
Mobile Phone Radiation Deflects Brain Energy Homeostasis and Prompts Human Food Ingestion

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Design
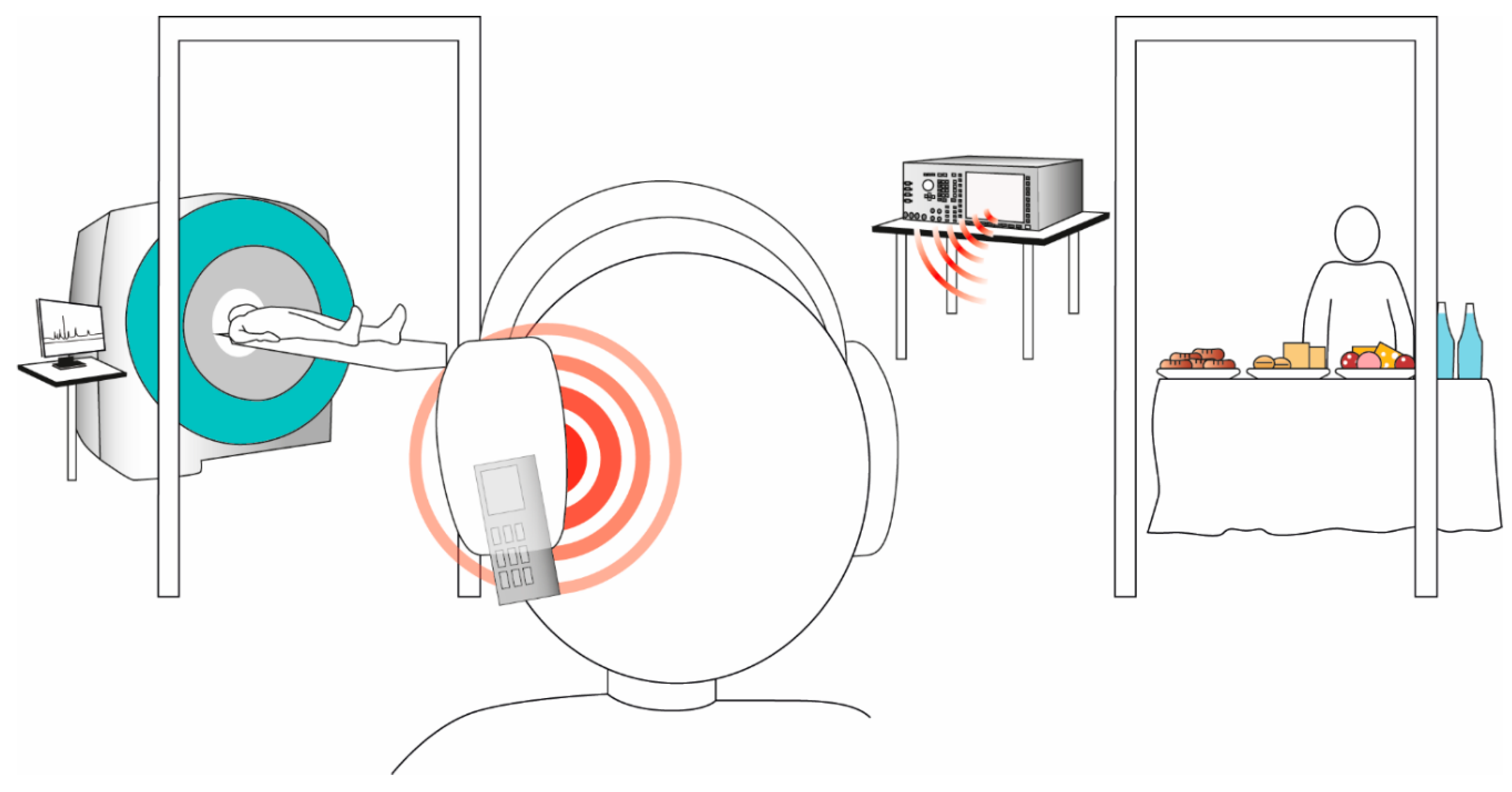
2.3. Mobile Phone Radiation Exposure
