
Evaluation of an Ionic Calcium Fiber Supplement and Its Impact on Bone Health Preservation in a Dietary Calcium Deficiency Mice Model

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Fourier Transform Infrared Spectroscopy
2.3. Apparent Calcium Solubility
2.4. Cytotoxicity and Mutagenicity Assays
2.5. Calcium Absorption In-Vivo Experiment
2.6. X-ray Analysis
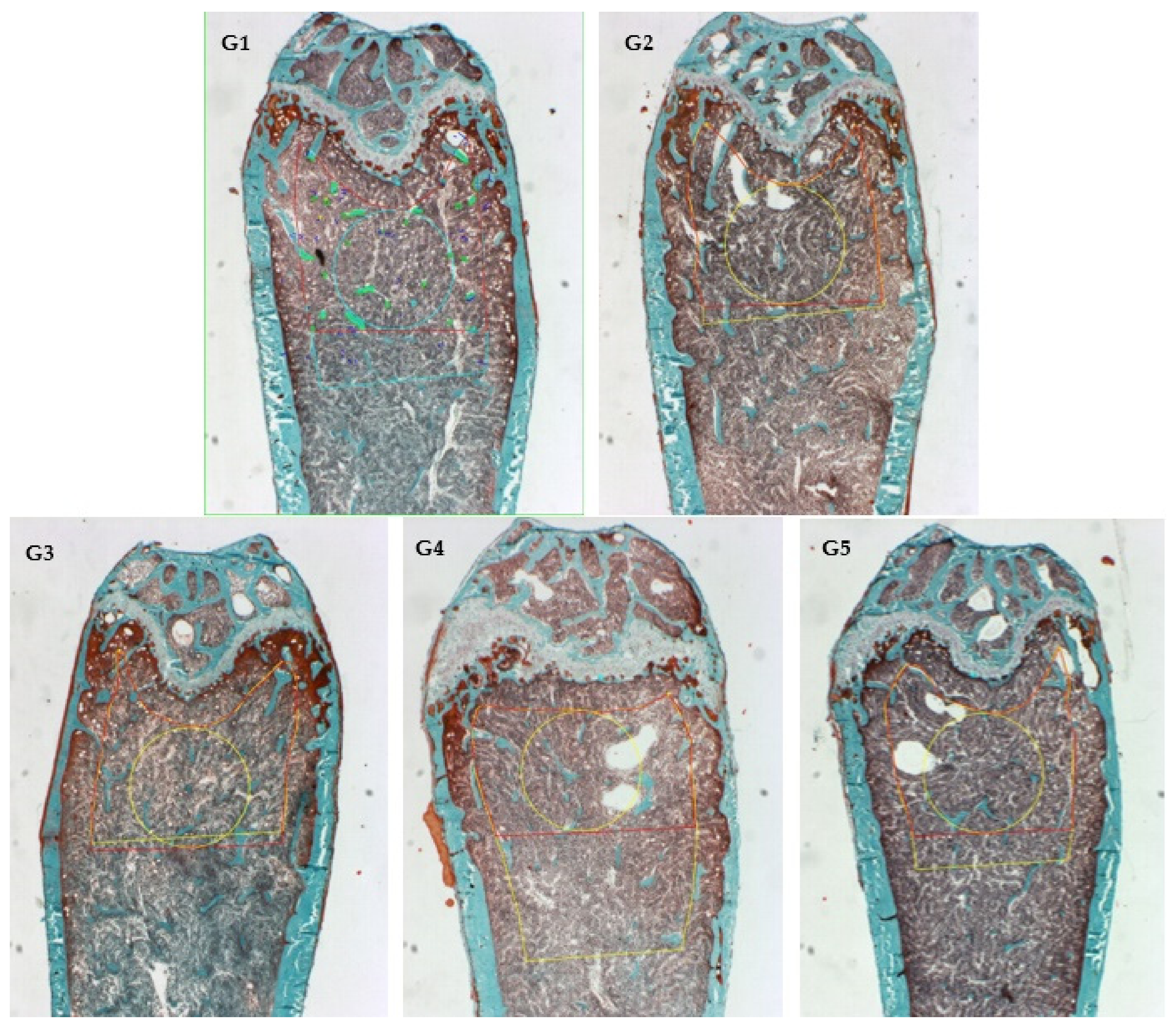
2.7. Histomorphometry Analyses (Goldner’s Trichrome Stain)
2.8. Statistical Analysis
3. Results and Discussion
3.1. Fourier Transform Infrared Spectroscopy
3.2. Apparent Calcium Solubility in the Supplements
3.3. Cytotoxicity and Mutagenicity In Vitro
3.4. Calcium Absorption In Vivo Mice Model
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Borciani, G.; Montalbano, G.; Baldini, N.; Cerqueni, G.; Vitale-Brovarone, C.; Ciapetti, G. Co–culture systems of osteoblasts and osteoclasts: Simulating in vitro bone remodeling in regenerative approaches. Acta Biomater. 2020, 108, 22–45. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; McDonald, J.M. Disorders of bone remodeling. Annu. Rev. Pathol. 2011, 6, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Wongdee, K.; Rodrat, M.; Teerapornpuntakit, J.; Krishnamra, N.; Charoenphandhu, N. Factors inhibiting intestinal calcium absorption: Hormones and luminal factors that prevent excessive calcium uptake. J. Physiol. Sci. 2019, 69, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Vannucci, L.; Fossi, C.; Quattrini, S.; Guasti, L.; Pampaloni, B.; Gronchi, G.; Giusti, F.; Romagnoli, C.; Cianferotti, L.; Marcucci, G.; et al. Calcium Intake in bone health: A focus on calcium-rich mineral waters. Nutrients 2018, 10, 1930. [Google Scholar] [CrossRef] [PubMed]
- Goss, S.; Lemons, K.; Kerstetter, J.; Bogner, R. Determination of calcium salt solubility with changes in pH and PCO2, simulating varying gastrointestinal environments. J. Pharm. Pharmacol. 2007, 59, 1485–1492. [Google Scholar] [CrossRef] [PubMed]
- Nordin, B.E.; Morris, H.A. The calcium deficiency model for osteoporosis. Nutr. Rev. 1989, 47, 65–72. [Google Scholar] [CrossRef]
- Nordin, B.E.C. Calcium and osteoporosis. Nutrition 1997, 13, 664–686. [Google Scholar] [CrossRef]
- Sipponen, P.; Härkönen, M. Hypochlorhydric stomach: A risk condition for calcium malabsorption and osteoporosis? Scand. J. Gastroenterol. 2010, 45, 133–138. [Google Scholar] [CrossRef]
