1. Introduction
Obesity has become a chronic and global epidemic disease because it is intimately related to a chronic low-grade inflammatory state and even exacerbates obesity-related diseases, particularly metabolic dysfunctions [
1]. In patients with obesity, adipose tissue releases adipokines that trigger and maintain chronic low-grade inflammation and increase oxidative stress in the adipose tissue [
2]. Controlling the progression of obesity and reducing weight have become important strategies in the treatment of numerous diseases, particularly metabolic diseases. Therefore, it is essential to develop safe and effective weight loss drugs to reduce the morbidity and mortality of various chronic diseases resulting from obesity.
White adipose tissue (WAT) is the primary organ for energy storage. WAT plays a crucial role in regulating lipid distribution and energy homeostasis in the body and is involved in the development of obesity. Adipose tissue is closely linked to metabolic health through the development of adipocyte hypertrophy and hyperplasia in which hypertrophic growth is a critical component in triggering adipose tissue dysfunction. Once energy intake exceeds consumption, the excess energy will be stored in the adipocytes in the form of triglycerides, leading to hypertrophic expansion. Acetyl-CoA carboxylase (ACC), an essential enzyme in the synthesis of fatty acids from acetyl-CoA, provides a substrate for triglyceride synthesis in adipocytes. Considering that phosphorylated acetyl-CoA carboxylase (p-ACC) loses catalytic capacity, promoting ACC phosphorylation could inhibit the fatty acid synthesis and reduce substrates required for triglyceride synthesis [
3]. In contrast to lipogenesis, the triglycerides in adipocytes break down and release free glycerol and fatty acids, providing a substrate source for hepatic gluconeogenesis and oxidative catabolism. Adipocyte triglyceride lipase (ATGL) and phosphorylated hormone-sensitive lipase (p-HSL) are critical in initiating and catalyzing the triglyceride breakdown step wisely. Therefore, promoting the function of lipolytic enzymes may increase triglyceride catabolism in adipocytes and reduce lipid accumulation.
Adipose tissue is also an endocrine organ capable of secreting a wide range of cytokines that are closely linked to a range of obesity-induced chronic diseases [
4]. Weight gain and obesity often result in the increased secretion of inflammatory factors and chemokines, including tumor necrosis factor alpha (TNFα), interleukin 6 (IL6), and monocyte chemoattractant protein-1 (MCP-1). These inflammatory factors and chemokines induce the accumulation of inflammatory cells such as pro-inflammatory M1-type macrophages in the adipose tissue [
5]. They also trigger persistent chronic low-grade inflammation, particularly in hypertrophic abdominal adipose tissue. Reducing adipose inflammation in patients with obesity has been explored as a potential therapeutic strategy to prevent obesity-related metabolic and vascular complications [
6,
7].
Ramulus Mori (Sangzhi) alkaloid (SZ-A) tablets are derived from
Morus alba L. (mulberry twig) and approved for the treatment of Type 2 diabetes (T2DM) in China (approval number Z20200002). In a multicenter, randomized, double-blind clinical trial, SZ-A exhibited excellent hypoglycaemic effect in the treatment of T2DM, with minimal gastrointestinal- side effects like flatulence during treatment [
8]. Previous studies have reported that SZ-A could restore diabetic β-cells, improve insulin resistance, and modulate the gut microbiota and intestinal barrier integrity in KKAy mice with signs of obesity and diabetes [
9,
10]. Additionally, it has been reported that SZ-A could alleviate non-alcoholic fatty liver disease (NAFLD) in high-fat diet (HFD)-induced obese mice [
11]. It was also demonstrated that SZ-A could inhibit body weight gain in HFD mice in a dose-dependent manner, with SZ-A at a dose of 400 mg/kg having the most significant effect. In addition, Gao et al. reported that SZ-A treatment showed anti-inflammatory effects in vitro by blocking p38 MAPK, ERK, and JNK signaling pathways in macrophages. Owing to the synergistic effect of SZ-A, its anti-inflammatory effect was higher than that of its components [
12]. The analysis of tissue distribution results suggested a high distribution of SZ-A in adipose tissue after oral administration [
13]. These results indicate the potential modulating effects of SZ-A on the adipose tissue.
In this study, we aimed to investigate the effects of SZ-A on adipose tissue metabolism and inflammation development in an HFD-fed mouse model and assessed whether SZ-A can be potentially used in treating lipid metabolism-related diseases from these results.
2. Materials and Methods
2.1. Chemical and Reagents
SZ-A powder was friendly presented by Beijing Wehand-Bio Pharmaceutical Co., Ltd. (Beijing, China). β-actin(4970), ACC(3676), p-ACC(11818), HSL(18381), p-HSL(4139), ATGL(2439), F4/80(70076), and CD86(19589) primary antibodies were purchased from Cell Signaling Technology (Danvers, MA, USA). The PPARα primary antibody (ab126285) was obtained from Abcam (Cambridge, MA, USA). Horseradish peroxidase (HRP)-conjugated anti-rabbit secondary antibodies were obtained from Zhong Shan-Golden Bridge Biological Technology Co., Ltd. (Beijing, China).
