

Protective Effects of Naringenin from Citrus sinensis (var. Valencia) Peels against CCl4-Induced Hepatic and Renal Injuries in Rats Assessed by Metabolomics, Histological and Biochemical Analyses

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Extraction and Purification of Naringenin
2.2. Chemicals and Drugs
2.3. Pharmacological Study
2.3.1. Animals
2.3.2. Induction of Acute Liver Toxicity
2.3.3. Biochemical Assays
2.3.4. Histopathologic Investigation
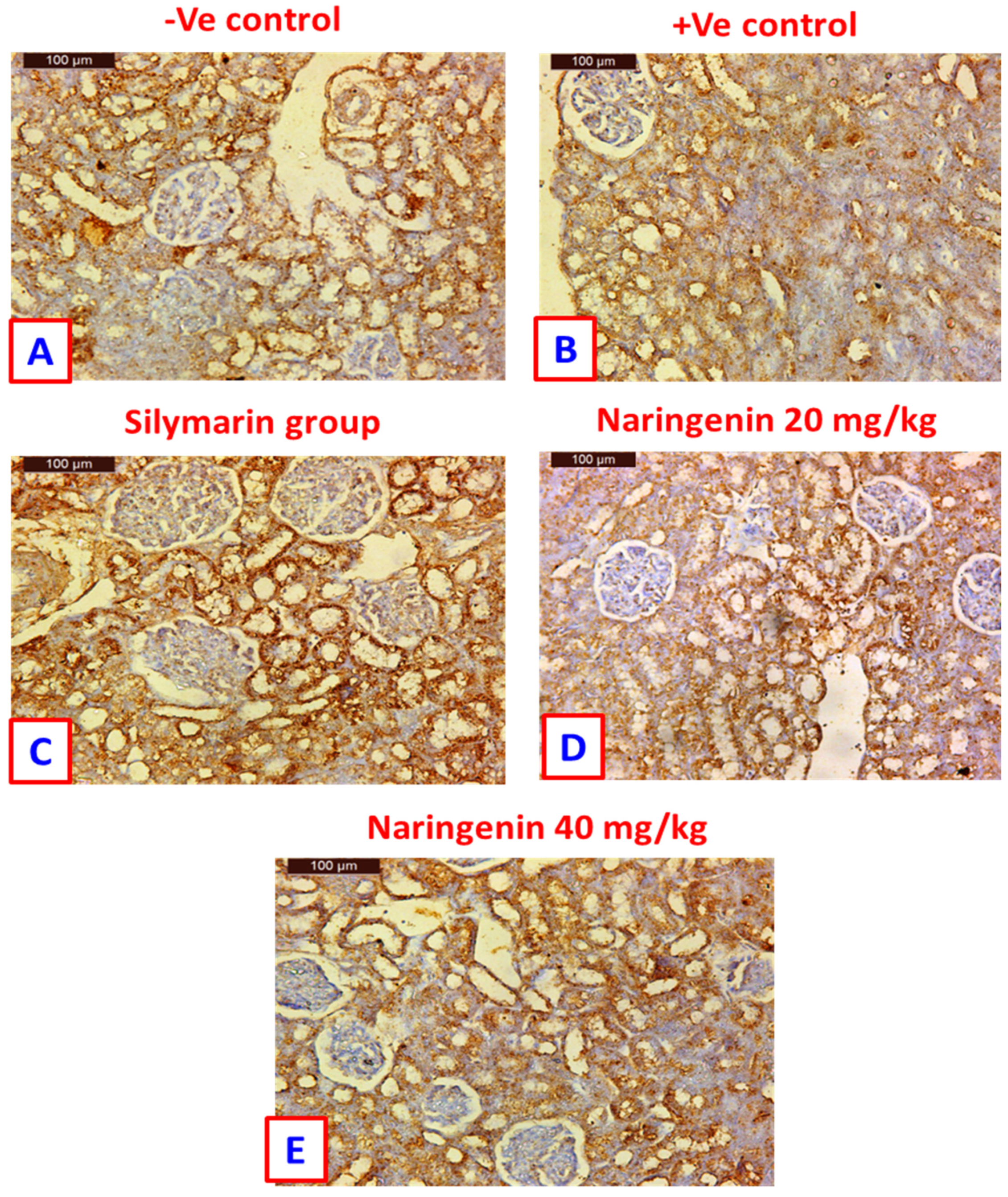
2.3.5. Immuno-Histochemical Assessment of Bcl-2 Expression in Kidneys and Liver
2.3.6. Preparation of Metabolomics Samples and GC–MS Analysis
2.3.7. GC–MS Analysis
2.3.8. Processing of GC–MS Data, Molecular Networking and Multivariate Data Analyses
2.3.9. Statistical Analysis
3. Results
3.1. Effects on Liver Enzymes
3.2. Effects on Kidney-Related Parameters
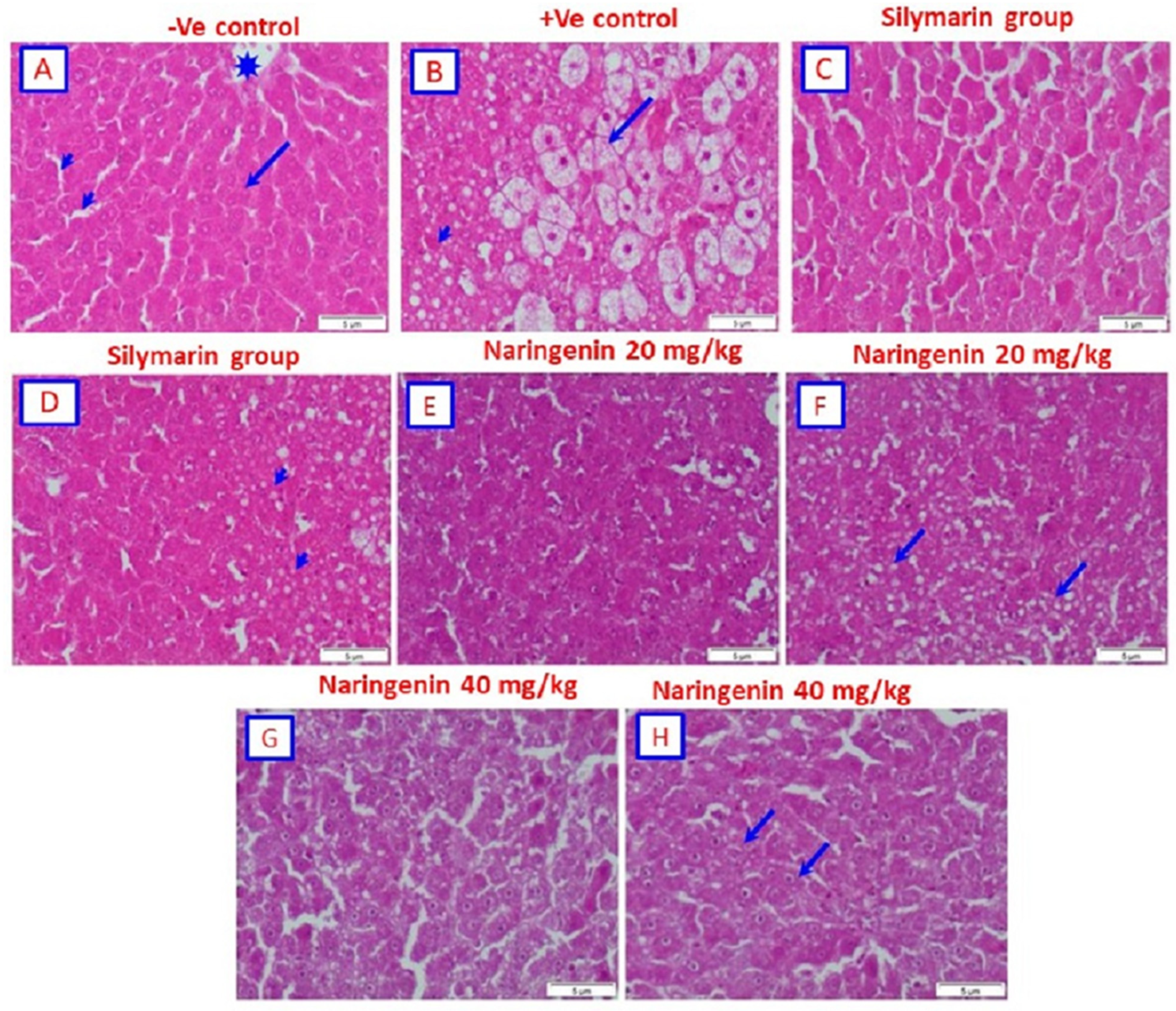
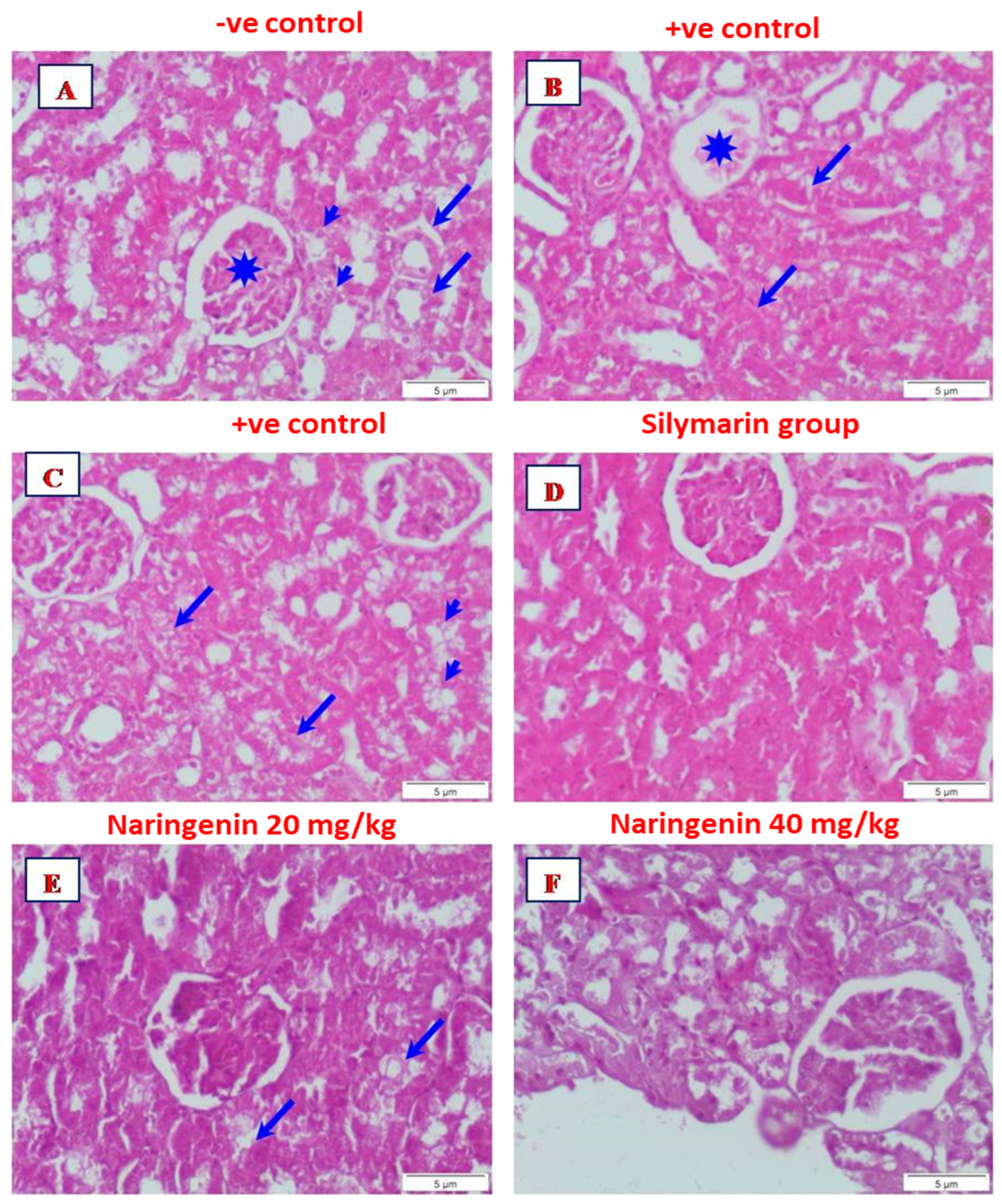
3.3. Histopathological Examination
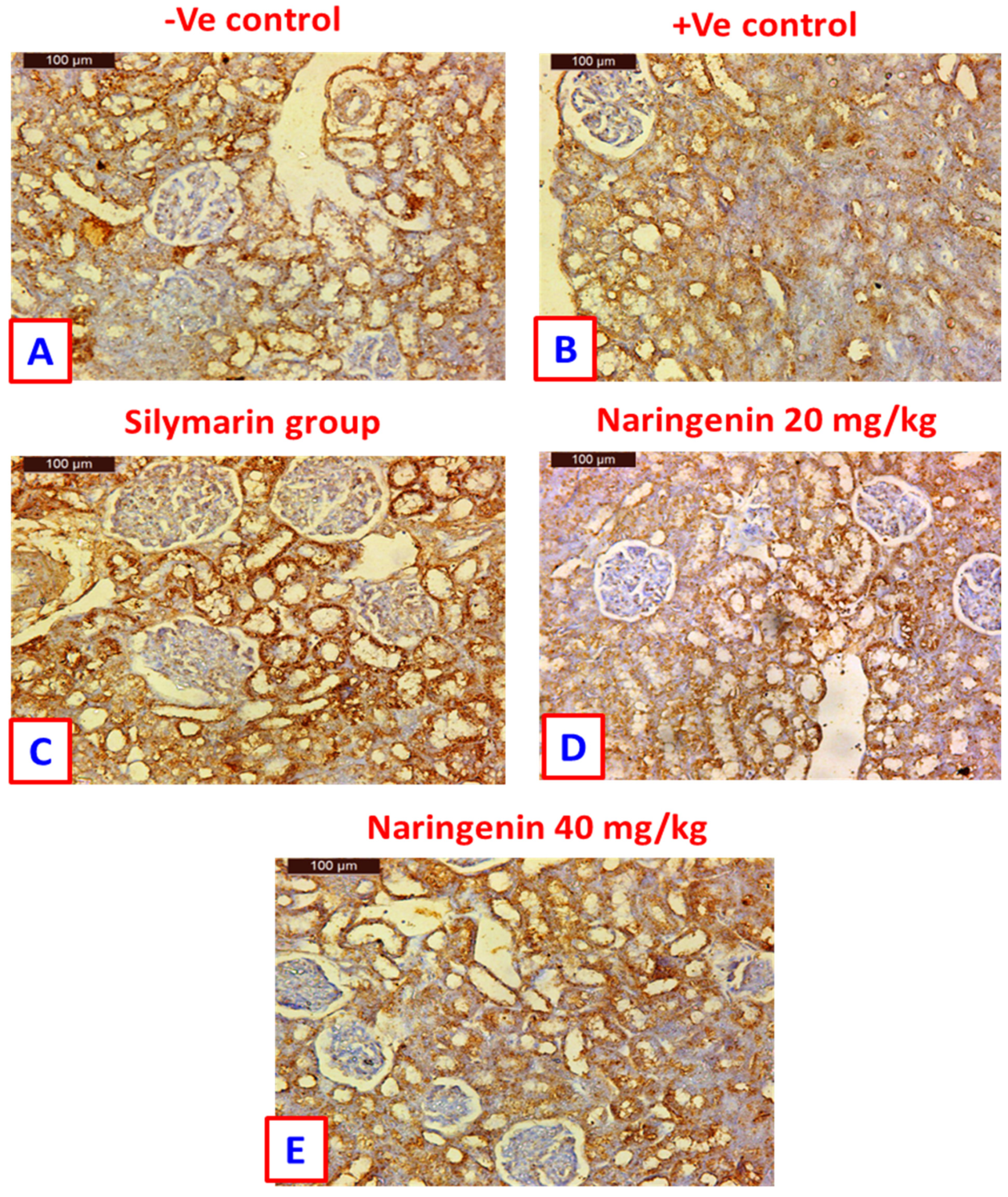
3.4. Bcl-2 Immuno-Histochemical Expression
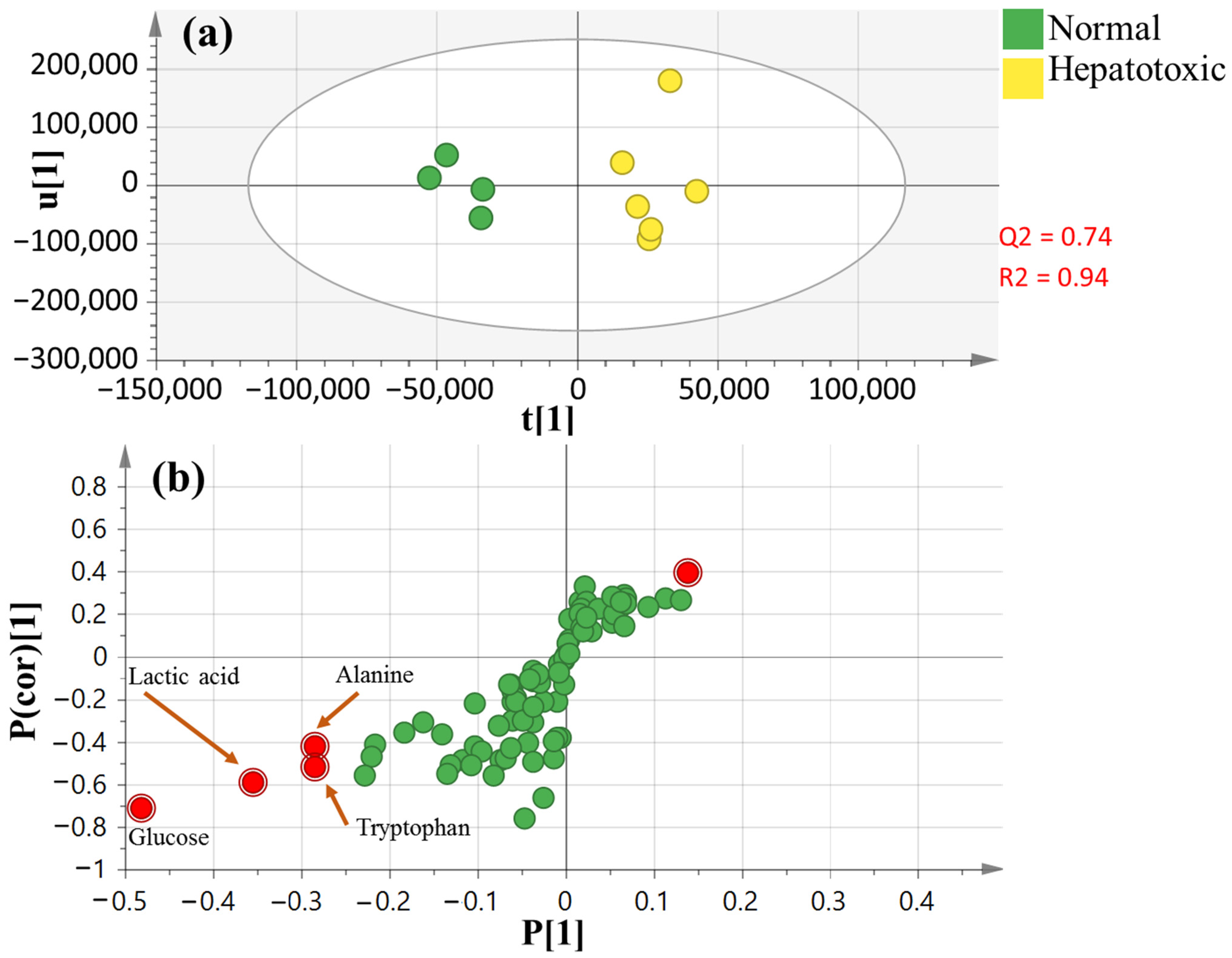
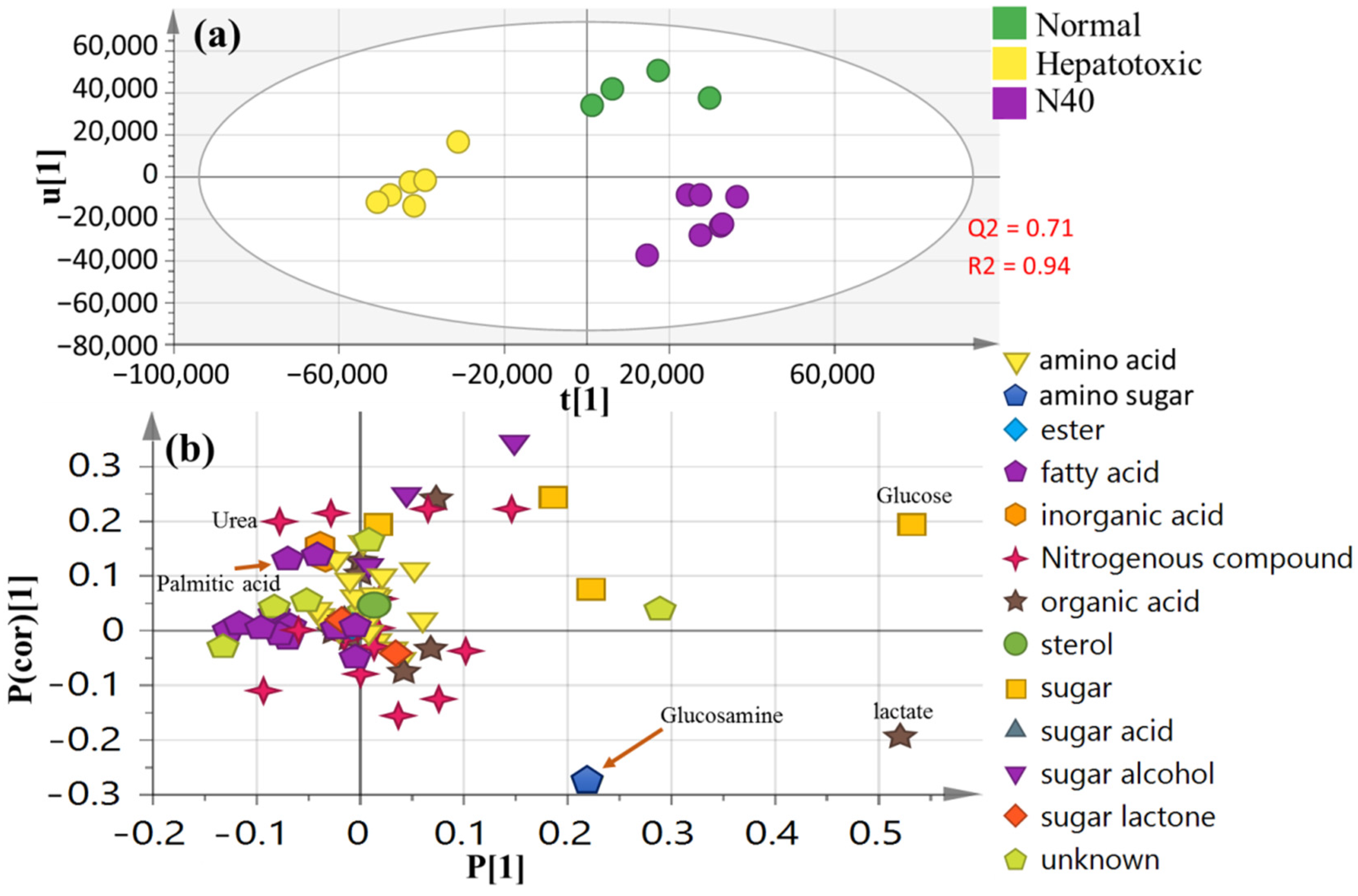
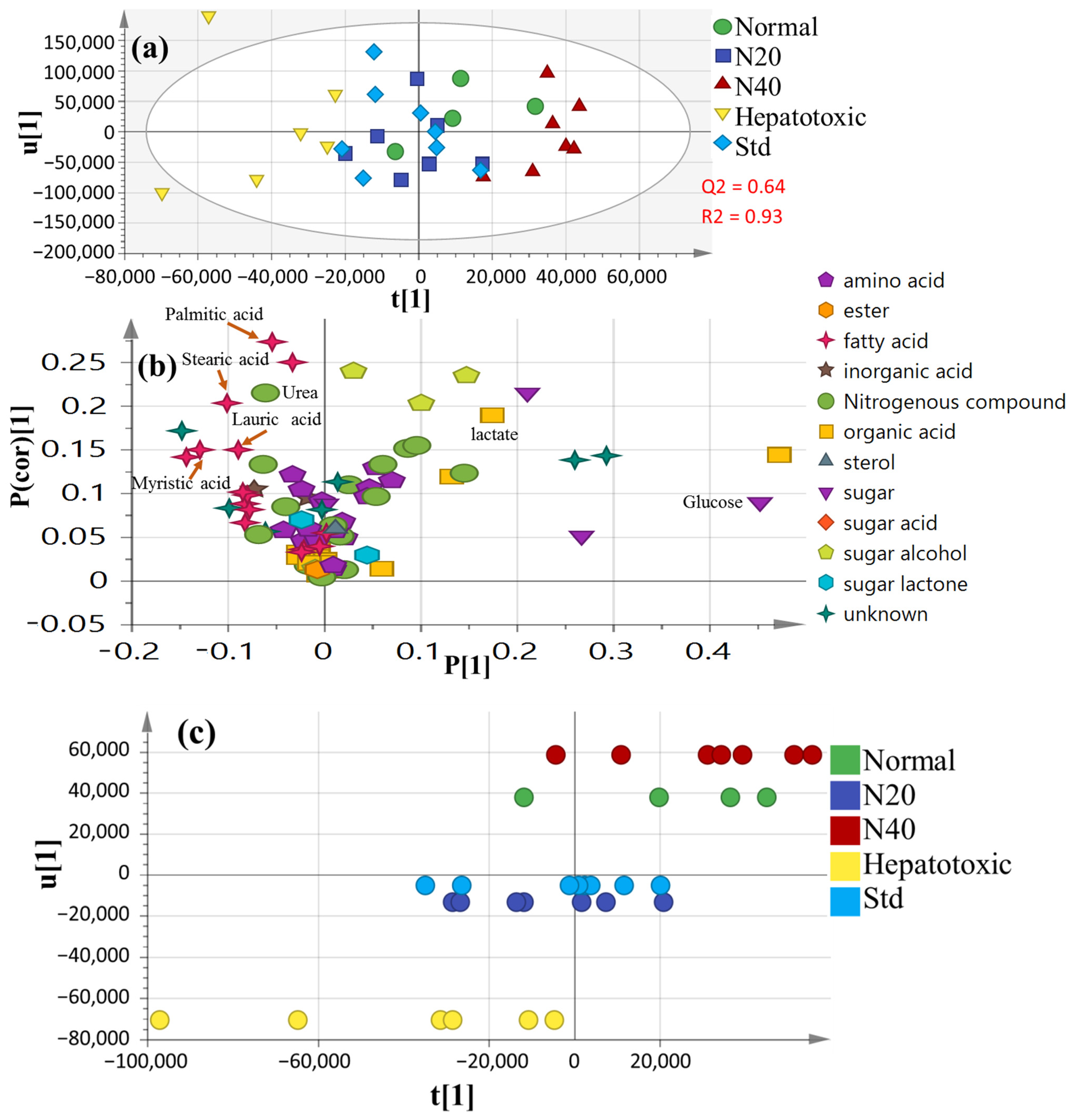
3.5. Serum Metabolite Profiling in CCl4-Induced Rat Livers and with Treatments as Analyzed Using GC–MS
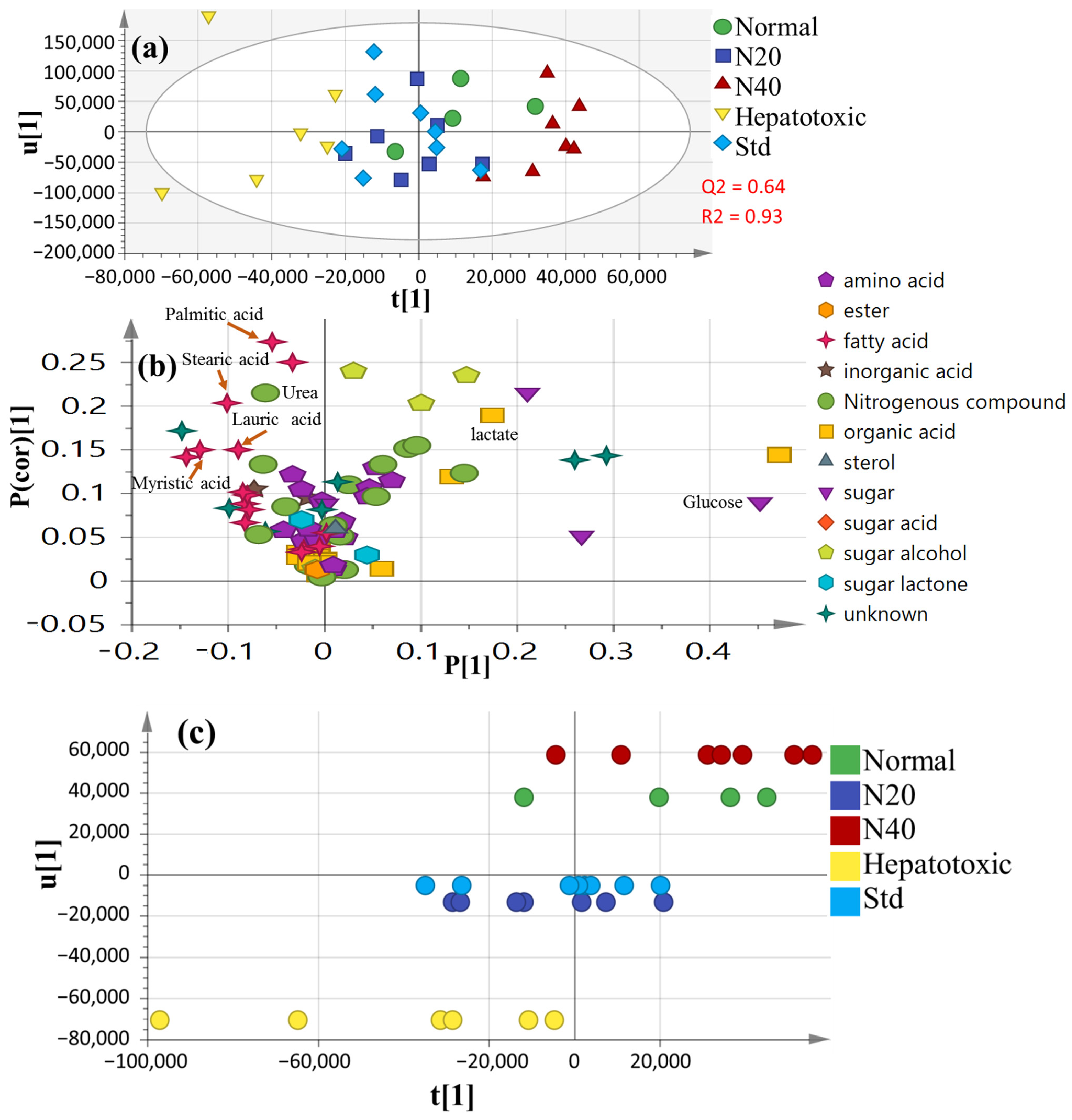
3.6. Multivariate Analysis of Collected Datasets from All Treatment Groups
3.7. OPLS-DA Analysis of Collected Dataset from CCl4-Induced Liver Injury versus Normal Control
3.8. OPLS-DA Analysis of Collected Dataset from Normal Control, CCl4-Intoxicated and Naringenin (40 mg/kg) Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gao, B.; Jeong, W.I.; Tian, Z. Liver: An organ with predominant innate immunity. Hepatology 2008, 47, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Al-Seeni, M.N.; El Rabey, H.A.; Zamzami, M.A.; Alnefayee, A.M. The hepatoprotective activity of olive oil and Nigella sativa oil against CCl4 induced hepatotoxicity in male rats. BMC Complem. Altern. 2016, 16, 438. [Google Scholar] [CrossRef] [PubMed]
- Asrani, S.K.; Devarbhavi, H.; Eaton, J.; Kamath, P.S. Burden of liver diseases in the world. J. Hepatol. 2019, 70, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Aquino, E.; Muriel, P. Benefcial effects of naringenin in liver diseases: Molecular mechanisms. World J. Gastroenterol. 2018, 24, 1679–1707. [Google Scholar] [CrossRef]
- Han, C.Y.; Sun, T.; Xv, G.P.; Wang, S.; Gu, J.G.; Liu, C.Y. Berberine ameliorates CCl4-induced liver injury in rats through regulation of the Nrf2-Keap1-ARE and p53 signaling pathways. Mol. Med. Rep. 2019, 20, 3095–3102. [Google Scholar] [CrossRef]
- Manibusan, M.K.; Odin, M.; Eastmond, D.A. Postulated carbon tetrachloride mode of action: A review. J. Environ. Sci. Health Part C 2007, 25, 185–209. [Google Scholar] [CrossRef]
- Navarro, V.J.; Khan, I.; Björnsson, E.; Seeff, L.B.; Serrano, J.; Hoofnagle, J.H. Liver injury from herbal and dietary supplements. Hepatology 2017, 65, 363–373. [Google Scholar] [CrossRef]