- Prentice, A. Diet, nutrition and the prevention of osteoporosis. Public Health Nutr. 2004, 7, 227–243. [Google Scholar] [CrossRef]
- Ezquerra, R. Dieta y osteoporosis. Endocrinol. Nutr. 2006, 53, 296–299. [Google Scholar] [CrossRef]
- Fitzpatrick, L.A. Secondary Causes of Osteoporosis. Mayo Clin. Proc. 2002, 77, 453–468. [Google Scholar] [CrossRef]
- Mauck, K.F.; Clarke, B.L. Diagnosis, Screening, Prevention, and Treatment of Osteoporosis. Concise Rev. Clin. 2006, 81, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Kim, H.; Kim, S. Effects of adolescents’ lifestyle habits and body composition on bone mineral density. Int. J. Environ. Res. Public Health 2021, 18, 6170. [Google Scholar] [CrossRef] [PubMed]
- Greer, F.R.; Krebs, N.F.; Baker, R.D.; Bhatia, J.J.S.; Heyman, M.B.; Lifshitz, F.; Blum-Kemelor, D.; Boland, M.P.; Dietz, W.; Hubbard, V.S.; et al. Optimizing bone health and calcium intakes of infants, children, and adolescents. Pediatrics 2006, 117, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.N.; Jepsen, K.J.; Khoury, B.; Graf, K.H.; Varani, J. Bone structure and function in male C57BL/6 mice: Effects of a high-fat Western-style diet with or without trace minerals. Bone Rep. 2016, 5, 141–149. [Google Scholar] [CrossRef]
- Marstein, H.S.; Godang, K.; Flatø, B.; Sjaastad, I.; Bollerslev, J.; Sanner, H. Bone mineral density and explanatory factors in children and adults with juvenile dermatomyositis at long term follow-up; a cross sectional study. Pediatr. Rheumatol. 2021, 19, 56. [Google Scholar] [CrossRef]
- Ratajczak, A.E.; Szymczak-Tomczak, A.; Rychter, A.M.; Zawada, A.; Dobrowolska, A.; Krela-Kaźmierczak, I. Impact of Cigarette Smoking on the Risk of Osteoporosis in Inflammatory Bowel Diseases. J. Clin. Med. 2021, 10, 1515. [Google Scholar] [CrossRef]
- Scholz-Ahrens, K.E.; Adolphi, B.; Rochat, F.; Barclay, D.V.; de Vrese, M.; Açil, Y.; Schrezenmeir, J. Effects of probiotics, prebiotics, and synbiotics on mineral metabolism in ovariectomized rats—Impact of bacterial mass, intestinal absorptive area and reduction of bone turn-over. NFS J. 2016, 3, 41–50. [Google Scholar] [CrossRef]
- Xue, Z.; Yu, J.; Zhao, M.; Kang, W.; Ma, Z. Effects of synbiotics on intestinal mucosal barrier in rat model. Clin. Nutr. 2017, 13, 12–21. [Google Scholar] [CrossRef][Green Version]