2.2. Animals Experimental and Treatment
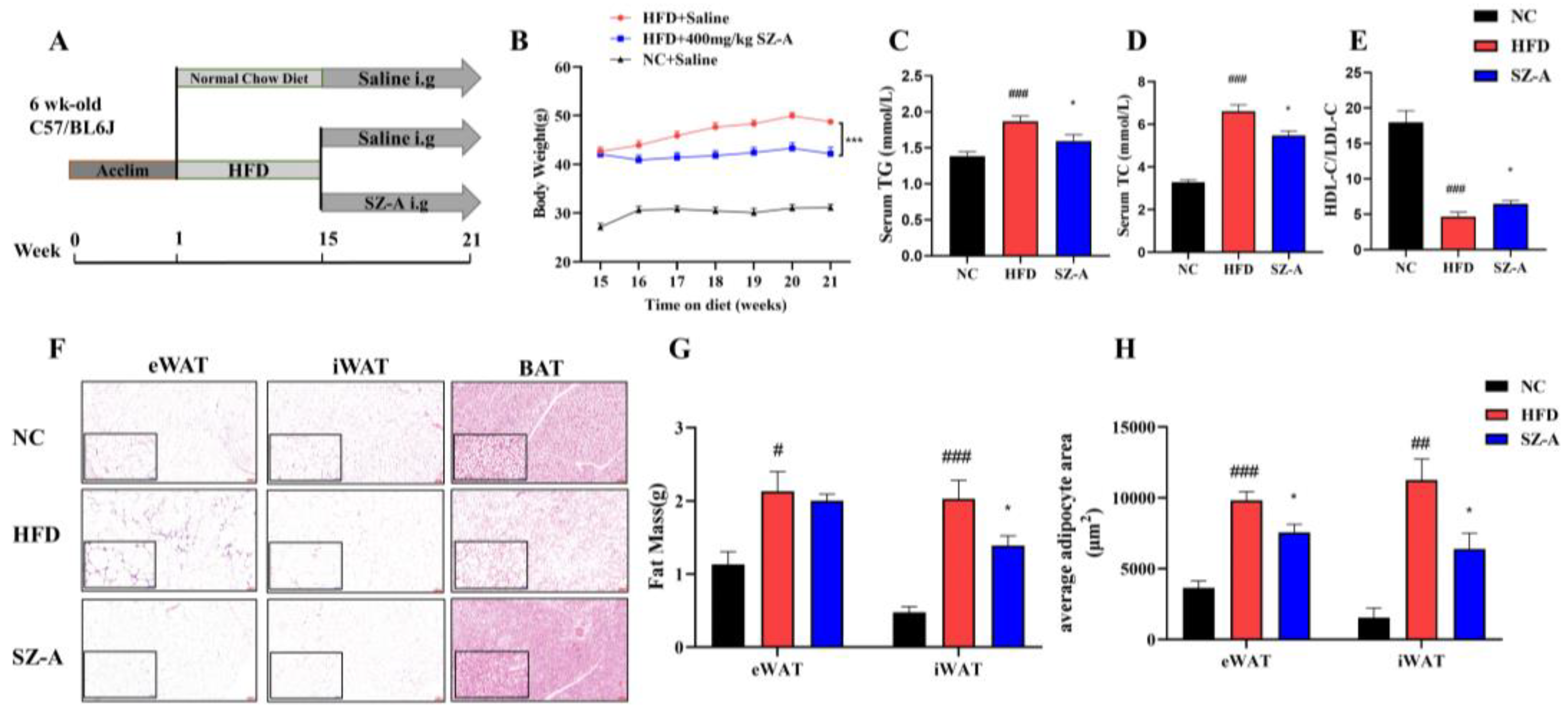
The Ethical review of experimental animal welfare was approved by the Beijing Laboratory Animal Research Center (ethical code: 2021050). The mice were housed at 24 °C with a 12 h light/dark cycle. Food and water were freely available. After 1 week of acclimatization to the environment, six-week-old male C57BL/6J mice were randomly divided into the following three groups: a normal control group (NC, n = 10), an HFD control group subjected to intragastric administration of saline (HFD, n = 10), and the SZ-A treatment group subjected to intragastric administration of SZ-A (SZ-A, n = 10). The NC was fed a maintenance feed, whereas the HFD groups were fed with 60 kcal% fat (cat. no. D12492; Research Diets Inc.) for 14 weeks. Subsequently, the mice were intragastrically administered either saline or SZ-A at a dose of 400 mg/kg/d for another 6 weeks. Mice in the HFD and SZ-A treatment groups were kept feeding an HFD until the end of the experiment. Weighed weekly during whole the experiment period. At the end of the experiment, the mice were fasted overnight from 17:00 the day before the autopsy until 9:00 the next day and then euthanized. Blood was collected by removing eyeballs, allowed to stand for 30 min at room temperature, and then centrifuged at 3000 rpm at 4 °C for 15 min to separate the serum. Adipose tissues, including epididymal adipose tissue (eWAT), inguinal adipose tissue (iWAT), and brown adipose tissue (BAT), were dissected, weighed, and kept in liquid nitrogen for 30 min. Finally, the tissues were stored at −80 °C.
