
Naringenin and Quercetin Exert Contradictory Cytoprotective and Cytotoxic Effects on Tamoxifen-Induced Apoptosis in HepG2 Cells


Abstract
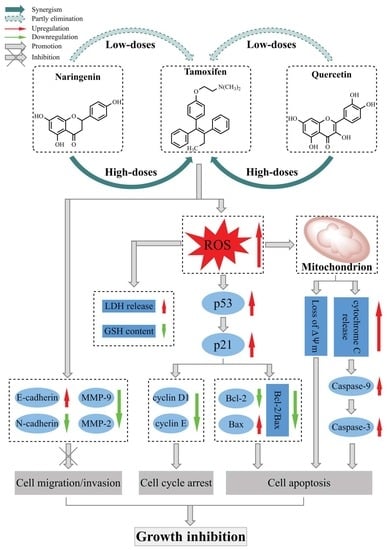
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Biological and Chemical Materials
2.2. Cell Culture and Chemical Treatment
2.3. Cell Proliferation and Cytotoxicity Evaluation
2.4. Wound Healing Assay
2.5. Transwell Assay
2.6. Cell Cycle and Apoptosis Assessment Using Flow Cytometry
2.7. Fluorescence Microscopy
2.8. Determination of LDH Leakage, GSH Content and Caspase Activity
2.9. ROS Determination
2.10. ΔΨm Determination
2.11. RNA Extraction and RT-qPCR
2.12. Statistical Analysis
3. Results
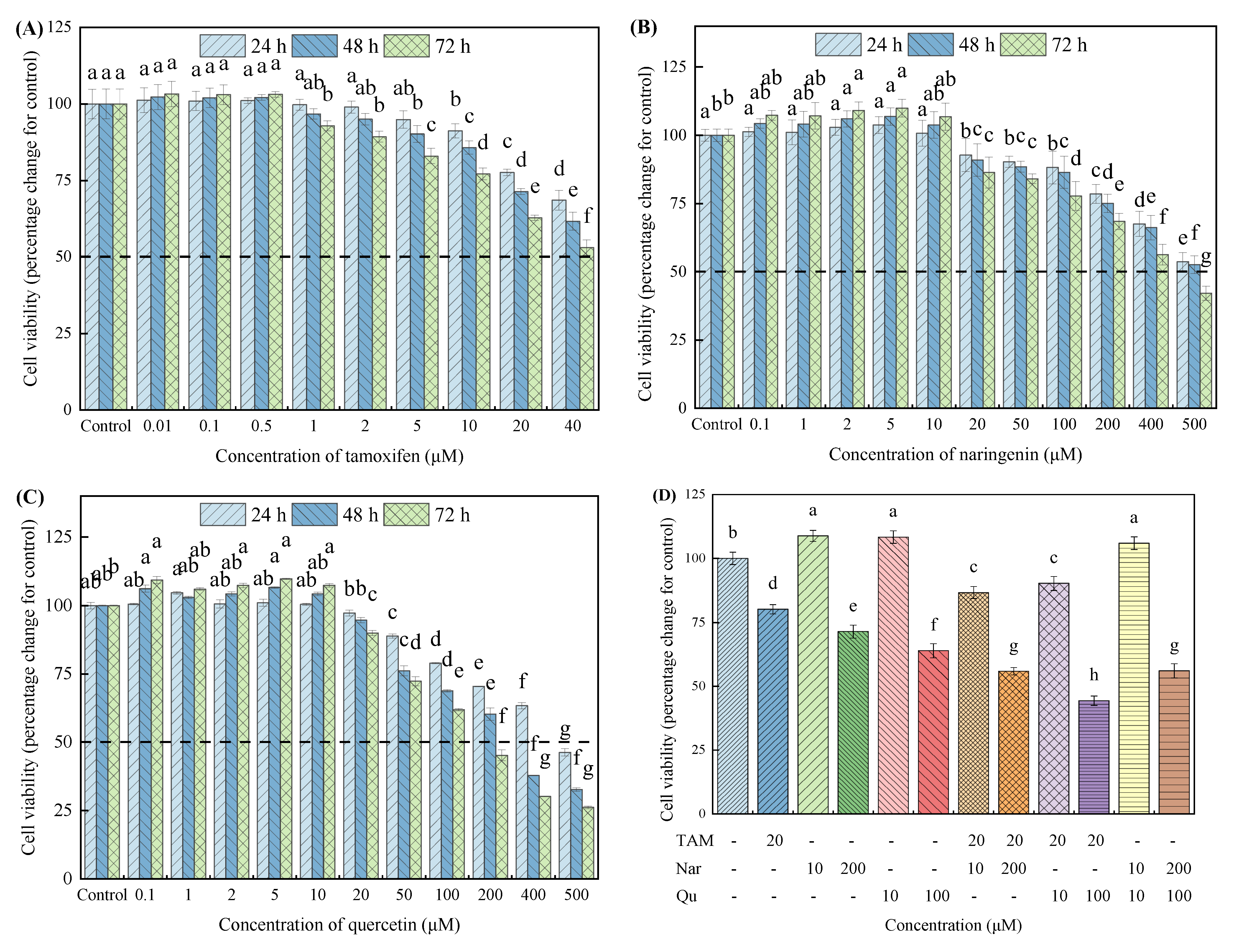
3.1. Tamoxifen, Naringenin and Quercetin Induced Morphological Changes and Regulated Cell Viability
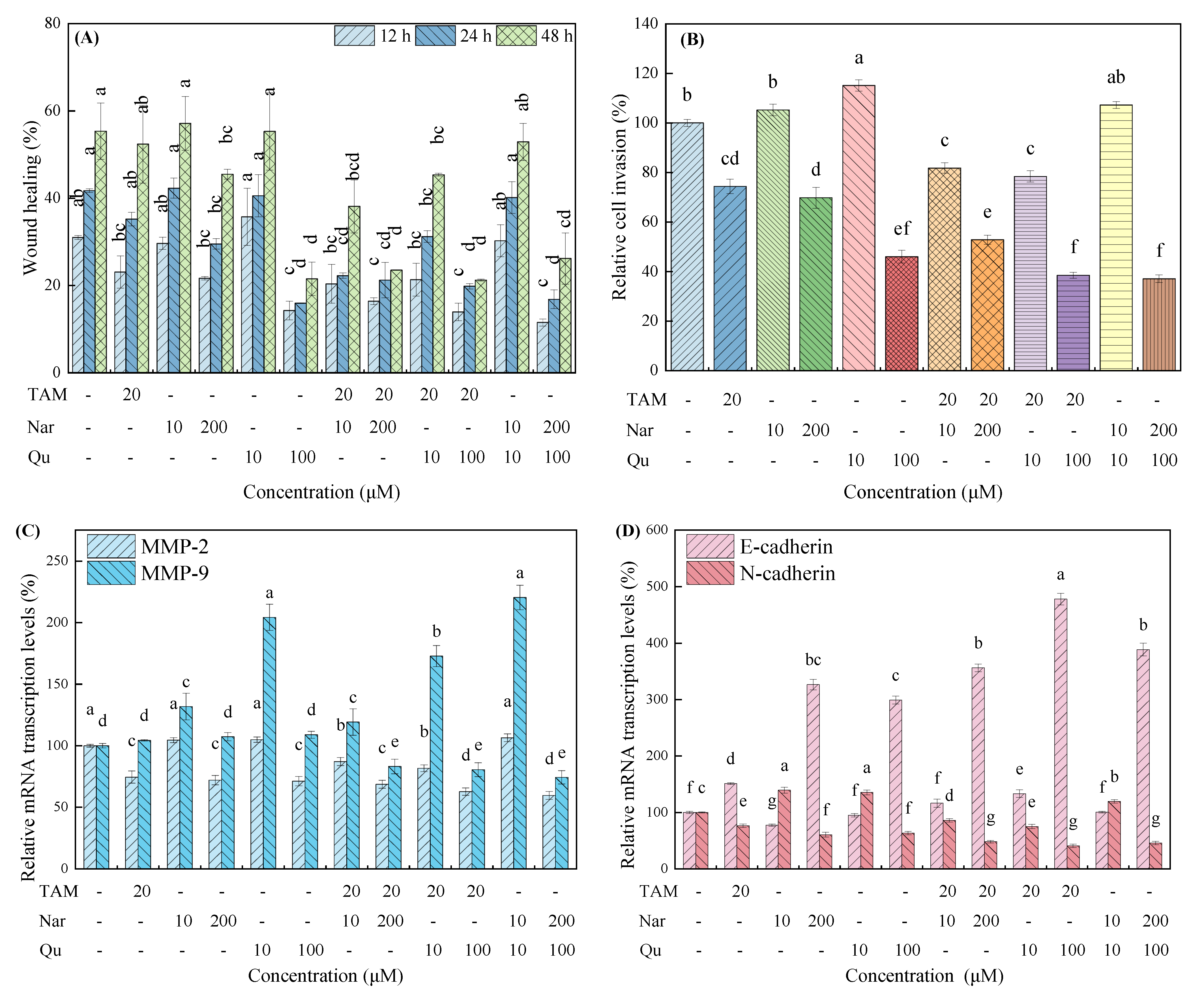
3.2. Tamoxifen, Naringenin and Quercetin Regulated Cell Migration and Invasion
3.3. Tamoxifen, Naringenin and Quercetin Regulated Cell Cycle Progression
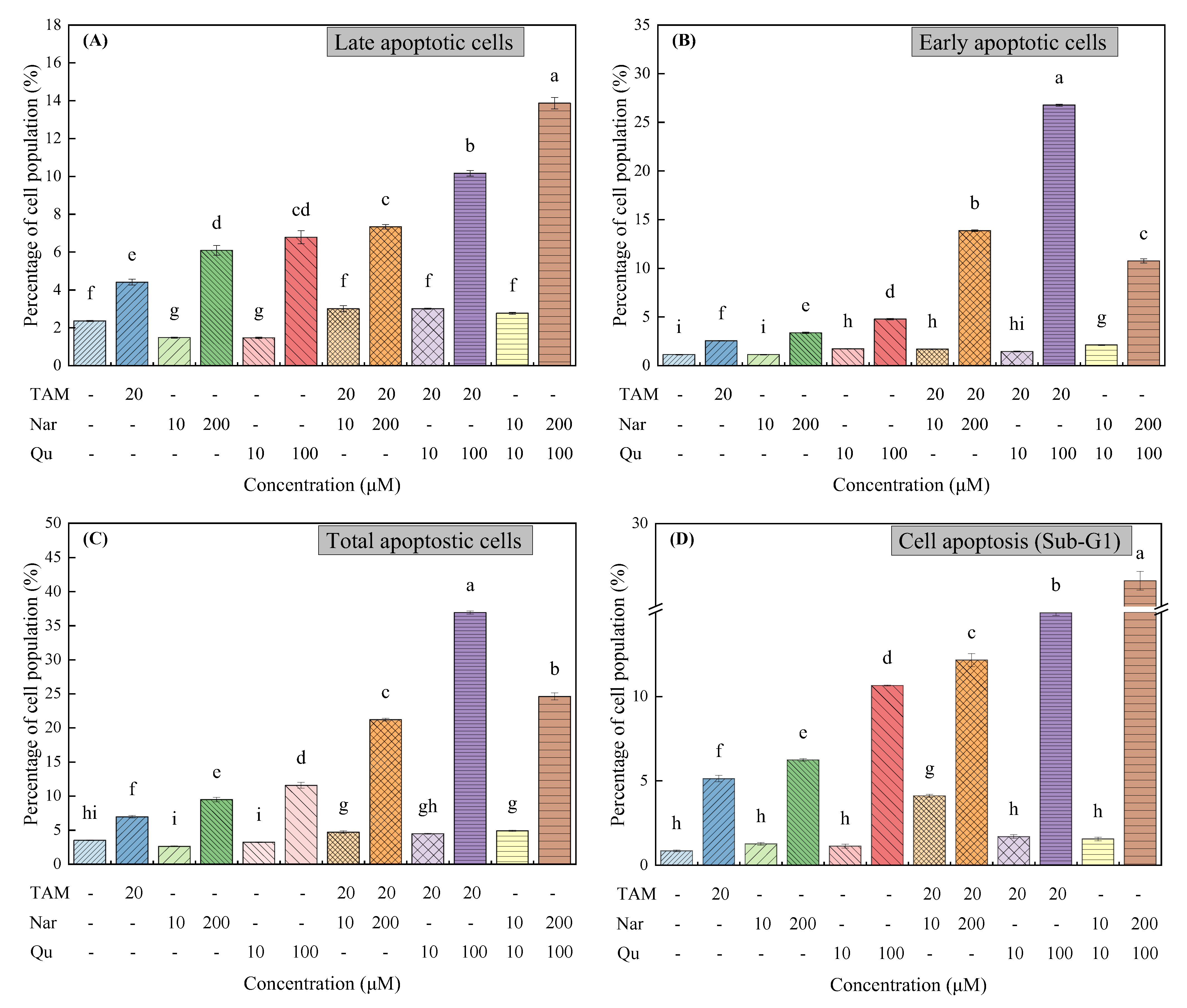
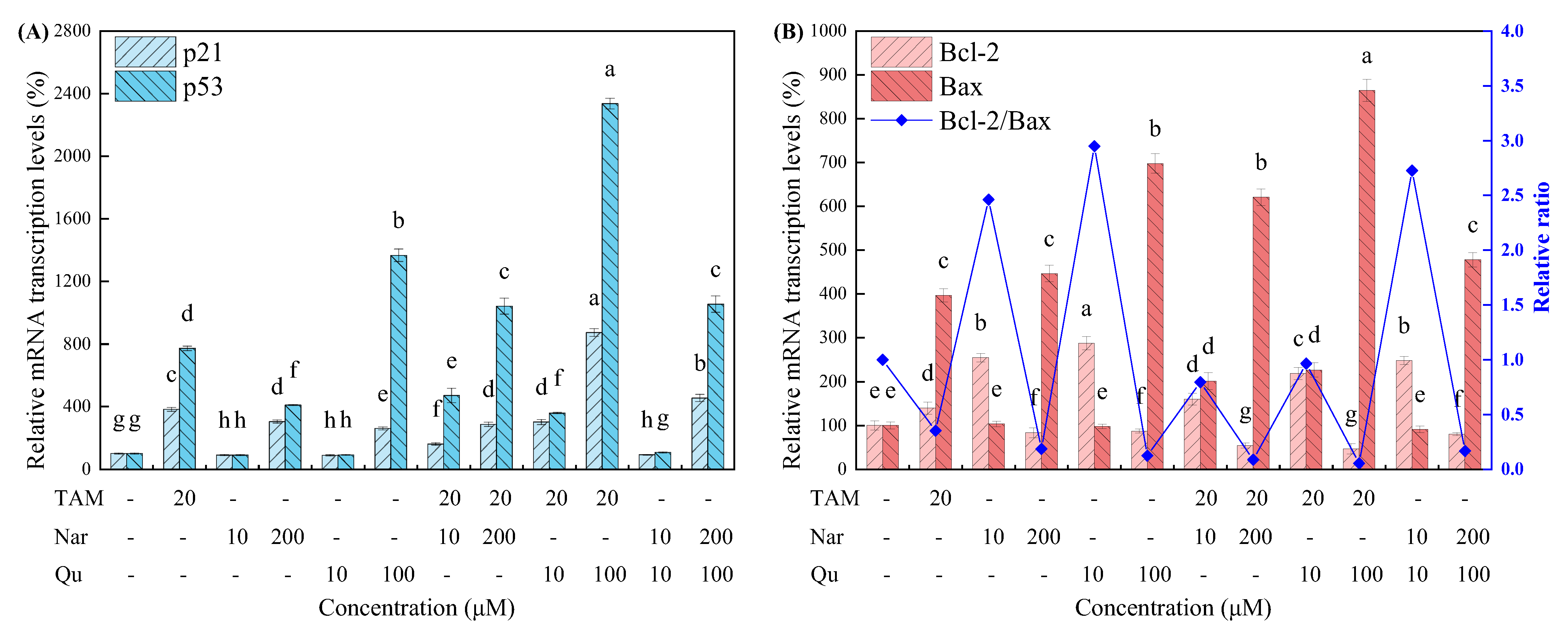
3.4. Tamoxifen, Naringenin and Quercetin Regulated Cell Apoptosis
3.5. Tamoxifen, Naringenin and Quercetin Regulated the Leakage of LDH and Glutathione Content
3.6. Tamoxifen, Naringenin and Quercetin Regulated the Activation of Caspases
3.7. Tamoxifen, Naringenin and Quercetin Regulated ΔΨm in HepG2 Cells
3.8. Tamoxifen, Naringenin and Quercetin Regulated ROS Generation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hejazi, I.I.; Khanam, R.; Mehdi, S.H.; Bhat, A.R.; Rizvi, M.M.A.; Thakur, S.C.; Athar, F. Antioxidative and anti-proliferative potential of Curculigo orchioides Gaertn in oxidative stress induced cytotoxicity: In vitro, ex vivo and in silico studies. Food Chem. Toxicol. 2018, 115, 244–259. [Google Scholar] [CrossRef] [PubMed]
