
Does a Higher Protein Diet Promote Satiety and Weight Loss Independent of Carbohydrate Content? An 8-Week Low-Energy Diet (LED) Intervention

 ,
, 
 , and
, and
Abstract

1. Introduction
2. Materials and Methods
2.1. Trial Design
2.2. Participants
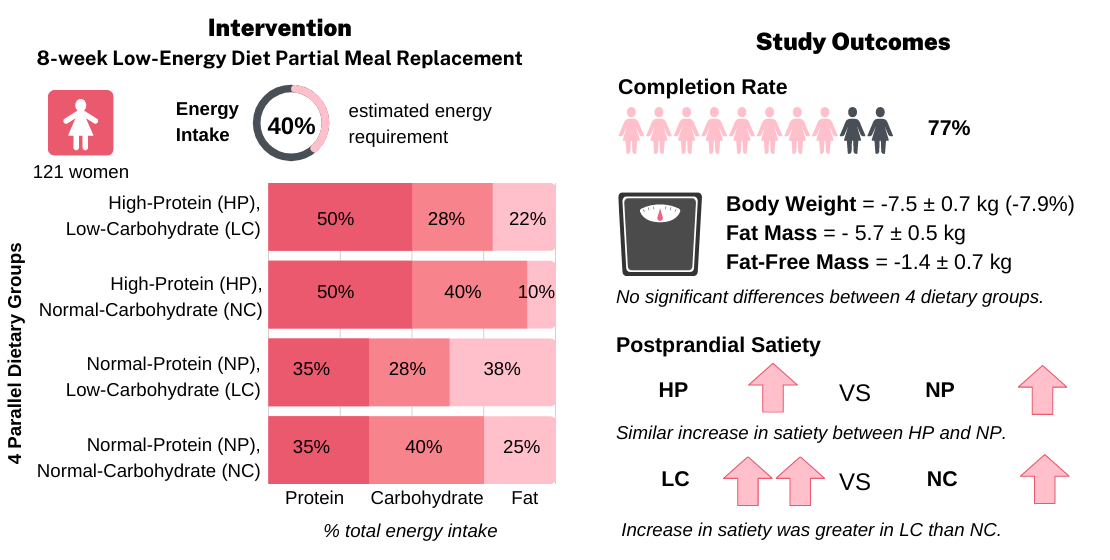
2.3. LED Intervention
= 0.4 × BMR (Harris-Benedict’s Equation for women) × Estimated Physical Activity Level (PAL)
= 0.4 × [655 + (9.6 × weight in kg) + (1.8 × height in cm) − (4.7 × age in years)] × 4.184 (conversion from kcal to kJ) × 1.375 (assumed undertaking light activity at work)
2.4. Dietary Intake Assessment
2.5. Anthropometry and Body Composition Measurements
2.6. Blood and Urine Samples
2.7. Appetite Assessment—Preload Postprandial Challenge Protocol
2.8. Randomisation and Blinding
2.9. Statistical Methods
3. Results
3.1. Participants
3.2. Dietary Intake
3.3. Body Weight
3.4. Anthropometry and Body Composition
3.5. Serum Lipids
3.6. Plasma Glucose and Glucoregulatory Peptides
3.7. Plasma Appetite-Related Gut Peptides
3.8. Plasma Amino Acids
3.9. Appetite Responses
3.10. Postprandial Appetite-Related Gut Peptides Responses
4. Discussion
4.1. Effect of LEDpdr Intervention of BW, FM, and FFM
4.2. Effect of LEDpdr Intervention on Appetite
4.3. Effect of LEDpdr Intervention on Biomarkers of Metabolic Health
4.4. Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Astbury, N.M.; Aveyard, P.; Nickless, A.; Hood, K.; Corfield, K.; Lowe, R.; Jebb, S.A. Doctor Referral of Overweight People to Low Energy total diet replacement Treatment (DROPLET): Pragmatic randomised controlled trial. BMJ 2018, 362, k3760. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.; Leeds, A.R. Very low-energy and low-energy formula diets: Effects on weight loss, obesity co-morbidities and type 2 diabetes remission—An update on the evidence for their use in clinical practice. Nutr. Bull. 2019, 44, 7–24. [Google Scholar] [CrossRef]
- Thom, G.; Lean, M. Is There an Optimal Diet for Weight Management and Metabolic Health? Gastroenterology 2017, 152, 1739–1751. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Sadeghirad, B.; Ball, G.D.C.; Da Costa, B.R.; Hitchcock, C.L.; Svendrovski, A.; Kiflen, R.; Quadri, K.; Kwon, H.Y.; Karamouzian, M.; et al. Comparison of dietary macronutrient patterns of 14 popular named dietary programmes for weight and cardiovascular risk factor reduction in adults: Systematic review and network meta-analysis of randomised trials. BMJ 2020, 369, m696. [Google Scholar] [CrossRef]
- San-Cristobal, R.; Navas-Carretero, S.; Martínez-González, M. Ángel; Ordovas, J.M.; Martínez, J.A. Contribution of macronutrients to obesity: Implications for precision nutrition. Nat. Rev. Endocrinol. 2020, 16, 305–320. [Google Scholar] [CrossRef]
- Westerterp-Plantenga, M.S.; Lemmens, S.G.; Westerterp, K.R. Dietary protein—Its role in satiety, energetics, weight loss and health. Br. J. Nutr. 2012, 108, S105–S112. [Google Scholar] [CrossRef]
- Leidy, H.J.; Clifton, P.M.; Astrup, A.; Wycherley, T.P.; Westerterp-Plantenga, M.S.; Luscombe-Marsh, N.D.; Woods, S.C.; Mattes, R.D. The role of protein in weight loss and maintenance. Am. J. Clin. Nutr. 2015, 101, 1320S–1329S. [Google Scholar] [CrossRef]
- Hansen, T.T.; Astrup, A.; Sjödin, A. Are Dietary Proteins the Key to Successful Body Weight Management? A Systematic Review and Meta-Analysis of Studies Assessing Body Weight Outcomes after Interventions with Increased Dietary Protein. Nutrients 2021, 13, 3193. [Google Scholar] [CrossRef]
- Magkos, F. The role of dietary protein in obesity. Rev. Endocr. Metab. Disord. 2020, 21, 329–340. [Google Scholar] [CrossRef]
- Soenen, S.; Bonomi, A.G.; Lemmens, S.G.; Scholte, J.; Thijssen, M.A.; van Berkum, F.; Westerterp-Plantenga, M.S. Relatively high-protein or ‘low-carb’ energy-restricted diets for body weight loss and body weight maintenance? Physiol. Behav. 2012, 107, 374–380. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products; Nutrition and Allergies (NDA). Guidance on the scientific requirements for health claims related to appetite ratings, weight management, and blood glucose concentrations. EFSA J. 2012, 10, 2604. [Google Scholar] [CrossRef]
