
PPARγ Gene as a Possible Link between Acquired and Congenital Lipodystrophy and its Modulation by Dietary Fatty Acids

 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
- Initial research on the most common types of lipodystrophies and the main genes involved in the development of congenital lipodystrophy as well as the genes implicated in acquired lipodystrophy.
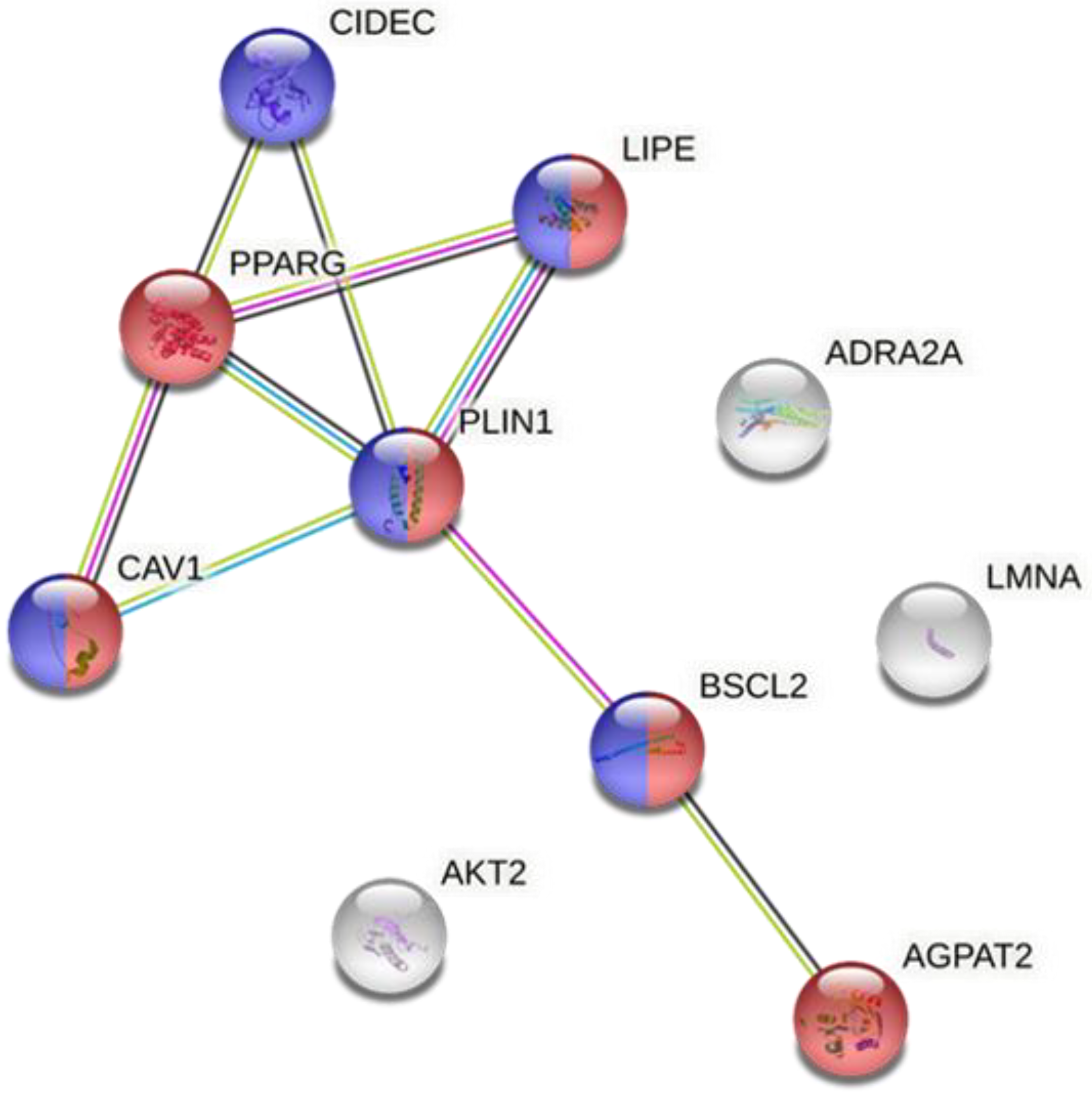
- Analysis of the proteins involved in lipodystrophy via the Search Tool for the Retrieval of Interacting Proteins 11 (STRING 11) (a database under a Creative Commons by 4.0’ license) with a minimum required interaction score (high confidence (0.700)) [7]. This analysis was performed to determine the interaction between the different proteins implicated in lipodystrophy and to identify which ones played a more relevant role and the main biological processes in which they were involved. In addition, it allowed us to determine the relationship between the implicated proteins in congenital and acquired lipodystrophy.
- A systematic review of the scientific evidence on the modulation of the expression and activity of the selected genes (PPARγ and Perilipin 1 (PLIN1)) by dietary lipids. The PubMed database was searched from January 2017 to January 2022. Due to the lack of studies analyzing the impact of diet on lipodystrophy, the following search strategy was used: ((PPARG) OR (peroxisome proliferator-activated receptor-gamma) OR (PLIN1) OR (PERILIPIN1)) AND ((oil) OR (fatty acid) OR (high-fat diet) OR (dietary lipid) OR (capric acid) OR (lauric acid) OR (myristic acid) OR (palmitic acid) OR (stearic acid) OR (arachidic acid) OR (behenic acid) OR (caprylic acid) OR (oleic acid) OR (linoleic acid) OR (eicosapentaenoic acid) OR (linolenic acid) OR (arachidonic acid) OR (docosatetraenoic acid) OR (palmitoleic acid)) NOT (review [Publication Type]). The inclusion criteria were as follows: (1) experimental model: cells, mice, rats and clinical trials; (2) intervention with dietary oils, fats or isolated fatty acids (i.e., conjugated linoleic, palm oil or omega-3 fatty acids); (3) analysis of PPARγ or PLIN1 gene/protein expression; (4) original papers (not reviews); (5) articles written in the English language. The eligibility for inclusion and exclusion criteria were evaluated by reading both (1) the title and abstract and (2) the full text (Supplementary Figure S1).