- Weng, C.J.; Yen, G.C. Flavonoids, a ubiquitous dietary phenolic subclass, exert extensive in vitro anti-invasive and in vivo anti-metastatic activities. Cancer Metastasis Rev. 2012, 31, 323–351. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kang, J.W.; Kim, M.S.; Bak, Y.; Park, Y.S.; Jung, K.Y.; Lim, Y.H.; Yoon, D.Y. The apoptotic effects of the flavonoid N101-2 in human cervical cancer cells. Toxicol. Vitr. 2012, 26, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Marino, M.; Pellegrini, M.; La Rosa, P.; Acconcia, F. Susceptibility of estrogen receptor rapid responses to xenoestrogens: Physiological outcomes. Steroids 2012, 77, 910–917. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, R.G.; Liberati, A.; Angiolini, C.; Pagliaro, L. Treatment of hepatocellular carcinoma: A systematic review of randomized controlled trials. Ann. Oncol. 1997, 8, 117–136. [Google Scholar] [CrossRef]
- Di Bisceglie, A.M.; Osmack, P.; Brunt, E.M. Chemoprevention of hepatocellular carcinoma: Use of tamoxifen in an animal model of hepatocarcinogenesis. J. Lab. Clin. Med. 2005, 145, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Nichols, H.B.; DeRoo, L.A.; Scharf, D.R.; Sandler, D.P. Risk-benefit profiles of women using tamoxifen for chemoprevention. J. Natl. Cancer Inst. 2015, 107, dju354. [Google Scholar] [CrossRef]