- Wang, Q.; Ou, Y.; Hu, G.; Wen, C.; Yue, S.; Chen, C.; Xu, L.; Xie, J.; Dai, H.; Xiao, H.; et al. Naringenin attenuates non-alcoholic fatty liver disease by down-regulating the NLRP3/NF-κB pathway in mice. Br. J. Pharmacol. 2020, 177, 1806–1821. [Google Scholar] [CrossRef]
- El-Kersh, D.M.; Ezzat, S.M.; Salama, M.M.; Salama, M.; Mahrous, E.; Attia, Y.; Ahmed, M.; Elmazar, M. Anti-estrogenic and anti-aromatase activities of citrus peels major compounds in breast cancer. Sci. Rep. 2021, 11, 7121. [Google Scholar] [CrossRef]
- Citrus-Market. 2017. Available online: https://farmfolio.net/articles/agricultural-citrus-markets-egypt/ (accessed on 17 May 2017).
- Nair, A.; Kurup, S.R.; Nair, A.S.; Baby, S. Citrus peels prevent cancer. Phytomedicine 2018, 50, 231–237. [Google Scholar] [CrossRef]
- Rawson, N.E.; Ho, C.-T.; Li, S. Efficacious anti-cancer property of flavonoids from citrus peels. Food Sci. Hum. Wellness 2014, 3, 104–109. [Google Scholar] [CrossRef]
- Salehi, B.; Fokou, P.V.T.; Sharifi-Rad, M.; Zucca, P.; Pezzani, R.; Martins, N.; Sharifi-Rad, J. The therapeutic potential of naringenin: A review of clinical trials. Pharmaceuticals 2019, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted metabolomics strategies-challenges and emerging directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, N.; Taleb, M.; Heiss, A.; Kropf, M.; Farag, M. GC-MS based metabolites profiling of nutrients and anti-nutrients in 10 Lathyrus seed genotypes: A prospect for phyto-equivalency and chemotaxonomy. Food Biosci. 2021, 42, 101183. [Google Scholar] [CrossRef]
- Elshamy, A.I.; Abdallah, H.M.I.; El Gendy, A.E.G.; El-Kashak, W.; Muscatello, B.; De Leo, M.; Pistelli, L. Evaluation of anti-inflammatory, antinociceptive, and antipyretic activities of Prunus persica var. nucipersica (Nectarine) Kernel. Plant. Med. 2019, 85, 1016–1023. [Google Scholar] [CrossRef]
- Nassar, M.I.; Mohamed, T.K.; Elshamy, A.I.; El-Toumy, S.A.; Abdel Lateef, A.M.; Farrag, A.R. Chemical constituents and anti-ulcerogenic potential of the scales of Cynara scolymus (artichoke) heads. J. Sci. Food Agric. 2013, 93, 2494–2501. [Google Scholar] [CrossRef]
- Esmaeili, M.A.; Alilou, M. Naringenin attenuates CCl4-induced hepatic inflammation by the activation of an Nrf2-mediated pathway in rats. Clin. Exp. Pharmacol. Physiol. 2014, 41, 416–422. [Google Scholar] [CrossRef]
- Ustuner, D.; Kolac, U.; Ustuner, M.; Tanrikut, C.; Koroglu, Z.; Donmez, D.; Ozen, H.; Ozden, H. Naringenin Ameliorate Carbon Tetrachloride-Induced Hepatic Damage Through Inhibition of Endoplasmic Reticulum Stress and Autophagy in Rats. J. Med. Food 2020, 23, 1192–1200. [Google Scholar] [CrossRef]
- Dong, D.; Xu, L.; Yin, L.; Qi, Y.; Peng, J. Naringin prevents carbon tetrachloride-induced acute liver injury in mice. J. Funct. Foods 2015, 12, 179–191. [Google Scholar] [CrossRef]
