]
- Wu, D.; Prives, C. Relevance of the p53–MDM2 axis to aging. Cell Death Differ. 2018, 25, 169–179. [Google Scholar] [CrossRef]
- Tomesz, A.; Szabo, L.; Molnar, R.; Deutsch, A.; Darago, R.; Mathe, D.; Budan, F.; Ghodratollah, N.; Varjas, T.; Nemeth, B. Effect of 7, 12-Dimethylbenz (α) anthracene on the Expression of miR-330, miR-29a, miR-9-1, miR-9-3 and the mTORC1 Gene in CBA/Ca Mice. In Vivo 2020, 34, 2337–2343. [Google Scholar] [CrossRef] [PubMed]
- Padlianah; Arif, M.; Yustisia, I. Blood chemistry profiles of DMBA-induced mammary tumor in female sprague dawley rats. In Proceedings of the International Conference on Bioinformatics and Nano-Medicine from Natural Resources for Biomedical Research: 3rd Annual Scientific Meeting for Biomedical Sciences, Malang, Indonesia, 21–23 November 2018; p. 020043. [Google Scholar]
- Mao, Z.; Zhang, W. Role of mTOR in glucose and lipid metabolism. Int. J. Mol. Sci. 2018, 19, 2043. [Google Scholar] [CrossRef] [PubMed]
- Yeo, E.-J. Hypoxia and aging. Exp. Mol. Med. 2019, 51, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, B.P.; Sinclair, D.A. Small molecule SIRT1 activators for the treatment of aging and age-related diseases. Trends Pharmacol. Sci. 2014, 35, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Khew-Goodall, Y.; Goodall, G.J. Myc-modulated miR-9 makes more metastases. Nat. Cell Biol. 2010, 12, 209–211. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, M.S.; Bilodeau, J.-F.; Larose, J.; Greffard, K.; Julien, P.; Barbier, O.; Rudkowska, I. Modulation of the biomarkers of inflammation and oxidative stress by ruminant trans fatty acids and dairy proteins in vascular endothelial cells (HUVEC). Prostaglandins Leukot. Essent. Fat. Acids 2017, 126, 64–71. [Google Scholar] [CrossRef]
- Zhang, K.; Yang, X.; Zhao, Q.; Li, Z.; Fu, F.; Zhang, H.; Zheng, M.; Zhang, S. Molecular mechanism of stem cell differentiation into adipocytes and adipocyte differentiation of malignant tumor. Stem Cells Int. 2020, 2020, 8892300. [Google Scholar] [CrossRef]
- Flores-Sierra, J.; Arredondo-Guerrero, M.; Cervantes-Paz, B.; Rodríguez-Ríos, D.; Alvarado-Caudillo, Y.; Nielsen, F.C.; Wrobel, K.; Wrobel, K.; Zaina, S.; Lund, G. The trans fatty acid elaidate affects the global DNA methylation profile of cultured cells and in vivo. Lipids Health Dis. 2016, 15, 1–7. [Google Scholar] [CrossRef]
- Menaa, F.; Menaa, A.; Menaa, B.; Tréton, J. Trans-fatty acids, dangerous bonds for health? A background review paper of their use, consumption, health implications and regulation in France. Eur. J. Nutr. 2013, 52, 1289–1302. [Google Scholar] [CrossRef]
- Papsdorf, K.; Brunet, A. Linking lipid metabolism to chromatin regulation in aging. Trends Cell Biol. 2019, 29, 97–116. [Google Scholar] [CrossRef]
- Hąc-Wydro, K.; Wydro, P. The influence of fatty acids on model cholesterol/phospholipid membranes. Chem. Phys. Lipids 2007, 150, 66–81. [Google Scholar] [CrossRef] [PubMed]