2.3. Serum Biochemical Parameters
Serum triglycerides (TGs), total cholesterol (TC), low-density lipoprotein cholesterol (LDL-C), and high-density lipoprotein cholesterol (HDL-C) levels were determined using an automatic biochemistry analyzer (TOSHIBA TBA-40FR, Tokyo, Japan) with appropriate commercial assay kits from Biosino Biotechnology and Science, Inc. (Beijing, China). Serum levels of tumor necrosis factor alpha (TNFα), interleukin 6 (IL6), fibrinogen activator inhibitor-1 (PAI-1), angiopoietin-2 (Ang-2), and leptin (LEP) were determined using a mouse Multifactor test kit (R&D Systems, Minneapolis, MN, USA). Serum adiponectin (ADPN) levels were determined using a mouse adiponectin ELISA kit (Abcam, Cambridge, MA, USA) following the manufacturer’s instructions.
2.4. Histopathological Evaluation and Immunohistochemistry Assay of the Adipose Tissue
Fresh eWAT, iWAT, and BAT from mice in the NC, HFD, and SZ-A groups were fixed in 4% paraformaldehyde solution, embedded in paraffin wax, and subsequently cut into 5-µm sections for hematoxylin-eosin (H&E) staining. For immunohistochemistry analysis, paraffin-embedded sections were dehydrated, antigen repaired, and incubated overnight at 4 °C with diluted primary antibody. Subsequently, the sections were rinsed in PBS and cultivated with a secondary antibody for 1 h at room temperature. This was followed by color development using ready-to-use diaminobenzidine (DAB) solution. F4/80 and CD86 were used to label macrophage and M1-type macrophage in adipose tissue, respectively. Histological images were taken using a microscope imager (Cytation5, Biotek, Winooski, VT, USA) at 100× and 400× magnification.
2.5. RNA Sequencing and Data Analysis
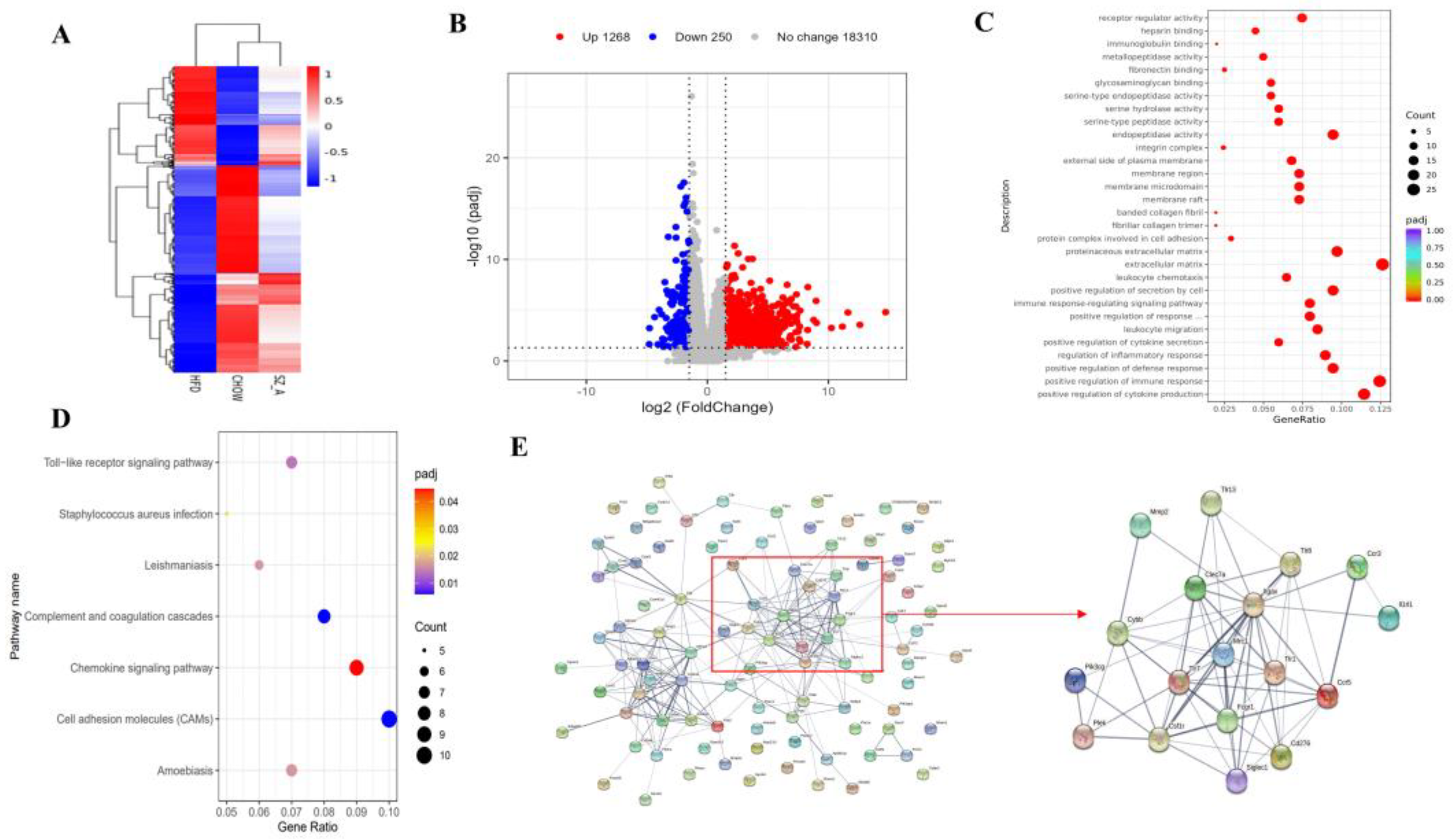
RNA extraction and sequencing analyses were performed by Novogene (Beijing, China). Briefly, isolating the total RNA from eWAT using TRIzol. The total amounts and integrity of RNA were assessed using the RNA Nano 6000 Assay Kit of the Bioanalyzer2100 system (Agilent Technologies, Santa Clara, CA, USA). Then, the extracted mRNA was purified from total RNA using poly-T oligo-attached magnetic bead, followed by fragmentation. The library fragments were purified with the AMPure XP system (Beckman Coulter, Beverly, MA, USA). The PCR product was purified by AMPure XP beads. Finally, the library was obtained. The Agilent Bioanalyzer 2100 system and qRT-PCR were used to assess the library quality. The library was sequenced using the Illumina Novaseq platform, and 150 bp paired-end reads were generated after cluster generation. RNA sequencing data were uploaded to the Gene Expression Omnibus (GEO) under accession number (GSE214618).
HISAT2 (version 2.0.5) software and paired-end clean reads were used to align the high-quality reads to the Ensemble mouse (mm10/GRCm38) reference genome. Differential expression analysis was performed by the DESeq2 R package (1.20.0), and the resulting
