
The Effects of Physical Activity on the Gut Microbiota and the Gut–Brain Axis in Preclinical and Human Models: A Narrative Review

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
Studies Selection
3. Results
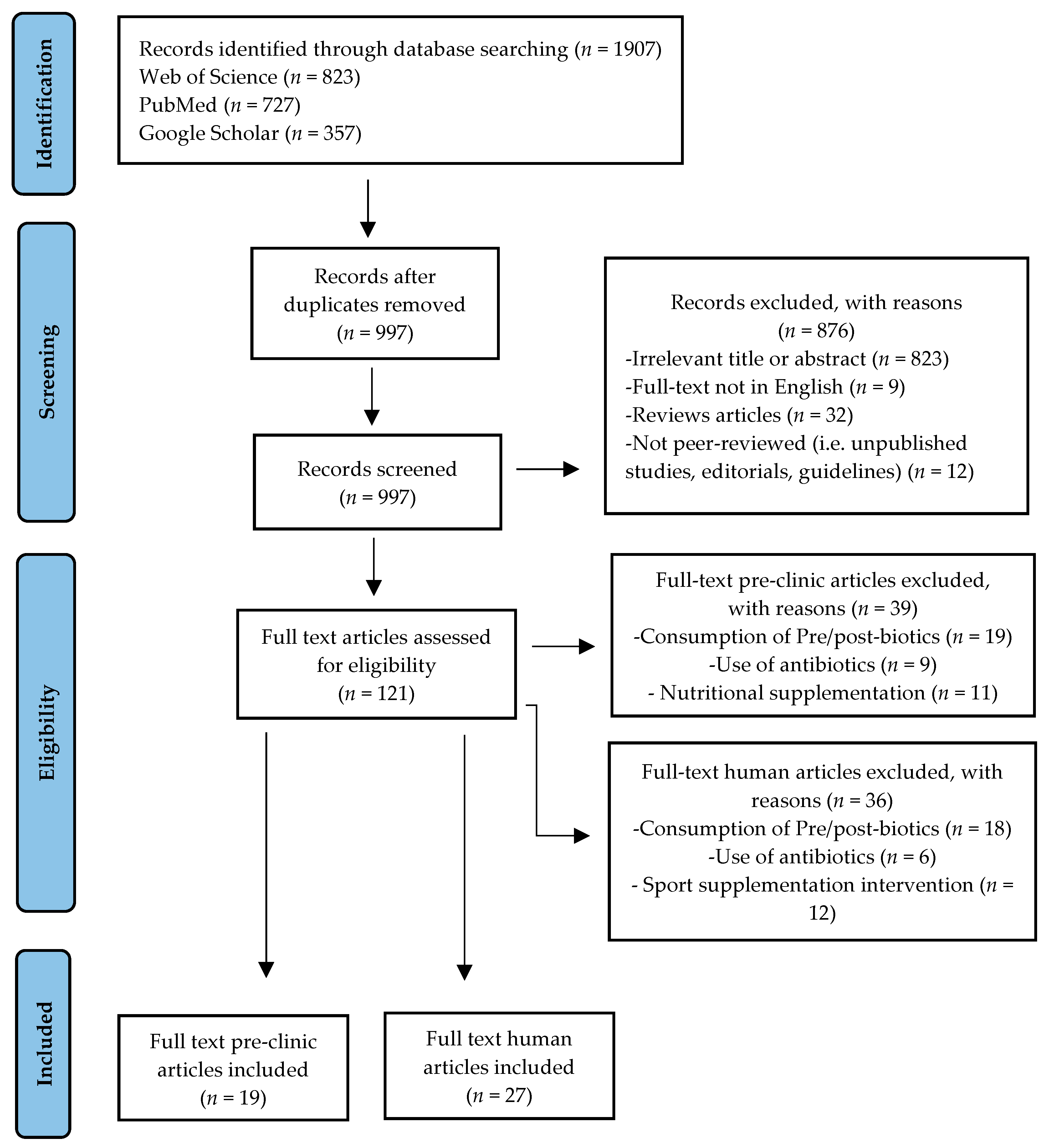
3.1. Identification of Studies
3.2. Studies Characteristics
4. Discussion
4.1. The Gut Microbiota
4.2. Physical Activity and Gut Microbiota in Preclinic Studies
4.3. Voluntary or Forced Exercise?
4.4. Exercise and Host Age
4.5. Exercise, Performance and Overtraining Mediated by Gut Microbiota
4.6. Physical Activity and Gut Microbiota in Human Studies
4.7. Influence of Physical Fitness on the Gut Microbiota
4.8. Endurance Activities and Gut Microbiota
4.9. Physical Activity and Bacterial Changes in the Elderly Population
4.10. Physical Activity, Gut Microbiota and Overweight
4.11. The Gut–Brain Axis Functional Basis and the Impact of Physical Activity
4.12. Physical Activity, Gut Microbiota and Cognitive Ability
5. Limitations and Future Directions
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, G.; Zhao, N.; Zhang, C.; Lam, Y.Y.; Zhao, L. Guild-based analysis for understanding gut microbiome in human health and diseases. Genome Med. 2021, 13, 22. [Google Scholar] [CrossRef]
- Belizário, J.E.; Napolitano, M. Human microbiomes and their roles in dysbiosis, common diseases, and novel therapeutic approaches. Front. Microbiol. 2015, 6, 1050. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.B.; Zhu, X.; Moan, E.; Murff, H.J.; Ness, R.M.; Seidner, D.L.; Sun, S.; Yu, C.; Dai, Q.; Fodor, A.A.; et al. Inter-niche and inter-individual variation in gut microbial community assessment using stool, rectal swab, and mucosal samples. Sci. Rep. 2018, 8, 4139. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- De la Cuesta-Zuluaga, J.; Kelley, S.T.; Chen, Y.; Escobar, J.S.; Mueller, N.T.; Ley, R.E.; McDonald, D.; Huang, S.; Swafford, A.D.; Knight, R.; et al. Age- and Sex-Dependent Patterns of Gut Microbial Diversity in Human Adults. mSystems 2019, 4, e00261-19. [Google Scholar] [CrossRef] [PubMed]
- Di Domenico, M.; Ballini, A.; Boccellino, M.; Scacco, S.; Lovero, R.; Charitos, I.A.; Santacroce, L. The Intestinal Microbiota May Be a Potential Theranostic Tool for Personalized Medicine. J. Pers. Med. 2022, 12, 523. [Google Scholar] [CrossRef]
- González, K.; Fuentes, J.; Márquez, J.L. Physical Inactivity, Sedentary Behavior and Chronic Diseases. Korean J. Fam. Med. 2017, 38, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Booth, F.W.; Laye, M.J.; Lees, S.J.; Rector, R.S.; Thyfault, J.P. Reduced physical activity and risk of chronic disease: The biology behind the consequences. Eur. J. Appl. Physiol. 2008, 102, 381–390. [Google Scholar] [CrossRef]