- Silva-Martínez, G.A.; Rodríguez-Ríos, D.; Alvarado-Caudillo, Y.; Vaquero, A.; Esteller, M.; Carmona, F.J.; Moran, S.; Nielsen, F.C.; Wickström-Lindholm, M.; Wrobel, K. Arachidonic and oleic acid exert distinct effects on the DNA methylome. Epigenetics 2016, 11, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Youssef, J.; Badr, M. Role of peroxisome proliferator-activated receptors in inflammation control. J. Biomed. Biotechnol. 2004, 2004, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Medeiros-de-Moraes, I.M.; Gonçalves-de-Albuquerque, C.F.; Kurz, A.R.; Oliveira, F.M.d.J.; Abreu, V.H.P.d.; Torres, R.C.; Carvalho, V.F.; Estato, V.; Bozza, P.T.; Sperandio, M. Omega-9 oleic acid, the main compound of olive oil, mitigates inflammation during experimental sepsis. Oxidative Med. Cell. Longev. 2018, 2018, 6053492. [Google Scholar] [CrossRef]
- Bento-Abreu, A.; Tabernero, A.; Medina, J.M. Peroxisome proliferator-activated receptor-alpha is required for the neurotrophic effect of oleic acid in neurons. J. Neurochem. 2007, 103, 871–881. [Google Scholar] [CrossRef]
- Fidaleo, M.; Fanelli, F.; Paola Ceru, M.; Moreno, S. Neuroprotective properties of peroxisome proliferator-activated receptor alpha (PPARα) and its lipid ligands. Curr. Med. Chem. 2014, 21, 2803–2821. [Google Scholar] [CrossRef]
- Najt, C.P.; Khan, S.A.; Heden, T.D.; Witthuhn, B.A.; Perez, M.; Heier, J.L.; Mead, L.E.; Franklin, M.P.; Karanja, K.K.; Graham, M.J. Lipid droplet-derived monounsaturated fatty acids traffic via PLIN5 to allosterically activate SIRT1. Mol. Cell 2020, 77, 810–824. e8. [Google Scholar] [CrossRef]
- Heo, J.; Lim, J.; Lee, S.; Jeong, J.; Kang, H.; Kim, Y.; Kang, J.W.; Yu, H.Y.; Jeong, E.M.; Kim, K. Sirt1 regulates DNA methylation and differentiation potential of embryonic stem cells by antagonizing Dnmt3l. Cell Rep. 2017, 18, 1930–1945. [Google Scholar] [CrossRef]
- Peng, L.; Yuan, Z.; Ling, H.; Fukasawa, K.; Robertson, K.; Olashaw, N.; Koomen, J.; Chen, J.; Lane, W.S.; Seto, E. SIRT1 deacetylates the DNA methyltransferase 1 (DNMT1) protein and alters its activities. Mol. Cell. Biol. 2011, 31, 4720–4734. [Google Scholar] [CrossRef]
- Notarnicola, M.; Tutino, V.; De Nunzio, V.; Dituri, F.; Caruso, M.G.; Giannelli, G. Dietary ω-3 polyunsaturated fatty acids inhibit tumor growth in transgenic ApcMin/+ mice, correlating with CB1 receptor up-regulation. Int. J. Mol. Sci. 2017, 18, 485. [Google Scholar] [CrossRef]
- Poole, C.J.; Lodh, A.; Choi, J.-H.; Van Riggelen, J. MYC deregulates TET1 and TET2 expression to control global DNA (hydroxy) methylation and gene expression to maintain a neoplastic phenotype in T-ALL. Epigenetics Chromatin 2019, 12, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.T.; Gu, W. SIRT1: Regulator of p53 deacetylation. Genes Cancer 2013, 4, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Vassiliou, E.K.; Gonzalez, A.; Garcia, C.; Tadros, J.H.; Chakraborty, G.; Toney, J.H. Oleic acid and peanut oil high in oleic acid reverse the inhibitory effect of insulin production of the inflammatory cytokine TNF-α both in vitro and in vivo systems. Lipids Health Dis. 2009, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bermúdez, B.; Pacheco, Y.M.; López, S.; Abia, R.; Muriana, F.J. Digestion and absorption of olive oil. Grasas y aceites 2004, 55, 1–10. [Google Scholar]