3. Results and Discussion
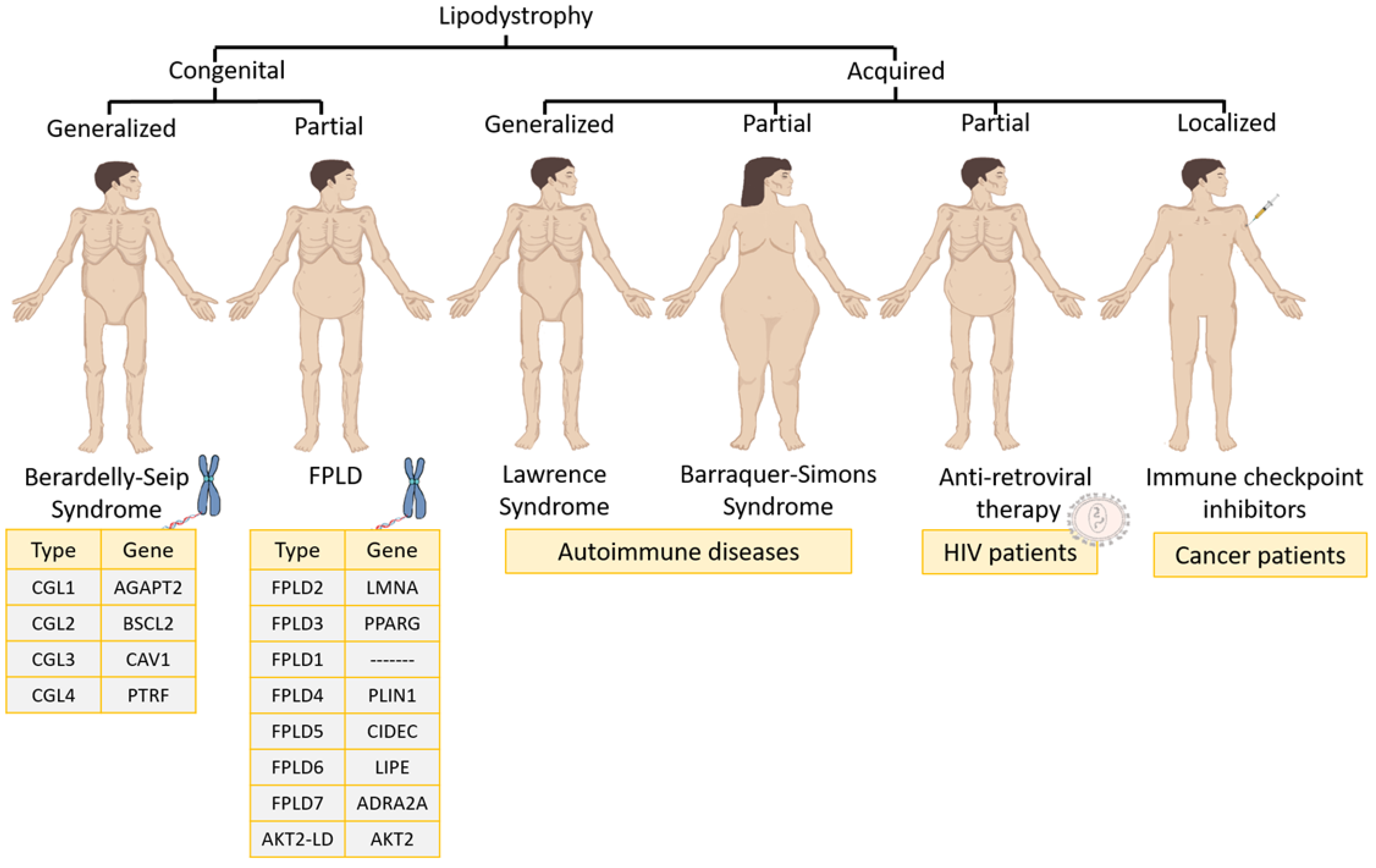
3.1. Lipodystrophy Syndromes Classification
3.1.1. Congenital Lipodystrophies
- Familial partial lipodystrophy (FPLD) is usually an autosomal dominant syndrome characterized by a selective loss of fat from the lower and upper extremities as well as the trunk [1]. During childhood, patients have a normal fat distribution but at puberty start to lose fat from the chest, anterior abdomen and extremities. There are eight varieties of FPLD: (1) FLPD2, the most common subtype (also called the Dunnigan type), which is characterized by mutations in the lamin A/C gene (LMNA); (2) FPLD3, the second most common subtype, which is based on mutations in the PPARγ gene; (3) FPLD1 (or the Kobberling type), whose genetic mutation is unknown; (4) FPDL4, which is characterized by heterozygous mutations in the PLIN1 gene; (5) FPDL5 and (6) FPDL6, both of which are autosomal recessive disorders in the cell death-inducing DFFA-like effector C (CIDEC) and lipase E (LIPE) genes, respectively; (7) FPDL7, which features a genetic mutation in the adrenoceptor alpha 2A (ADRA2A) gene; and (8) AKT2-linked lipodystrophy (AKT2-LD), which is based on a mutation in the (AKT Serine/Threonine Kinase 2) AKT2 gene [1].
- Congenital generalized lipodystrophy (CGL), or Berardinelli–Seip syndrome, is an autosomal recessive disease that is distinguished by the absence of AT both at birth and in early childhood [1]. Four distinct subtypes exist depending on the gene that is altered: (1) CGL1 is the most common subtype, and the associated altered gene is 1-Acylglycerol-3-Phosphate O-Acyltransferase 2 (AGPAT2), which is involved in triglyceride biosynthesis; (2) CGL2 is the second most common subtype and the altered gene in that case is lipid droplet biogenesis associated (BSCL2), which plays a relevant role in adipocyte differentiation and small lipid droplet fusion in adipocytes; (3) CGL3 has only been reported in one patient and the altered gene was caveolin 1 (CAV1), which translocates fatty acids to lipid droplets; (4) CGL4 has been reported in 20 patients and is very close to CGL3 because its gene (caveolae-associated protein 1) is regulated by CAV1 expression [1].
3.1.2. Acquired Lipodystrophies
- Acquired generalized lipodystrophy or Lawrence syndrome is characterized by the generalized loss of subcutaneous fat. The loss of fat usually begins in childhood or adolescence [1]. Most patients have related autoimmune diseases [8,9,10,11,12,13]. In certain autoimmune diseases, the role of the PPARγ gene is essential to modulate inflammation. In fact, therapy is based on agonists of the PPARγ gene [14].
- Acquired partial lipodystrophy, or Barraquer–Simons syndrome, is characterized by a gradual loss of subcutaneous fat from the upper trunk, upper extremities, neck and face. In the case of females, after puberty, excess fat can accumulate in the lower extremities, hips and lower abdomen. It is often related to autoimmune diseases and this syndrome affects mostly women [1,15,16,17].
- Antiretroviral therapy-induced lipodystrophy occurs in patients infected with human immunodeficiency virus who, after 2–4 years of treatment with ART, start to have an increased accumulation of both intra-abdominal and upper trunk fat, while they lose subcutaneous fat in the lower and upper extremities [1,18]. This is related to PPARγ protein downregulation. However, by stopping ART, PPARγ protein expression is restored in macrophages and adipocytes [4].
- Recent cases of acquired lipodystrophy have been associated with the use of immune checkpoint inhibitors to treat cancer [19,20,21]. Among them, childhood cancer survivors transplanted with hematopoietic stem cells and treated with chemotherapy developed acquired lipodystrophy over time [22]. Furthermore, other types of cancer, such as craniopharyngioma, may lead to chronic inflammatory demyelinating polyneuropathy together with acquired lipodystrophy [23]. This pathology is associated with certain cancer therapies against programmed cell death protein 1 (PD-1), which promotes apoptosis in antigen-specific T cells in lymph nodes and leads to adverse events of an immune nature [24].
3.2. Relationship of Genes Involved in the Development of Lipodystrophy
- BSCL2: a protein expressed mainly in AT, which is involved in lipid droplet biogenesis, in the regulation of energy homeostasis and adipocyte differentiation [25].
- LIPE or hormone-sensitive lipase, which promotes the hydrolysis of triglycerides stored in lipid droplets during adipocyte differentiation [26].
- CAV1: a protein located in lipid droplets of adipocytes which has a key role in cholesterol homeostasis, endothelial transcytosis and cellular metabolism [27].
- LMNA: a protein involved in telomere dynamics, the nuclear membrane, chromatin organization and nuclear assembly [28].
- AKT2: a kinase involved in processes such as angiogenesis, cell growth, proliferation and metabolism [29].
- ADRA2A: a receptor involved in the inhibition of adenylate cyclase induced by catecholamine [30].
- PPARγ: a nuclear receptor that controls insulin sensitivity, glucose metabolism and adipocyte differentiation. PPARγ protein is a major adipogenic factor [31].
- AGPAT2: an acyltransferase involved in the transformation of lysophosphatidic acid into phosphatidic acid, which belongs to the triglyceride biosynthetic pathway [32].
- CIDEC: a protein that modulates triglyceride storage by restricting lipolysis and is involved in the enlargement of lipid droplets [31].