- Brandt, S.; Heller, H.; Schuster, K.D.; Grote, J. Tamoxifen induces suppression of cell viability and apoptosis in the human hepatoblastoma cell line HepG2 via down-regulation of telomerase activity. Liver Int. 2004, 24, 46–54. [Google Scholar] [CrossRef]
- Chow, P.K. Tamoxifen does not improve overall survival in people with advanced-stage hepatocellular carcinoma. Cancer Treat. Rev. 2005, 31, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Villa, E.; Colantoni, A.; Grottola, A.; Ferretti, I.; Buttafoco, P.; Bertani, H.; De Maria, N.; Manenti, F. Variant estrogen receptors and their role in liver disease. Mol. Cell. Endocrinol. 2002, 193, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Kalra, M.; Mayes, J.; Assefa, S.; Kaul, A.K.; Kaul, R. Role of sex steroid receptors in pathobiology of hepatocellular carcinoma. World J. Gastroenterol. 2008, 14, 5945–5961. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.T.; Chang, C.W.; Wei, R.J.; Wang, S.N. Progesterone and related compounds in hepatocellular carcinoma: Basic and clinical aspects. Biomed. Res. Int. 2013, 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Huang, B.; Liu, J.; Wu, X.; Luo, N.; Wang, X.; Zheng, X.; Pan, X. Combinatorial anti-proliferative effects of tamoxifen and naringenin: The role of four estrogen receptor subtypes. Toxicology 2018, 410, 231–246. [Google Scholar] [CrossRef]
- Zhou, J.; Fang, L.; Liao, J.; Li, L.; Yao, W.; Xiong, Z.; Zhou, X. Investigation of the anti-cancer effect of quercetin on HepG2 cells in vivo. PLoS ONE 2017, 12, e0172838. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Huang, Z.; Shu, Y.; Jin, S.; Ge, H. Tamoxifen inhibits proliferation and induces apoptosis in human hepatocellular carcinoma cell line HepG2 via down-regulation of survivin expression. Biomed. Pharmacother. 2009, 63, 375–379. [Google Scholar] [CrossRef]