- Espinosa-Andrews, H.; Urías-Silvas, J.E.; Morales-Hernández, N. The role of agave fructans in health and food applications: A review. Trends Food Sci. Technol. 2021, 114, 585–598. [Google Scholar] [CrossRef]
- Raschka, L.; Daniel, H. Mechanisms underlying the effects of inulin-type fructans on calcium absorption in the large intestine of rats. Bone 2005, 37, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Hara, H. Various nondigestible saccharides open a paracellular calcium transport pathway with the induction of intracellular calcium signaling in human intestinal Caco-2 cells. J. Nutr. 2004, 134, 1935–1941. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Huerta, M.; Lizárraga-Grimes, V.L.; Castro-Torres, I.G.; Tinoco-Méndez, M.; Macías-Rosales, L.; Sánchez-Bartéz, F.; Tapia-Pérez, G.G.; Romero-Romero, L.; Gracia-Mora, M.I. Functional Effects of Prebiotic Fructans in Colon Cancer and Calcium Metabolism in Animal Models. Biomed. Res. Int. 2017, 2017, 9758982. [Google Scholar] [CrossRef] [PubMed]
- García-Vieyra, M.I.; Del Real, A.; López, M.G. Agave fructans: Their effect on mineral absorption and bone mineral content. J. Med. Food 2014, 17, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Van den Heuvel, E.G.H.M.; Muys, T.; van Dokkum, W.; Schaafsma, G. Oligofructose stimulates calcium absorption in adolescents. Am. J. Clin. Nutr. 1999, 69, 544–548. [Google Scholar] [CrossRef] [PubMed]
- Calame, W.; Weseler, A.R.; Viebke, C.; Flynn, C.; Siemensma, A.D. Gum arabic establishes prebiotic functionality in healthy human volunteers in a dose-dependent manner. Br. J. Nutr. 2008, 100, 1269–1275. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-W.; Park, Y.-S.; Jung, J.-S.; Shin, W.-S. Chitosan oligosaccharides, dp 2-8, have prebiotic effect on the Bifidobacterium bifidium and Lactobacillus sp. Anaerobe 2002, 8, 319–324. [Google Scholar] [CrossRef]
- Espinosa-Andrews, H.; García-Márquez, E. Soluble Calcium Stabilised in an Anionic-Cationic Polymer and Fructans. U.S. Patent and Trademark Office US10709160B2, 14 July 2020. [Google Scholar]
- Metrohm Calcium and magnesium in rock extracts. 2020. Available online: https://www.metrohm.com/en-gb/applications/AN-C-040 (accessed on 25 January 2020).