- PLIN1: a modulator of the lipid metabolism in adipocytes which protects lipid droplets from breakdown by HSL, and its interaction with CIDEC promotes the enlargement of lipid droplets [33].
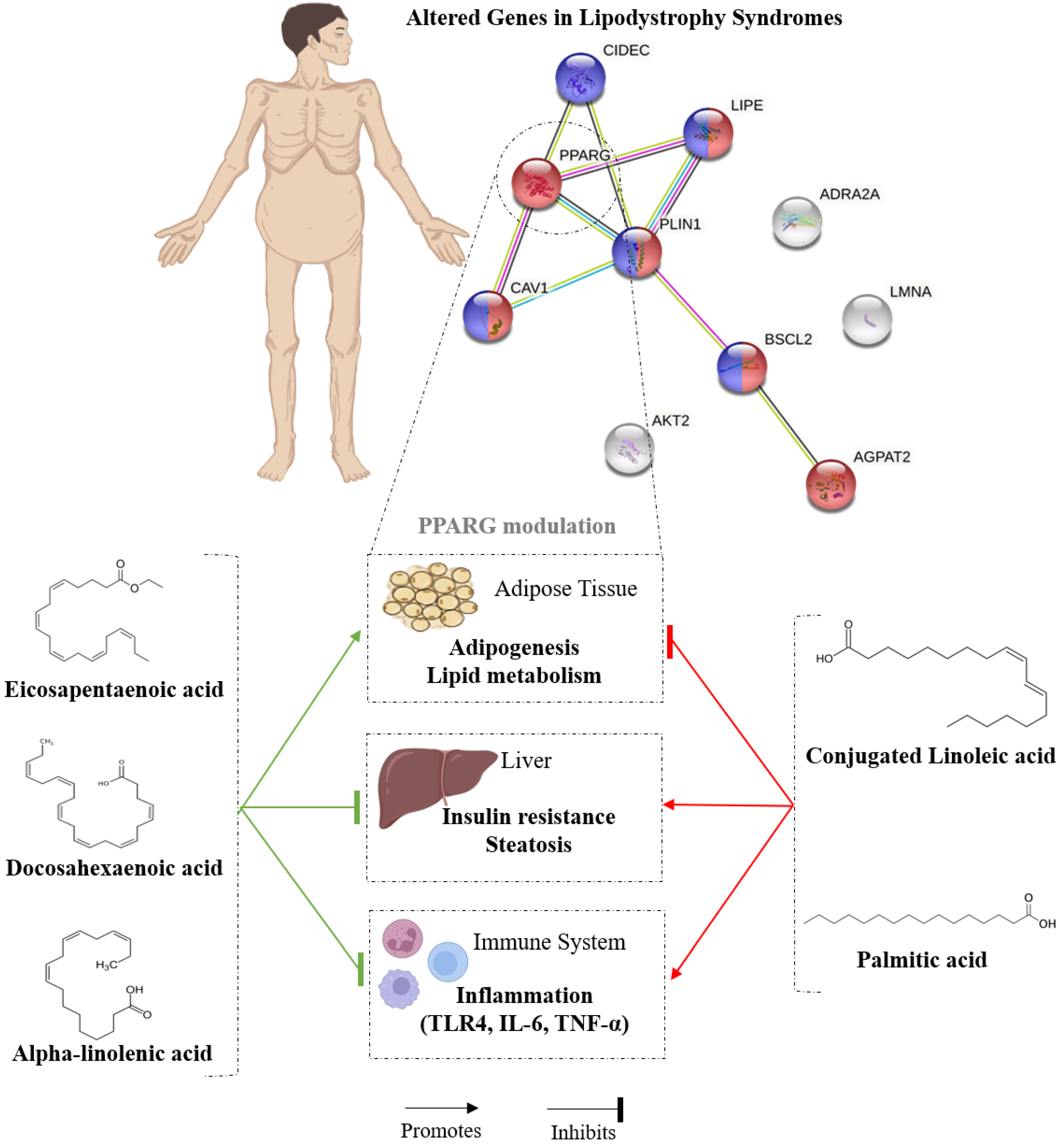
3.3. Nutrigenomic Effects of Dietary Lipids on PPARγ and PLIN1
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Hussain, I.; Garg, A. Lipodystrophy Syndromes. Endocrinol. Metab. Clin. N. Am. 2016, 45, 783–797. [Google Scholar] [CrossRef]
- Brown, R.J.; Araujo-Vilar, D.; Cheung, P.T.; Dunger, D.; Garg, A.; Jack, M.; Mungai, L.; Oral, E.A.; Patni, N.; Rother, K.I.; et al. The Diagnosis and Management of Lipodystrophy Syndromes: A Multi-Society Practice Guideline. J. Clin. Endocrinol. Metab. 2016, 101, 4500–4511. [Google Scholar] [CrossRef]
- Akinci, B.; Oral, E.A.; Neidert, A.; Rus, D.; Cheng, W.Y.; Thompson-Leduc, P.; Cheung, H.C.; Bradt, P.; Foss de Freitas, M.C.; Montenegro, R.M.; et al. Comorbidities and Survival in Patients with Lipodystrophy: An International Chart Review Study. J. Clin. Endocrinol. Metab. 2019, 104, 5120–5135. [Google Scholar] [CrossRef]
- Caron, M.; Vigouroux, C.; Bastard, J.-P.; Capeau, J. Antiretroviral-Related Adipocyte Dysfunction and Lipodystrophy in HIV-Infected Patients: Alteration of the PPARγ-Dependent Pathways. PPAR Res. 2009, 2009, 507141. [Google Scholar] [CrossRef]
- Pinel, A.; Rigaudière, J.P.; Morio, B.; Capel, F. Adipose Tissue Dysfunctions in Response to an Obesogenic Diet Are Reduced in Mice after Transgenerational Supplementation with Omega 3 Fatty Acids. Metabolites 2021, 11, 838. [Google Scholar] [CrossRef]
- Amin, K.A.; Homeida, A.M.; El Mazoudy, R.H.; Hashim, K.S.; Garelnabi, M. Dietary Lipids in Health and Disease. J. Lipids 2019, 5729498. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2020, 49, D605–D612. [Google Scholar] [CrossRef]
- Sharawat, I.K.; Yadav, J.; Dawman, L. Multiple sites acquired lipodystrophy in two siblings: A rare adverse effect of intramuscular triamcinolone. BMJ Case Rep. 2019, 12, e231017. [Google Scholar] [CrossRef]
- Corvillo, F.; Aparicio, V.; López-Lera, A.; Garrido, S.; Araújo-Vilar, D.; de Miguel, M.P.; López-Trascasa, M. Autoantibodies Against Perilipin 1 as a Cause of Acquired Generalized Lipodystrophy. Front. Immunol. 2018, 9, 2142. [Google Scholar] [CrossRef]
- Childs, C.E.; Calder, P.C.; Miles, E.A. Diet and Immune Function. Nutrients 2019, 11, 1933. [Google Scholar] [CrossRef]
- Manzel, A.; Muller, D.N.; Hafler, D.A.; Erdman, S.E.; Linker, R.A.; Kleinewietfeld, M. Role of “Western diet” in inflammatory autoimmune diseases. Curr. Allergy Asthma Rep. 2014, 14, 404. [Google Scholar] [CrossRef]
- Vieira, S.M.; Pagovich, O.E.; Kriegel, M.A. Diet, microbiota and autoimmune diseases. Lupus 2014, 23, 518–526. [Google Scholar] [CrossRef]
- Sahinoz, M.; Khairi, S.; Cuttitta, A.; Brady, G.F.; Rupani, A.; Meral, R.; Tayeh, M.K.; Thomas, P.; Riebschleger, M.; Camelo-Piragua, S.; et al. Potential association of LMNA-associated generalized lipodystrophy with juvenile dermatomyositis. Clin. Diabetes Endocrinol. 2018, 4, 6. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, J.; Luo, S.; Zhan, Y.; Lu, Q. The roles of PPARγ and its agonists in autoimmune diseases: A comprehensive review. J. Autoimmun. 2020, 113, 102510. [Google Scholar] [CrossRef]
- Corvillo, F.; Nozal, P.; López-Lera, A.; De Miguel, M.P.; Piñero-Fernández, J.A.; De Lucas, R.; García-Concepción, M.D.; Beato, M.J.; Araújo-Vilar, D.; López-Trascasa, M. Evidence of ongoing complement activation on adipose tissue from an 11-year-old girl with Barraquer–Simons syndrome. J. Dermatol. 2020, 47, 1439–1444. [Google Scholar] [CrossRef]
- Corvillo, F.; Ceccarini, G.; Nozal, P.; Magno, S.; Pelosini, C.; Garrido, S.; López-Lera, A.; Moraru, M.; Vilches, C.; Fornaciari, S.; et al. Immunological features of patients affected by Barraquer-Simons syndrome. Orphanet. J. Rare Dis. 2020, 15, 9. [Google Scholar] [CrossRef]
- Heidemann, L.N.; Thomsen, J.B.; Sørensen, J.A. Barraquer-Simons syndrome: A unique patient’s perspective on diagnosis, disease progression and recontouring treatment. BMJ Case Rep. 2016, 2016, bcr2016216134. [Google Scholar] [CrossRef]
- Akita, S.; Suzuki, K.; Yoshimoto, H.; Ohtsuru, A.; Hirano, A.; Yamashita, S. Cellular Mechanism Underlying Highly-Active or Antiretroviral Therapy-Induced Lipodystrophy: Atazanavir, a Protease Inhibitor, Compromises Adipogenic Conversion of Adipose-Derived Stem/Progenitor Cells through Accelerating ER Stress-Mediated Cell Death in Differentiating Adipocytes. Int. J. Mol. Sci. 2021, 22, 2114. [Google Scholar]
- Gnanendran, S.S.; Miller, J.A.; Archer, C.A.; Jain, S.V.; Hwang, S.J.E.; Peters, G.; Miller, A. Acquired lipodystrophy associated with immune checkpoint inhibitors. Melanoma Res. 2020, 30, 599–602. [Google Scholar] [CrossRef]
- Hosokawa, M.; Shibata, H.; Hosokawa, T.; Irie, J.; Ito, H.; Hasegawa, T. Acquired partial lipodystrophy with metabolic disease in children following hematopoietic stem cell transplantation: A report of two cases and a review of the literature. J. Pediatr. Endocrinol. Metab. 2019, 32, 537–541. [Google Scholar] [CrossRef]
- Falcao, C.K.; Cabral, M.C.S.; Mota, J.M.; Arbache, S.T.; Costa-Riquetto, A.D.; Muniz, D.Q.B.; Cury-Martins, J.; Almeida, M.Q.; Kaczemorska, P.C.; Nery, M.; et al. Acquired Lipodystrophy Associated with Nivolumab in a Patient with Advanced Renal Cell Carcinoma. J. Clin. Endocrinol. Metab. 2019, 104, 3245–3248. [Google Scholar] [CrossRef]