- Nakbi, A.; Tayeb, W.; Dabbou, S.; Issaoui, M.; Grissa, A.K.; Attia, N.; Hammami, M. Dietary olive oil effect on antioxidant status and fatty acid profile in the erythrocyte of 2, 4-D-exposed rats. Lipids Health Dis. 2010, 9, 1–10. [Google Scholar] [CrossRef]
- Seiquer, I.; Rueda, A.; Olalla, M.; Cabrera-Vique, C. Assessing the bioavailability of polyphenols and antioxidant properties of extra virgin argan oil by simulated digestion and Caco-2 cell assays. Comparative study with extra virgin olive oil. Food Chem. 2015, 188, 496–503. [Google Scholar] [CrossRef]
- Ferreri, C.; Masi, A.; Sansone, A.; Giacometti, G.; Larocca, A.V.; Menounou, G.; Scanferlato, R.; Tortorella, S.; Rota, D.; Conti, M. Fatty acids in membranes as homeostatic, metabolic and nutritional biomarkers: Recent advancements in analytics and diagnostics. Diagnostics 2017, 7, 1. [Google Scholar] [CrossRef]
- Hornedo-Ortega, R.; Cerezo, A.B.; De Pablos, R.M.; Krisa, S.; Richard, T.; García-Parrilla, M.C.; Troncoso, A.M. Phenolic compounds characteristic of the mediterranean diet in mitigating microglia-mediated neuroinflammation. Front. Cell. Neurosci. 2018, 373. [Google Scholar] [CrossRef]
- Giusti, L.; Angeloni, C.; Barbalace, M.C.; Lacerenza, S.; Ciregia, F.; Ronci, M.; Urbani, A.; Manera, C.; Digiacomo, M.; Macchia, M. A proteomic approach to uncover neuroprotective mechanisms of oleocanthal against oxidative stress. Int. J. Mol. Sci. 2018, 19, 2329. [Google Scholar] [CrossRef]
- Kouka, P.; Tsakiri, G.; Tzortzi, D.; Dimopoulou, S.; Sarikaki, G.; Stathopoulos, P.; Veskoukis, A.S.; Halabalaki, M.; Skaltsounis, A.-L.; Kouretas, D. The polyphenolic composition of extracts derived from different greek extra virgin olive oils is correlated with their antioxidant potency. Oxidative Med. Cell. Longev. 2019, 2019, 1870965. [Google Scholar] [CrossRef]
- López-Miranda, J.; Pérez-Jiménez, F.; Ros, E.; De Caterina, R.; Badimón, L.; Covas, M.I.; Escrich, E.; Ordovás, J.M.; Soriguer, F.; Abia, R. Olive oil and health: Summary of the II international conference on olive oil and health consensus report, Jaén and Córdoba (Spain) 2008. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Malliou, F.; Andreadou, I.; Gonzalez, F.J.; Lazou, A.; Xepapadaki, E.; Vallianou, I.; Lambrinidis, G.; Mikros, E.; Marselos, M.; Skaltsounis, A.-L. The olive constituent oleuropein, as a PPARα agonist, markedly reduces serum triglycerides. J. Nutr. Biochem. 2018, 59, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Khanfar, M.A.; Bardaweel, S.K.; Akl, M.R.; El Sayed, K.A. Olive oil-derived oleocanthal as potent inhibitor of mammalian target of rapamycin: Biological evaluation and molecular modeling studies. Phytother. Res. 2015, 29, 1776–1782. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Chen, H.; Ren, S.; Xia, M.; Zhu, J.; Liu, Y.; Zhang, L.; Tang, L.; Sun, L.; Liu, H. Aberrant DNA methylation of mTOR pathway genes promotes inflammatory activation of immune cells in diabetic kidney disease. Kidney Int. 2019, 96, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef]
- Badr, K.F.; Abi-Antoun, T.E. Isoprostanes and the kidney. Antioxid. Redox Signal. 2005, 7, 236–243. [Google Scholar] [CrossRef]
