
Weizmannia coagulans BC2000 Plus Ellagic Acid Inhibits High-Fat-Induced Insulin Resistance by Remodeling the Gut Microbiota and Activating the Hepatic Autophagy Pathway in Mice

 ,
,
Abstract
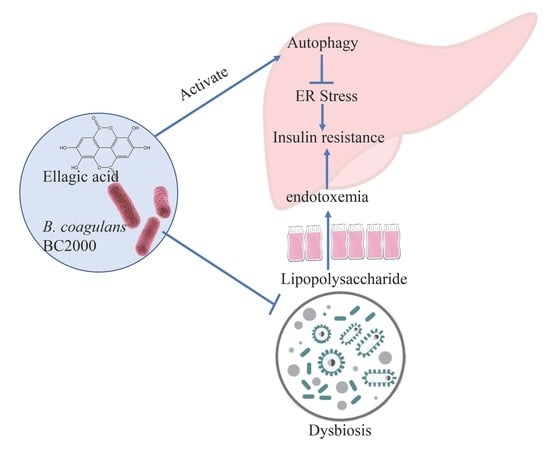
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Evaluation of the Probiotic Properties of W. coagulans
2.3. Experimental Design and Animals
2.4. Tissue Sample Collection
2.5. Determination of Insulin Resistance Indicators
2.6. Determination of Inflammatory Cytokine Indicators
2.7. High-Throughput Sequencing of 16S rDNA in Colonic Contents
2.8. Mouse Liver RNA Sequencing and Gene Enrichment Analysis
2.9. Statistical Analysis
3. Results
3.1. Probiotic Properties of W. coagulans
3.2. Impact of W. coagulans and EA on Body Weight of High-Fat Diet-Fed Mice
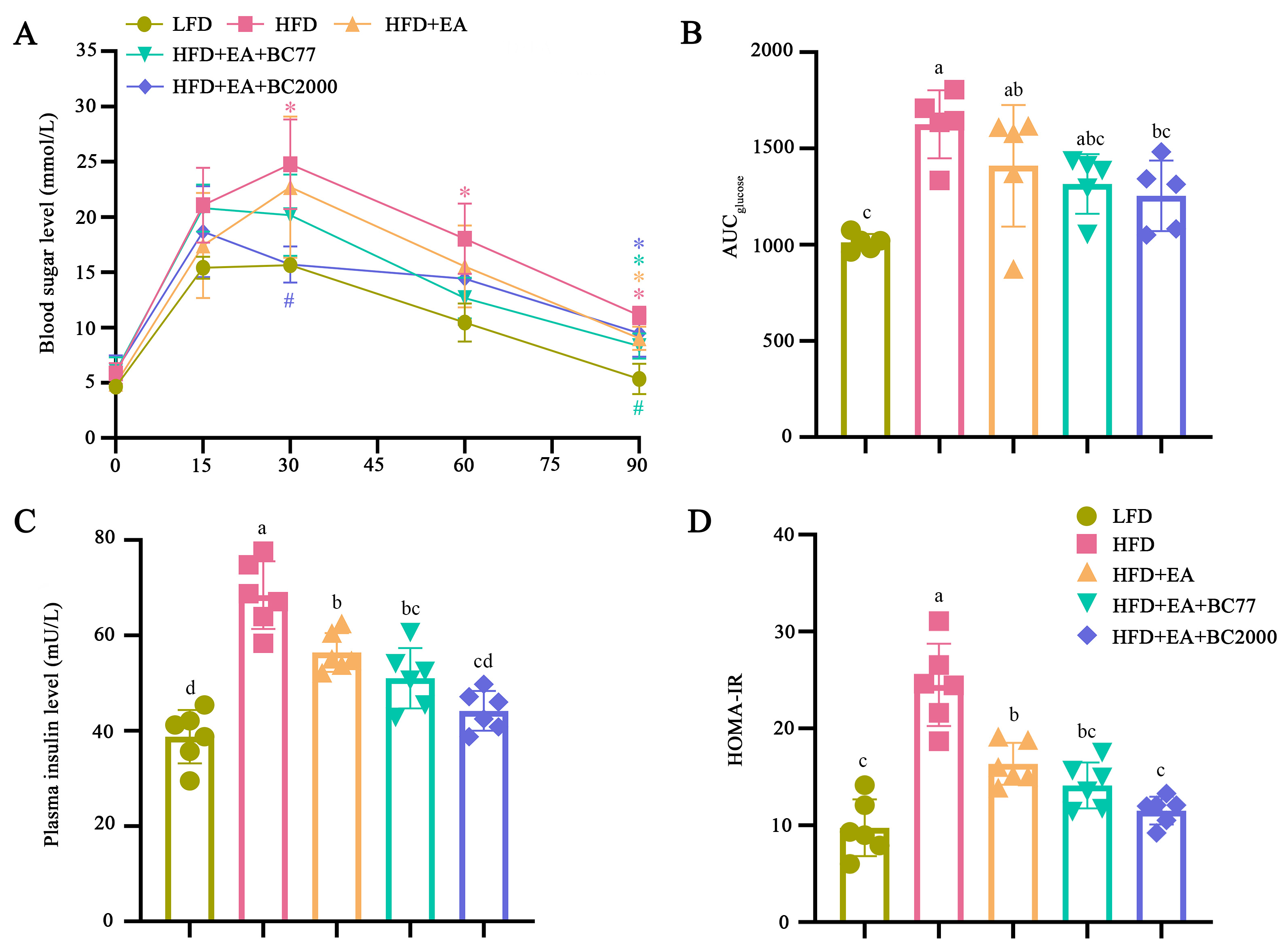
3.3. Impact of W. coagulans and EA on Insulin Resistance in the High-Fat Diet-Fed Mice
3.4. Impact of W. coagulans and EA on Inflammatory Cytokines in Obese Mice
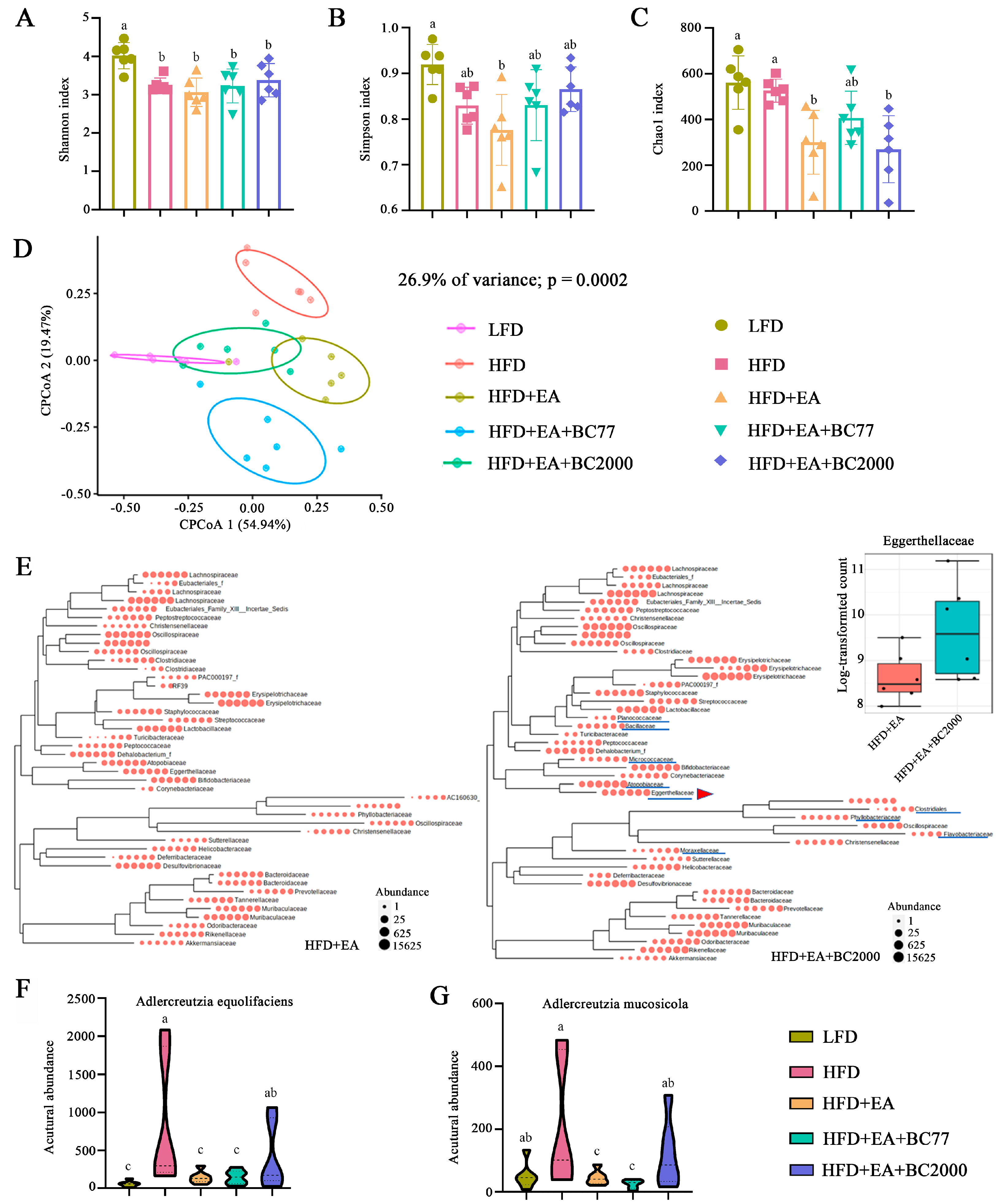
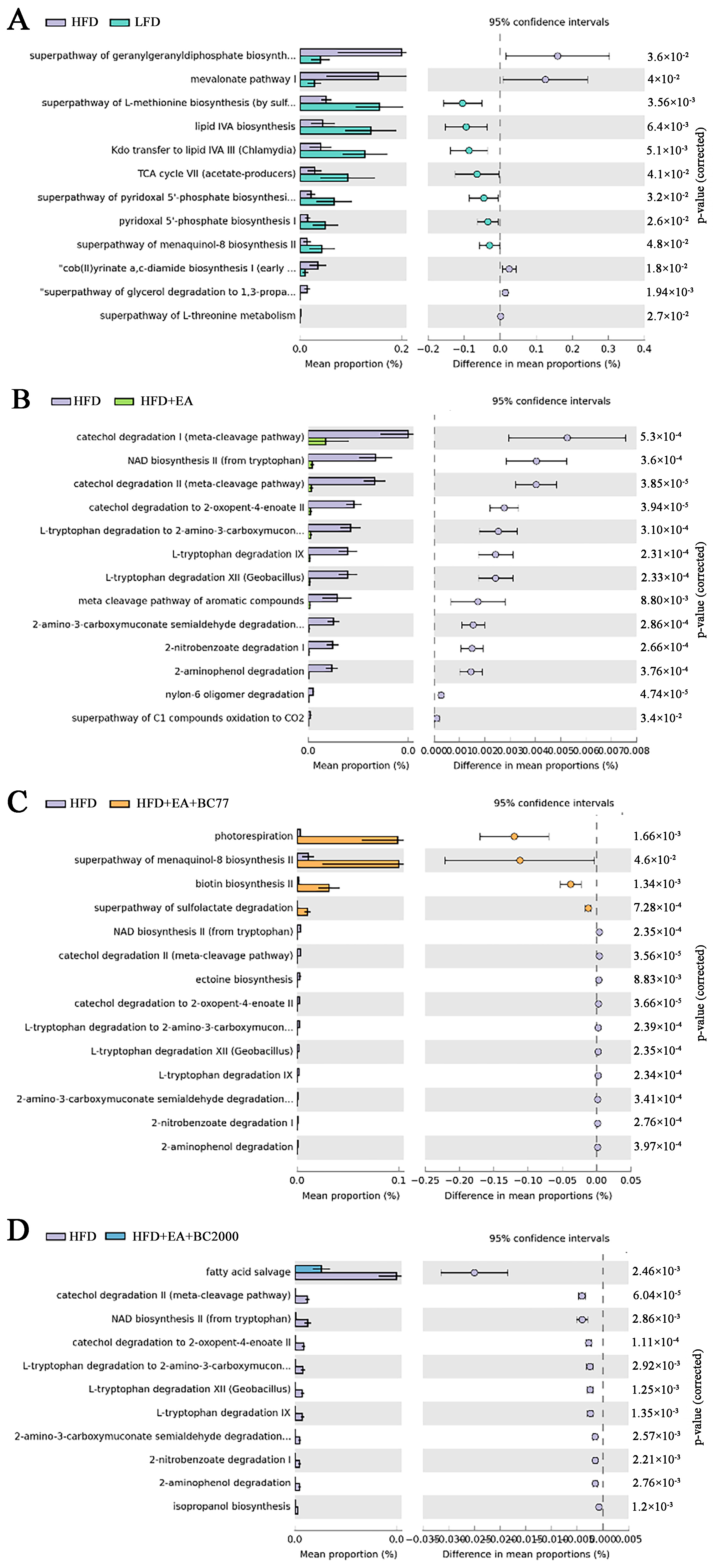
3.5. Impact of W. coagulans and EA on Cecum Microbiota
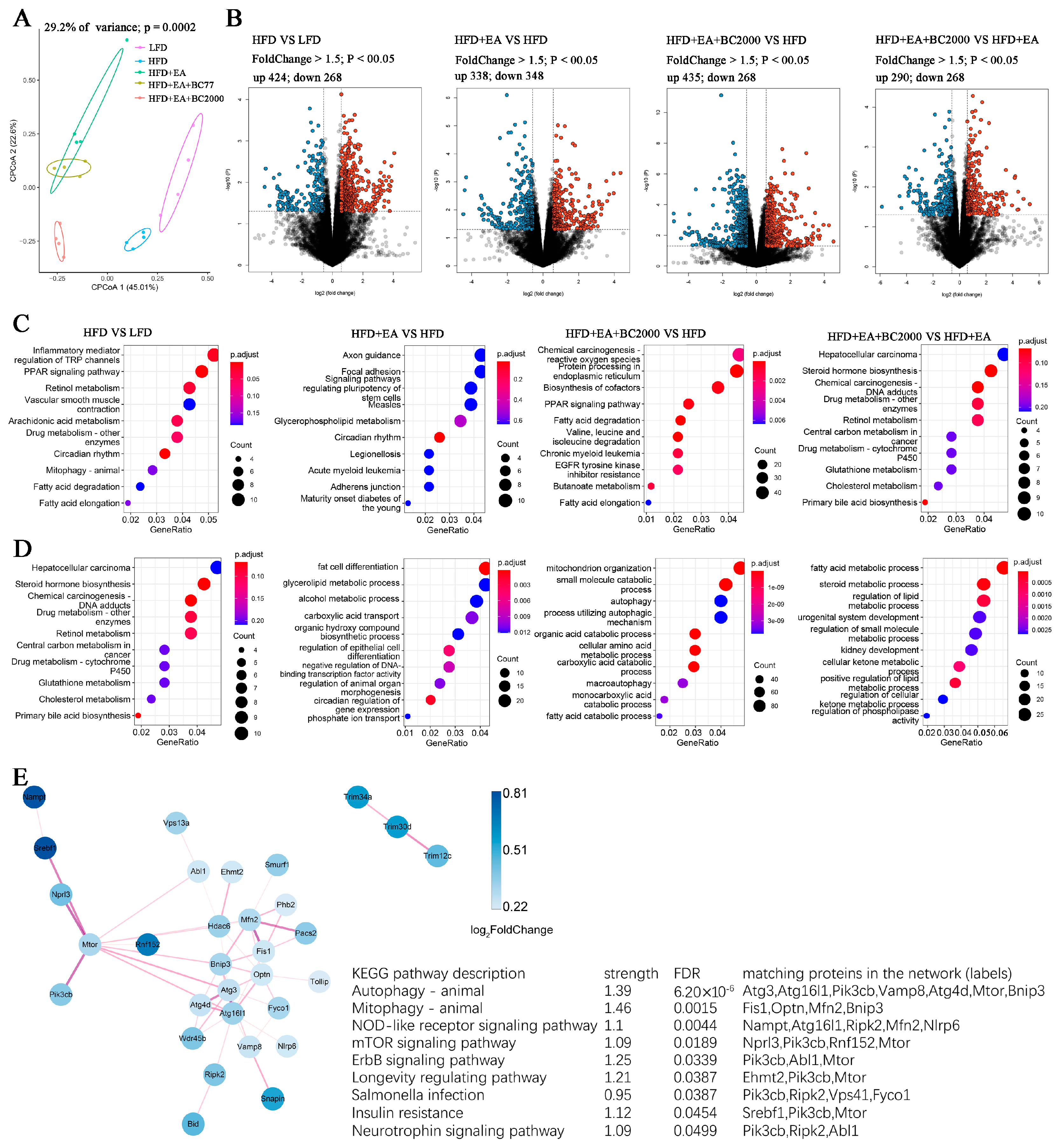
3.6. Impact of BC2000 on Gene Expression in Mouse Liver
3.7. Insulin Signaling Gene Expression
3.8. Differential Gene KEGG Enrichment Analysis
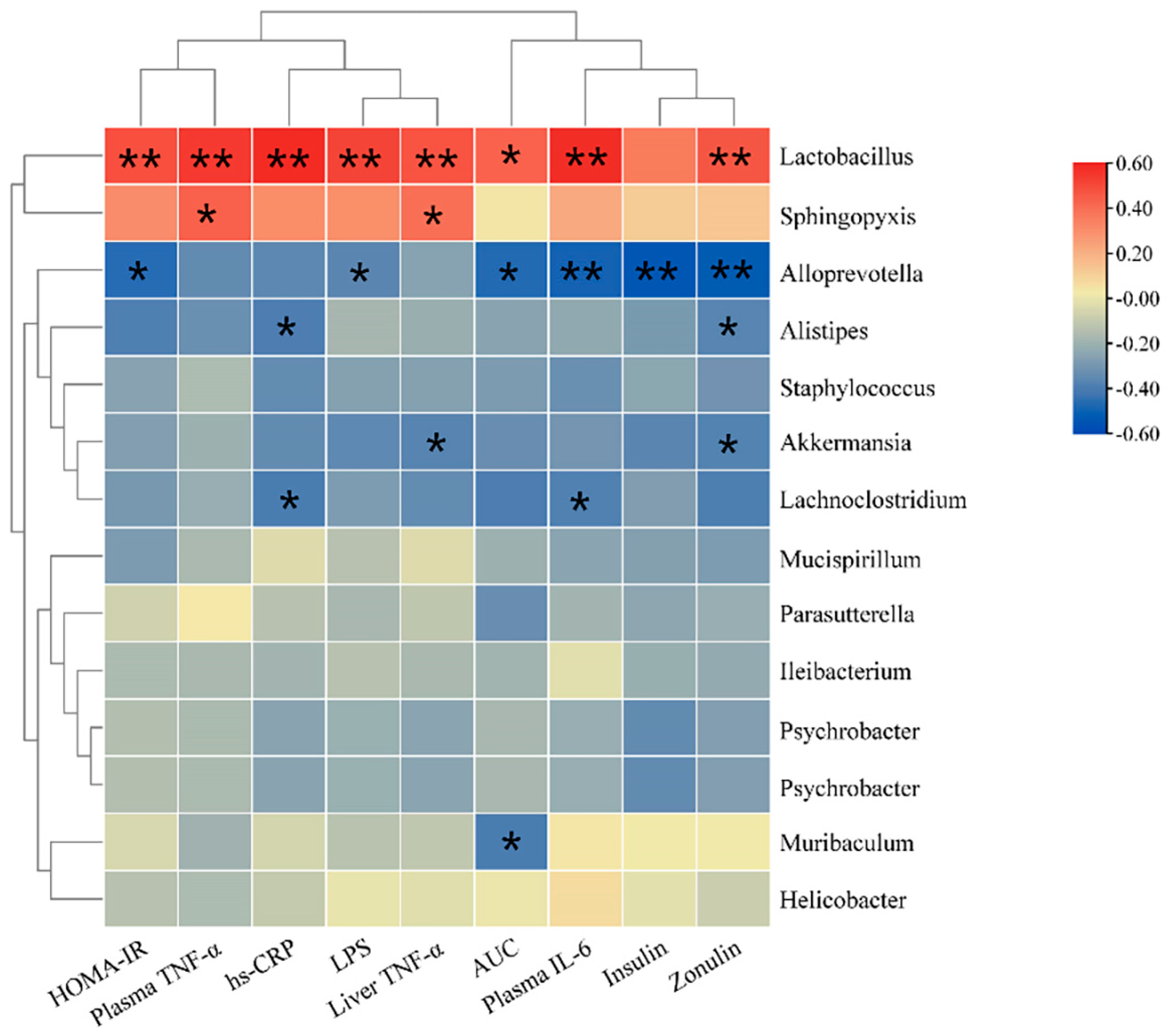
3.9. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yalcin, G.; Demirbag, C.; Bahsi, I.; Ozgul, L.; Alkaya, D.B.; Onurlu, H.I.; Seyhan, S.A. Determination of Ellagic Acid in the Wastes of Walnut, Chestnut, and Pomegranate Grown in Turkey. In Advances in Plant Phenolics: From Chemistry to Human Health; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2018; Volume 1286, pp. 5–81. ISBN 9780841232969. [Google Scholar]
- Zhang, Y.; Zhang, Y.; Halemahebai, G.; Tian, L.; Dong, H.; Aisker, G. Urolithin A, a pomegranate metabolite, protects pancreatic β cells from apoptosis by activating autophagy. J. Ethnopharmacol. 2021, 272, 113628. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.L.; Zhang, Y.; Gulihaixia, H.; Bahetibieke, T.; Zhang, Y.Z. Urolithin A Activates Autophagy to Improve Liver Insulin Resistance in Diabetic Mice. Chinese Tradit. Herb. Drugs 2020, 51, 710–716. [Google Scholar]