- Nagayama, A.; Ashida, K.; Moritaka, K.; Hidaka, M.; Gobaru, M.; Tanaka, S.; Hasuzawa, N.; Akasu, S.; Goto, Y.; Motomura, S.; et al. Metreleptin Supplementation for Improving Lipid and Glycemic Profiles in Acquired Diabetes Lipodystrophy: A Case Report. J. Endocr. Soc. 2019, 3, 2179–2183. [Google Scholar] [CrossRef]
- Lockemer, H.E.; Sumpter, K.M.; Cope-Yokoyama, S.; Garg, A. A novel paraneoplastic syndrome with acquired lipodystrophy and chronic inflammatory demyelinating polyneuropathy in an adolescent male with craniopharyngioma. J. Pediatr. Endocrinol. Metab. 2018, 31, 479–483. [Google Scholar] [CrossRef]
- Eigentler, T.; Lomberg, D.; Machann, J.; Stefan, N. Lipodystrophic Nonalcoholic Fatty Liver Disease Induced by Immune Checkpoint Blockade. Ann. Intern Med. 2020, 172, 836–837. [Google Scholar] [CrossRef]
- Zhou, H.; Xu, C.; Lee, H.; Yoon, Y.; Chen, W. Berardinelli–Seip congenital lipodystrophy 2/SEIPIN determines brown adipose tissue maintenance and thermogenic programing. Mol. Metab. 2020, 36, 100971. [Google Scholar] [CrossRef]
- Sollier, C.; Capel, E.; Aguilhon, C.; Smirnov, V.; Auclair, M.; Douillard, C.; Ladsous, M.; Defoort-Dhellemmes, S.; Gorwood, J.; Braud, L.; et al. LIPE-related lipodystrophic syndrome: Clinical features and disease modeling using adipose stem cells. Eur. J. Endocrinol. 2021, 184, 155–168. [Google Scholar] [CrossRef]
- Schrauwen, I.; Szelinger, S.; Siniard, A.L.; Kurdoglu, A.; Corneveaux, J.J.; Malenica, I.; Richholt, R.; Van Camp, G.; De Both, M.; Swaminathan, S.; et al. A Frame-Shift Mutation in CAV1 Is Associated with a Severe Neonatal Progeroid and Lipodystrophy Syndrome. PLoS ONE 2015, 10, e0131797. [Google Scholar] [CrossRef]
- Vigouroux, C.; Guénantin, A.C.; Vatier, C.; Capel, E.; Le Dour, C.; Afonso, P.; Bidault, G.; Béréziat, V.; Lascols, O.; Capeau, J.; et al. Lipodystrophic syndromes due to LMNA mutations: Recent developments on biomolecular aspects, pathophysiological hypotheses and therapeutic perspectives. Nucleus 2018, 9, 235–248. [Google Scholar] [CrossRef]
- Shearin, A.L.; Monks, B.R.; Seale, P.; Birnbaum, M.J. Lack of AKT in adipocytes causes severe lipodystrophy. Mol. Metab. 2016, 5, 472–479. [Google Scholar] [CrossRef]
- Garg, A.; Sankella, S.; Xing, C.; Agarwal, A.K. Whole-exome sequencing identifies ADRA2A mutation in atypical familial partial lipodystrophy. JCI Insight 2016, 1. [Google Scholar] [CrossRef]
- Zammouri, J.; Vatier, C.; Capel, E.; Auclair, M.; Storey-London, C.; Bismuth, E.; Mosbah, H.; Donadille, B.; Janmaat, S.; Fève, B.; et al. Molecular and Cellular Bases of Lipodystrophy Syndromes. Front. Endocrinol. 2021, 12, 803189. [Google Scholar] [CrossRef] [PubMed]
- Ceccarini, G.; Magno, S.; Pelosini, C.; Ferrari, F.; Sessa, M.R.; Scabia, G.; Maffei, M.; Jéru, I.; Lascols, O.; Vigouroux, C.; et al. Congenital Generalized Lipoatrophy (Berardinelli-Seip Syndrome) Type 1: Description of Novel AGPAT2 Homozygous Variants Showing the Highly Heterogeneous Presentation of the Disease. Front. Endocrinol. 2020, 11, 39. [Google Scholar] [CrossRef] [PubMed]
- Jéru, I.; Vantyghem, M.-C.; Bismuth, E.; Cervera, P.; Barraud, S.; Auclair, M.; Vatier, C.; Lascols, O.; Savage, D.B.; Vigouroux, C. Diagnostic Challenge in PLIN1-Associated Familial Partial Lipodystrophy. J. Clin. Endocrinol. Metab. 2019, 104, 6025–6032. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Quintela, A.; Churruca, I.; Portillo, M.P. The role of dietary fat in adipose tissue metabolism. Public Health Nutr. 2007, 10, 1126–1131. [Google Scholar] [CrossRef]
- Cheng, A.M.; Handa, P.; Tateya, S.; Schwartz, J.; Tang, C.; Mitra, P.; Oram, J.F.; Chait, A.; Kim, F. Apolipoprotein A-I attenuates palmitate-mediated NF-κB activation by reducing Toll-like receptor-4 recruitment into lipid rafts. PLoS ONE 2012, 7, e33917. [Google Scholar] [CrossRef]
- Sanon, V.P.; Handelsman, Y.; Pham, S.V.; Chilton, R. Cardiac Manifestations of Congenital Generalized Lipodystrophy. Clin. Diabetes 2016, 34, 181–186. [Google Scholar] [CrossRef]
- Sun, X.; Sun, X.; Meng, H.; Wu, J.; Guo, X.; Du, L.; Wu, H. Krill Oil Inhibits NLRP3 Inflammasome Activation in the Prevention of the Pathological Injuries of Diabetic Cardiomyopathy. Nutrients 2022, 14, 368. [Google Scholar] [CrossRef]
- Kong, S.; Cai, B.; Nie, Q. PGC-1α affects skeletal muscle and adipose tissue development by regulating mitochondrial biogenesis. Mol. Genet. Genomic. 2022, 297, 621–633. [Google Scholar] [CrossRef]
- Hashemzadeh, A.A.; Nasoohi, N.; Raygan, F.; Aghadavod, E.; Akbari, E.; Taghizadeh, M.; Memarzadeh, M.R.; Asemi, Z. Flaxseed Oil Supplementation Improve Gene Expression Levels of PPAR-γ, LP(a), IL-1 and TNF-α in Type 2 Diabetic Patients with Coronary Heart Disease. Lipids 2017, 52, 907–915. [Google Scholar] [CrossRef]
- Szalai, K.; Tempfli, K.; Zsédely, E.; Lakatos, E.; Gáspárdy, A.; Bali Papp, Á. Linseed oil supplementation affects fatty acid desaturase 2, peroxisome proliferator activated receptor gamma, and insulin-like growth factor 1 gene expression in turkeys (Meleagris gallopavo). Anim. Biosci. 2021, 34, 662–669. [Google Scholar] [CrossRef]
- Brandt, A.; Rajcic, D.; Jin, C.J.; Sánchez, V.; Engstler, A.J.; Jung, F.; Nier, A.; Baumann, A.; Bergheim, I. Fortifying diet with rapeseed oil instead of butterfat attenuates the progression of diet-induced non-alcoholic fatty liver disease (NAFLD) and impairment of glucose tolerance. Metabolism 2020, 109, 154283. [Google Scholar] [CrossRef]
- Uriho, A.; Yang, S.; Tang, X.; Liu, C.S.; Wang, S.; Cong, Y.; Zhang, J.; Zhou, P. Benefits of blended oil consumption over other sources of lipids on the cardiovascular system in obese rats. Food Funct. 2019, 10, 5290–5301. [Google Scholar] [CrossRef]