2.4. 31Phosphorus Magnetic Resonance Spectroscopy Measurements
2.5. Statistical Analysis
3. Results
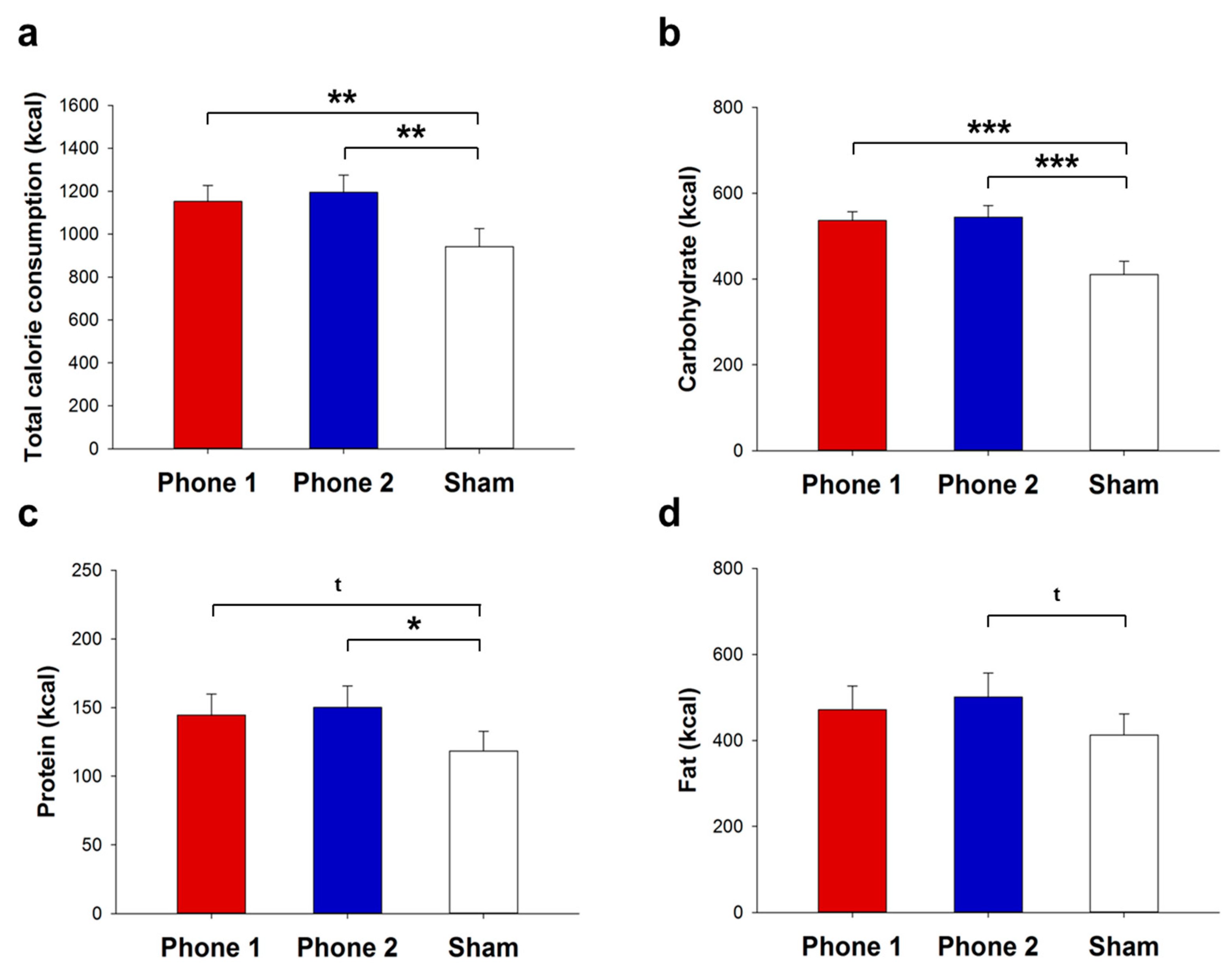
3.1. Food Ingestion
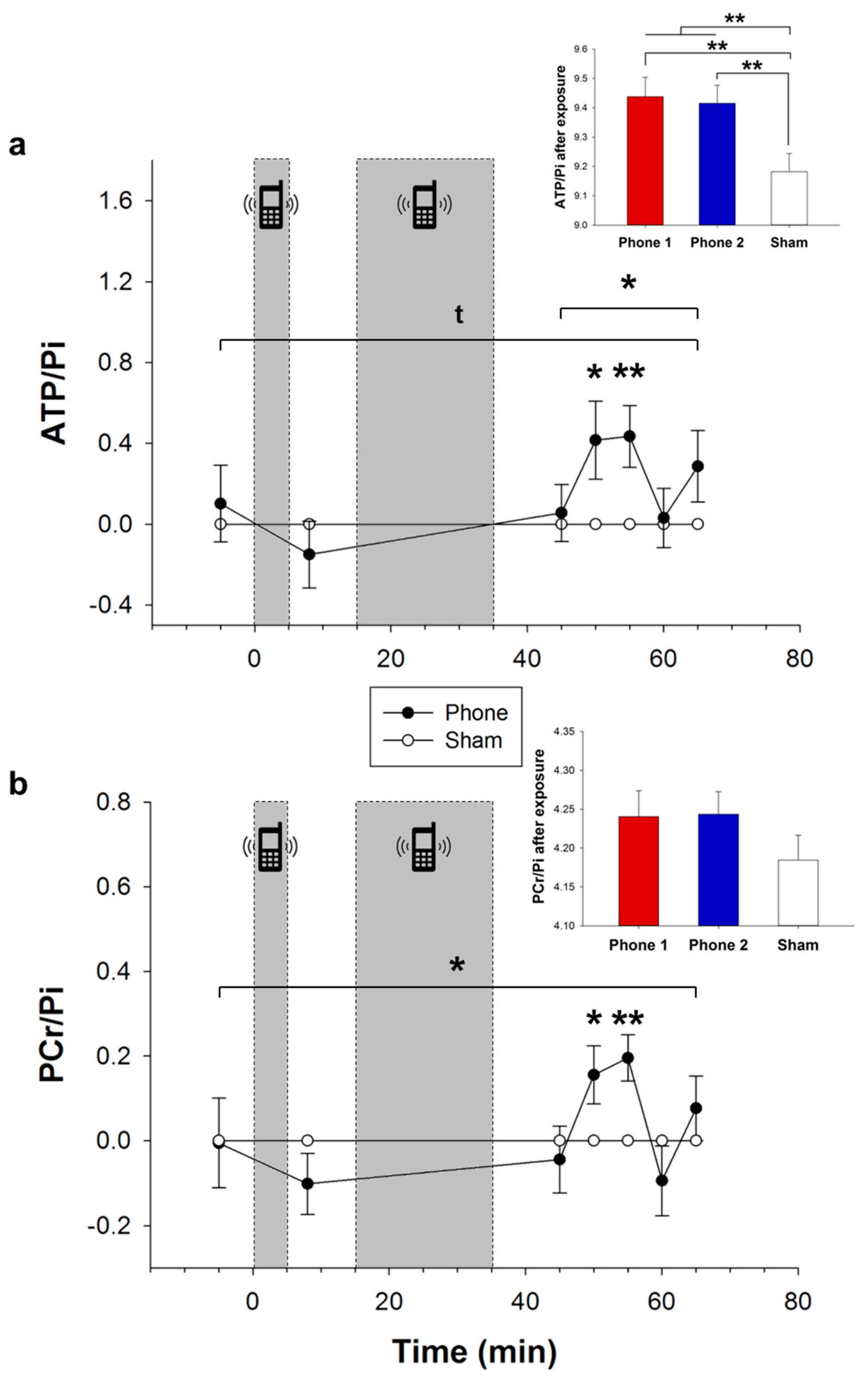
3.2. Cerebral High-Energy Phosphate Metabolism
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Statista, Number of Smartphone Users from 2016 to 2021. Available online: https://www.statista.com/statistics/330695/number-of-smartphone-users-worldwide/ (accessed on 8 August 2021).