- Zhao, R.; Bu, W.; Chen, Y.; Chen, X. The Dose-Response Associations of Sedentary Time with Chronic Diseases and the Risk for All-Cause Mortality Affected by Different Health Status: A Systematic Review and Meta-Analysis. J. Nutr. Health Aging 2020, 24, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Cerdá, B.; Pérez, M.; Pérez-Santiago, J.D.; Tornero-Aguilera, J.F.; González-Soltero, R.; Larrosa, M. Gut Microbiota Modification: Another Piece in the Puzzle of the Benefits of Physical Exercise in Health? Front. Physiol. 2016, 7, 51. [Google Scholar] [CrossRef] [PubMed]
- Youngblut, N.D.; Reischer, G.H.; Walters, W.; Schuster, N.; Walzer, C.; Stalder, G.; Ley, R.E.; Farnleitner, A.H. Host diet and evolutionary history explain different aspects of gut microbiome diversity among vertebrate clades. Nat. Commun. 2019, 10, 2200. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.; Mach, N. Exercise-induced stress behavior, gut-microbiota-brain axis and diet: A systematic review for athletes. J. Int. Soc. Sports Nutr. 2016, 13, 43. [Google Scholar] [CrossRef]
- Campaniello, D.; Corbo, M.R.; Sinigaglia, M.; Speranza, B.; Racioppo, A.; Altieri, C.; Bevilacqua, A. How Diet and Physical Activity Modulate Gut Microbiota: Evidence, and Perspectives. Nutrients 2022, 14, 2456. [Google Scholar] [CrossRef] [PubMed]
- Cataldi, S.; Bonavolontà, V.; Poli, L.; Clemente, F.M.; De Candia, M.; Carvutto, R.; Silva, A.F.; Badicu, G.; Greco, G.; Fischetti, F. The Relationship between Physical Activity, Physical Exercise, and Human Gut Microbiota in Healthy and Unhealthy Subjects: A Systematic Review. Biology 2022, 11, 479. [Google Scholar] [CrossRef] [PubMed]
- Hanage, W.P. Microbiology: Microbiome science needs a healthy dose of scepticism. Nature 2014, 512, 247–248. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D. Human gut microbiome: Hopes, threats and promises. Gut 2018, 67, 1716–1725. [Google Scholar] [CrossRef] [PubMed]
- Brooks, A.W.; Priya, S.; Blekhman, R.; Bordenstein, S.R. Gut microbiota diversity across ethnicities in the United States. PLoS Biol. 2018, 16, e2006842. [Google Scholar] [CrossRef]
- Yang, W.; Cong, Y. Gut microbiota-derived metabolites in the regulation of host immune responses and immune-related inflammatory diseases. Cell Mol. Immunol. 2021, 18, 866–877. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, F.; Liu, S.; Liu, G.; Yang, X.; Gao, W.; Fan, K.; Zhao, H.; Ma, J. Significance of gastrointestinal tract in the therapeutic mechanisms of exercise in depression: Synchronism between brain and intestine through GBA. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2020, 103, 109971. [Google Scholar] [CrossRef] [PubMed]
- Dalton, A.; Mermier, C.; Zuhl, M. Exercise influence on the microbiome-gut-brain axis. Gut Microbes 2019, 10, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Narrative Review Checklist. Available online: https://www.elsevier.com/__data/promis_misc/ANDJ%20Narrative%20Review%20Checklist.pdf (accessed on 29 April 2022).