- Institute of Medicine. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein and Amino Acids; National Academies Press: Washington, DC, USA, 2005. [Google Scholar] [CrossRef]
- Westerterp-Plantenga, M.S. How are normal, high- or low-protein diets defined? Br. J. Nutr. 2007, 97, 217–218. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bosse, J.D.; Dixon, B.M. Dietary protein in weight management: A review proposing protein spread and change theories. Nutr. Metab. 2012, 9, 81. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, K.M.B.; Pizato, N.; Botelho, P.B.; Dutra, E.S.; Gonçalves, V.S.S. Dietary protein and appetite sensations in individuals with overweight and obesity: A systematic review. Eur. J. Nutr. 2020, 59, 2317–2332. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; O’Connor, L.E.; Sands, L.; Slebodnik, M.B.; Campbell, W.W. Effects of dietary protein intake on body composition changes after weight loss in older adults: A systematic review and meta-analysis. Nutr. Rev. 2016, 74, 210–224. [Google Scholar] [CrossRef]
- Soenen, S.; Martens, E.A.P.; Hochstenbach-Waelen, A.; Lemmens, S.G.; Westerterp-Plantenga, M.S. Normal Protein Intake Is Required for Body Weight Loss and Weight Maintenance, and Elevated Protein Intake for Additional Preservation of Resting Energy Expenditure and Fat Free Mass. J. Nutr. 2013, 143, 591–596. [Google Scholar] [CrossRef]
- Westman, E.C. Is dietary carbohydrate essential for human nutrition? Am. J. Clin. Nutr. 2002, 75, 951–953. [Google Scholar] [CrossRef]
- Goldenberg, J.Z.; Day, A.; Brinkworth, G.D.; Sato, J.; Yamada, S.; Jönsson, T.; Beardsley, J.; Johnson, J.A.; Thabane, L.; Johnston, B.C. Efficacy and safety of low and very low carbohydrate diets for type 2 diabetes remission: Systematic review and meta-analysis of published and unpublished randomized trial data. BMJ 2021, 372, m4743. [Google Scholar] [CrossRef]
- Paoli, A.; Rubini, A.; Volek, J.S.; Grimaldi, K.A. Beyond weight loss: A review of the therapeutic uses of very-low-carbohydrate (ketogenic) diets. Eur. J. Clin. Nutr. 2013, 67, 789–796. [Google Scholar] [CrossRef]
- Ludwig, D.S. The Ketogenic Diet: Evidence for Optimism but High-Quality Research Needed. J. Nutr. 2019, 150, 1354–1359. [Google Scholar] [CrossRef]
- Gibson, A.A.; Seimon, R.V.; Lee, C.M.Y.; Ayre, J.; Franklin, J.; Markovic, T.P.; Caterson, I.D.; Sainsbury, A. Do ketogenic diets really suppress appetite? A systematic review and meta-analysis. Obes. Rev. 2014, 16, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Gibson, A.A.; Franklin, J.; Pattinson, A.L.; Cheng, Z.G.Y.; Samman, S.; Markovic, T.P.; Sainsbury, A. Comparison of Very Low Energy Diet Products Available in Australia and How to Tailor Them to Optimise Protein Content for Younger and Older Adult Men and Women. Healthcare 2016, 4, 71. [Google Scholar] [CrossRef] [PubMed]
- Weigle, D.S.; Breen, P.A.; Matthys, C.C.; Callahan, H.S.; Meeuws, K.E.; Burden, V.R.; Purnell, J.Q. A high-protein diet induces sustained reductions in appetite, ad libitum caloric intake, and body weight despite compensatory changes in diurnal plasma leptin and ghrelin concentrations. Am. J. Clin. Nutr. 2005, 82, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Brindal, E.; Hendrie, G.A.; Taylor, P.; Freyne, J.; Noakes, M. Cohort Analysis of a 24-Week Randomized Controlled Trial to Assess the Efficacy of a Novel, Partial Meal Replacement Program Targeting Weight Loss and Risk Factor Reduction in Overweight/Obese Adults. Nutrients 2016, 8, 265. [Google Scholar] [CrossRef]
- Drummen, M.; Tischmann, L.; Gatta-Cherifi, B.; Adam, T.; Westerterp-Plantenga, M. Dietary Protein and Energy Balance in Relation to Obesity and Co-morbidities. Front. Endocrinol. 2018, 9, 443. [Google Scholar] [CrossRef]
- Mars, M.; Stafleu, A.; de Graaf, C. Use of satiety peptides in assessing the satiating capacity of foods. Physiol. Behav. 2012, 105, 483–488. [Google Scholar] [CrossRef]
- Lim, J.J.; Poppitt, S.D. How Satiating Are the ’Satiety’ Peptides: A Problem of Pharmacology versus Physiology in the Development of Novel Foods for Regulation of Food Intake. Nutrients 2019, 11, 1517. [Google Scholar] [CrossRef]