- Liu, S.H.; Chiu, C.Y.; Wang, L.P.; Chiang, M.T. Omega-3 Fatty Acids-Enriched Fish Oil Activates AMPK/PGC-1α Signaling and Prevents Obesity-Related Skeletal Muscle Wasting. Mar. Drugs 2019, 17, 380. [Google Scholar] [CrossRef] [PubMed]
- Naeini, Z.; Toupchian, O.; Vatannejad, A.; Sotoudeh, G.; Teimouri, M.; Ghorbani, M.; Nasli-Esfahani, E.; Koohdani, F. Effects of DHA-enriched fish oil on gene expression levels of p53 and NF-κB and PPAR-γ activity in PBMCs of patients with T2DM: A randomized, double-blind, clinical trial. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Hajri, T.; Zaiou, M.; Fungwe, T.V.; Ouguerram, K.; Besong, S. Epigenetic Regulation of Peroxisome Proliferator-Activated Receptor Gamma Mediates High-Fat Diet-Induced Non-Alcoholic Fatty Liver Disease. Cells 2021, 10, 1355. [Google Scholar] [CrossRef] [PubMed]
- Perfilyev, A.; Dahlman, I.; Gillberg, L.; Rosqvist, F.; Iggman, D.; Volkov, P.; Nilsson, E.; Risérus, U.; Ling, C. Impact of polyunsaturated and saturated fat overfeeding on the DNA-methylation pattern in human adipose tissue: A randomized controlled trial. Am. J. Clin. Nutr. 2017, 105, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhou, N.; Zhu, X.; Min, C.; Zhou, W.; Li, X. n-3 PUFAs protect against adiposity and fatty liver by promoting browning in postnatally overfed male rats: A role for NRG4. J. Nutr. Biochem. 2021, 93, 108628. [Google Scholar] [CrossRef]
- Zayed, E.A.; AinShoka, A.A.; El Shazly, K.A.; Abd El Latif, H.A. Improvement of insulin resistance via increase of GLUT4 and PPARγ in metabolic syndrome-induced rats treated with omega-3 fatty acid or l-carnitine. J. Biochem. Mol. Toxicol. 2018, 32, e22218. [Google Scholar] [CrossRef]
- Moradi, S.; Alivand, M.; KhajeBishak, Y.; AsghariJafarabadi, M.; Alipour, M.; Chilibeck, P.D.; Alipour, B. The effect of omega3 fatty acid supplementation on PPARγ and UCP2 expressions, resting energy expenditure, and appetite in athletes. BMC Sports Sci. Med. Rehabil. 2021, 13, 48. [Google Scholar] [CrossRef]
- Eraky, S.M.; Ramadan, N.M. Effects of omega-3 fatty acids and metformin combination on diabetic cardiomyopathy in rats through autophagic pathway. J. Nutr. Biochem. 2021, 97, 108798. [Google Scholar] [CrossRef]
- Zheng, J.S.; Chen, J.; Wang, L.; Yang, H.; Fang, L.; Yu, Y.; Yuan, L.; Feng, J.; Li, K.; Tang, J.; et al. Replication of a Gene-Diet Interaction at CD36, NOS3 and PPARG in Response to Omega-3 Fatty Acid Supplements on Blood Lipids: A Double-Blind Randomized Controlled Trial. EBioMedicine 2018, 31, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Bae, M.; Park, Y.K.; Lee, J.Y. n-3 PUFAs inhibit TGFβ1-induced profibrogenic gene expression by ameliorating the repression of PPARγ in hepatic stellate cells. J. Nutr. Biochem. 2020, 85, 108452. [Google Scholar] [CrossRef] [PubMed]
- Pandurangan, S.B.; Al-Maiman, S.A.; Al-Harbi, L.N.; Alshatwi, A.A. Beneficial Fatty Acid Ratio of Salvia hispanica L. (Chia Seed) Potentially Inhibits Adipocyte Hypertrophy, and Decreases Adipokines Expression and Inflammation in Macrophage. Foods 2020, 9, 368. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Takabatake, Y.; Minami, S.; Sakai, S.; Fujimura, R.; Takahashi, A.; Namba-Hamano, T.; Matsuda, J.; Kimura, T.; Matsui, I.; et al. Eicosapentaenoic acid attenuates renal lipotoxicity by restoring autophagic flux. Autophagy 2021, 17, 1700–1713. [Google Scholar] [CrossRef] [PubMed]
- Illesca, P.; Valenzuela, R.; Espinosa, A.; Echeverría, F.; Soto-Alarcon, S.; Campos, C.; Rodriguez, A.; Vargas, R.; Magrone, T.; Videla, L.A. Protective Effects of Eicosapentaenoic Acid Plus Hydroxytyrosol Supplementation Against White Adipose Tissue Abnormalities in Mice Fed a High-Fat Diet. Molecules 2020, 25, 4433. [Google Scholar] [CrossRef] [PubMed]
- Pahlavani, M.; Ramalingam, L.; Miller, E.K.; Scoggin, S.; Menikdiwela, K.R.; Kalupahana, N.S.; Festuccia, W.T.; Moustaid-Moussa, N. Eicosapentaenoic Acid Reduces Adiposity, Glucose Intolerance and Increases Oxygen Consumption Independently of Uncoupling Protein 1. Mol. Nutr. Food Res. 2019, 63, e1800821. [Google Scholar] [CrossRef]
- Sergi, D.; Luscombe-Marsh, N.; Heilbronn, L.K.; Birch-Machin, M.; Naumovski, N.; Lionetti, L.; Proud, C.G.; Abeywardena, M.Y.; O’Callaghan, N. The Inhibition of Metabolic Inflammation by EPA Is Associated with Enhanced Mitochondrial Fusion and Insulin Signaling in Human Primary Myotubes. J. Nutr. 2021, 151, 810–819. [Google Scholar] [CrossRef]
- Riera-Heredia, N.; Lutfi, E.; Gutiérrez, J.; Navarro, I.; Capilla, E. Fatty acids from fish or vegetable oils promote the adipogenic fate of mesenchymal stem cells derived from gilthead sea bream bone potentially through different pathways. PLoS ONE 2019, 14, e0215926. [Google Scholar] [CrossRef]
- Martins, F.F.; Aguila, M.B.; Mandarim-de-Lacerda, C.A. Eicosapentaenoic and docosapentaenoic acids lessen the expression of PPARγ/Cidec affecting adipogenesis in cultured 3T3-L1 adipocytes. Acta Histochem. 2020, 122, 151504. [Google Scholar] [CrossRef]
- Lauvai, J.; Becker, A.K.; Lehnert, K.; Schumacher, M.; Hieronimus, B.; Vetter, W.; Graeve, L. The Furan Fatty Acid 9M5 Acts as a Partial Ligand to Peroxisome Proliferator-Activated Receptor gamma and Enhances Adipogenesis in 3T3-L1 Preadipocytes. Lipids 2019, 54, 277–288. [Google Scholar] [CrossRef]