p-values were adjusted using Benjamini and Hochberg’s approach to control the false discovery rate. Transcripts with a padj < 0.05 and |log2(foldchange)| > 1.5 were set as the threshold for differentially expressed genes (DEGs). Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) function enrichment analyses of DEGs were implemented by Novogene (Beijing, China). Furthermore, an analysis of interactions among the top 100 DEGs (based on experimental and curated databases) and functional enrichment analysis (concerning biological processes, cell components, UniProt keywords, and KEGG or Reactome pathways) was performed according to the STRING database (
https://string-db.org; version 11.5, accessed on 31 August 2022).
2.6. Western Blot
The epididymal adipose tissue samples were homogenized in ice-cold lysis buffer containing complete tablets, mini EASYpack (Roche, Basel, Switzerland), and 1 mmol/L phenylmethanesulfonyl fluoride (Solarbio, Beijing, China) for 30 min. The homogenate was centrifuged at 4 °C with a speed of 12,000× g for 15 min and the supernatant was collected. Protein concentration was determined using the BCA assay (Thermo Fisher Scientific, Waltham, MA, USA). Next, the proteins were separated using 12% SDS-PAGE and transferred to a polyvinylidene difluoride membrane. The membranes were blocked with 5% skimmed milk in tris-buffered saline with Tween 20 (TBST) for 2 h at room temperature, then incubated with the following primary antibodies: β-actin (1:1000), p-ACC (1:1000), ACC (1:1000), p-HSL (1:1000), HSL (1:1000), PPARα (1:1000), and ATGL (1:1000) overnight at 4 °C. Subsequently, the membranes were rinsed using TBST three times and incubated with HRP-conjugated anti-rabbit secondary antibody (1:5000) for 1 h at room temperature. The protein expression bands were detected using enhanced chemiluminescence (Easybio, Beijing, China) and visualized using a Tanon-4600SF chemiluminescence imager (Tanon Science & Technology Co., Ltd., Shanghai, China).
2.7. Real-Time Polymerase Chain Reaction Analysis
Total RNA was isolated from eWAT using the TRIzol reagent (Invitrogen, CA, USA) and the RNA was quantified using a Nano-300 Micro-Spectrophotometer (AllSheng, Hangzhou, China). cDNA was synthesized from 1 µg of RNA using a reverse transcription system (Promega, Madison, WI, USA), following the manufacturer’s instructions. Real-time RT-PCR was performed using a protocol consisting of 40 cycles of 30 s at 95 °C, 10 s at 95 °C, and 30 s at 60 °C. Melting curve analysis was used to determine the purity of the PCR products. The 2−∆∆CT method was used to calculate the relative expression of each gene. The expression level of each gene was normalized to that of the internal control-
Ppia. The used primers in this experiment are listed in
Table S1 in the Supplementary Information.
2.8. Software and Statistical Analysis
The relative protein expressions, adipocyte area, and positive area were quantified using ImageJ software (NIH, Bethesda, MD, USA). Graphs were generated using Prism 8 software (GraphPad Software Inc., San Diego, CA, USA). Data are expressed as mean ± standard error of the mean (SEM). Multiple groups were compared using one-way or two-way analysis of variance (ANOVA) followed by Tukey’s test, depending on the experiment. The two experimental groups were evaluated using a t-test. Statistical significance was defined as p < 0.05.