- Swinburn, B.A.; Sacks, G.; Hall, K.D.; McPherson, K.; Finegood, D.T.; Moodie, M.L.; Gortmaker, S.L. The global obesity pandemic: Shaped by global drivers and local environments. Lancet 2011, 37, 804–814. [Google Scholar] [CrossRef]
- Collaborators, G.B.D.R.F. Global burden of 87 risk factors in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of disease study 2019. Lancet 2020, 396, 1223–1249. [Google Scholar]
- WHO. Fact Sheet-Obesity and Overweight. Updated 2020. Available online: https://www.who.int/en/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 6 June 2020).
- Wada, K.; Yamakawa, M.; Konishi, K.; Goto, Y.; Mizuta, F.; Koda, S.; Uji, T.; Tamura, T.; Nakamura, K.; Tsuji, M.; et al. Associations of cell phone use and screen viewing with overweight in children. Child Obes. 2019, 15, 417–425. [Google Scholar] [CrossRef]
- Kim, K.H.; Kabir, E.; Jahan, S.A. The use of cell phone and insight into its potential human health impacts. Environ. Monit. Assess. 2016, 188, 221. [Google Scholar] [CrossRef]
- Fernandez, C.; de Salles, A.A.; Sears, M.E.; Morris, R.D.; Davis, D.L. Absorption of wireless radiation in the child versus adult brain and eye from cell phone conversation or virtual reality. Environ. Res. 2018, 167, 694–699. [Google Scholar] [CrossRef]
- Cardis, E.; Deltour, I.; Mann, S.; Moissonnier, M.; Taki, M.; Varsier, N.; Wake, K.; Wiart, J. Distribution of RF energy emitted by mobile phones in anatomical structures of the brain. Phys. Med. Biol. 2008, 53, 2771–2783. [Google Scholar] [CrossRef]
- Cardis, E.; Varsier, N.; Bowman, J.D.; Deltour, I.; Figuerola, J.; Mann, S.; Moissonnier, M.; Taki, M.; Vecchia, P.; Villegas, R.; et al. Estimation of RF energy absorbed in the brain from mobile phones in the Interphone Study. Occup. Environ. Med. 2011, 68, 686–693. [Google Scholar] [CrossRef]
- Gultekin, D.H.; Moeller, L. NMR imaging of cell phone radiation absorption in brain tissue. Proc. Natl. Acad. Sci. USA 2013, 110, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Tomasi, D.; Wang, G.J.; Vaska, P.; Fowler, J.S.; Telang, F.; Alexoff, D.; Logan, J.; Wong, C. Effects of cell phone radiofrequency signal exposure on brain glucose metabolism. JAMA 2011, 305, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, F.; Curcio, G.; Pasqualetti, P.; De Gennaro, L.; Fini, R.; Rossini, P.M. Mobile phone emissions and human brain excitability. Ann. Neurol. 2006, 60, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Roggeveen, S.; van Os, J.; Viechtbauer, W.; Lousberg, R. EEG changes due to experimentally induced 3G mobile phone radiation. PLoS ONE 2015, 10, e0129496. [Google Scholar]
- Vecchio, F.; Buffo, P.; Sergio, S.; Iacoviello, D.; Rossini, P.M.; Babiloni, C. Mobile phone emission modulates event-related desynchronization of alpha rhythms and cognitive-motor performance in healthy humans. Clin. Neurophysiol. 2012, 123, 121–128. [Google Scholar] [CrossRef]
- Birks, L.E.; van Wel, L.; Liorni, I.; Pierotti, L.; Guxens, M.; Huss, A.; Foerster, M.; Capstick, M.; Eeftens, M.; El Marroun, H.; et al. Radiofrequency electromagnetic fields from mobile communication: Description of modeled dose in brain regions and the body in European children and adolescents. Environ. Res. 2021, 193, 110505. [Google Scholar] [CrossRef] [PubMed]