- Katsnelson, G.; Ceddia, R.B. Docosahexaenoic and eicosapentaenoic fatty acids differentially regulate glucose and fatty acid metabolism in L6 rat skeletal muscle cells. Am. J. Physiol. Cell Physiol. 2020, 319, C1120–C1129. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Li, C.; Lv, Y.; Zhang, Y.; Amakye, W.K.; Mao, L. DHA increases adiponectin expression more effectively than EPA at relative low concentrations by regulating PPARγ and its phosphorylation at Ser273 in 3T3-L1 adipocytes. Nutr. Metab. 2017, 14, 52. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Lei, J.; Schoemaker, M.H.; Ma, B.; Zhong, Y.; Lambers, T.T.; Van Tol, E.A.F.; Zhou, Y.; Nie, T.; Wu, D. Long-chain polyunsaturated fatty acids and extensively hydrolyzed casein-induced browning in a Ucp-1 reporter mouse model of obesity. Food Funct. 2018, 9, 2362–2373. [Google Scholar] [CrossRef]
- Dirandeh, E.; Ghorbanalinia, M.; Roodbari, A.R.; Colazo, M.G. Effects of dietary conjugated linoleic acid on metabolic status, BW and expression of genes related to lipid metabolism in adipose tissue of dairy cows during peripartum. Animal 2021, 15, 100105. [Google Scholar] [CrossRef] [PubMed]
- Di Cristofano, M.; Ferramosca, A.; Di Giacomo, M.; Fusco, C.; Boscaino, F.; Luongo, D.; Aufiero, V.R.; Maurano, F.; Cocca, E.; Mazzarella, G.; et al. Mechanisms underlying the hormetic effect of conjugated linoleic acid: Focus on Nrf2, mitochondria and NADPH oxidases. Free Radic. Biol. Med. 2021, 167, 276–286. [Google Scholar] [CrossRef]
- Abdelatty, A.M.; Mohamed, S.A.; Moustafa, M.M.A.; Al-Mokaddem, A.K.; Baker, M.R.; Elolimy, A.A.; Elmedany, S.A.; Hussein, S.; Farid, O.A.A.; Sakr, O.G.; et al. Nutrigenomic effect of conjugated linoleic acid on growth and meat quality indices of growing rabbit. PLoS ONE 2019, 14, e0222404. [Google Scholar] [CrossRef]
- Barone, R.; Sangiorgi, C.; Marino Gammazza, A.; D’Amico, D.; Salerno, M.; Cappello, F.; Pomara, C.; Zummo, G.; Farina, F.; Di Felice, V.; et al. Effects of Conjugated Linoleic Acid Associated With Endurance Exercise on Muscle Fibres and Peroxisome Proliferator-Activated Receptor γ Coactivator 1 α Isoforms. J. Cell. Physiol. 2017, 232, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.J.; Ko, Y.J.; Kang, S.K.; Kim, W.S.; Cho, C.S.; Choi, Y.J. Additive anti-inflammation by a combination of conjugated linoleic acid and α-lipoic acid through molecular interaction between both compounds. Food Sci. Biotechnol. 2020, 29, 419–429. [Google Scholar] [CrossRef]
- Han, H.; Li, X.; Guo, Y.; Zheng, M.; Xue, T.; Wang, L. Plant sterol ester of α-linolenic acid ameliorates high-fat diet-induced nonalcoholic fatty liver disease in mice: Association with regulating mitochondrial dysfunction and oxidative stress via activating AMPK signaling. Food Funct. 2021, 12, 2171–2188. [Google Scholar] [CrossRef]
- Li, Y.; Rong, Y.; Bao, L.; Nie, B.; Ren, G.; Zheng, C.; Amin, R.; Arnold, R.D.; Jeganathan, R.B.; Huggins, K.W. Suppression of adipocyte differentiation and lipid accumulation by stearidonic acid (SDA) in 3T3-L1 cells. Lipids Health Dis. 2017, 16, 181. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Q.; Nogoy, K.M.C.; Sun, J.; Sun, B.; Wang, Y.; Tang, L.; Yu, J.; Jin, X.; Li, X.; et al. Effect of palmitoleic acid on the differentiation of bovine skeletal muscle satellite cells. J. Anim. Sci. Technol. 2021, 63, 919–933. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Li, X.; Feng, Y.; Han, J.; Feng, Z.; Li, X.; Wen, Y. The Nuclear Orphan Receptor Nur77 Alleviates Palmitate-induced Fat Accumulation by Down-regulating G0S2 in HepG2 Cells. Sci. Rep. 2018, 8, 4809. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.M.; Ni, X.X.; Xu, Q.Y.; Wang, Q.; Li, X.Y.; Hua, J. Regulation of lipid-induced macrophage polarization through modulating peroxisome proliferator-activated receptor-gamma activity affects hepatic lipid metabolism via a Toll-like receptor 4/NF-κB signaling pathway. J. Gastroenterol. Hepatol. 2020, 35, 1998–2008. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Intervention | Duration | Study Design | Outcome | References | |
|---|---|---|---|---|---|
| Krill oil (KO) | AING-93 G diet for control and 1.5% KO-containing high-fat diet | 24 weeks | In vivo Diabetic C57BL/6 mice | KO increases cardiac PGC1-α protein expression in cardiac tissue, reducing apoptosis of cardiomyocytes and negatively regulating the NRLP3 inflammasome | [37] |
| Flaxseed Oil | Placebo or 1000 mg flaxseed oil supplements (400 mg α-linolenic acid) (twice a day) | 12 weeks | Randomized double-blind placebo-controlled trial Diabetic patients with coronary heart disease (40–85 years) n = 60 | Flaxseed oil supplementation up-regulates PPARγ gene expression and reduces TNF-α levels | [39] |
| Linseed oil (LO) | A basic diet or a basic diet supplemented with 0.5% or 1% of LO | 22 weeks | In vivo Meleagris gallopavo n = 140 | LO supplementation leads to higher levels of PPARγ in adipose tissue | [40] |
| Rapeseed oil | Control diets with 12% of energy from soybean oil or 25% of energy from butterfat. Diet with 12% of energy from rapeseed oil or a diet with rapeseed oil, fructose and cholesterol | 13 weeks | In vivo C57Bl/6J mice with NAFLD n = 28 | Rapeseed oil exerts protective effects against NAFLD by increasing PPARγ activity, lowering portal endotoxin levels and attenuating TLR4 signal | [41] |
| Blended oil | Low-fat, middle. A fat and high-fat diet based on peanut oil, lard oil diet or blended oil (canola oil, corn oil, olive oil, peanut oil, sunflower oil) | 12 weeks | In vivo Wistar rats n = 90 | A blended oil diet enhances genes related to lipid catabolism such as PPARγ in the liver, due mainly to their n-6/n-3 PUFAs ratio. Blended oil reduces TNF-α and C-reactive protein in serum | [42] |