4. Discussion
It has been verified that obesity can cause chronic low-grade inflammation and increase the risk of systemic metabolic dysfunction associated with obesity-linked disorders. In addition to being an energy storage organ, the adipose tissue is also a critical endocrine organ, which can release various bioactive substances with pro-inflammatory or anti-inflammatory activities, such as adipose-derived secreted factors and adipokines. SZ-A, an extract of Morus alba L., was approved to treat T2DM in 2020. Besides, SZ-A also showed potential in decreasing weight gain in HFD-mice and in vitro anti-inflammatory. In this study, mice were fed with HFD for 14 weeks to create an obesity model. The most effective dose was selected at 400 mg/kg SZ-A, and this dose was used to conduct an in-depth study of the effects of SZ-A on the regulation of lipid metabolism and inflammation of adipose tissue.
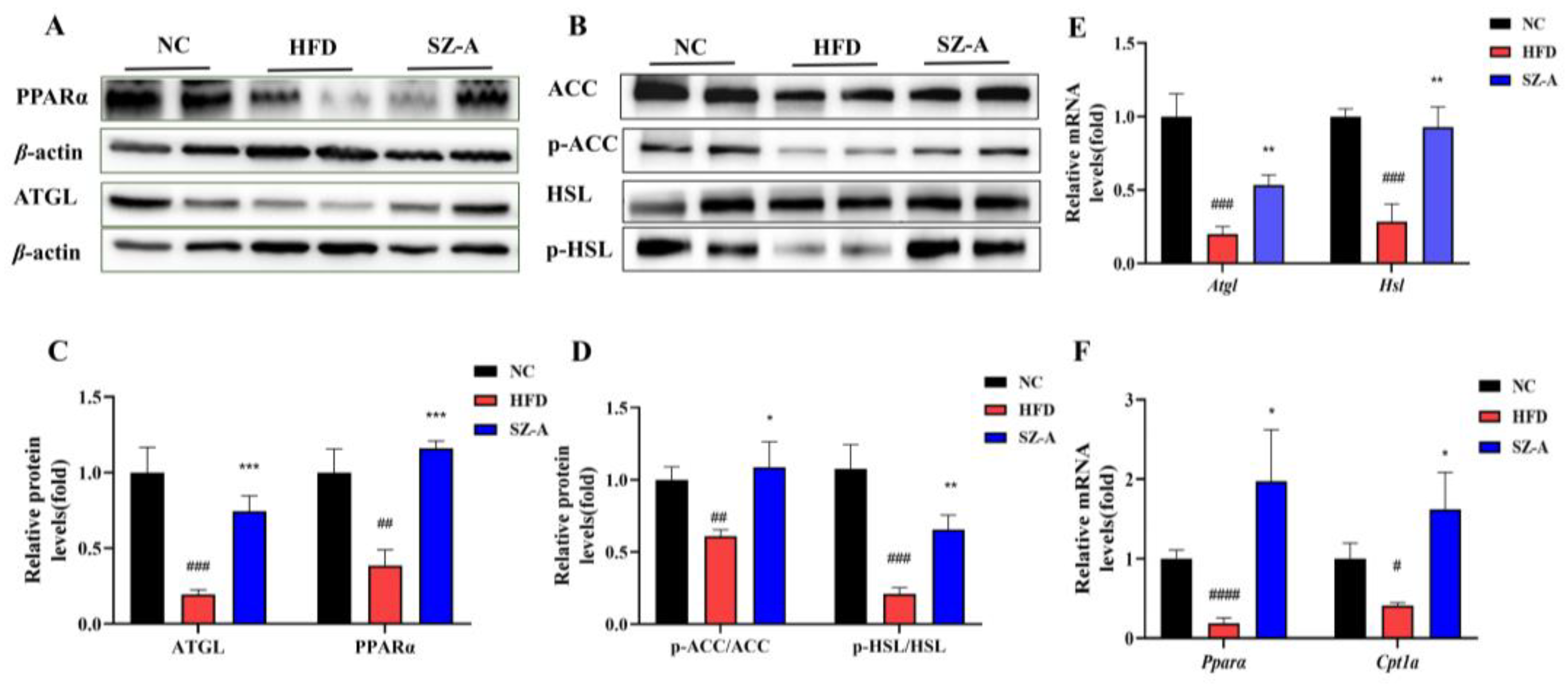
Firstly, the adipose tissue of HFD-induced obese mice in pathological sections showed that SZ-A significantly reduced the size of adipocytes in both WAT and BAT. It is well known that central obesity is dangerous to human health [
14]. Therefore, in this study, we selected the eWAT (abdominal adipose representative adipose tissue) from obese mice for lipid metabolism-related analysis. Our findings suggest that SZ-A inhibits the catalytic capacity of ACC in epididymal adipose tissue by promoting its phosphorylation, thereby inhibiting the de novo synthesis of fatty acids. As the core enzyme that catalyzes lipolysis in adipocytes, ATGL specifically catalyzes triglyceride catabolism, whereas HSL is primarily responsible for catalyzing diglyceride catabolism. It has been reported that lipolysis is reduced in ATGL-deficient mice, and lipid accumulation occurs in almost all tissues throughout the body [
15]. The catalytic activity of HSL is primarily related to its phosphorylation site. Phosphorylation of the Ser563 site has been found to stimulate HSL activity and may enhance its catalytic activity by promoting the translocation of HSL to lipid droplets [
16]. In this study, SZ-A significantly promoted the protein expression of ATGL, promoted the phosphorylation of HSL (Ser563), and increased the mRNA levels of both proteins. In addition, SZ-A promoted the protein expression and transcription of
Pparα as well as the transcriptional level of
Cpt1a. PPARα is a transcription factor that belongs to the member of the nuclear hormone receptor family and is a fatty-acid sensor. Activation of PPARα increases the catabolic oxidation of fatty acids and facilitates the transport of long-chain fatty acids to mitochondria via CPT1A, which in turn increases the fatty acids’ oxidative catabolism [
17]. Accordingly, the results of this study suggest that SZ-A reduces fatty acid synthesis at its source as well as enhances fatty acid catabolism and oxidation by promoting the breakdown of triglycerides and diglycerides in eWAT, thereby reducing lipid accumulation, achieving weight loss, and improving lipid metabolism.