- Arul, D.; Subramanian, P. Naringenin (citrus flavonone) induces growth inhibition, cell cycle arrest and apoptosis in human hepatocellular carcinoma cells. Pathol. Oncol. Res. 2013, 19, 763–770. [Google Scholar] [CrossRef]
- Ahamad, M.S.; Siddiqui, S.; Jafri, A.; Ahmad, S.; Afzal, M.; Arshad, M. Induction of apoptosis and antiproliferative activity of naringenin in human epidermoid carcinoma cell through ROS generation and cell cycle arrest. PLoS ONE 2014, 9, e110003. [Google Scholar] [CrossRef]
- Tan, J.; Wang, B.; Zhu, L. Regulation of Survivin and Bcl-2 in HepG2 Cell Apoptosis Induced by Quercetin. Chem. Biodivers. 2009, 6, 1101–1110. [Google Scholar] [CrossRef]
- Xu, Z.; Liu, J.; Jianxin, C.; Yongliang, Z.; Pan, X. 17β-Estradiol inhibits testosterone-induced cell proliferation in HepG2 by modulating the relative ratios of 3 estrogen receptor isoforms to the androgen receptor. J. Cell. Biochem. 2018, 119, 8659–8671. [Google Scholar] [CrossRef]
- Xin, L.; Wang, J.; Zhang, L.W.; Che, B.; Dong, G.; Fan, G.; Cheng, K. Development of HSPA1A promoter-driven luciferase reporter gene assays in human cells for assessing the oxidative damage induced by silver nanoparticles. Toxicol. Appl. Pharmacol. 2016, 304, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Pinti, M.; Troiano, L.; Nasi, M.; Ferraresi, R.; Dobrucki, J.; Cossarizza, A. Hepatoma HepG2 cells as a model for in vitro studies on mitochondrial toxicity of antiviral drugs: Which correlation with the patient? J. Biol. Regul. Homeost. Agents 2003, 17, 166–171. [Google Scholar] [PubMed]