- Marins, J.S.R.; Sassone, L.M.; Fidel, S.R.; Ribeiro, D.A. In vitro genotoxicity and cytotoxicity in murine fibroblasts exposed to EDTA, NaOCl, MTAD and citric acid. Braz. Dent. J. 2012, 23, 527–533. [Google Scholar] [CrossRef]
- Maron, D.M.; Ames, B.N. Revised methods for the Salmonella mutagenicity test. Mutat. Res. Mutagen. Relat. Subj. 1983, 113, 173–215. [Google Scholar] [CrossRef]
- Dempster, D.W.; Compston, J.E.; Drezner, M.K.; Glorieux, F.H.; Kanis, J.A.; Malluche, H.; Meunier, P.J.; Ott, S.M.; Recker, R.R.; Parfitt, A.M. Standardized nomenclature, symbols, and units for bone histomorphometry: A 2012 update of the report of the ASBMR Histomorphometry Nomenclature Committee. J. Bone Miner. Res. 2013, 28, 2–17. [Google Scholar] [CrossRef]
- Rodriguez-Blanco, J.D.; Shaw, S.; Benning, L.G. The kinetics and mechanisms of amorphous calcium carbonate (ACC) crystallization to calcite, viavaterite. Nanoscale 2011, 3, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Mourya, V.K.; Inamdar, N.N. Trimethyl chitosan and its applications in drug delivery. J. Mater. Sci. Mater. Med. 2009, 20, 1057–1079. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Andrews, H.; Sandoval-Castilla, O.; Vázquez-Torres, H.; Vernon-Carter, E.J.; Lobato-Calleros, C. Determination of the gum Arabic–chitosan interactions by Fourier Transform Infrared Spectroscopy and characterization of the microstructure and rheological features of their coacervates. Carbohydr. Polym. 2010, 79, 541–546. [Google Scholar] [CrossRef]
- Velázquez-Martínez, R.J.; González-Cervantes, M.R.; Hernández-Gallegos, A.M.; Mendiola, C.R.; Aparicio, R.A.; Ocampo, L.M. Prebiotic Potential of Agave angustifolia Haw Fructans with Different Degrees of Polymerization. Molecules 2014, 19, 12660–12675. [Google Scholar] [CrossRef]
- Vaisman, N.; Shaltiel, G.; Daniely, M.; Meiron, O.E.; Shechter, A.; Abrams, S.A.; Niv, E.; Shapira, Y.; Sagi, A. Increased calcium absorption from synthetic stable amorphous calcium carbonate: Double-blind randomized crossover clinical trial in postmenopausal women. J. Bone Miner. Res. 2014, 29, 2203–2209. [Google Scholar] [CrossRef]
- Michalski, R. Ion Chromatography Applications in Wastewater Analysis. Separations 2018, 5, 16. [Google Scholar] [CrossRef]
- Md Noh, M.F.; Gunasegavan, R.D.; Mustafa Khalid, N.; Balasubramaniam, V.; Mustar, S.; Abd Rashed, A. Recent Techniques in Nutrient Analysis for Food Composition Database. Molecules 2020, 25, 4567. [Google Scholar] [CrossRef]
- Pasika, W.M. Polysaccharide Polyelectrolytes; ACS Publications: Washington, DC, USA, 1977; ISBN 1947-5918. [Google Scholar]
- Lopez, C.G. Entanglement properties of polyelectrolytes in salt-free and excess-salt solutions. ACS Macro Lett. 2019, 8, 979–983. [Google Scholar] [CrossRef]
- Carr, C.J.; Shangraw, R.F. Nutritional and pharmaceutical aspects of calcium supplementation. Am. Pharm. 1987, 27, 49–56. [Google Scholar] [CrossRef]
- Rodrigues, S.; Dionísio, M.; López, C.R.; Grenha, A. Biocompatibility of Chitosan Carriers with Application in Drug Delivery. J. Funct. Biomater. 2012, 3, 615–641. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, R.; García-Carvajal, Z.Y.; Jiménez-Palomar, I.; Jiménez-Avalos, J.A.; Espinosa-Andrews, H. Development of gelatin/chitosan/PVA hydrogels: Thermal stability, water state, viscoelasticity, and cytotoxicity assays. J. Appl. Polym. Sci. 2019, 136, 47149. [Google Scholar] [CrossRef]