- Korompokis, K.; Östman, E.; Dougkas, A. The impact of liquid preloads varying in macronutrient content on postprandial kinetics of amino acids relative to appetite in healthy adults. Appetite 2016, 107, 511–520. [Google Scholar] [CrossRef]
- Veldhorst, M.A.; Nieuwenhuizen, A.G.; Hochstenbach-Waelen, A.; Westerterp, K.R.; Engelen, M.P.; Brummer, R.-J.M.; Deutz, N.E.; Westerterp-Plantenga, M.S. A breakfast with alpha-lactalbumin, gelatin, or gelatin+TRP lowers energy intake at lunch compared with a breakfast with casein, soy, whey, or whey-GMP. Clin. Nutr. 2009, 28, 147–155. [Google Scholar] [CrossRef]
- Camacho-Barcia, L.; García-Gavilán, J.; Papandreou, C.; Hansen, T.; Harrold, J.; Finlayson, G.; Blundell, J.; Sjödin, A.; Halford, J.; Bulló, M. Circulating Metabolites Associated with Postprandial Satiety in Overweight/Obese Participants: The SATIN Study. Nutrients 2021, 13, 549. [Google Scholar] [CrossRef]
- Lim, J.J.; Sequeira, I.R.; Yip, W.C.; Lu, L.W.; Barnett, D.; Cameron-Smith, D.; Poppitt, S.D. Postprandial glycine as a biomarker of satiety: A dose-rising randomised control trial of whey protein in overweight women. Appetite 2021, 169, 105871. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, M.; Lewis, G.D.; Ericson, U.; Orho-Melander, M.; Hedblad, B.; Engstrom, G.; Ostling, G.; Clish, C.; Wang, T.J.; Gerszten, R.E.; et al. A diabetes-predictive amino acid score and future cardiovascular disease. Eur. Heart J. 2013, 34, 1982–1989. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; McGarrah, R.W.; Herman, M.A.; Bain, J.R.; Shah, S.H.; Newgard, C.B. Insulin action, type 2 diabetes, and branched-chain amino acids: A two-way street. Mol. Metab. 2021, 52, 101261. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Health New Zealand. Clinical Guidelines for Weight Management in New Zealand Adults; Ministry of Health New Zealand: Wellington, New Zealand, 2017. [Google Scholar]
- Jebb, S.A.; Astbury, N.M.; Tearne, S.; Nickless, A.; Aveyard, P. Doctor Referral of Overweight People to a Low-Energy Treatment (DROPLET) in primary care using total diet replacement products: A protocol for a randomised controlled trial. BMJ Open 2017, 7, e016709. [Google Scholar] [CrossRef]
- Maroni, B.J.; Steinman, T.I.; Mitch, W.E. A method for estimating nitrogen intake of patients with chronic renal failure. Kidney Int. 1985, 27, 58–65. [Google Scholar] [CrossRef]
- Altman, D.G.; Bland, J.M. Measurement in Medicine: The Analysis of Method Comparison Studies. J. R. Stat. Soc. Ser. D 1983, 32, 307–317. [Google Scholar] [CrossRef]
- Cohen, S.; Michaud, D. Synthesis of a Fluorescent Derivatizing Reagent, 6-Aminoquinolyl-N-Hydroxysuccinimidyl Carbamate, and Its Application for the Analysis of Hydrolysate Amino Acids via High-Performance Liquid Chromatography. Anal. Biochem. 1993, 211, 279–287. [Google Scholar] [CrossRef]
- Prodhan, U.K.; Pundir, S.; Chiang, V.S.-C.; Milan, A.M.; Barnett, M.P.G.; Smith, G.C.; Markworth, J.F.; Knowles, S.O.; Cameron-Smith, D. Comparable Postprandial Amino Acid and Gastrointestinal Hormone Responses to Beef Steak Cooked Using Different Methods: A Randomised Crossover Trial. Nutrients 2020, 12, 380. [Google Scholar] [CrossRef]
- Blundell, J.; de Graaf, C.; Hulshof, T.; Jebb, S.; Livingstone, B.; Lluch, A.; Mela, D.; Salah, S.; Schuring, E.; van der Knaap, H.; et al. Appetite control: Methodological aspects of the evaluation of foods. Obes. Rev. 2010, 11, 251–270. [Google Scholar] [CrossRef]
- Gibbons, C.; Hopkins, M.; Beaulieu, K.; Oustric, P.; Blundell, J.E. Issues in Measuring and Interpreting Human Appetite (Satiety/Satiation) and Its Contribution to Obesity. Curr. Obes. Rep. 2019, 8, 77–87. [Google Scholar] [CrossRef]
- Wiessing, K.R.; Xin, L.; McGill, A.-T.; Budgett, S.C.; Strik, C.M.; Poppitt, S.D. Sensitivity of ad libitum meals to detect changes in hunger. Restricted-item or multi-item testmeals in the design of preload appetite studies. Appetite 2012, 58, 1076–1082. [Google Scholar] [CrossRef] [PubMed]
- Randomizer. Available online: www.randomizer.org (accessed on 13 February 2021).