- Lin, A.X.; Chan, G.; Hu, Y.; Ouyang, D.; Ung, C.O.L.; Shi, L.; Hu, H. Internationalization of traditional Chinese medicine: Current international market, internationalization challenges and prospective suggestions. J. Chin. Med. 2018, 13, 1–6. [Google Scholar] [CrossRef]
- Dhuley, J.N.; Naik, S.R. Proective effect of Rhinax, a herbal formulation against CCl4-induced liver injury and survival in rats. J. Ethnopharmacol. 1997, 56, 59–164. [Google Scholar] [CrossRef]
- Adil, M.; Kandhare, A.D.; Ghosh, P.; Venkata, S.; Raygude, K.S.; Bodhankar, S.L. Ameliorative effect of naringenin in acetaminophen induced hepatic and renal toxicity in laboratory rats: Role of FXR and KIM-1. Ren. Fail. 2016, 38, 1007–1020. [Google Scholar] [CrossRef] [PubMed]
- Elsawy, H.; Algefare, A.I.; Alfwuaires, M.; Khalil, M.; Elmenshawy, O.M.; Sedky, A.; Abdel-Moneim, A.M. Naringenin alleviates methotrexate-induced liver injury in male albino rats and enhances its antitumor efficacy in HepG2 cells. Biosci. Rep. 2020, 40, BSR20193686. [Google Scholar] [CrossRef] [PubMed]
- Ghobadi, M.; Mirazi, N.; Alaei, H.; Moradkhani, S.; Rajaei, Z.; Monsef, A. Effects of lactulose and silymarin on liver enzymes in cirrhotic rats. Can. J. Physiol. Pharmacol. 2017, 95, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Ozen, O.A.; Kus, M.A.; Kus, I.; Alkoc, O.A.; Songur, A. Protective effects of melatonin against formaldehyde-induced oxidative damage and apoptosis in rat testes: An immunohistochemical and biochemical study. Syst. Biol. Reprod. Med. 2008, 54, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Han, Y.; Yang, H.; Li, S. Effects of SSTF on the expression of apoptosis-associated gene Bcl-2 and Bax by cardiomyocytes induced by H2O2. Guangdong Med. J. 2005, 28, 1590–1591. [Google Scholar]
- Ammar, N.M.; Hassan, H.A.; Mohammed, M.A.; Serag, A.; Abd El-Alim, S.H.; Elmotasem, H.; El Raey, M.; El Gendy, A.N.; Sobeh, M.; Abdel-Hamid, A.H. Metabolomic profiling to reveal the therapeutic potency of Posidonia oceanica nanoparticles in diabetic rats. RSC Adv. 2021, 11, 8398–8410. [Google Scholar] [CrossRef]
- Farag, M.A.; Khattab, A.R.; Shamma, S.; Afifi, S.M. Profiling of primary metabolites and volatile determinants in mahlab cherry (Prunus mahaleb L.) seeds in the context of its different varieties and roasting as analyzed using chemometric tools. Foods 2021, 10, 728. [Google Scholar] [CrossRef]
- Afifi, S.M.; El-Mahis, A.; Heiss, A.G.; Farag, M.A. Gas chromatography–mass spectrometry-based classification of 12 fennel (Foeniculum vulgare Miller) varieties based on their aroma profiles and estragole levels as analyzed using chemometric tools. ACS Omega 2021, 6, 5775–5785. [Google Scholar] [CrossRef]
- Rudnicki, M.; Silveira, M.; Pereira, T.V.; Oliveira, M.R.; Reginatto, F.H.; Dal-Pizzol, F.; Moreira, J.C. Protective effects of Passiflora alata extract pretreatment on carbon tetrachloride induced oxidative damage in rats. Food Chem. Toxicol. 2007, 45, 656–661. [Google Scholar] [CrossRef]