- Xu, D.; Li, L.; Liu, L.; Dong, H.; Deng, Q.; Yang, X.; Song, E.; Song, Y. Polychlorinated biphenyl quinone induces mitochondrial-mediated and caspase-dependent apoptosis in HepG2 cells. Environ. Toxicol. 2015, 30, 1063–1072. [Google Scholar] [CrossRef]
- Ke, N.; Wang, X.; Xu, X.; Abassi, Y.A. The xCELLigence system for real-time and label-free monitoring of cell viability. Methods Mol. Biol. 2011, 740, 33–43. [Google Scholar] [PubMed]
- Grada, A.; Otero-Vinas, M.; Prieto-Castrillo, F.; Obagi, Z.; Falanga, V. Research techniques made simple: Analysis of collective cell migration using the wound healing assay. J. Investig. Dermatol. 2017, 137, e11. [Google Scholar] [CrossRef]
- Guo, M.; Suo, Y.; Gao, Q.; Du, H.; Zeng, W.; Wang, Y.; Hu, X.; Jiang, X. The protective mechanism of Ginkgolides and Ginkgo flavonoids on the TNF-α induced apoptosis of rat hippocampal neurons and its mechanisms in vitro. Heliyon 2015, 1, e00020. [Google Scholar] [CrossRef]
- Lei, B.; Xu, J.; Peng, W.; Wen, Y.; Zeng, X.; Yu, Z.; Wang, Y.; Chen, T. In vitro profiling of toxicity and endocrine disrupting effects of bisphenol analogues by employing MCF-7 cells and two-hybrid yeast bioassay. Environ. Toxicol. 2017, 32, 278–289. [Google Scholar] [CrossRef]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Guo, R.; Wang, T.; Shen, H.; Ge, H.M.; Sun, J.; Huang, Z.H.; Shu, Y.Q. Involvement of mTOR and survivin inhibition in tamoxifen-induced apoptosis in human hepatoblastoma cell line HepG2. Biomed. Pharmacother. 2010, 64, 249–253. [Google Scholar] [CrossRef]