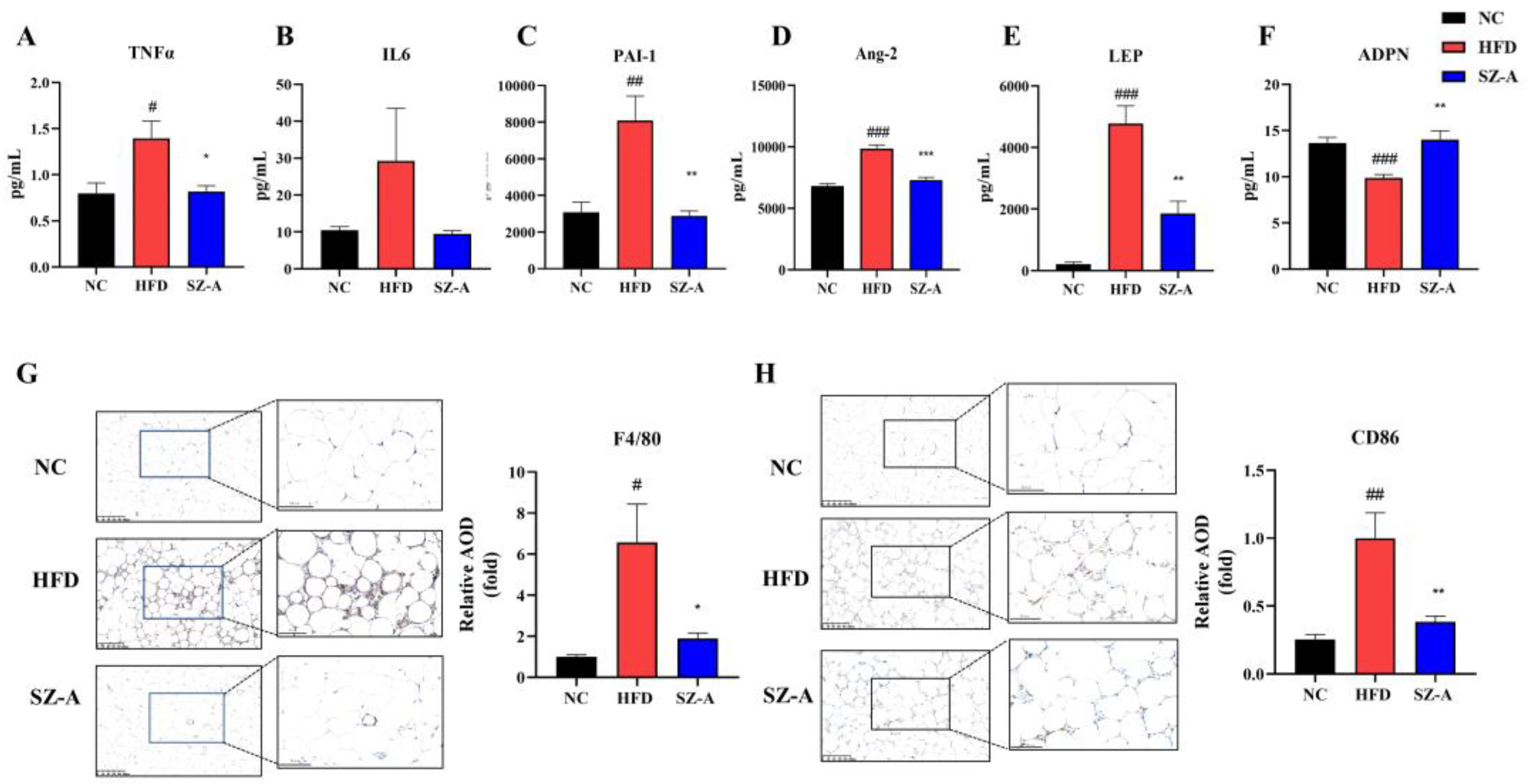
As a critical endocrine organ, the adipose tissue secretes many adipokines and inflammatory factors. Obesity triggers chronic inflammation, which is a sign of adipose tissue dysfunction and metabolic dysregulation. TNF-α and IL6 are two major pro-inflammatory factors in adipose tissue, and they are usually expressed at elevated levels in the serum and adipose tissue of obese individuals and involved in obesity-related systemic insulin resistance [
4]. Compared with the non-obese population, PAI1 and Ang-2 levels were obviously higher in the obese population. PAI-1 is a member of the serine protease inhibitor family and plays an important regulatory part in the fibrinolytic process and thrombosis. Although the liver is the primary source of PAI-1, some studies have demonstrated that visceral fat in obese people is the predominant cause of elevated PAI-1 levels. Moreover, the elevated PAI-1 expression triggers cardiovascular diseases, such as atherosclerosis [
18]. Ang-2 is mainly secreted by adipose tissue, and while getting obese, its level would be increased. Ang-2 plays a substantial role in the development of obesity-type hypertension and insulin resistance [
19]. Clinical data show that serum LEP levels are elevated and ADPN levels are reduced in obese individuals. LEP primarily acts on the central nervous system and regulates energy metabolism by suppressing appetite. Circulating LEP levels are proportionally correlated with fat mass, and obese individuals have higher levels of LEP but no anorexic response, a phenomenon known as leptin resistance [
20]. Furthermore, LEP has pro-inflammatory activity, and its elevated levels are associated with insulin resistance and liver fibrosis in obese individuals [
21]. To date, ADPN is an adipokine that is negatively associated with obesity, but its levels are higher in healthy people who have gained weight and are lower in a metabolically disturbed state of obesity. Increased ADPN expression is expected to increase insulin sensitivity and reduce cardiovascular risk in obese people [
22]. In this study, the results of the serum multi-factor assay showed that the SZ-A-treated group showed noteworthy lower serum levels of TNFα, IL6, PAI1, Ang-2, and LEP and higher serum expression levels of ADPN compared to the HFD group. SZ-A exhibits excellent in ameliorating HFD-induced organismal and adipose tissue inflammation in obese mice.