- De Lima, R.; Feitosa, L.; Pereira, A.d.E.S.; De Moura, M.R.; Aouada, F.A.; Mattoso, L.H.C.; Fraceto, L.F. Evaluation of the Genotoxicity of Chitosan Nanoparticles for Use in Food Packaging Films. J. Food Sci. 2010, 75, N89–N96. [Google Scholar] [CrossRef] [PubMed]
- Somerville, J.M.; Aspden, R.M.; Armour, K.E.; Armour, K.J.; Reid, D.M. Growth of C57BL/6 mice and the material and mechanical properties of cortical bone from the tibia. Calcif. Tissue Int. 2004, 74, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Tordoff, M.G. No effect of dietary calcium on body weight of lean and obese mice and rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286, R669–R677. [Google Scholar] [CrossRef][Green Version]
- McNamara, L.M. Bone as a Material. In Comprehensive Biomaterials II; Elsevier: Amsterdam, The Netherlands, 2017; Volume 2, pp. 202–227. [Google Scholar] [CrossRef]
- Berman, A.G.; Damrath, J.G.; Hatch, J.; Pulliam, A.N.; Powell, K.M.; Hinton, M.; Wallace, J.M. Effects of Raloxifene and tibial loading on bone mass and mechanics in male and female mice. Connect. Tissue Res. 2021, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Brent, M.B.; Thomsen, J.S.; Brüel, A. Short-term glucocorticoid excess blunts abaloparatide-induced increase in femoral bone mass and strength in mice. Sci. Rep. 2021, 11, 12258. [Google Scholar] [CrossRef]
- Li, J.; Wang, W.; Li, M.; Liu, L. Repair of segmental bone defect using tissue engineered heterogeneous deproteinized bone doped with lithium. Sci. Rep. 2021, 11, 4819. [Google Scholar] [CrossRef]
- Ferretti, M.; Cavani, F.; Smargiassi, A.; Roli, L.; Palumbo, C. Mineral and Skeletal Homeostasis Influence the Manner of Bone Loss in Metabolic Osteoporosis due to Calcium-Deprived Diet in Different Sites of Rat Vertebra and Femur. BioMed Res. Int. 2015, 2015, 304178. [Google Scholar] [CrossRef] [PubMed]
- Yeom, H.; Blanchard, S.; Kim, S.; Zunt, S.; Chu, T.-M.G. Correlation between micro-computed tomography and histomorphometry for assessment of new bone formation in a calvarial experimental model. J. Craniofac. Surg. 2008, 19, 446–452. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrera-Rodríguez, S.E.; García-Márquez, E.; Padilla-Camberos, E.; Espinosa-Andrews, H. Evaluation of an Ionic Calcium Fiber Supplement and Its Impact on Bone Health Preservation in a Dietary Calcium Deficiency Mice Model. Nutrients 2022, 14, 422. https://doi.org/10.3390/nu14030422
Herrera-Rodríguez SE, García-Márquez E, Padilla-Camberos E, Espinosa-Andrews H. Evaluation of an Ionic Calcium Fiber Supplement and Its Impact on Bone Health Preservation in a Dietary Calcium Deficiency Mice Model. Nutrients. 2022; 14(3):422. https://doi.org/10.3390/nu14030422
Chicago/Turabian StyleHerrera-Rodríguez, Sara Elisa, Eristeo García-Márquez, Eduardo Padilla-Camberos, and Hugo Espinosa-Andrews. 2022. "Evaluation of an Ionic Calcium Fiber Supplement and Its Impact on Bone Health Preservation in a Dietary Calcium Deficiency Mice Model" Nutrients 14, no. 3: 422. https://doi.org/10.3390/nu14030422
APA StyleHerrera-Rodríguez, S. E., García-Márquez, E., Padilla-Camberos, E., & Espinosa-Andrews, H. (2022). Evaluation of an Ionic Calcium Fiber Supplement and Its Impact on Bone Health Preservation in a Dietary Calcium Deficiency Mice Model. Nutrients, 14(3), 422. https://doi.org/10.3390/nu14030422