- Desai, S.N.; Patel, D.K.; Devkar, R.V.; Patel, P.V.; Ramachandran, A. Hepatoprotective potential of polyphenol rich extract of Murraya koenigii L.: An in vivo study. Food Chem. Toxicol. 2012, 50, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Ramaiah, S.K. A toxicologist guide to the diagnostic interpretation ofhepatic biochemical parameters. Food Chem. Toxicol. 2007, 45, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Panahi Kokhdan, E.; Ahmadi, K.; Sadeghi, H.; Sadeghi, H.; Dadgary, F.; Danaei, N.; Aghamaali, M.R. Hepatoprotective effect of Stachys pilifera ethanol extract in carbon tetrachloride-induce hepatotoxicity in rats. Pharm. Biol. 2017, 55, 1389–1393. [Google Scholar] [CrossRef] [PubMed]
- Ullah, H.; Khan, A.; Baig, M.W.; Ullah, N.; Ahmed, N.; Tipu, M.; Hussain Ali, H.; Salman Khan, S. Poncirin attenuates CCL4-induced liver injury through inhibition of oxidative stress and inflammatory cytokines in mice. BMC Complement. Med. Ther. 2020, 20, 115. [Google Scholar] [CrossRef]
- Makni, M.; Chtourou, Y.; Garoui, E.M.; Boudawara, T.; Fetoui, H. Carbon tetrachloride-induced nephrotoxicity and DNA damage in rats: Protective role of vanillin. Hum. Exp. Toxicol. 2012, 31, 844–852. [Google Scholar] [CrossRef]
- Jaramillo-Juárez, F.; Rodríguez-Vázquez, M.L.; Rincón-Sánchez, A.R.; Consolación Martínez, M.; Ortiz, G.; Llamas, J.; Anibal Posadas, F.; Reyes, J.L. Acute renal failure induced by carbon tetrachloride in rats with hepatic cirrhosis. Ann. Hepatol. 2008, 7, 331–338. [Google Scholar] [CrossRef]
- Radulović, N.S.; Randjelović, P.J.; Stojanović, N.M.; Ilić, I.R.; Miltojević, A.B.; Stojković, M.B.; Ilić, M. Effect of two esters of N-methylanthranilic acid from Rutaceae species on impaired kidney morphology and function in rats caused by CCl4. Life Sci. 2015, 135, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Olagunjua, J.; Adeneyeb, A.; Fagbohunkac, B.; Bisugac, N.; Ketikuc, A.; Benebod, A.; Olufowobic, O.; Adeoyec, A.; Alimic, M.; Adelekec, A. Nephroprotective activities of the aqueous seed extract of Carica papaya Linn. in carbon tetrachloride induced renal injured Wistar rats: A dose- and time-dependent study. Biol. Med. 2009, 2009, 11–19. [Google Scholar]
- Yilmaz-Ozden, T.; Can, A.; Karatug, A.; Pala-Kara, Z.; Okyar, A.; Bolkent, S. Carbon tetrachloride-induced kidney damage and protective effect of Amaranthus lividus L. in rats. Toxicol. Ind. Health 2016, 32, 1143–1152. [Google Scholar] [CrossRef]
- Baker, A.J.; Mooney, A.; Hughes, J.; Lombardi, D.; Johnson, R.; Savill, J. Mesangial cell apoptosis: The major mechanism for resolution of glomerular hypercellularity in experimental mesangial proliferative nephritis. J. Clin. Investig. 1994, 94, 2105–2116. [Google Scholar] [CrossRef]
- Yang, B.; Johnson, T.S.; Thomas, G.L.; Watson, P.F.; Wagner, B.; Furness, P.N.; El Nahas, A.M. A shift in the Bax/Bcl-2 balance may activate caspase-3 and modulate apoptosis in experimental glomerulonephritis. Kidney Int. 2002, 62, 1301–1313. [Google Scholar] [CrossRef] [PubMed]