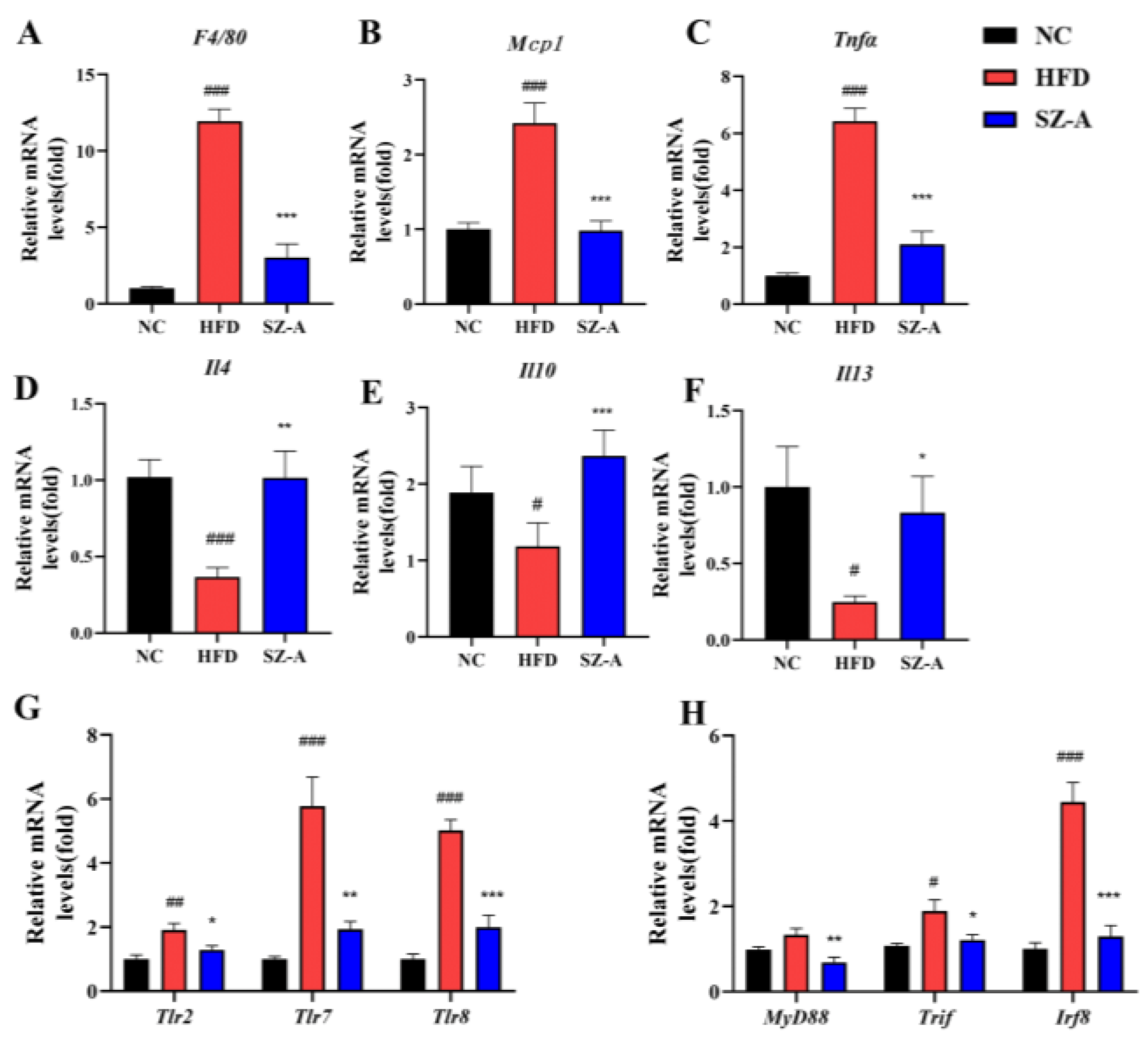
Obesity usually induces macrophage accumulation and inflammation in adipose tissues. Adipocytes in obese people secrete more pro-inflammatory and chemotactic factors, causing large numbers of macrophages to recruit into adipose tissue and triggering persistent low-grade chronic inflammation. Low-grade chronic inflammation is always affiliated with chronic diseases, such as insulin resistance, NAFLD, and cardiovascular pathology. The development of adipose tissue inflammation is largely dependent on inflammatory cells, such as macrophages recruited to the adipose tissue, particularly pro-inflammatory M1-type macrophages. M1-type macrophages have been reported to be the major generator of pro-inflammatory cytokines and involve dead or dying adipocytes in adipose tissue, modeling a typical coronal structure [
23]. The results of the eWAT transcriptome analysis in mice display that SZ-A has a regulatory role in inflammatory genes and pathways in epididymal fat. Therefore, we examined inflammatory factors and cells in the eWAT of HFD-induced obese mice. The results of the immunohistochemical analysis showed that SZ-A treatment could remarkably reduce the counts of both total macrophages and M1-type cells. SZ-A treatment significantly down-regulated the transcription of the pro-inflammatory factors
Tnfα and
Mcp1, which may be related to the reduction of the count of macrophages in adipose tissue after SZ-A treatment. This further suggested that SZ-A achieved the amelioration of improving adipose tissue inflammation by reducing macrophage infiltration in adipose tissue, which in turn significantly inhibited the expression of pro-inflammatory factors
Tnfα and
Mcp1 and up-regulated the gene transcription of anti-inflammatory factors
Il4,
Il10, and
Il13.Numerous studies have confirmed that Toll-like pattern recognition receptors play an essential role in mediating the proinflammatory effects of free fatty acids [