| Fish oil | High-fat diet or a high-fat diet with 5% of fish oil (30.6% of EPA and 19.2% of DHA) | 8 weeks | In vivo Sprague-Dawley rats n = 18 | High-fat diet in fish oil may reverse the protein expression in soleus muscle of PPARγ and PGC-1α protein, improving lipid metabolism | [43] |
| DHA-enriched fish oil | Soft gels of 2400 mg of fish oil with 600 mg of n-3 PUFAs or paraffin oil. Four gels per day | 8 weeks | A double-blind randomized controlled trial Patients with DMT2 n = 50 | DHA-rich fish oil increases PPAR-γ activity in PBMCs | [44] |
| Palm oil | Low-fat diet or a high-fat diet based on palm oil (60%) | 12 weeks | In vivo C57BL6 mice n = 32 | A high-fat diet based on palm oil increases the hepatic expression of VLDLR, CD36 and PPARγ, leading to excess hepatic lipid accumulation | [45] |
| Palm oil and sunflower oil | High-calorie muffins that contain refined sunflower oil or refined palm oil. | 7 weeks | A double-blind randomized controlled trial Healthy adults (20–38 years) n = 31 | After overfeeding, the methylation of PGC-1α and TNF-α in adipose tissue was increased | [46] |
| Intervention | Duration | Study Design | Outcome | References | |
|---|---|---|---|---|---|
| n-3 PUFAs | A high-fat diet rich in saturated fats or n-3 PUFAs or a normal diet. | 13 weeks | In vivo male Sprague-Dawley n = 30 | n-3 PUFAs induce browning in white adipose tissue by the PPARγ pathway, and EPA increases PPARγ in HPAs. | [47] |
| 0, 5, 10 and 20 μmol/L of EPA | Data not shown | In vitro Human preadipocytes | |||
| n-3 PUFAs | Chow diet, high-fat diet + 260 mg/kg n-3 PUFA, high-fat diet + 100 mg/kg metformin or high-fat diet + 200 mg/kg L-carnitine | 12 weeks | In vivo Sprague Dawley rats n = 30 | n-3 PUFAs decrease body weight, glucose and insulin and increase adiponectin by GLUT4 and PPARγ regulation | [48] |
| n-3 PUFA | Supplement of two Omega 3 gel capsules per day (240 mg of DHA and 360 mg EPA) or placebo (paraffin oil capsules) | 3 weeks | A double-blind randomized controlled trial Peripheral blood mononuclear cells of athletes (PBMCs) n = 36 | Omega 3 supplementation leads to the up-regulation of PPARγ protein levels in the blood | [49] |
| n-3 PUFAs | Normocaloric diet or a high-fat diet supplemented with metformin, 300 mg/kg/d of n-3 PUFAs or a combination of both. | 8 weeks | In vivo diabetic Sprague-Dawley rats n = 38 | n-3 PUFAs alone or in combination with metformin can suppress metabolic changes related to diabetes by increasing the gene expression of PGC-1α | [50] |
| n-3 PUFAS | Four capsules/day of 0.5 g of n-3 PUFAs from fish oil, 0.62 g of α-linolenic acid from flaxseed and corn oil as a control | 180 days | A double-blind randomized controlled trial Patients with DMT2 (35–80 years) n = 185 | T2D patients with different genotypes at PPARγ, NOS3 and CD36 respond differentially to the intervention of omega-3 supplements in blood lipid profiles | [51] |
| n-3 PUFAs | Treatment with palmitic acid, oleic acid, linoleic acid, ALA, EPA and DHA at 50 μM | 24 h | In vitro LX-2 cells Primary human and mouse hepatic stellate cells | α-linolenic acid, EPA and DHA prevent liver fibrosis development by PPARγ regulation | [52] |
| n-6:n-3 PUFAs | 0.1, 0.2 and 0.4 μg/mL of chia seed extract fatty acid with a ratio of 3:1 of omega 3 and omega 6 | 14 days | In vitro Human bone marrow-derived mesenchymal stem cells PBMCs from healthy volunteer | Chia seed extracts fatty acid, decreases lipid accumulation and enhances mitochondrial fatty acid oxidation in mature adipocytes by increasing the expression of PGC-1α. Furthermore, this treatment suppresses macrophage recruitment in adipocytes and reduces protein levels of TNF-α | [53] |
| Intervention | Duration | Study Design | Outcome | References | |
|---|---|---|---|---|---|
| EPA | Normocaloric diet, high-fat diet or high-fat diet mixed with 3.6% of EPA (weight/weight) | 8 weeks | In vivo GFP-MAP1LC3 transgenic, atg5-cKO (Atg5F/F; Kap-Cre) and atg5-iKO (Atg5F/F; Ndrg1-Cre) mice n = 21 | EPA promotes lipid droplet formation and the transfer of fatty acid from them to the mitochondria for beta-oxidation, attenuating the lipotoxicity induced by a high-fat diet | [54] |
| EPA | Control diet or a high-fat diet supplemented with saline, EPA (50 mg/kg) or hidroxytyrosol (5 mg/kg) or a combination of both. | 12 weeks | In vivo C57BL/6J mice n = 80 | Supplementation with EPA improved the activity and mRNA levels of PPAR-γ and reduced the levels of TNFα and IL-6 in white adipose tissue | [55] |
| EPA | High-fat diet or a high-fat diet supplemented with 36 g/kg of EPA-enriched fish oil | 14 weeks | In vivo Wild type and UCP1 knockout C57BL/6J mice n = 40 | EPA can rescue glucose tolerance in UCP1 knockout mice via PGC1-α | [56] |
| EPA | 500 μM of palmitic acid, 500 μM of EPA or 500 μM of palmitic acid combined with 100 μM of EPA | 24 h | In vitro Human primary myotubes Human primary myoblasts derived from the abdominal rectus muscles of male individuals 31.00 ± 5.67 years | EPA increases the expression of PGC1-α and is coupled with the inhibition of the inflammatory response induced by palmitic acid. | [57] |
| EPA | EPA, DHA, linoleic acid or α-linolenic acid at 1, 10 or 100 μM | 6, 24, 48 and 72 h | In vitro Primary cultures of gilthead sea bream bone-derived MSCs | EPA treatment up-regulates the genes involved in adipogenesis such as PPAR-γ | [58] |
| EPA/DHA | 10 μM of EPA, 50 μM of DHA or 100 μM of EPA + 50 μM of DHA | 24 h | In vitro Murine 3T3-L1 cell line | EPA and DHA, alone or combined, modulate the adipogenesis of adipocytes via PPARγ-CIDEC suppression | [59] |