- Cardenas, H.; Vieth, E.; Lee, J.; Segar, M.; Liu, Y.; Nephew, K.P.; Matei, D. TGF-β induces global changes in DNA methylation during the epithelial-to-mesenchymal transition in ovarian cancer cells. Epigenetics 2014, 9, 1461–1472. [Google Scholar] [CrossRef]
- Lavoie, G.; Estève, P.-O.; Laulan, N.B.; Pradhan, S.; St-Pierre, Y. PKC isoforms interact with and phosphorylate DNMT1. BMC Biol. 2011, 9, 1–15. [Google Scholar] [CrossRef]
- Massagué, J.; Blain, S.W.; Lo, R.S. TGFβ signaling in growth control, cancer, and heritable disorders. Cell 2000, 103, 295–309. [Google Scholar] [CrossRef]
- Florl, A.R.; Löwer, R.; Schmitz-Dräger, B.; Schulz, W. DNA methylation and expression of LINE-1 and HERV-K provirus sequences in urothelial and renal cell carcinomas. Br. J. Cancer 1999, 80, 1312–1321. [Google Scholar] [CrossRef]
- Kelley, D.S.; Bartolini, G.L.; Newman, J.W.; Vemuri, M.; Mackey, B.E. Fatty acid composition of liver, adipose tissue, spleen, and heart of mice fed diets containing t10, c12-, and c9, t11-conjugated linoleic acid. Prostaglandins Leukot. Essent. Fatty Acids 2006, 74, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.; Hwang, J.-T.; Shin, E.J.; Hur, H.J.; Kang, K.; Choi, H.-K.; Chung, M.-Y.; Chung, S.; Sung, M.J.; Park, J.-H. Genome-wide analysis of DNA methylation identifies novel differentially methylated regions associated with lipid accumulation improved by ethanol extracts of Allium tubersosum and Capsella bursa-pastoris in a cell model. PLoS ONE 2019, 14, e0217877. [Google Scholar] [CrossRef] [PubMed]
- Lambert, M.-P.; Ancey, P.-B.; Esposti, D.D.; Cros, M.-P.; Sklias, A.; Scoazec, J.-Y.; Durantel, D.; Hernandez-Vargas, H.; Herceg, Z. Aberrant DNA methylation of imprinted loci in hepatocellular carcinoma and after in vitro exposure to common risk factors. Clin. Epigenetics 2015, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sheng, M.; McFadden, G.; Greenberg, M.E. Membrane depolarization and calcium induce c-fos transcription via phosphorylation of transcription factor CREB. Neuron 1990, 4, 571–582. [Google Scholar] [CrossRef]
- Choi, E.; Uyeno, S.; Nishida, N.; Okumoto, T.; Fujimura, S.; Aoki, Y.; Nata, M.; Sagisaka, K.; Fukuda, Y.; Nakao, K. Alterations of c-fos gene methylation in the processes of aging and tumorigenesis in human liver. Mutat. Res./Fundam. Mol. Mech. Mutagenesis 1996, 354, 123–128. [Google Scholar] [CrossRef]
- Rodríguez-Miguel, C.; Moral, R.; Escrich, R.; Vela, E.; Solanas, M.; Escrich, E. The role of dietary extra virgin olive oil and corn oil on the alteration of epigenetic patterns in the rat DMBA-induced breast cancer model. PLoS ONE 2015, 10, e0138980. [Google Scholar] [CrossRef]
- Barone, M.; Notarnicola, M.; Caruso, M.G.; Scavo, M.P.; Viggiani, M.T.; Tutino, V.; Polimeno, L.; Pesetti, B.; Di Leo, A.; Francavilla, A. Olive oil and omega-3 polyunsaturated fatty acids suppress intestinal polyp growth by modulating the apoptotic process in ApcMin/+ mice. Carcinogenesis 2014, 35, 1613–1619. [Google Scholar] [CrossRef]
- Menendez, J.A.; Joven, J.; Aragonès, G.; Barrajón-Catalán, E.; Beltrán-Debón, R.; Borrás-Linares, I.; Camps, J.; Corominas-Faja, B.; Cufí, S.; Fernández-Arroyo, S. Xenohormetic and anti-aging activity of secoiridoid polyphenols present in extra virgin olive oil: A new family of gerosuppressant agents. Cell Cycle 2013, 12, 555–578. [Google Scholar] [CrossRef]