- Ebersohn, K.; Coetzee, P.; Venter, E.H. An improved method for determining virucidal efficacy of a chemical disinfectant using an electrical impedance assay. J. Virol. Methods 2014, 199, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Stewart, Z.A.; Westfall, M.D.; Pietenpol, J.A. Cell-cycle dysregulation and anticancer therapy. Trends Pharmacol. Sci. 2003, 24, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, Y.; Zhang, L.; Chen, L.; Du, Y.; Ye, T.; Shi, X. Naringenin exerts anti-angiogenic effects in human endothelial cells: Involvement of ERRα/VEGF/KDR signaling pathway. Fitoterapia 2016, 111, 78–86. [Google Scholar] [CrossRef]
- Maji, S.; Panda, S.; Samal, S.K.; Shriwas, O.; Rath, R.; Pellecchia, M.; Emdad, L.; Das, S.K.; Fisher, P.B.; Dash, R. Chapter Three—Bcl-2 Antiapoptotic Family Proteins and Chemoresistance in Cancer. In Advances in Cancer Research; Tew, K.D., Fisher, P.B., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 37–75. [Google Scholar]
- Tanigawa, S.; Fujii, M.; Hou, D.X. Stabilization of p53 is involved in quercetin-induced cell cycle arrest and apoptosis in HepG2 cells. Biosci. Biotechnol. Biochem. 2008, 72, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.Y.; Park, C.; Lee, J.H.; Chung, K.T.; Kwon, T.K.; Kim, G.Y.; Choi, B.T.; Choi, Y.H. Naringenin-induced apoptosis is attenuated by Bcl-2 but restored by the small molecule Bcl-2 inhibitor, HA 14-1, in human leukemia U937 cells. Toxicol. In Vitro 2009, 23, 259–265. [Google Scholar] [CrossRef]