- Taub, R. Liver regeneration: From myth to mechanism. Nat. Rev. Mol. Cell Biol. 2004, 5, 836–847. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wen, P.H.; Zhang, X.; Dai, Y.; He, Q. Breviscapine ameliorates CCl4-induced liver injury in mice through inhibiting inflammatory apoptotic response and ROS generation. Int. J. Mol. Med. 2018, 42, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Safhi, M.M. Nephroprotective Effect of zingerone against CCl4-Induced renal toxicity in swiss albino mice: Molecular mechanism. Oxidative Med. Cell. Longev. 2018, 2018, 2474831. [Google Scholar] [CrossRef]
- Chung, T.W.; Li, S.; Lin, C.C.; Tsai, S.W. Antinociceptive and anti-inflammatory effects of the citrus flavanone naringenin. Tzu-Chi Med. J. 2019, 31, 81–85. [Google Scholar] [CrossRef]
- Jin, C.Y.; Park, C.; Lee, J.H.; Chung, K.T.; Kwon, T.K.; Kim, G.Y.; Choi, B.T.; Choi, Y.H. Naringenin-induced apoptosis is attenuated by Bcl-2 but restored by the small molecule Bcl-2 inhibitor, HA 14-1, in human leukemia U937 cells. Toxicol. Vitr. 2009, 23, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Manchope, M.F.; Calixto-Campos, C.; Coelho-Silva, L.; Zarpelon, A.C.; Pinho-Ribeiro, F.A.; Georgetti, S.R. Naringenin Inhibits Superoxide Anion-Induced Inflammatory Pain: Role of Oxidative Stress, Cytokines, Nrf-2 and the NO−cGMP−PKG−KATP Channel Signaling Pathway. PLoS ONE 2016, 11, e0153015. [Google Scholar] [CrossRef]
- Zeng, L.; Zhen, Y.; Chen, Y.; Zou, L.; Zhang, Y.; Hu, F.; Feng, J.; Shen, J.; Wei, B. Naringin inhibits growth and induces apoptosis by a mechanism dependent on reduced activation of NF-κB/COX-2-caspase-1 pathway in HeLa cervical cancer cells. Int. J. Oncol. 2014, 45, 1929–1936. [Google Scholar] [CrossRef]
- Ahamad, M.S.; Siddiqui, S.; Jafri, A.; Ahmad, S.; Afzal, M.; Arshad, M. Induction of Apoptosis and Antiproliferative Activity of Naringenin in Human Epidermoid Carcinoma Cell through ROS Generation and Cell Cycle Arrest. PLoS ONE 2014, 9, e110003. [Google Scholar] [CrossRef]
- Park, J.H.; Jin, C.Y.; Lee, B.K.; Kim, G.Y.; Choi, Y.H.; Jeong, Y.K. Naringenin induces apoptosis through downregulation of Akt and caspase-3 activation in human leukemia THP-1 cells. Food Chem. Toxicol. 2008, 46, 3684–3690. [Google Scholar] [CrossRef]
- Wali, A.F.; Rashid, S.; Rashid, S.M.; Ansari, M.A.; Khan, M.R.; Haq, N.; Alhareth, D.Y.; Ahmad, A.; Rehman, M.U. Naringenin regulates doxorubicin-induced liver dysfunction: Impact on oxidative stress and inflammation. Plants 2020, 9, 550. [Google Scholar] [CrossRef] [PubMed]
- El-Desoky, A.H.; Abdel-Rahman, R.F.; Ahmed, O.K.; El-Beltagi, H.S.; Hattori, M. Anti-inflammatory and antioxidant activities of naringin isolated from Carissa carandas L.: In vitro and in vivo evidence. Phytomedicine 2018, 42, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hu, C.; Li, H.; Wu, L.; Xiong, Y.; Tang, X.; Deng, S. Metabolic profiling of fatty acids in Tripterygium wilfordii multiglucoside- and triptolide-induced liver-injured rats. Open Life Sci. 2021, 6, 184–197. [Google Scholar] [CrossRef] [PubMed]