- Cortés-Martín, A.; Selma, M.V.; Tomás-Barberán, F.A.; González-Sarrías, A.; Espín, J.C. Where to Look into the Puzzle of Polyphenols and Health? The Postbiotics and Gut Microbiota Associated with Human Metabotypes. Mol. Nutr. Food Res. 2020, 64, 1900952. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; García-Villalba, R.; González-Sarrías, A.; Selma, M.V.; Espín, J.C. Ellagic Acid Metabolism by Human Gut Microbiota: Consistent Observation of Three Urolithin Phenotypes in Intervention Trials, Independent of Food Source, Age, and Health Status. J. Agric. Food Chem. 2014, 62, 6535–6538. [Google Scholar] [CrossRef]
- Selma, M.V.; Beltrán, D.; García-Villalba, R.; Espín, J.C.; Tomás-Barberán, F.A. Description of urolithin production capacity from ellagic acid of two human intestinal Gordonibacter species. Food Funct. 2014, 5, 1779–1784. [Google Scholar] [CrossRef]
- Gaya, P.; Peirotén, Á.; Medina, M.; Álvarez, I.; Landete, J.M. Bifidobacterium pseudocatenulatum INIA P815: The first bacterium able to produce urolithins A and B from ellagic acid. J. Funct. Foods 2018, 45, 95–99. [Google Scholar] [CrossRef]
- Beltrán, D.; Romo-Vaquero, M.; Espín, J.C.; Tomás-Barberán, F.A.; Selma, M.V. Ellagibacter isourolithinifaciens gen. nov., sp. nov., a new member of the family Eggerthellaceae, isolated from human gut. Int. J. Syst. Evol. Microbiol. 2018, 68, 1707–1712. [Google Scholar] [CrossRef]
- Tripathi, M.K.; Giri, S.K. Probiotic functional foods: Survival of probiotics during processing and storage. J. Funct. Foods 2014, 9, 225–241. [Google Scholar] [CrossRef]
- Konuray, G.; Erginkaya, Z. Potential Use of Bacillus coagulans in the Food Industry. Foods 2018, 7, 92. [Google Scholar] [CrossRef] [PubMed]
- Abhari, K.; Shekarforoush, S.S.; Sajedianfard, J.; Hosseinzadeh, S.; Nazifi, S. The Effects of Probiotic, Prebiotic and Synbiotic Diets Containing Bacillus Coagulans and Inulin on Rat Intestinal Microbiota. Iran. J. Vet. Res. 2015, 16, 267–273. [Google Scholar] [PubMed]
- Maathuis, A.J.H.; Keller, D.; Farmer, S. Survival and metabolic activity of the GanedenBC30 strain of Bacillus coagulans in a dynamic in vitro model of the stomach and small intestine. Benef. Microbes 2010, 1, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Li, X.; Song, P.; Xu, L. Optimal cut-off values for the homeostasis model assessment of insulin resistance (HOMA-IR) and pre-diabetes screening: Developments in research and prospects for the future. Drug Discov. Ther. 2015, 9, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-X.; Qin, Y.; Chen, T.; Lu, M.; Qian, X.; Guo, X.; Bai, Y. A practical guide to amplicon and metagenomic analysis of microbiome data. Protein Cell 2021, 12, 315–330. [Google Scholar] [CrossRef]