- Christensen, P.; Bliddal, H.; Riecke, B.F.; Leeds, A.R.; Astrup, A. Comparison of a low-energy diet and a very low-energy diet in sedentary obese individuals: A pragmatic randomized controlled trial. Clin. Obes. 2011, 1, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Christensen, P.; Larsen, T.M.; Westerterp-Plantenga, M.; Macdonald, I.; Martinez, J.A.; Handjiev, S.; Poppitt, S.; Hansen, S.; Ritz, C.; Astrup, A.; et al. Men and women respond differently to rapid weight loss: Metabolic outcomes of a multi-centre intervention study after a low-energy diet in 2500 overweight, individuals with pre-diabetes (PREVIEW). Diabetes Obes. Metab. 2018, 20, 2840–2851. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.T.; Hjorth, M.F.; Sandby, K.; Andersen, S.V.; Astrup, A.; Ritz, C.; Bulló, M.; Camacho-Barcía, M.L.; García-Gavilán, J.F.; Salas-Salvadó, J.; et al. Predictors of successful weight loss with relative maintenance of fat-free mass in individuals with overweight and obesity on an 8-week low-energy diet. Br. J. Nutr. 2019, 122, 468–479. [Google Scholar] [CrossRef]
- Leslie, W.S.; Taylor, R.; Harris, L.; Lean, M.E.J. Weight losses with low-energy formula diets in obese patients with and without type 2 diabetes: Systematic review and meta-analysis. Int. J. Obes. 2017, 41, 96–101. [Google Scholar] [CrossRef]
- Soenen, S.; Hochstenbach-Waelen, A.; Westerterp-Plantenga, M.S. Efficacy of α-Lactalbumin and Milk Protein on Weight Loss and Body Composition during Energy Restriction. Obesity 2011, 19, 370–379. [Google Scholar] [CrossRef]
- Weinheimer, E.M.; Sands, L.P.; Campbell, W.W. A systematic review of the separate and combined effects of energy restriction and exercise on fat-free mass in middle-aged and older adults: Implications for sarcopenic obesity. Nutr. Rev. 2010, 68, 375–388. [Google Scholar] [CrossRef]
- Wycherley, T.P.; Moran, L.J.; Clifton, P.M.; Noakes, M.; Brinkworth, G.D. Effects of energy-restricted high-protein, low-fat compared with standard-protein, low-fat diets: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2012, 96, 1281–1298. [Google Scholar] [CrossRef]
- Clifton, P.; Condo, D.; Keogh, J. Long term weight maintenance after advice to consume low carbohydrate, higher protein diets—A systematic review and meta analysis. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 224–235. [Google Scholar] [CrossRef]
- Vogtschmidt, Y.D.; Raben, A.; Faber, I.; de Wilde, C.; Lovegrove, J.A.; Givens, D.I.; Pfeiffer, A.F.; Soedamah-Muthu, S.S. Is protein the forgotten ingredient: Effects of higher compared to lower protein diets on cardiometabolic risk factors. A systematic review and meta-analysis of randomised controlled trials. Atherosclerosis 2021, 328, 124–135. [Google Scholar] [CrossRef]
- Andriessen, C.; Christensen, P.; Nielsen, L.V.; Ritz, C.; Astrup, A.; Larsen, T.M.; Martinez, J.A.; Saris, W.H.; van Baak, M.A.; Papadaki, A.; et al. Weight loss decreases self-reported appetite and alters food preferences in overweight and obese adults: Observational data from the DiOGenes study. Appetite 2018, 125, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Lyngstad, A.; Nymo, S.; Coutinho, S.R.; Rehfeld, J.F.; Truby, H.; Kulseng, B.; Martins, C. Investigating the effect of sex and ketosis on weight-loss-induced changes in appetite. Am. J. Clin. Nutr. 2019, 109, 1511–1518. [Google Scholar] [CrossRef] [PubMed]