- Wiart, J.; Hadjem, A.; Gadi, N.; Bloch, I.; Wong, M.F.; Pradier, A.; Lautru, D.; Hanna, V.F.; Dale, C. Modeling of RF head exposure in children. Bioelectromagnetics 2005, 26, S19–S30. [Google Scholar] [CrossRef]
- Christ, A.; Gosselin, M.-C.; Christopoulou, M.; Kühn, S.; Kuster, N. Age-dependent tissue-specific exposure of cell phone users. Phys. Med. Biol. 2010, 55, 1767–1783. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M.W.; Porte, D., Jr. Diabetes, obesity, and the brain. Science 2005, 307, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Lam, T.K. Neuronal regulation of homeostasis by nutrient sensing. Nat. Med. 2010, 16, 392–395. [Google Scholar] [CrossRef]
- Tripathi, R.; Banerjee, S.K.; Nirala, J.P.; Mathur, R. Simultaneous exposure to electromagnetic field from mobile phone and unimpeded fructose drinking during pre-peri-and post-pubertal stages perturbs the hypothalamic and hepatic regulation of energy homeostasis by early adulthood: Experimental evidence. Environ. Sci. Pollut. Res. Int. 2021, 1–14. [Google Scholar] [CrossRef]
- Sommer, A.M.; Streckert, J.; Bitz, A.K.; Hansen, V.W.; Lerchl, A. No effects of GSM-modulated 900 MHz electromagnetic fields on survival rate and spontaneous development of lymphoma in female AKR/J mice. BMC Cancer 2004, 4, 77. [Google Scholar] [CrossRef]
- Lerchl, A.; Kruger, H.; Niehaus, M.; Streckert, J.R.; Bitz, A.K.; Hansen, V. Effects of mobile phone electromagnetic fields at nonthermal SAR values on melatonin and body weight of Djungarian hamsters (Phodopus sungorus). J. Pineal. Res. 2008, 44, 267–672. [Google Scholar] [CrossRef]
- Pelletier, A.; Delanaud, S.; Decima, P.; Thuroczy, G.; de Seze, R.; Cerri, M.; Bach, V.; Libert, J.P.; Loos, N. Effects of chronic exposure to radiofrequency electromagnetic fields on energy balance in developing rats. Environ. Sci. Pollut. Res. Int. 2013, 20, 2735–2746. [Google Scholar] [CrossRef]
- Sanders, A.P.; Joines, W.T.; Allis, J.W. Effects of continuous-wave, pulsed, and sinusoidal-amplitude-modulated microwaves on brain energy metabolism. Bioelectromagnetics 1985, 6, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Jauch-Chara, K.; Friedrich, A.; Rezmer, M.; Melchert, U.H.; Scholand-Engler, H.G.; Hallschmid, M.; Oltmanns, K.M. Intranasal insulin suppresses food intake via enhancement of brain energy levels in humans. Diabetes 2012, 61, 2261–2268. [Google Scholar] [CrossRef]
- Jauch-Chara, K.; Oltmanns, K.M. Obesity—A neuropsychological disease? Systematic review and neuropsychological model. Prog. Neurobiol. 2014, 114, 84–101. [Google Scholar] [CrossRef]
- Wardzinski, E.K.; Hyzy, C.; Duysen, K.; Melchert, U.H.; Jauch-Chara, K.; Oltmanns, K.M. Hypocaloric dieting unsettles the neuroenergetic homeostasis in humans. Nutrients 2021, 13, 3433. [Google Scholar] [CrossRef] [PubMed]
- Spaeth, A.M.; Dinges, D.F.; Goel, N. Effects of experimental sleep restriction on weight gain, caloric intake, and meal timing in healthy adults. Sleep 2013, 36, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Kracht, C.L.; Chaput, J.P.; Martin, C.K.; Champagne, C.M.; Katzmarzyk, P.T.; Staiano, A.E. Associations of sleep with food cravings, diet, and obesity in adolescence. Nutrients 2019, 11, 2899. [Google Scholar] [CrossRef]