- Clarke, S.F.; Murphy, E.F.; O’Sullivan, O.; Lucey, A.J.; Humphreys, M.; Hogan, A.; Hayes, P.; O’Reilly, M.; Jeffery, I.B.; Wood-Martin, R.; et al. Exercise and associated dietary extremes impact on gut microbial diversity. Gut 2014, 63, 1913–1920. [Google Scholar] [CrossRef] [PubMed]
- Estaki, M.; Pither, J.; Baumeister, P.; Little, J.P.; Gill, S.K.; Ghosh, S.; Ahmadi-Vand, Z.; Marsden, K.R.; Gibson, D.L. Cardiorespiratory fitness as a predictor of intestinal microbial diversity and distinct metagenomic functions. Microbiome 2016, 4, 42. [Google Scholar] [CrossRef] [PubMed]
- Bressa, C.; Bailén-Andrino, M.; Pérez-Santiago, J.; González-Soltero, R.; Pérez, M.; Montalvo-Lominchar, M.G.; Maté-Muñoz, J.L.; Domínguez, R.; Moreno, D.; Larrosa, M. Differences in gut microbiota profile between women with active lifestyle and sedentary women. PLoS ONE 2017, 12, e0171352. [Google Scholar]
- Mörkl, S.; Lackner, S.; Müller, W.; Gorkiewicz, G.; Kashofer, K.; Oberascher, A.; Painold, A.; Holl, A.; Holzer, P.; Meinitzer, A.; et al. Gut microbiota and body composition in anorexia nervosa inpatients in comparison to athletes, overweight, obese, and normal weight controls. Int. J. Eat. Disord. 2017, 50, 1421–1431. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Shi, Y.; Wiklund, P.; Tan, X.; Wu, N.; Zhang, X.; Tikkanen, O.; Zhang, C.; Munukka, E.; Cheng, S. The Association between Cardiorespiratory Fitness and Gut Microbiota Composition in Premenopausal Women. Nutrients 2017, 9, 792. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.; Bautista, E.J.; Nguyen, H.; Hanson, B.M.; Chen, L.; Lek, S.H.; Sodergren, E.; Weinstock, G. Community characteristics of the gut microbiomes of competitive cyclists. Microbiome 2017, 5, 98. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, J.A.; Ptacek, T.S.; Carter, S.J.; Liu, N.; Kumar, R.; Hyndman, L.; Lefkowitz, E.J.; Morrow, C.D.; Rogers, L.Q. Gut microbiota composition associated with alterations in cardiorespiratory fitness and psychosocial outcomes among breast cancer survivors. Supportive Care Cancer 2017, 25, 1563–1570. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, Z.; Hu, B.; Huang, W.; Yuan, C.; Zou, L. Response of Gut Microbiota to Metabolite Changes Induced by Endurance Exercise. Front. Microbiol. 2018, 9, 765. [Google Scholar] [CrossRef] [PubMed]
- Barton, W.; Penney, N.C.; Cronin, O.; Garcia-Perez, I.; Molloy, M.G.; Holmes, E.; Shanahan, F.; Cotter, P.D.; O’Sullivan, O. The microbiome of professional athletes differs from that of more sedentary subjects in composition and particularly at the functional metabolic level. Gut 2018, 67, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.M.; Mailing, L.J.; Niemiro, G.M.; Moore, R.; Cook, M.D.; White, B.A.; Holscher, H.D.; Woods, J.A. Exercise Alters Gut Microbiota Composition and Function in Lean and Obese Humans. Med. Sci. Sports Exerc. 