- Adam, T.C.M.; Lejeune, M.P.G.M.; Westerterp-Plantenga, M.S. Nutrient-stimulated glucagon-like peptide 1 release after body-weight loss and weight maintenance in human subjects. Br. J. Nutr. 2006, 95, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Sumithran, P.; Prendergast, L.A.; Delbridge, E.; Purcell, K.; Shulkes, A.; Kriketos, A.D.; Proietto, J. Ketosis and appetite-mediating nutrients and hormones after weight loss. Eur. J. Clin. Nutr. 2013, 67, 759–764. [Google Scholar] [CrossRef]
- Nymo, S.; Coutinho, S.R.; Jørgensen, J.; Rehfeld, J.F.; Truby, H.; Kulseng, B.; Martins, C. Timeline of changes in appetite during weight loss with a ketogenic diet. Int. J. Obes. 2017, 41, 1224–1231. [Google Scholar] [CrossRef]
- Halliday, T.M.; Polsky, S.; Schoen, J.A.; Legget, K.T.; Tregellas, J.R.; Williamson, K.M.; Cornier, M. Comparison of surgical versus diet-induced weight loss on appetite regulation and metabolic health outcomes. Physiol. Rep. 2019, 7, e14048. [Google Scholar] [CrossRef]
- Deemer, S.; Plaisance, E.P.; Martins, C. Impact of ketosis on appetite regulation—A review. Nutr. Res. 2020, 77, 1–11. [Google Scholar] [CrossRef]
- Sumithran, P.; Prendergast, L.A.; Delbridge, E.; Purcell, K.; Shulkes, A.; Kriketos, A.; Proietto, J. Long-Term Persistence of Hormonal Adaptations to Weight Loss. N. Engl. J. Med. 2011, 365, 1597–1604. [Google Scholar] [CrossRef]
- Sumithran, P.; Proietto, J. The defence of body weight: A physiological basis for weight regain after weight loss. Clin. Sci. 2012, 124, 231–241. [Google Scholar] [CrossRef]
- Hansen, T.T.; Andersen, S.V.; Astrup, A.; Blundell, J.; Sjödin, A. Is reducing appetite beneficial for body weight management in the context of overweight and obesity? A systematic review and meta-analysis from clinical trials assessing body weight management after exposure to satiety enhancing and/or hunger reducing products. Obes. Rev. 2019, 20, 983–997. [Google Scholar] [CrossRef]
- Martins, C.; Nymo, S.; Truby, H.; Rehfeld, J.F.; Hunter, G.R.; Gower, B.A. Association Between Ketosis and Changes in Appetite Markers with Weight Loss Following a Very Low-Energy Diet. Obesity 2020, 28, 2331–2338. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.; Roekenes, J.; Hunter, G.R.; Gower, B.A. Association between ketosis and metabolic adaptation at the level of resting metabolic rate. Clin. Nutr. 2021, 40, 4824–4829. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.; Yoshino, J.; Kelly, S.C.; Reeds, D.N.; Okunade, A.; Patterson, B.W.; Klein, S.; Mittendorfer, B. High-Protein Intake during Weight Loss Therapy Eliminates the Weight-Loss-Induced Improvement in Insulin Action in Obese Postmenopausal Women. Cell Rep. 2016, 17, 849–861. [Google Scholar] [CrossRef] [PubMed]
- Raben, A.; Vestentoft, P.S.; Brand-Miller, J.; Jalo, E.; Drummen, M.; Simpson, L.; Martinez, J.A.; Handjieva-Darlenska, T.; Stratton, G.; Huttunen-Lenz, M.; et al. The PREVIEW intervention study: Results from a 3-year randomized 2 x 2 factorial multinational trial investigating the role of protein, glycaemic index and physical activity for prevention of type 2 diabetes. Diabetes Obes. Metab. 2021, 23, 324–337. [Google Scholar] [CrossRef]
- Rietman, A.; Schwarz, J.; Tome, D.; Kok, F.J.; Mensink, M. High dietary protein intake, reducing or eliciting insulin resistance? Eur. J. Clin. Nutr. 2014, 68, 973–979. [Google Scholar] [CrossRef]
- Veldhorst, M.A.B.; Westerterp, K.R.; Westerterp-Plantenga, M.S. Gluconeogenesis and protein-induced satiety. Br. J. Nutr. 2012, 107, 595–600. [Google Scholar] [CrossRef]