- Mota, M.; Banini, B.A.; Cazanave, S.C.; Sanyal, A.J. Molecular mechanisms of lipotoxicity and glucotoxicity in nonalcoholic fatty liver disease. Metabolism 2016, 65, 1049–1061. [Google Scholar] [CrossRef] [PubMed]
- Abaffy, T.; Möller, M.G.; Riemer, D.; Milikowski, C.; DeFazio, R.A. Comparative analysis of volatile metabolomics signals from melanoma and benign skin: A pilot study. Metabolomics 2013, 9, 998–1008. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.D.; Cardin, S.; Edgerton, D.; Cherrington, A. Splanchnic free fatty acid kinetics. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E1140. [Google Scholar] [CrossRef]
- Ke, X.-H.; Wang, C.-G.; Luo, W.-Z.; Wang, J.; Li, B.; Lv, J.-P.; Dong, R.-J.; Ge, D.-Y.; Han, Y.; Yang, Y.-J.; et al. Metabolomic study to determine the mechanism underlying the effects of Sagittaria sagittifolia polysaccharide on isoniazid- and rifampicin-induced hepatotoxicity in mice. Molecules 2018, 23, 3087. [Google Scholar] [CrossRef]
- Angulo, P.; Lindor, K.D. Non-alcoholic fatty liver disease. J. Gastroenterol. Hepatol. 2002, 17, S186–S190. [Google Scholar] [CrossRef]
- Pessayre, D.; Berson, A.; Fromenty, B.; Mansouri, A. Mitochondria in steatohepatitis. Semin. Liver Dis. 2001, 21, 57–69. [Google Scholar] [CrossRef]
- Moravcová, A.; Červinková, Z.; Kučera, O.; Mezera, V.; Rychtrmoc, D.; Lotková, H. The effect of oleic and palmitic acid on induction of steatosis and cytotoxicity on rat hepatocytes in primary culture. Physiol. Res. 2015, 5, S627. [Google Scholar] [CrossRef] [PubMed]
- Puri, P.; Wiest, M.M.; Cheung, O.; Mirshahi, F.; Sargeant, C.; Min, H.K.; Contos, M.J.; Sterling, R.K.; Fuchs, M.; Zhou, H.; et al. The plasma lipidomic signature of nonalcoholic steatohepatitis. Hepatology 2009, 50, 1827–1838. [Google Scholar] [CrossRef] [PubMed]
- Speziali, G.; Liesinger, L.; Gindlhuber, J.; Leopold, C.; Pucher, B.; Brandi, J.; Castagna, A.; Tomin, T.; Krenn, P.; Thallinger, G.; et al. Myristic acid induces proteomic and secretomic changes associated with steatosis, cytoskeleton remodeling, endoplasmic reticulum stress, protein turnover and exosome release in HepG2 cells. J Proteom. 2018, 181, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Ebbesson, S.O.; Voruganti, V.S.; Higgins, P.B.; Fabsitz, R.; Ebbesson, L.O.; Laston, S.; Harris, W.; Kennish, J.; Umans, B.; Wang, H.; et al. Fatty acids linked to cardiovascular mortality are associated with risk factors. Int. J. Circumpolar Health 2015, 74, 28055. [Google Scholar] [CrossRef] [PubMed]