- Hoffman, C.; Siddiqui, N.Y.; Fields, I.; Gregory, W.T.; Simon, H.M.; Mooney, M.A.; Wolfe, A.J.; Karstens, L. Species-Level Resolution of Female Bladder Microbiota from 16S rRNA Amplicon Sequencing. mSystems 2021, 6, e00518-21. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef] [PubMed]
- Yonker, L.M.; Gilboa, T.; Ogata, A.F.; Senussi, Y.; Lazarovits, R.; Boribong, B.P.; Bartsch, Y.C.; Loiselle, M.; Rivas, M.N.; Porritt, R.A.; et al. Multisystem inflammatory syndrome in children is driven by zonulin-dependent loss of gut mucosal barrier. J. Clin. Investig. 2021, 131, 149633. [Google Scholar] [CrossRef] [PubMed]
- Stevens, B.R.; Goel, R.; Seungbum, K.; Richards, E.M.; Holbert, R.C.; Pepine, C.J.; Raizada, M.K. Increased human intestinal barrier permeability plasma biomarkers zonulin and FABP2 correlated with plasma LPS and altered gut microbiome in anxiety or depression. Gut 2018, 67, 1555–1557. [Google Scholar] [CrossRef] [PubMed]
- Selma, M.V.; Beltrán, D.; Luna, M.C.; Romo-Vaquero, M.; Garcia-Villalba, R.; Mira, A.; Espín, J.C.; Tomás-Barberán, F.A. Isolation of Human Intestinal Bacteria Capable of Producing the Bioactive Metabolite Isourolithin A from Ellagic Acid. Front. Microbiol. 2017, 8, 1521. [Google Scholar] [CrossRef]
- Albasher, G.; Alkahtani, S.; Al-Harbi, L.N. Urolithin A prevents streptozotocin-induced diabetic cardiomyopathy in rats by activating SIRT1. Saudi J. Biol. Sci. 2022, 29, 1210–1220. [Google Scholar] [CrossRef]
- Andreux, P.A.; Blanco-Bose, W.; Ryu, D.; Burdet, F.; Ibberson, M.; Aebischer, P.; Auwerx, J.; Singh, A.; Rinsch, C. The mitophagy activator urolithin A is safe and induces a molecular signature of improved mitochondrial and cellular health in humans. Nat. Metab. 2019, 1, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Daveri, E.; Cremonini, E.; Mastaloudis, A.; Hester, S.N.; Wood, S.M.; Waterhouse, A.L.; Anderson, M.; Fraga, C.G.; Oteiza, P.I. Cyanidin and delphinidin modulate inflammation and altered redox signaling improving insulin resistance in high fat-fed mice. Redox Biol. 2018, 18, 16–24. [Google Scholar] [CrossRef]