- Santos, H.O.; Lavie, C.J. Weight loss and its influence on high-density lipoprotein cholesterol (HDL-C) concentrations: A noble clinical hesitation. Clin. Nutr. ESPEN 2021, 42, 90–92. [Google Scholar] [CrossRef]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A Branched-Chain Amino Acid-Related Metabolic Signature that Differentiates Obese and Lean Humans and Contributes to Insulin Resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef]
- Zhou, Y.; Qiu, L.; Xiao, Q.; Wang, Y.; Meng, X.; Xu, R.; Wang, S.; Na, R. Obesity and diabetes related plasma amino acid alterations. Clin. Biochem. 2013, 46, 1447–1452. [Google Scholar] [CrossRef]
- Boulet, M.M.; Chevrier, G.; Grenier-Larouche, T.; Pelletier, M.; Nadeau, M.; Scarpa, J.; Prehn, C.; Marette, A.; Adamski, J.; Tchernof, A. Alterations of plasma metabolite profiles related to adipose tissue distribution and cardiometabolic risk. Am. J. Physiol. Metab. 2015, 309, E736–E746. [Google Scholar] [CrossRef]
- Takashina, C.; Tsujino, I.; Watanabe, T.; Sakaue, S.; Ikeda, D.; Yamada, A.; Sato, T.; Ohira, H.; Otsuka, Y.; Oyama-Manabe, N.; et al. Associations among the plasma amino acid profile, obesity, and glucose metabolism in Japanese adults with normal glucose tolerance. Nutr. Metab. 2016, 13, 5. [Google Scholar] [CrossRef]
- Yamakado, M.; Tanaka, T.; Nagao, K.; Ishizaka, Y.; Mitushima, T.; Tani, M.; Toda, A.; Toda, E.; Okada, M.; Miyano, H.; et al. Plasma amino acid profile is associated with visceral fat accumulation in obese Japanese subjects. Clin. Obes. 2012, 2, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Perez-Cornago, A.; Brennan, L.; Ibero-Baraibar, I.; Hermsdorff, H.H.M.; O’Gorman, A.; Zulet, M.A.; Martínez, J.A. Metabolomics identifies changes in fatty acid and amino acid profiles in serum of overweight older adults following a weight loss intervention. J. Physiol. Biochem. 2014, 70, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Ceglarek, U.; Huang, T.; Li, L.; Rood, J.; Ryan, D.H.; Bray, G.A.; Sacks, F.M.; Schwarzfuchs, D.; Thiery, J.; et al. Weight-loss diets and 2-y changes in circulating amino acids in 2 randomized intervention trials1–3. Am. J. Clin. Nutr. 2016, 103, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Geidenstam, N.; Magnusson, M.; Danielsson, A.P.H.; Gerszten, R.E.; Wang, T.; Reinius, L.E.; Mulder, H.; Melander, O.; Ridderstråle, M. Amino Acid Signatures to Evaluate the Beneficial Effects of Weight Loss. Int. J. Endocrinol. 2017, 2017, 6490473. [Google Scholar] [CrossRef]
- Prodhan, U.; Milan, A.; Silvestre, M.; Christensen, P.; Raben, A.; Fogelholm, M.; Poppitt, S.; Cameron-Smith, D. Differential Trajectories in Altered Insulin Sensitivity Following Weight Loss and Their Impact on Circulatory Amino Acids: Results from the PREVIEW: New Zealand Sub-study (OR27-07-19). Curr. Dev. Nutr. 2019, 3, nzz046-OR27. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products; Nutrition and Allergies (NDA). Scientific Opinion on the essential composition of total diet replacements for weight control. EFSA J. 2015, 13, 3957. [Google Scholar] [CrossRef]
- Mitra, S.R.; Tan, P.Y. Effect of an individualised high-protein, energy-restricted diet on anthropometric and cardio-metabolic parameters in overweight and obese Malaysian adults: A 6-month randomised controlled study. Br. J. Nutr. 2019, 121, 1002–1017. [Google Scholar] [CrossRef]
- Skov, A.; Toubro, S.; Rønn, B.; Holm, L.; Astrup, A. Randomized trial on protein vs carbohydrate in ad libitum fat reduced diet for the treatment of obesity. Int. J. Obes. 1999, 23, 528–536. [Google Scholar] [CrossRef]