2018, 50, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Munukka, E.; Ahtiainen, J.P.; Puigbó, P.; Jalkanen, S.; Pahkala, K.; Keskitalo, A.; Kujala, U.M.; Pietilä, S.; Hollmén, M.; Elo, L.; et al. Six-Week Endurance Exercise Alters Gut Metagenome That Is not Reflected in Systemic Metabolism in Over-weight Women. Front. Microbiol. 2018, 9, 2323. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, H.; Tanisawa, K.; Sun, X.; Kubo, T.; Hoshino, Y.; Hosokawa, M.; Takeyama, H.; Higuchi, M. Effects of short-term endurance exercise on gut microbiota in elderly men. Physiol. Rep. 2018, 6, e13935. [Google Scholar] [CrossRef] [PubMed]
- Durk, R.P.; Castillo, E.; Márquez-Magaña, L.; Grosicki, G.J.; Bolter, N.D.; Lee, C.M.; Bagley, J.R. Gut Microbiota Composition Is Related to Cardiorespiratory Fitness in Healthy Young Adults. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Scheiman, J.; Luber, J.M.; Chavkin, T.A.; MacDonald, T.; Tung, A.; Pham, L.D.; Wibowo, M.C.; Wurth, R.C.; Punthambaker, S.; Tierney, B.T.; et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nat. Med. 2019, 25, 1104–1109. [Google Scholar] [CrossRef] [PubMed]
- Keohane, D.M.; Woods, T.; O’Connor, P.; Underwood, S.; Cronin, O.; Whiston, R.; O’Sullivan, O.; Cotter, P.; Shanahan, F.; Molloy, M.G.M. Four men in a boat: Ultra-endurance exercise alters the gut microbiome. J. Sci. Med. Sport 2019, 22, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Morita, E.; Yokoyama, H.; Imai, D.; Takeda, R.; Ota, A.; Kawai, E.; Hisada, T.; Emoto, M.; Suzuki, Y.; Okazaki, K. Aerobic Exercise Training with Brisk Walking Increases Intestinal Bacteroides in Healthy Elderly Women. Nutrients 2019, 11, 868. [Google Scholar] [CrossRef] [PubMed]
- Kern, T.; Blond, M.B.; Hansen, T.H.; Rosenkilde, M.; Quist, J.S.; Gram, A.S.; Ekstrøm, C.T.; Hansen, T.; Stallknecht, B. Structured exercise alters the gut microbiota in humans with overweight and obesity-A randomized controlled trial. Int. J. Obes. 2020, 44, 125–135. [Google Scholar] [CrossRef]
- Castellanos, N.; Diez, G.G.; Antúnez-Almagro, C.; Bressa, C.; Bailén, M.; González-Soltero, R.; Pérez, M.; Larrosa, M. Key Bacteria in the Gut Microbiota Network for the Transition between Sedentary and Active Lifestyle. Microorganisms 2020, 8, 785. [Google Scholar] [CrossRef]
- Quiroga, R.; Nistal, E.; Estébanez, B.; Porras, D.; Juárez-Fernández, M.; Martínez-Flórez, S.; García-Mediavilla, M.V.; de Paz, J.A.; González-Gallego, J.; Sánchez-Campos, S.; et al. Exercise training modulates the gut microbiota profile and impairs inflammatory signaling pathways in obese children. Exp. Mol. Med. 2020, 52, 1048–1061. [Google Scholar] [CrossRef] [PubMed]
- Rettedal, E.A.; Cree, J.; Adams, S.E.; MacRae, C.; Skidmore, P.; Cameron-Smith, D.; Gant, N.; Blenkiron, C.; Merry, T.L. Short-term high-intensity interval training exercise does not affect gut bacterial community diversity or composition of lean and overweight men. Exp. Physiol. 2020, 105, 1268–1279. [Google Scholar] [CrossRef] [PubMed]