- Zhao, Q.; Fan, Y.C.; Zhao, J.; Gao, S.; Zhao, Z.H.; Wang, K. DNA methylation patterns of peroxisome proliferator-activated receptor gamma gene associated with liver fibrosis and inflammation in chronic hepatitis B. J. Viral Hepat. 2013, 20, 430–437. [Google Scholar] [CrossRef]
- Jurado-Ruiz, E.; Álvarez-Amor, L.; Varela, L.M.; Berná, G.; Parra-Camacho, M.S.; Oliveras-Lopez, M.J.; Martínez-Force, E.; Rojas, A.; Hmadcha, A.; Soria, B. Extra virgin olive oil diet intervention improves insulin resistance and islet performance in diet-induced diabetes in mice. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Rowland, J.; Akbarov, A.; Eales, J.; Xu, X.; Dormer, J.P.; Guo, H.; Denniff, M.; Jiang, X.; Ranjzad, P.; Nazgiewicz, A. Uncovering genetic mechanisms of kidney aging through transcriptomics, genomics, and epigenomics. Kidney Int. 2019, 95, 624–635. [Google Scholar] [CrossRef] [PubMed]
- Teng, C.T. Lactoferrin gene expression and regulation: An overview. Biochem. and Cell Biol. 2002, 80, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.R.; Bryan, J.N.; Esebua, M.; Amos-Landgraf, J.; May, T.J. Testis specific Y-like 5: Gene expression, methylation and implications for drug sensitivity in prostate carcinoma. BMC Cancer 2017, 17, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Epping, M.T.; Meijer, L.A.; Krijgsman, O.; Bos, J.L.; Pandolfi, P.P.; Bernards, R. TSPYL5 suppresses p53 levels and function by physical interaction with USP7. Nat. Cell Biol. 2011, 13, 102–108. [Google Scholar] [CrossRef]
- Nasr, A.F.; Nutini, M.; Palombo, B.; Guerra, E.; Alberti, S. Mutations of TP53 induce loss of DNA methylation and amplification of the TROP1 gene. Oncogene 2003, 22, 1668–1677. [Google Scholar] [CrossRef]
- Tiwari, B.; Jones, A.E.; Caillet, C.J.; Das, S.; Royer, S.K.; Abrams, J.M. p53 directly represses human LINE1 transposons. Genes Dev. 2020, 34, 1439–1451. [Google Scholar] [CrossRef]
- Qiu, X.; Hu, B.; Huang, Y.; Deng, Y.; Wang, X.; Zheng, F. Hypermethylation of ACP1, BMP4, and TSPYL5 in hepatocellular carcinoma and their potential clinical significance. Dig. Dis. Sci. 2016, 61, 149–157. [Google Scholar] [CrossRef]
- Letchoumy, P.V.; Chandra Mohan, K.; Stegeman, J.; Gelboin, H.; Hara, Y.; Nagini, S. In vitro antioxidative potential of lactoferrin and black tea polyphenols and protective effects in vivo on carcinogen activation, DNA damage, proliferation, invasion, and angiogenesis during experimental oral carcinogenesis. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2008, 17, 193–203. [Google Scholar] [CrossRef]
- Ando, K.; Hasegawa, K.; Shindo, K.I.; Furusawa, T.; Fujino, T.; Kikugawa, K.; Nakano, H.; Takeuchi, O.; Akira, S.; Akiyama, T. Human lactoferrin activates NF-κB through the Toll-like receptor 4 pathway while it interferes with the lipopolysaccharide-stimulated TLR4 signaling. FEBS J. 2010, 277, 2051–2066. [Google Scholar] [CrossRef]
- Kouka, P.; Tekos, F.; Papoutsaki, Z.; Stathopoulos, P.; Halabalaki, M.; Tsantarliotou, M.; Zervos, I.; Nepka, C.; Liesivuori, J.; Rakitskii, V.N. Olive oil with high polyphenolic content induces both beneficial and harmful alterations on rat redox status depending on the tissue. Toxicol. Rep. 2020, 7, 421–432. [Google Scholar] [CrossRef]