- Waters, N.J.; Waterfield, C.J.; Farrant, R.D.; Holmes, E.; Nicholson, J.K. Metabonomic deconvolution of embedded toxicity: Application to thioacetamide hepato- and nephrotoxicity. Chem. Res. Toxicol. 2005, 18, 639–654. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.T.; Li, P.; Wang, J.S.; Zhang, Q.; Xu, D.; Yang, M.; Kong, L. Protection of baicalin against lipopolysaccharide induced liver and kidney injuries based on 1H NMR metabolomic profiling. Toxicol. Res. 2016, 5, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Panasyuk, G.; Espeillac, C.; Chauvin, C.; Pradelli, L.A.; Horie, Y.; Suzuki, A.; Annicotte, J.S.; Fajas, L.; Foretz, M.; Verdeguer, F.; et al. PPARγ contributes to PKM2 and HK2 expression in fatty liver. Nat. Commun. 2012, 14, 672. [Google Scholar] [CrossRef]
- Lu, X.; Zhao, Q.; Tian, Y.; Xiao, S.; Jin, T.; Fan, X. A metabonomic characterization of (+)-usnic acid-induced liver injury by gas chromatography-mass spectrometry-based metabolic profiling of the plasma and liver in rat. Int. J. Toxicol. 2011, 30, 478–491. [Google Scholar] [CrossRef]
- Mu, L.; Hu, G.; Liu, J.; Chen, Y.; Cui, W.; Qiao, L. Protective effects of naringenin in a rat model of sepsis-triggered acute kidney injury via activation of antioxidant enzymes and reduction in urinary angiotensinogen. Med. Sci. Monit. 2019, 25, 5986–5991. [Google Scholar] [CrossRef]
- Akang, E.N.; Dosumu, O.; Okoko, I.E.; Faniyan, O.; Oremosu, A.; Akanmu, A.S. Microscopic and biochemical changes on liver and kidney of Wistar rats on combination antiretroviral therapy: The impact of naringenin and quercetin. Toxicol. Res. 2020, 9, 601–608. [Google Scholar] [CrossRef]
- Gnanasoundari, M.; Pari, L. Impact of naringenin on oxytetracycline-mediated oxidative damage in kidney of rats. Ren. Fail. 2006, 28, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Wei, D.; Wang, J.; Dong, G.; Zhang, Q.; Yang, M.; Kong, L. Chronic toxicity of crude ricinine in rats assessed by 1H NMR metabolomics analysis. RSC Adv. 2015, 5, 27018–27028. [Google Scholar] [CrossRef]
- Xia, D.; Lai, X.; Wu, K.; Zhou, P.; Li, L.; Guo, Z.; Xu, S. Metabolomics study of fasudil on cisplatin-induced kidney injury. Biosci. Rep. 2019, 39, BSR20192940. [Google Scholar] [CrossRef]
- Li, Z.; Li, Y.; Lu, L.; Yang, Z.; Xue, W.; Tian, X.; Zhang, X. 1H-NMR based serum metabolomics study to investigate hepatoprotective effect of qin-jiao on carbon tetrachloride-induced acute hepatotoxicity in rats. Evid. -Based Complement. Altern. Med. 2017, 2017, 6091589. [Google Scholar] [CrossRef]
- Zira, A.; Kostidis, S.; Theocharis, S.; Sigala, F.; Engelsen, S.B.; Andreadou, I.; Mikros, E. 1H NMR-based metabonomics approach in a rat model of acute liver injury and regeneration induced by CCl4 administration. Toxicology 2013, 303, 115–124. [Google Scholar] [CrossRef]
- Gao, J.; Qin, X.J.; Jiang, H.; Jiang, H.; Chen, J.F.; Wang, T.; Zhang, T.; Xu, S.Z.; Song, J.M. Detecting serum and urine metabolic profile changes of CCl4-liver fibrosis in rats at 12 weeks based on gas chromatography-mass spectrometry. Exp. Ther. Med. 2017, 14, 1496–1504. [Google Scholar] [CrossRef]
- Manco, M.; Nobili, V. Beta-cell glucose sensitivity in patients with liver fibrosis. Gut 2008, 57, 1023. [Google Scholar]