- Chiou, W.-C.; Chang, B.-H.; Tien, H.-H.; Cai, Y.-L.; Fan, Y.-C.; Chen, W.-J.; Chu, H.-F.; Chen, Y.-H.; Huang, C. Synbiotic Intervention with an Adlay-Based Prebiotic and Probiotics Improved Diet-Induced Metabolic Disturbance in Mice by Modulation of the Gut Microbiota. Nutrients 2021, 13, 3161. [Google Scholar] [CrossRef]
- Ávalos, Y.; Hernández-Cáceres, M.P.; Lagos, P.; Pinto-Nuñez, D.; Rivera, P.; Burgos, P.; Díaz-Castro, F.; Joy-Immediato, M.; Venegas-Zamora, L.; Lopez-Gallardo, E.; et al. Palmitic acid control of ciliogenesis modulates insulin signaling in hypothalamic neurons through an autophagy-dependent mechanism. Cell Death Dis. 2022, 13, 659. [Google Scholar] [CrossRef] [PubMed]
- Ryu, D.; Mouchiroud, L.; Andreux, P.A.; Katsyuba, E.; Moullan, N.; Nicolet-Dit-Félix, A.A.; Williams, E.G.; Jha, P.; Lo Sasso, G.; Huzard, D.; et al. Urolithin A induces mitophagy and prolongs lifespan in C. elegans and increases muscle function in rodents. Nat. Med. 2016, 22, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-R.; Zhang, M.-H.; Chen, Y.-J.; Sun, Y.-L.; Gao, Z.-M.; Li, Z.-J.; Zhang, G.-P.; Qin, Y.; Dai, X.-Y.; Yu, X.-Y.; et al. Urolithin A ameliorates obesity-induced metabolic cardiomyopathy in mice via mitophagy activation. Acta Pharmacol. Sin. 2022. online ahead of print. [Google Scholar] [CrossRef]
- Tuohetaerbaike, B.; Zhang, Y.; Tian, Y.; Zhang, N.N.; Kang, J.; Mao, X.; Zhang, Y.; Li, X. Pancreas protective effects of Urolithin A on type 2 diabetic mice induced by high fat and streptozotocin via regulating autophagy and AKT/mTOR signaling pathway. J. Ethnopharmacol. 2020, 250, 112479. [Google Scholar] [CrossRef] [PubMed]
- Maruo, T.; Sakamoto, M.; Ito, C.; Toda, T.; Benno, Y. Adlercreutzia equolifaciens gen. nov., sp. nov., an equol-producing bacterium isolated from human faeces, and emended description of the genus Eggerthella. Int. J. Syst. Evol. Microbiol. 2008, 58, 1221–1227. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Martín, A.; García-Villalba, R.; García-Mantrana, I.; Rodríguez-Varela, A.; Romo-Vaquero, M.; Collado, M.C.; Tomás-Barberán, F.A.; Espín, J.C.; Selma, M.V. Urolithins in Human Breast Milk after Walnut Intake and Kinetics of Gordonibacter Colonization in Newly Born: The Role of Mothers’ Urolithin Metabotypes. J. Agric. Food Chem. 2020, 68, 12606–12616. [Google Scholar] [CrossRef] [PubMed]