- Sobinoff, A.P.; Mahony, M.; Nixon, B.; Roman, S.D.; McLaughlin, E.A. Understanding the Villain: DMBA-induced preantral ovotoxicity involves selective follicular destruction and primordial follicle activation through PI3K/Akt and mTOR signaling. Toxicol. Sci. 2011, 123, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Tikoo, K.; Kumar, P.; Gupta, J. Rosiglitazone synergizes anticancer activity of cisplatin and reduces its nephrotoxicity in 7, 12-dimethyl benz {a} anthracene (DMBA) induced breast cancer rats. BMC Cancer 2009, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.; Van Meter, M.; Ablaeva, J.; Ke, Z.; Gonzalez, R.S.; Taguchi, T.; De Cecco, M.; Leonova, K.I.; Kogan, V.; Helfand, S.L. LINE1 derepression in aged wild-type and SIRT6-deficient mice drives inflammation. Cell Metab. 2019, 29, 871–885.e5. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Lee, Y.D.; Wagers, A.J. Stem cell aging: Mechanisms, regulators and therapeutic opportunities. Nat. Med. 2014, 20, 870–880. [Google Scholar] [CrossRef]
- Fontana, L.; Partridge, L.; Longo, V.D. Extending healthy life span—from yeast to humans. Science 2010, 328, 321–326. [Google Scholar] [CrossRef]




| Name of the Group | ip. DMBA | Daily Dose/Animal | Manufacturer | Latin/Scientific Names |
|---|---|---|---|---|
| negative control | – | |||
| positive control | + | Sigma Aldrich Ltd. | dimethylbenz[a]anthracene | |
| EVOO | + | 0.3 g | Agraria Riva Del Garda SCA | Oleum virgineum |
| TFA | + | 0.3 g | Sigma Aldrich Ltd. | trans-3-hexadecenoic acid |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szabo, L.; Molnar, R.; Tomesz, A.; Deutsch, A.; Darago, R.; Varjas, T.; Ritter, Z.; Szentpeteri, J.L.; Andreidesz, K.; Mathe, D.; et al. Olive Oil Improves While Trans Fatty Acids Further Aggravate the Hypomethylation of LINE-1 Retrotransposon DNA in an Environmental Carcinogen Model. Nutrients 2022, 14, 908. https://doi.org/10.3390/nu14040908
Szabo L, Molnar R, Tomesz A, Deutsch A, Darago R, Varjas T, Ritter Z, Szentpeteri JL, Andreidesz K, Mathe D, et al. Olive Oil Improves While Trans Fatty Acids Further Aggravate the Hypomethylation of LINE-1 Retrotransposon DNA in an Environmental Carcinogen Model. Nutrients. 2022; 14(4):908. https://doi.org/10.3390/nu14040908
Chicago/Turabian StyleSzabo, Laszlo, Richard Molnar, Andras Tomesz, Arpad Deutsch, Richard Darago, Timea Varjas, Zsombor Ritter, Jozsef L. Szentpeteri, Kitti Andreidesz, Domokos Mathe, and et al. 2022. "Olive Oil Improves While Trans Fatty Acids Further Aggravate the Hypomethylation of LINE-1 Retrotransposon DNA in an Environmental Carcinogen Model" Nutrients 14, no. 4: 908. https://doi.org/10.3390/nu14040908
APA StyleSzabo, L., Molnar, R., Tomesz, A., Deutsch, A., Darago, R., Varjas, T., Ritter, Z., Szentpeteri, J. L., Andreidesz, K., Mathe, D., Hegedüs, I., Sik, A., Budan, F., & Kiss, I. (2022). Olive Oil Improves While Trans Fatty Acids Further Aggravate the Hypomethylation of LINE-1 Retrotransposon DNA in an Environmental Carcinogen Model. Nutrients, 14(4), 908. https://doi.org/10.3390/nu14040908