- Zant, R.; Melter, M.; Beck, D.; Ameres, M.; Knoppke, B.; Kunkel, J. Glucose metabolism and associated outcome after pediatric liver transplantation. Transplant. Proc. 2016, 48, 2709–2713. [Google Scholar] [CrossRef]
- Campo, G.M.; Avenoso, A.; Campo, S.; D’Ascola, A.; Ferlazzo, A.M.; Calatroni, A. The antioxidant and antifibrogenic effects of the glycosaminoglycans hyaluronic acid and chondroitin-4-sulphate in a subchronic rat model of carbon tetrachloride-induced liver fibrogenesis. Chem. Biol. Interact. 2004, 148, 125–138. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Liver Enzymes | Kidney Markers | |||
|---|---|---|---|---|---|
| AST (U/L) | ALT (U/L) | Serum Creatinine (mg/dL) | Urea (mg/dL) | Uric Acid (mg/dL) | |
| Negative control | 95.37 ± 7.13 b | 46.51 ± 1.68 b | 0.89 ± 0.037 b | 10.22 ± 0.38 b | 7.49 ± 0.25 b |
| Positive control | 250.30 ± 12.25 a | 137.50 ± 5.82 a | 1.71 ± 0.11 a | 24.56 ± 0.79 a | 12.43 ± 0.22 a |
| Silymarin (50 mg/kg) | 157.10 ± 6.58 a,b | 100.41 ± 7.76 a,b | 0.85 ± 0.01 b | 11.07 ± 0.47 b | 6.97 ± 0.24 b |
| Naringenin (20 mg/kg) | 162.90 ± 5.35 a,b | 115.50 ± 1.43 a,b | 0.77 ± 0.02 b | 15.8 ± 0.73 a,b | 7.89 ± 0.22 b |
| Naringenin (40 mg/kg) | 152.60 ± 12.82 a,b | 108.10 ± 4.64 a,b | 0.73 ± 0.02 b | 14.63 ± 0.32 a,b | 7.46 ± 0.14 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ammar, N.M.; Hassan, H.A.; Abdallah, H.M.I.; Afifi, S.M.; Elgamal, A.M.; Farrag, A.R.H.; El-Gendy, A.E.-N.G.; Farag, M.A.; Elshamy, A.I. Protective Effects of Naringenin from Citrus sinensis (var. Valencia) Peels against CCl4-Induced Hepatic and Renal Injuries in Rats Assessed by Metabolomics, Histological and Biochemical Analyses. Nutrients 2022, 14, 841. https://doi.org/10.3390/nu14040841
Ammar NM, Hassan HA, Abdallah HMI, Afifi SM, Elgamal AM, Farrag ARH, El-Gendy AE-NG, Farag MA, Elshamy AI. Protective Effects of Naringenin from Citrus sinensis (var. Valencia) Peels against CCl4-Induced Hepatic and Renal Injuries in Rats Assessed by Metabolomics, Histological and Biochemical Analyses. Nutrients. 2022; 14(4):841. https://doi.org/10.3390/nu14040841
Chicago/Turabian StyleAmmar, Naglaa M., Heba A. Hassan, Heba M. I. Abdallah, Sherif M. Afifi, Abdelbaset M. Elgamal, Abdel Razik H. Farrag, Abd El-Nasser G. El-Gendy, Mohamed A. Farag, and Abdelsamed I. Elshamy. 2022. "Protective Effects of Naringenin from Citrus sinensis (var. Valencia) Peels against CCl4-Induced Hepatic and Renal Injuries in Rats Assessed by Metabolomics, Histological and Biochemical Analyses" Nutrients 14, no. 4: 841. https://doi.org/10.3390/nu14040841
APA StyleAmmar, N. M., Hassan, H. A., Abdallah, H. M. I., Afifi, S. M., Elgamal, A. M., Farrag, A. R. H., El-Gendy, A. E.-N. G., Farag, M. A., & Elshamy, A. I. (2022). Protective Effects of Naringenin from Citrus sinensis (var. Valencia) Peels against CCl4-Induced Hepatic and Renal Injuries in Rats Assessed by Metabolomics, Histological and Biochemical Analyses. Nutrients, 14(4), 841. https://doi.org/10.3390/nu14040841