- Kistenmacher, A.; Manneck, S.; Wardzinski, E.K.; Martens, J.C.; Gohla, G.; Melchert, U.H.; Jauch-Chara, K.; Oltmanns, K.M. Persistent blood glucose reduction upon repeated transcranial electric stimulation in men. Brain Stimul. 2017, 10, 780–786. [Google Scholar] [CrossRef]
- Wardzinski, E.K.; Friedrichsen, L.; Dannenberger, S.; Kistenmacher, A.; Melchert, U.H.; Jauch-Chara, K.; Oltmanns, K.M. Double transcranial direct current stimulation of the brain increases cerebral energy levels and systemic glucose tolerance in men. J. Neuroendocrinol. 2019, 31, e12688. [Google Scholar] [CrossRef]
- Binkofski, F.; Loebig, M.; Jauch-Chara, K.; Bergmann, S.; Melchert, U.H.; Scholand-Engler, H.G.; Schweiger, U.; Pellerin, L.; Oltmanns, K.M. Brain energy consumption induced by electrical stimulation promotes systemic glucose uptake. Biol. Psychiatry 2011, 70, 690–695. [Google Scholar] [CrossRef]
- Kistenmacher, A.; Goetsch, J.; Ullmann, D.; Wardzinski, E.K.; Melchert, U.H.; Jauch-Chara, K.; Oltmanns, K.M. Psychosocial stress promotes food intake and enhances the neuroenergetic level in men. Stress 2018, 21, 538–547. [Google Scholar] [CrossRef]
- Cassioli, D.; Durantini, A. Measurements, modeling and simulations of the UWB propagation channel based on direct-sequence channel sounding. Wirel. Commun. Mob. Comput. 2005, 5, 513–523. [Google Scholar] [CrossRef]
- Dudoyer, S.; Deniau, V.; Adriano, R.; Ben Slimen, M.N.; Rioult, J.; Meyniel, B.; Berbineau, M. Study of the susceptibility of the GSM-R communications face to the electromagnetic interferences of the rail environment. IEEE Trans. Electromagn. Compat. 2012, 54, 667–676. [Google Scholar] [CrossRef]
- Bachert-Baumann, P.; Ermark, F.; Zabel, H.J.; Sauter, R.; Semmler, W.; Lorenz, W.J. In vivo nuclear Overhauser effect in 31P-(1H) double-resonance experiments in a 1.5-T whole-body MR system. Magn. Reson. Med. 1990, 15, 165–172. [Google Scholar] [CrossRef]
- Barker, P.B.; Golay, X.; Artemov, D.; Ouwerkerk, R.; Smith, M.A.; Shaka, A.J. Broadband proton decoupling for in vivo brain spectroscopy in humans. Magn. Reson. Med. 2001, 45, 226–232. [Google Scholar] [CrossRef]
- Vanhamme, L.; van den Boogaart, A.; Van Huffel, S. Improved method for accurate and efficient quantification of MRS data with use of prior knowledge. J. Magn. Reson. 1997, 129, 35–43. [Google Scholar] [CrossRef]
- Schmoller, A.; Hass, T.; Strugovshchikova, O.; Melchert, U.H.; Scholand-Engler, H.G.; Peters, A.; Schweiger, U.; Hohagen, F.; Oltmanns, K.M. Evidence for a relationship between body mass and energy metabolism in the human brain. J. Cereb. Blood Flow Metab. 2010, 30, 1403–1410. [Google Scholar] [CrossRef]
- Wardzinski, E.K.; Kistenmacher, A.; Melchert, U.H.; Jauch-Chara, K.; Oltmanns, K.M. Impaired brain energy gain upon a glucose load in obesity. Metabolism 2018, 85, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Lovely, R.H.; Mizumori, S.J.Y.; Johnson, R.B.; Guy, A.W. Subtle consequences of exposure to weak microwave fields: Are there nonthermal effects? A2-ADAIR, ELEANOR R. In Microwaves and Thermoregulation; Academic Press: San Diego, CA, USA, 1983; pp. 401–429. [Google Scholar]
- Taberski, K.; Klose, M.; Grote, K.; El Ouardi, A.; Streckert, J.; Hansen, V.W.; Lerchl, A. Noninvasive assessment of metabolic effects of exposure to 900 MHz electromagnetic fields on Djungarian Hamsters (Phodopus sungorus). Radiat. Res. 2014, 181, 617–622. [Google Scholar] [CrossRef]