- García-Villalba, R.; Tomás-Barberán, F.A.; Iglesias-Aguirre, C.E.; Giménez-Bastida, J.A.; González-Sarrías, A.; Selma, M.V.; Espín, J.C. Ellagitannins, urolithins, and neuroprotection: Human evidence and the possible link to the gut microbiota. Mol. Asp. Med. 2022, 101109. [Google Scholar] [CrossRef] [PubMed]
- Abdulrahman, A.O.; Alzubaidi, M.Y.; Nadeem, M.S.; Khan, J.A.; Rather, I.A.; Khan, M.I. Effects of urolithins on obesity-associated gut dysbiosis in rats fed on a high-fat diet. Int. J. Food Sci. Nutr. 2021, 72, 923–934. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Daza, M.-C.; Roquim, M.; Dudonné, S.; Pilon, G.; Levy, E.; Marette, A.; Roy, D.; Desjardins, Y. Berry Polyphenols and Fibers Modulate Distinct Microbial Metabolic Functions and Gut Microbiota Enterotype-Like Clustering in Obese Mice. Front. Microbiol. 2020, 11, 2032. [Google Scholar] [CrossRef]
- Al Khalaf, A.K.; Abdulrahman, A.O.; Kaleem, M.; Nur, S.M.; Asseri, A.H.; Choudhry, H.; Khan, M.I. Comparative Analysis of the Impact of Urolithins on the Composition of the Gut Microbiota in Normal-Diet Fed Rats. Nutrients 2021, 13, 3885. [Google Scholar] [CrossRef]
- Ren, S.; Wang, C.; Chen, A.; Lv, W.; Gao, R. The Probiotic Lactobacillus paracasei Ameliorates Diarrhea Cause by Escherichia coli O(8)via Gut Microbiota Modulation1. Front. Nutr. 2022, 9, 878808. [Google Scholar] [CrossRef]
- Kim, Y.A.; Keogh, J.B.; Clifton, P.M. Probiotics, prebiotics, synbiotics and insulin sensitivity. Nutr. Res. Rev. 2018, 31, 35–51. [Google Scholar] [CrossRef]
- Anhê, F.F.; Nachbar, R.T.; Varin, T.V.; Vilela, V.; Dudonné, S.; Pilon, G.; Fournier, M.; Lecours, M.-A.; Desjardins, Y.; Roy, D.; et al. A polyphenol-rich cranberry extract reverses insulin resistance and hepatic steatosis independently of body weight loss. Mol. Metab. 2017, 6, 1563–1573. [Google Scholar] [CrossRef]
- Urtasun, R.; Díaz-Gómez, J.; Araña, M.; Pajares, M.J.; Oneca, M.; Torre, P.; Jiménez, M.; Munilla, G.; Barajas, M.; Encío, I. A Combination of Apple Vinegar Drink with Bacillus coagulans Ameliorates High Fat Diet-Induced Body Weight Gain, Insulin Resistance and Hepatic Steatosis. Nutrients 2020, 12, 2504. [Google Scholar] [CrossRef]