24]. In this study, transcriptome analysis showed that SZ-A could down-regulate the transcription of genes in the Toll-like receptor pathway. TLRs are innate immune pattern recognition receptors expressed in different cells and tissues. Upon binding to ligands, TLRs undergo conformational changes, forming homo- or heterodimers that subsequently recruit bridging proteins, such as MyD88 and TIRF, and initiate intracellular signaling cascades. In adipose tissue, the TLR signaling pathway is important for regulating immune responses [
25]. Activation of the TLR2 signaling pathway has been reported to be associated with chronic inflammation, which triggers the development of insulin resistance in adipocytes. TLR2 expression is increased in the muscle and adipose tissue of HFD mice, and TLR2 gene silencing improves insulin resistance in these mice. This implies that TLR2 is an important regulator of the inflammatory and metabolic pathways of HFD-induced obesity [
26]. TLR7 and TLR8 are expressed in adipocyte endosomes and lysosomes, respectively. They are up-regulated in the adipose tissue and promote metabolic inflammation in obese individuals [
27,
28]. In this study, the RT-PCR results of eWAT indicated that SZ-A significantly inhibited the transcription of
Tlr2,
Tlr7, and
Tlr8 and their downstream bridging proteins, including
MyD88,
Tirf, and
Irf8. Therefore, SZ-A may have a regulatory effect on the Toll-like receptor pathway, and we speculated that the reduction in TLR signaling may be due to a decrease in adipose tissue infiltrated macrophages.
5. Conclusions
In this study, SZ-A (six weeks of treatment) improved lipid metabolism and inhibited weight gain in HFD-induced obese mice by inhibiting fatty acid synthase and increasing lipolytic enzyme expression to inhibit fat accumulation. In addition, SZ-A was seen to have a beneficial effect on obesity-induced chronic inflammation of adipose tissue. SZ-A can reduce the expression of pro-inflammatory factors such as PAI1, Ang-2, TNFα, IL6, and LEP; reduce the recruitment of macrophages, especially M1 macrophages; inhibit the transcription of Tlr2, Tlr7, Tlr8, and their downstream bridging proteins MyD88, Tirf, and Irf8; and up-regulate the expression of anti-inflammatory factors Il4, Il10, and Il13 at the level of mRNA.
Thus, SZ-A, a natural anti-diabetic agent, has been shown to be effective in improving disorders of lipid metabolism and adipose tissue inflammation in HFD-induced obese mice.



 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