- Lajunen, H.R.; Keski-Rahkonen, A.; Pulkkinen, L.; Rose, R.J.; Rissanen, A.; Kaprio, J. Are computer and cell phone use associated with body mass index and overweight? A population study among twin adolescents. BMC Public Health 2007, 7, 24. [Google Scholar] [CrossRef]
- Jauch-Chara, K.; Kistenmacher, A.; Herzog, N.; Schwarz, M.; Schweiger, U.; Oltmanns, K.M. Repetitive electric brain stimulation reduces food intake in humans. Am. J. Clin. Nutr. 2014, 100, 1003–1009. [Google Scholar] [CrossRef]
- Kim, J.H.; Chung, K.H.; Hwang, Y.R.; Park, H.R.; Kim, H.J.; Kim, H.-G.; Kim, H.R. Exposure to RF-EMF alters postsynaptic structure and hinders neurite outgrowth in developing hippocampal neurons of early postnatal mice. Int. J. Mol. Sci. 2021, 22, 5340. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, R.J.; Francis, H.M. The hippocampus and the regulation of human food intake. Psychol. Bull. 2017, 143, 1011–1032. [Google Scholar] [CrossRef]
- Pocai, A.; Lam, T.K.; Gutierrez-Juarez, R.; Obici, S.; Schwartz, G.J.; Bryan, J.; Aguilar-Bryan, L.; Rossetti, L. Hypothalamic K(ATP) channels control hepatic glucose production. Nature 2005, 434, 1026–1031. [Google Scholar] [CrossRef]
- Schwartz, M.W.; Woods, S.C.; Porte, D.; Seeley, R.J.; Baskin, D.G. Central nervous system control of food intake. Nature 2000, 404, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Yang, H.; Chang, R.; Yin, P.; Yang, Y.; Yang, W.; Huang, S.; Gaertig, M.A.; Li, S.; Li, X.-J. MANF regulates hypothalamic control of food intake and body weight. Nat. Commun. 2017, 8, 579. [Google Scholar] [CrossRef]
- Gandhi, O.P.; Lazzi, G.; Furse, C.M. Electromagnetic absorption in the human head and neck for mobile telephones at 835 and 1900 MHz. IEEE Trans. Microw. Theory Tech. 1996, 44, 1884–1897. [Google Scholar] [CrossRef]
- Tsoy, A.; Saliev, T.; Abzhanova, E.; Turgambayeva, A.; Kaiyrlykyzy, A.; Akishev, M.; Saparbayev, S.; Umbayev, B.; Askarova, S. The effects of mobile phone radiofrequency electromagnetic fields on beta-amyloid-induced oxidative stress in human and rat primary astrocytes. Neuroscience 2019, 408, 46–57. [Google Scholar] [CrossRef]
- Teissie, J.; Knox, B.E.; Tsong, T.Y.; Wehrle, J. Synthesis of adenosine triphosphate in respiration-inhibited submitochondrial particles induced by microsecond electric pulses. Proc. Natl. Acad. Sci. USA 1981, 78, 7473–7477. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Sharma, S.; Shrivastava, S.; Singhal, P.K.; Shukla, S. Mobile phone induced cognitive and neurochemical consequences. J. Chem. Neuroanat. 2019, 102, 101684. [Google Scholar] [CrossRef]
- Singh, K.V.; Gautam, R.; Meena, R.; Nirala, J.P.; Jha, S.K.; Rajamani, P. Effect of mobile phone radiation on oxidative stress, inflammatory response, and contextual fear memory in Wistar rat. Environ. Sci. Pollut. Res. Int. 2020, 27, 19340–19351. [Google Scholar] [CrossRef]
- Durusoy, R.; Hassoy, H.; Ozkurt, A.; Karababa, A.O. Mobile phone use, school electromagnetic field levels and related symptoms: A cross-sectional survey among 2150 high school students in Izmir. Environ. Health 2017, 16, 51. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.T.; Chang, Y.H.; Chen, C.C.; Ko, M.C.; Li, C.Y. Mobile phone use and health symptoms in children. J. Formos. Med. Assoc. 2015, 114, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Rideout, V. The Common Sense Census: Media Use by Kids Age Zero to Eight; Common Sense Media: San Francisco, CA, USA, 2017; pp. 263–283. [Google Scholar]