- Weigert, C.; Hennige, A.M.; Lehmann, R.; Brodbeck, K.; Baumgartner, F.; Schaüble, M.; Häring, H.U.; Schleicher, E.D. Direct Cross-talk of Interleukin-6 and Insulin Signal Transduction via Insulin Receptor Substrate-1 in Skeletal Muscle Cells. J. Biol. Chem. 2006, 281, 7060–7067. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Ishtiaq, S.M.; Rashid, H.; Hussain, Z.; Arshad, M.I.; Khan, J.A. Adiponectin and PPAR: A setup for intricate crosstalk between obesity and non-alcoholic fatty liver disease. Rev. Endocr. Metab. Disord. 2019, 20, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, T.; Yamauchi, T.; Iwabu, M.; Okada-Iwabu, M.; Nakamura, M. Adiponectin, Adiponectin Receptors, and Epigenetic Regulation of Adipogenesis. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Małodobra-Mazur, M.; Cierzniak, A.; Ryba, M.; Sozański, T.; Piórecki, N.; Kucharska, A.Z. Cornus mas L. Increases Glucose Uptake and the Expression of PPARG in Insulin-Resistant Adipocytes. Nutrients 2022, 14, 2307. [Google Scholar] [CrossRef] [PubMed]
- Małodobra-Mazur, M.; Cierzniak, A.; Pawełka, D.; Kaliszewski, K.; Rudnicki, J.; Dobosz, T. Metabolic Differences between Subcutaneous and Visceral Adipocytes Differentiated with an Excess of Saturated and Monounsaturated Fatty Acids. Genes 2020, 11, 1092. [Google Scholar] [CrossRef]
- Zhang, H.; Qi, R.; Zeng, Y.; Tsao, R.; Mine, Y. Chinese Sweet Leaf Tea (Rubus suavissimus) Mitigates LPS-Induced Low-Grade Chronic Inflammation and Reduces the Risk of Metabolic Disorders in a C57BL/6J Mouse Model. J. Agric. Food Chem. 2020, 68, 138–146. [Google Scholar] [CrossRef]
- Kuang, H.; Wei, C.H.; Wang, T.; Eastep, J.; Li, Y.; Chen, G. Vitamin A Status Affects Weight Gain and Hepatic Glucose Metabolism in Rats Fed a High-Fat Diet. Biochem. Cell Biol. 2019, 97, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Y.; Chen, G. Vitamin A status affects the plasma parameters and regulation of hepatic genes in streptozotocin-induced diabetic rats. Biochimie 2017, 137, 1–11. [Google Scholar] [CrossRef]
- Cabrera-Valladares, G.; German, M.S.; Matschinsky, F.M.; Wang, J.; Fernandez-Mejia, C. Effect of Retinoic Acid on Glucokinase Activity and Gene Expression and on Insulin Secretion in Primary Cultures of Pancreatic Islets. Endocrinology 1999, 140, 3091–3096. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Woo, S.M.; Jang, H.; Kang, M.; Kim, S.-Y. Cancer depends on fatty acids for ATP production: A possible link between cancer and obesity. Semin. Cancer Biol. 2022. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | LFD | HFD | HFD+EA | HFD+EA+BC77 | HFD+EA+BC2000 |
|---|---|---|---|---|---|