- Hebert, J.R.; Clemow, L.; Pbert, L.; Ockene, I.S.; Ockene, J.K. Social Desirability Bias in Dietary Self-Report May Compromise the Validity of Dietary Intake Measures. Int. J. Epidemiol. 1995, 24, 389–398. [Google Scholar] [CrossRef]
- Bingham, S.A. Urine Nitrogen as a Biomarker for the Validation of Dietary Protein Intake. J. Nutr. 2003, 133 (Suppl. S3), 921S–924S. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Higher Protein (HP) | Normal Protein (NP) | |||
|---|---|---|---|---|
| Lower CHO (LC) | Normal CHO (NC) | Lower CHO (LC) | Normal CHO (NC) | |
| Protein | 50%en | 50%en | 35%en | 35%en |
| CHO | 28%en | 40%en | 28%en | 40%en |
| Fat | 22%en | 10%en | 38%en | 25%en |
| Characteristics | HPLC (n = 31) | HPNC (n = 29) | NPLC (n = 31) | NPNC (n = 30) | p-Value |
|---|---|---|---|---|---|
| Age (years) | 39.0 ± 10.1 | 37.4 ± 12.0 | 41.2 ± 10.2 | 41.3 ± 12.4 | 0.472 |
| BW (kg) | 95.5 ± 11.3 | 95.6 ± 12.4 | 93.9 ± 12.8 | 95.3 ± 15.8 | 0.956 |
| Height (m) | 1.63 ± 0.07 | 1.64 ± 0.07 | 1.63 ± 0.08 | 1.64 ± 0.07 | 0.863 |
| BMI (kg/m2) | 35.9 ± 4.0 | 35.3 ± 3.5 | 35.3 ± 4.0 | 35.2 ± 4.4 | 0.898 |
| Waist circumference (cm) | 100.7 ± 7.5 | 101.6 ± 8.9 | 100.5 ± 8.8 | 100.1 ± 10.1 | 0.935 |
| Hip circumference (cm) | 120.8 ± 9.4 | 121.3 ± 8.8 | 119.5 ± 7.4 | 118.5 ± 10.8 | 0.627 |
| WHR | 0.84 ± 0.07 | 0.84 ± 0.04 | 0.84 ± 0.06 | 0.84 ± 0.06 | 0.910 |
| SBP (mmHg) | 119 ± 13 | 115 ± 13 | 115 ± 13 | 116 ± 13 | 0.684 |
| DBP (mmHg) | 65 ± 6 a | 62 ± 6 a,b | 61 ± 6 b | 64 ± 7 a,b | 0.040 |
| FM (kg) | 44.6 ± 7.8 | 44.5 ± 8.4 | 44.0 ± 7.5 | 44.6 ± 10.6 | 0.990 |
| Percentage FM (%) | 46.8 ± 4.7 | 46.6 ± 4.0 | 47.0 ± 2.9 | 46.7 ± 4.5 | 0.981 |
| FFM (kg) | 50.3 ± 6.2 | 50.5 ± 5.9 | 49.3 ± 6.2 | 50.2 ± 6.9 | 0.884 |
| Analysis Methods | Week | HPLC | HPNC | NPLC | NPNC | p-Value | ||
|---|---|---|---|---|---|---|---|---|
| Protein×Week | CHO×Week | Protein×CHO×Week | ||||||
| Multiple Imputation 1 | 0 | 95.1 ± 0.7 | 95.1 ± 0.7 | 95.0 ± 0.7 | 95.1 ± 0.7 | 0.733 | 0.454 | 0.928 |
| 4 | 89.5 ± 0.8 | 90.9 ± 0.8 | 89.6 ± 0.7 | 90.4 ± 1.0 | ||||
| 8 | 87.2 ± 1.3 | 88.8 ± 0.9 | 86.6 ± 0.8 | 87.6 ± 1.2 | ||||
| Observed Cases 2 | 0 | 94.0 ± 1.8 | 96.3 ± 2.4 | 93.5 ± 2.5 | 94.1 ± 3.7 | 0.432 | 0.116 | 0.469 |
| 4 | 88.7 ± 1.8 | 91.5 ± 2.6 | 88.0 ± 2.2 | 89.0 ± 3.7 | ||||
| 8 | 85.7 ± 2.0 | 89.6 ± 2.8 | 85.2 ± 2.2 | 86.1 ± 3.8 | ||||
| Variables | Baseline | Change from Week 0 | p-Value | |
|---|---|---|---|---|
| Week 0 | Week 4 | Week 8 | Week | |
| Anthropometry | ||||
| BMI (kg/m2) | 35.4 ± 0.1 | −1.8 ± 0.2 *** | −2.8 ± 0.2 *** | <0.001 |
| Waist (cm) | 100.7 ± 0.4 | −4.0 ± 0.6 *** | −7.1 ± 0.7 *** | <0.001 |
| WHR | 0.840 ± 0.004 | −0.003 ± 0.005 ns | −0.019 ± 0.008 ns | 0.038 |
| SBP (mm Hg) | 116.3 ± 0.7 | −4.0 ± 1.0 *** | −7.4 ± 1.0 *** | <0.001 |
| DBP (mm Hg) | 62.7 ± 0.4 | −2.6 ± 0.5 *** | −3.4 ± 0.5 *** | <0.001 |
| Body composition | ||||
| FM (kg) | 44.4 ± 0.2 | NA | −5.7 ± 0.5 *** | <0.001 |
| FM (% body mass) | 46.8 ± 0.3 | NA | −2.7 ± 0.5 *** | <0.001 |
| FFM (kg) | 50.1 ± 0.4 | NA | −1.4 ± 0.7 ns | 0.065 |
| Serum lipids | ||||
| Total-C (mM) | 5.29 ± 0.06 | NA | −0.91 ± 0.11 *** | <0.001 |
| HDL-C (mM) | 1.44 ± 0.02 | NA | −0.21 ± 0.03 *** | <0.001 |
| Total:HDL-C (mM) | 3.95 ± 0.10 | NA | −0.22 ± 0.15 ns | 0.203 |
| LDL-C (mM) | 3.45 ± 0.05 | NA | −0.61 ± 0.08 *** | <0.001 |
| Trig (mM) | 1.52 ± 0.08 | NA | −0.54 ± 0.12 *** | 0.001 |
| NEFA (mM) | 0.49 ± 0.02 | NA | 0.17 ± 0.03 *** | <0.001 |
| Glucose and glucoregulatory peptides | ||||
| Glucose (mM) | 5.67 ± 0.05 | −0.31 ± 0.07 *** | −0.36 ± 0.08 *** | <0.001 |
| Insulin (pg/mL) | 724.2 (CI: 646.1, 812.1) | −231.1 (CI: −334.6, −127.9) *** | −185.3 (CI: −284.3, −86.2) *** | <0.001 |