- Kabali, H.K.; Irigoyen, M.M.; Nunez-Davis, R.; Budacki, J.G.; Mohanty, S.H.; Leister, K.P.; Bonner, R.L., Jr. Exposure and use of mobile media devices by young children. Pediatrics 2015, 136, 1044–1050. [Google Scholar] [CrossRef]
- Reilly, J.J.; El-Hamdouchi, A.; Diouf, A.; Monyeki, A.; Somda, S.A. Determining the worldwide prevalence of obesity. Lancet 2018, 391, 1773–1774. [Google Scholar] [CrossRef]
- Mondal, S.P.; Padmini, T.N. Specific absorption rate (SAR), ways to find in mobile devices, its measurement and side effects on human. In Proceedings of the 2019 International Conference on Vision Towards Emerging Trends in Communication and Networking (ViTECoN), Vellore, India, 30–31 March 2019; pp. 1–4. [Google Scholar]
- International Commission on Non-Ionizing Radiation Protection. Guidelines for limiting exposure to electromagnetic fields (100 kHz to 300 GHz). Health Phys. 2020, 118, 483–524. [Google Scholar] [CrossRef]
- Fenton, S.; Burrows, T.L.; Skinner, J.A.; Duncan, M.J. The influence of sleep health on dietary intake: A systematic review and meta-analysis of intervention studies. J. Hum. Nutr. Diet. 2021, 34, 273–285. [Google Scholar] [CrossRef]
- Dworak, M.; Kim, T.; McCarley, R.W.; Basheer, R. Sleep, brain energy levels, and food intake: Relationship between hypothalamic ATP concentrations, food intake, and body weight during sleep-wake and sleep deprivation in rats. Somnologie 2011, 15, 111–117. [Google Scholar] [CrossRef][Green Version]
- López-Cepero, A.; O’Neill, J.; Tamez, M.; Falcón, L.M.; Tucker, K.L.; Rodríguez-Orengo, J.F.; Mattei, J. Associations between perceived stress and dietary intake in adults in Puerto Rico. J. Acad. Nutr. Diet. 2021, 121, 762–769. [Google Scholar] [CrossRef]
- van Deventer, E.; van Rongen, E.; Saunders, R. WHO research agenda for radiofrequency fields. Bioelectromagnetics 2011, 32, 417–421. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wardzinski, E.K.; Jauch-Chara, K.; Haars, S.; Melchert, U.H.; Scholand-Engler, H.G.; Oltmanns, K.M. Mobile Phone Radiation Deflects Brain Energy Homeostasis and Prompts Human Food Ingestion. Nutrients 2022, 14, 339. https://doi.org/10.3390/nu14020339
Wardzinski EK, Jauch-Chara K, Haars S, Melchert UH, Scholand-Engler HG, Oltmanns KM. Mobile Phone Radiation Deflects Brain Energy Homeostasis and Prompts Human Food Ingestion. Nutrients. 2022; 14(2):339. https://doi.org/10.3390/nu14020339
Chicago/Turabian StyleWardzinski, Ewelina K., Kamila Jauch-Chara, Sarah Haars, Uwe H. Melchert, Harald G. Scholand-Engler, and Kerstin M. Oltmanns. 2022. "Mobile Phone Radiation Deflects Brain Energy Homeostasis and Prompts Human Food Ingestion" Nutrients 14, no. 2: 339. https://doi.org/10.3390/nu14020339
APA StyleWardzinski, E. K., Jauch-Chara, K., Haars, S., Melchert, U. H., Scholand-Engler, H. G., & Oltmanns, K. M. (2022). Mobile Phone Radiation Deflects Brain Energy Homeostasis and Prompts Human Food Ingestion. Nutrients, 14(2), 339. https://doi.org/10.3390/nu14020339