| Casein | 189.6 | 231 | 231 | 231 | 231 |
| L-Cystine | 2.8 | 3 | 3 | 3 | 3 |
| Corn Starch | 479.8 | 83.9 | 83.6 | 83.5 | 83.5 |
| Maltodextrin | 118.5 | 115 | 115 | 115 | 115 |
| Sucrose | 65.2 | 199.3 | 199.3 | 199.3 | 199.3 |
| Cellulose | 47.4 | 58 | 58 | 58 | 58 |
| Soybean oil | 23.7 | 29 | 29 | 29 | 29 |
| Lard | 19 | 204.7 | 204.7 | 204.7 | 204.7 |
| Mineral Mix | 9.5 | 12 | 12 | 12 | 12 |
| Dicalcium Phosphate | 12.3 | 15 | 15 | 15 | 15 |
| Calcium Carbonate | 5.2 | 6.3 | 6.3 | 6.3 | 6.3 |
| Potassium Citrate·H2O | 15.6 | 19 | 19 | 19 | 19 |
| Vitamin Mix | 9.5 | 12 | 12 | 12 | 12 |
| Choline Bitartrate | 1.9 | 2 | 2 | 2 | 2 |
| Cholesterol | 0 | 9.8 | 9.8 | 9.8 | 9.8 |
| Ellagic acid | 0 | 0 | 0.3 | 0.3 | 0.3 |
| Probiotic powder | 0 | 0 | 0 | 0.1 | 0.1 |
| Total (g) | 1000 | 1000 | 1000 | 1000 | 1000 |
| Calories (kcal/g) | |||||
| Protein | 20% | 20% | 20% | 20% | 20% |
| Carbohydrate | 70% | 35% | 35% | 35% | 35% |
| Fat | 10% | 45% | 45% | 45% | 45% |
| Total | 100 | 100 | 100 | 100 | 100 |
| ID | HFD vs. LFD | HFD+EA vs. HFD | HFD+EA+BC77 vs. HFD | HFD+EA+BC2000 vs. HFD | ||||
|---|---|---|---|---|---|---|---|---|
| log2FC | p | log2FC | p | log2FC | p | log2FC | p | |
| Alloprevotella | 2.87 | <0.01 | –2.57 | <0.01 | –2.18 | 0.02 | –2.55 | 0.01 |
| Muribaculum | –1.42 | 0.03 | 1.40 | 0.04 | 2.13 | <0.01 | 2.28 | <0.01 |
| Limosilactobacillus | 8.08 | <0.01 | –2.43 | <0.01 | –3.55 | <0.01 | ||
| Alistipes | –2.24 | <0.01 | 2.50 | <0.01 | 3.86 | <0.01 | ||
| Akkermansia | –5.89 | <0.01 | ||||||
| Lactobacillus | 6.14 | <0.01 | –2.00 | <0.01 | ||||
| Sphingopyxis | –5.05 | <0.01 | –3.93 | <0.01 | –5.46 | <0.01 | ||
| Ileibacterium | 3.48 | <0.01 | –4.09 | <0.01 | 4.20 | <0.01 | ||
| Helicobacter | –2.21 | 0.02 | 3.68 | <0.01 | 2.39 | 0.03 | 5.03 | <0.01 |
| Sporobacter | 2.86 | <0.01 | 1.88 | 0.02 | 1.92 | 0.04 | ||
| Parasutterella | –2.70 | <0.01 | –2.57 | <0.01 | –2.71 | <0.01 | ||
| Acetatifactor | 3.93 | <0.01 | 4.63 | <0.01 | 4.18 | <0.01 | ||
| Mucispirillum | –2.57 | 0.02 | 2.72 | <0.01 | 3.35 | <0.01 | ||
| Lachnoclostridium | 1.84 | 0.01 | 1.77 | 0.01 | 2.29 | <0.01 | ||
| Weizmannia | 2.70 | <0.01 | 5.47 | <0.01 | ||||
| Gordonibacter | 3.88 | <0.01 | –3.28 | <0.01 | ||||
| Psychrobacter | –3.22 | 0.01 | –3.78 | <0.01 | –2.46 | 0.03 | ||
| Bifidobacterium | 2.60 | <0.01 | –2.05 | <0.01 | –2.67 | <0.01 | ||