- Wójtowicz, A.K.; Szychowski, K.A.; Wnuk, A.; Kajta, M. Dibutyl phthalate (DBP)-induced apoptosis and neurotoxicity are mediated via the aryl hydrocarbon receptor (AhR) but not by estrogen receptor alpha (ERalpha), estrogen receptor beta (ERbeta), or peroxisome proliferator-activated receptor gamma (PPARgamma) in mouse cortical neurons. Neurotox. Res. 2017, 31, 77–89. [Google Scholar]
- Holder, A.L.; Goth-Goldstein, R.; Lucas, D.; Koshland, C.P. Particle-induced artifacts in the MTT and LDH viability assays. Chem. Res. Toxicol. 2012, 25, 1885–1892. [Google Scholar] [CrossRef]
- Cunha-Oliveira, T.; Ferreira, L.L.; Coelho, A.R.; Deus, C.M.; Oliveira, P.J. Doxorubicin triggers bioenergetic failure and p53 activation in mouse stem cell-derived cardiomyocytes. Toxicol. Appl. Pharmacol. 2018, 348, 1–13. [Google Scholar] [CrossRef]
- Taylor, W.R.; Stark, G.R. Regulation of the G2/M transition by p53. Oncogene 2001, 20, 1803–1815. [Google Scholar] [CrossRef]
- Hatkevich, T.; Ramos, J.; Santos-Sanchez, I.; Patel, Y.M. A naringenin-tamoxifen combination impairs cell proliferation and survival of MCF-7 breast cancer cells. Exp. Cell Res. 2014, 327, 331–339. [Google Scholar] [CrossRef]
- Adams, J.M.; Cory, S. The Bcl-2 protein family: Arbiters of cell survival. Science 1998, 281, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.M.; Jung, C.-H.; Kim, J.; Hwang, S.-G.; Park, J.K.; Um, H.-D. The p53/p21 complex regulates cancer cell invasion and apoptosis by targeting Bcl-2 family proteins. Cancer Res. 2017, 77, 3092–3100. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Fu, J.; Yin, X.; Qu, C.; Yang, C.; He, H.; Ni, J. Induction of apoptosis in HepaRG cell line by aloe-emodin through generation of reactive oxygen species and the mitochondrial pathway. Cell. Physiol. Biochem. 2017, 42, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Ola, M.S.; Nawaz, M.; Ahsan, H. Role of Bcl-2 family proteins and caspases in the regulation of apoptosis. Mol. Cell. Biochem. 2011, 351, 41–58. [Google Scholar] [CrossRef]