| Glucagon (pg/mL) | 49.0 (CI: 45.6, 53.0) | −7.4 (CI: −12.4, −2.5) ** | −10.3 (CI: −15.9, −4.7) *** | <0.001 |
| GIP (pg/mL) | 45.6 (CI: 41.7–49.9) | −7.9 (CI: −14.1, −1.7) * | −6.9 (CI: −13.1, −0.8) ns | <0.001 |
| Gut peptides | ||||
| GLP-1 (pg/mL) | 172.9 (CI: 158.5, 188.6) | −7.2 (CI: −26.1, 11.8) ns | −27.4 (CI: −48.6, −6.3) * | 0.051 |
| PYY (pg/mL) | 37.6 (CI: 32.7, 43.3) | −5.4 (CI: −10.5, −0.2) ns | −2.8 (CI: −7.6, 2.1) ns | 0.113 |
| Branched-chain amino acids | ||||
| Leucine (µM) | 121.0 ± 1.4 | −1.3 ± 2.4 ns | 1.4 ± 2.5 ns | 0.515 |
| Isoleucine (µM) | 65.1 ± 1.2 | 1.2 ± 1.8 ns | 1.4 ± 1.7 ns | 0.701 |
| Valine (µM) | 236.3 ± 3.8 | −1.4 ± 5.0 ns | 2.9 ± 4.7 ns | 0.645 |
| Other essential amino acids | ||||
| Phenylalanine (µM) | 55.8 ± 0.7 | −2.5 ± 1.0 * | −3.2 ± 1.2 * | 0.025 |
| Methionine (µM) | 27.9 ± 0.5 | −1.6 ± 0.6 * | −2.2 ± 0.7 ** | 0.006 |
| Lysine (µM) | 83.8 ± 1.2 | 0.4 ± 1.8 ns | −3.3 ± 1.7 ns | 0.086 |
| Histidine (µM) | 53.9 ± 1.2 | −1.7 ± 1.7 ns | −0.4 ± 1.8 ns | 0.597 |
| Threonine (µM) | 111.7 ± 1.9 | −1.6 ± 2.6 ns | −8.6 ± 2.8 ** | 0.003 |
| Tryptophan (µM) | 39.7 ± 0.7 | −1.7 ± 1.1 ns | −3.4 ± 1.4 * | 0.053 |
| Non-essential amino acids | ||||
| Glycine (µM) | 275.5 ± 6.1 | 60.2 ± 9.1 *** | −8.0 ± 9.3 ns | <0.001 |
| Aspartic acid (µM) | 5.5 ± 0.3 | −1.6 ± 0.4 *** | −1.4 ± 0.4 ** | <0.001 |
| Asparagine (µM) | 48.0 ± 0.6 | −0.1 ± 0.9 ns | −2.4 ± 1.0 * | 0.016 |
| Glutamic acid (µM) | 55.0 ± 1.8 | −12.5 ± 2.7 *** | −12.1 ± 2.8 *** | <0.001 |
| Glutamine (µM) | 534.7 ± 5.0 | −5.2 ± 7.2 ns | −9.9 ± 7.5 ns | 0.458 |
| Arginine (µM) | 76.2 ± 1.7 | −0.4 ± 2.6 ns | −1.9 ± 2.5 ns | 0.738 |
| Alanine (µM) | 404.6 ± 5.3 | −59.5 ± 7.8 *** | −70.2 ± 7.8 *** | <0.001 |
| Serine (µM) | 105.5 ± 1.7 | 10.7 ± 2.4 *** | 14.6 ± 2.6 *** | <0.001 |
| Tyrosine (µM) | 66.9 ± 0.9 | −7.6 ± 1.2 *** | −8.9 ± 1.4 *** | <0.001 |
| Proline (µM) | 208.1 ± 3.6 | −17.5 ± 5.3 ** | −39.0 ± 5.8 *** | <0.001 |
| Non-proteogenic amino acids | ||||
| Hydroxyproline (µM) | 13.8 ± 0.5 | −1.8 ± 0.7 * | −3.2 ± 0.7 *** | <0.001 |
| Taurine (µM) | 114.7 ± 4.7 | 1.2 ± 7.1 ns | −15.9 ± 6.9 ns | 0.033 |
| Citrulline (µM) | 29.4 ± 0.3 | −2.3 ± 0.5 *** | −2.1 ± 0.5 *** | <0.001 |
| Ornithine (µM) | 38.9 ± 0.9 | −3.8 ± 1.3 * | −2.7 ± 1.2 ns | 0.010 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, J.J.; Liu, Y.; Lu, L.W.; Barnett, D.; Sequeira, I.R.; Poppitt, S.D. Does a Higher Protein Diet Promote Satiety and Weight Loss Independent of Carbohydrate Content? An 8-Week Low-Energy Diet (LED) Intervention. Nutrients 2022, 14, 538. https://doi.org/10.3390/nu14030538
Lim JJ, Liu Y, Lu LW, Barnett D, Sequeira IR, Poppitt SD. Does a Higher Protein Diet Promote Satiety and Weight Loss Independent of Carbohydrate Content? An 8-Week Low-Energy Diet (LED) Intervention. Nutrients. 2022; 14(3):538. https://doi.org/10.3390/nu14030538
Chicago/Turabian StyleLim, Jia Jiet, Yutong Liu, Louise Weiwei Lu, Daniel Barnett, Ivana R. Sequeira, and Sally D. Poppitt. 2022. "Does a Higher Protein Diet Promote Satiety and Weight Loss Independent of Carbohydrate Content? An 8-Week Low-Energy Diet (LED) Intervention" Nutrients 14, no. 3: 538. https://doi.org/10.3390/nu14030538
APA StyleLim, J. J., Liu, Y., Lu, L. W., Barnett, D., Sequeira, I. R., & Poppitt, S. D. (2022). Does a Higher Protein Diet Promote Satiety and Weight Loss Independent of Carbohydrate Content? An 8-Week Low-Energy Diet (LED) Intervention. Nutrients, 14(3), 538. https://doi.org/10.3390/nu14030538