| Frisingicoccus | 2.34 | <0.01 | –1.51 | 0.03 | –2.62 | <0.01 | ||
| Anaerosporobacter | 2.54 | <0.01 | –2.15 | <0.01 | –1.86 | 0.01 | ||
| Turicibacter | –3.19 | <0.01 | 2.98 | 0.01 | ||||
| Sporobacter | 2.86 | <0.01 | 1.88 | 0.02 | 1.92 | 0.04 | ||
| Parasutterella | –2.70 | <0.01 | –2.57 | <0.01 | –2.71 | <0.01 | ||
| Acetatifactor | 3.93 | <0.01 | 4.63 | <0.01 | 4.18 | <0.01 | ||
| Amedibacillus | –2.68 | 0.01 | –2.13 | 0.04 | ||||
| ID | HFD vs. LFD | HFD+EA vs. HFD | HFD+EA+BC77 vs. HFD | HFD+EA+BC2000 vs. HFD | ||||
|---|---|---|---|---|---|---|---|---|
| log2FC | p | log2FC | p | log2FC | p | log2FC | p | |
| PPARG | 0.17 | 0.73 | –0.27 | 0.33 | –0.08 | 0.76 | 0.82 | 0.00 |
| PIK3CA | –0.02 | 0.90 | –0.14 | 0.47 | –0.26 | 0.07 | –0.37 | 0.00 |
| PIK3CB | –0.24 | 0.20 | 0.30 | 0.14 | 0.23 | 0.11 | 0.39 | 0.01 |
| PIK3R1 | –0.08 | 0.70 | 0.24 | 0.30 | 0.59 | 0.00 | 0.64 | 0.00 |
| AKT1 | –0.08 | 0.65 | 0.10 | 0.61 | 0.02 | 0.90 | 0.02 | 0.86 |
| AKT3 | –0.10 | 0.74 | –0.07 | 0.79 | 0.12 | 0.58 | 0.06 | 0.78 |
| mTOR | 0.11 | 0.54 | 0.34 | 0.07 | 0.33 | 0.02 | 0.29 | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, L.; Dang, H.; Wu, J.; Yuan, L.; Chen, X.; Yao, J. Weizmannia coagulans BC2000 Plus Ellagic Acid Inhibits High-Fat-Induced Insulin Resistance by Remodeling the Gut Microbiota and Activating the Hepatic Autophagy Pathway in Mice. Nutrients 2022, 14, 4206. https://doi.org/10.3390/nu14194206
Jin L, Dang H, Wu J, Yuan L, Chen X, Yao J. Weizmannia coagulans BC2000 Plus Ellagic Acid Inhibits High-Fat-Induced Insulin Resistance by Remodeling the Gut Microbiota and Activating the Hepatic Autophagy Pathway in Mice. Nutrients. 2022; 14(19):4206. https://doi.org/10.3390/nu14194206
Chicago/Turabian StyleJin, Long, Hongyang Dang, Jinyong Wu, Lixia Yuan, Xiangsong Chen, and Jianming Yao. 2022. "Weizmannia coagulans BC2000 Plus Ellagic Acid Inhibits High-Fat-Induced Insulin Resistance by Remodeling the Gut Microbiota and Activating the Hepatic Autophagy Pathway in Mice" Nutrients 14, no. 19: 4206. https://doi.org/10.3390/nu14194206
APA StyleJin, L., Dang, H., Wu, J., Yuan, L., Chen, X., & Yao, J. (2022). Weizmannia coagulans BC2000 Plus Ellagic Acid Inhibits High-Fat-Induced Insulin Resistance by Remodeling the Gut Microbiota and Activating the Hepatic Autophagy Pathway in Mice. Nutrients, 14(19), 4206. https://doi.org/10.3390/nu14194206