- Liou, G.-Y.; Storz, P. Reactive oxygen species in cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef]
- Park, H.J.; Choi, Y.J.; Lee, J.H.; Nam, M.J. Naringenin causes ASK1-induced apoptosis via reactive oxygen species in human pancreatic cancer cells. Food Chem. Toxicol. 2017, 99, 1–8. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Z.; Jia, Y.; Liu, J.; Ren, X.; Yang, X.; Xia, X.; Pan, X. Naringenin and Quercetin Exert Contradictory Cytoprotective and Cytotoxic Effects on Tamoxifen-Induced Apoptosis in HepG2 Cells. Nutrients 2022, 14, 5394. https://doi.org/10.3390/nu14245394
Xu Z, Jia Y, Liu J, Ren X, Yang X, Xia X, Pan X. Naringenin and Quercetin Exert Contradictory Cytoprotective and Cytotoxic Effects on Tamoxifen-Induced Apoptosis in HepG2 Cells. Nutrients. 2022; 14(24):5394. https://doi.org/10.3390/nu14245394
Chicago/Turabian StyleXu, Zhixiang, Yue Jia, Jun Liu, Xiaomin Ren, Xiaoxia Yang, Xueshan Xia, and Xuejun Pan. 2022. "Naringenin and Quercetin Exert Contradictory Cytoprotective and Cytotoxic Effects on Tamoxifen-Induced Apoptosis in HepG2 Cells" Nutrients 14, no. 24: 5394. https://doi.org/10.3390/nu14245394
APA StyleXu, Z., Jia, Y., Liu, J., Ren, X., Yang, X., Xia, X., & Pan, X. (2022). Naringenin and Quercetin Exert Contradictory Cytoprotective and Cytotoxic Effects on Tamoxifen-Induced Apoptosis in HepG2 Cells. Nutrients, 14(24), 5394. https://doi.org/10.3390/nu14245394