| EPA/DHA | 10 mM of DHA or 10 mM of EPA | 24, 72 or 120 h | In vitro Rat L6 skeletal muscle cells | DHA increases the expression of PGC1-α, regulating metabolism and fat oxidation more effectively than EPA, and reduces the expression of TNFαR | [61] |
| EPA/DHA | EPA and DHA at 25, 50, 100 or 200 μmol/L | 24 h | In vitro 3T3-L1 mouse preadipocytes | DHA led to an increase in PPARγ expression and secretes adiponectin at relatively low concentrations | [62] |
| DHA + ARA | A low-fat diet, high-fat diet, high-fat diet supplemented with ARA + DHA, high-fat diet supplemented with eHC or high-fat diet supplemented with ARA + DHA + eHC | 12 weeks | In vivo Ucp1-2A-luciferase knock-in C57BL/6J n = 60 | ARA + DHA supplementation improves metabolic flexibility and attenuates adipose tissue dysfunction during a high-fat diet as well as systemic inflammation-reducing IL-6 and TNF-α in adipose tissue | [63] |
| EPA/DHA/9M5 | Treatment with 10 μM of 9M5, 50 μM of EPA or 50 μM of DHA alone or in different combinations. | 48 h | In vitro 3T3-L1 preadipocytes | 9M5 increases the protein expression of PPARγ and lipid accumulation during the differentiation process of 3T3-L1 preadipocytes into adipocytes | [60] |
| Intervention | Duration | Study Design | Outcome | References | |
|---|---|---|---|---|---|
| CLA | 7.5 g/d of CLA or 8.78 g/d of palm oil as a control | 3 weeks | In vivo Holstein cows n = 16 | CLA decreases the mRNA abundance of genes related to fatty acid oxidation and lipolysis and increases the mRNA abundance of genes related to lipogenesis, such as PPARγ, in the adipose tissue of dairy cows | [64] |
| CLA | Supplementation 5 days/week with 600, 2000 or 3000 mg/kg of CLA or fish oil doses of 600 or 3000 mg/kg | 4 weeks | In vivo BALB/c mice n = 30 | Fish oil increases mitochondrial respiration in the liver. High doses of CLA produce steatogenic effects and promote lipid accumulation by downregulating PCG-1α | [65] |
| CLA | A basal diet with 1% of oleic acid, a diet supplemented with 0.5% of CLA and 1% of oleic acid or a diet supplemented with a 1% isomer-mix of CLA | 8 weeks | In vivo V-line rabbits n = 75 | Dietary supplementation of CLA produces a lower fat percentage via PPARγ regulation | [66] |
| CLA | Placebo sedentary/trained or CLA sedentary/trained (84% CLA, 12% oleic acid, 3% stearic acid, 0.5% palmitic acid, 0.5% linoleic acid) | 6 weeks | In vivo BALB/c mice n = 32 | CLA does not stimulate mitochondrial biogenesis or PCG-1α expression | [67] |
| CLA + α-lipoic acid | CLA at 0, 25 or 50 μM, α-lipoic acid at 0, 25 or 50 μM or a combination of both. | 24 h | In vitro Murine macrophage RAW264.7 cells | CLA α-lipoic acid increases the expression of PPARγ and shows anti-inflammatory activity through ERK1 | [68] |
| Intervention | Duration | Study Design | Outcome | References | |
|---|---|---|---|---|---|
| Sterol ester of α-linolenic acid | Control diet, a high-fat diet rich in cholesterol or one based on plant sterol or ALA acid or sterol ester of ALA | 16 weeks | In vivo C57BL/6J mice n = 50 | Sterol ester of α-linolenic acid stimulates mitochondrial biogenesis by PCG-1α regulation and reduces oxidative stress in the NAFLD. | [69] |
| 0.9 mM of oleic acid, 0.1 mM of plant sterol, 0.1 mM of α-linolenic acid or 0.1 mM sterol ester of ALA | 24 h | In vitro HepG2 cells | |||
| Stearidonic acid | 50 or 200 μM of stearidonic acid, EPA, DHA or ALA. | 24, 75, and 144 h | In vitro 3T3-L1 preadipocytes | Stearidonic acid can inhibit adipocyte differentiation and reduce lipid accumulation by downregulating PPARγ | [70] |
| Palmitoleic acid (POA) | 50 μM, 100 μM and 200 μM of POA | 96 h | In vitro Bovine satellite cells (BSC) | All treatments of POA increased the protein expression of PPARγ and induced transdifferentiation of BSC into adipocytes. After treatment, PLIN1 is up-regulated | [71] |
| Palmitic acid | Palmitate at 50, 100 or 200 mM | 24 h | In vitro HepG2 cells | Palmitate-induced lipid accumulation and suppressed lipolysis in HepG2 cells via PPARγ stimulation | [72] |
| Palmitic acid/ DHA | Normocaloric diet or high-fat diet with rosiglitazone | 12 weeks | In vivo C57BL/6 mice | Palmitic acid induces the M1 polarized macrophage, which promotes lipid accumulation in hepatocytes via PPARγ and enhances the expression of IL-6 and TNF-α; DHA promotes M2 phenotype | [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-García, C.; Sánchez-Quesada, C.; Martínez-Ramírez, M.J.; Gaforio, J.J. PPARγ Gene as a Possible Link between Acquired and Congenital Lipodystrophy and its Modulation by Dietary Fatty Acids. Nutrients 2022, 14, 4742. https://doi.org/10.3390/nu14224742
Rodríguez-García C, Sánchez-Quesada C, Martínez-Ramírez MJ, Gaforio JJ. PPARγ Gene as a Possible Link between Acquired and Congenital Lipodystrophy and its Modulation by Dietary Fatty Acids. Nutrients. 2022; 14(22):4742. https://doi.org/10.3390/nu14224742
Chicago/Turabian StyleRodríguez-García, Carmen, Cristina Sánchez-Quesada, María José Martínez-Ramírez, and José J. Gaforio. 2022. "PPARγ Gene as a Possible Link between Acquired and Congenital Lipodystrophy and its Modulation by Dietary Fatty Acids" Nutrients 14, no. 22: 4742. https://doi.org/10.3390/nu14224742
APA StyleRodríguez-García, C., Sánchez-Quesada, C., Martínez-Ramírez, M. J., & Gaforio, J. J. (2022). PPARγ Gene as a Possible Link between Acquired and Congenital Lipodystrophy and its Modulation by Dietary Fatty Acids. Nutrients, 14(22), 4742. https://doi.org/10.3390/nu14224742