- Fart, F.; Rajan, S.K.; Wall, R.; Rangel, I.; Ganda-Mall, J.P.; Tingö, L.; Brummer, R.J.; Repsilber, D.; Schoultz, I.; Lindqvist, C.M. Differences in Gut Microbiome Composition between Senior Orienteering Athletes and Community-Dwelling Older Adults. Nutrients 2020, 12, 2610. [Google Scholar] [CrossRef] [PubMed]
- Bycura, D.; Santos, A.C.; Shiffer, A.; Kyman, S.; Winfree, K.; Sutliffe, J.; Pearson, T.; Sonderegger, D.; Cope, E.; Caporaso, J.G. Impact of Different Exercise Modalities on the Human Gut Microbiome. Sports 2021, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhong, F.; Wen, X.; Yang, M.; Lai, H.Y.; Momma, H.; Cheng, L.; Sun, X.; Nagatomi, R.; Huang, C. Effect of an 8-week Exercise Training on Gut Microbiota in Physically Inactive Older Women. Int. J. Sports Med. 2021, 42, 610–623. [Google Scholar] [CrossRef] [PubMed]
- Moitinho-Silva, L.; Wegener, M.; May, S.; Schrinner, F.; Akhtar, A.; Boysen, T.J.; Schaffer, E.; Hansen, C.; Schmidt, T.; Ruhlemann, M.C.; et al. Short-term physical exercise impacts on the human holobiont obtained by a randomised intervention study. BMC Microbiol. 2021, 21, 162. [Google Scholar] [CrossRef] [PubMed]
- Morishima, S.; Aoi, W.; Kawamura, A.; Kawase, T.; Takagi, T.; Naito, Y.; Tsukahara, T.; Inoue, R. Intensive, prolonged exercise seemingly causes gut dysbiosis in female endurance runners. J. Clin. Biochem. Nutr. 2021, 68, 253–258. [Google Scholar] [CrossRef]
- Erlandson, K.M.; Liu, J.; Johnson, R.; Dillon, S.; Jankowski, C.M.; Kroehl, M.; Robertson, C.E.; Frank, D.N.; Tuncil, Y.; Higgins, J.; et al. An exercise intervention alters stool microbiota and metabolites among older, sedentary adults. Ther. Adv. Infect. Dis. 2021, 8, 20499361211027067. [Google Scholar] [CrossRef] [PubMed]
- Šoltys, K.; Lendvorský, L.; Hric, I.; Baranovičová, E.; Penesová, A.; Mikula, I.; Bohmer, M.; Budiš, J.; Vávrová, S.; Grones, J.; et al. Strenuous Physical Training, Physical Fitness, Body Composition and Bacteroides to Prevotella Ratio in the Gut of Elderly Athletes. Front. Physiol. 2021, 12, 670989. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Inoue, R.; Tsukahara, T.; Ushida, K.; Chiji, H.; Matsubara, N.; Hara, H. Voluntary running exercise alters microbiota composition and increases n-butyrate concentration in the rat cecum. Biosci. Biotechnol. Biochem. 2008, 72, 572–576. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.J.; Eum, S.Y.; Rampersaud, E.; Daunert, S.; Abreu, M.T.; Toborek, M. Exercise attenuates PCB-induced changes in the mouse gut microbiome. Environ. Health Perspect. 2013, 121, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Queipo-Ortuño, M.I.; Seoane, L.M.; Murri, M.; Pardo, M.; Gomez-Zumaquero, J.M.; Cardona, F.; Casanueva, F.; Tinahones, F.J. Gut microbiota composition in male rat models under different nutritional status and physical activity and its association with serum leptin and ghrelin levels. PLoS ONE 2013, 8, e65465. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.S.; Jeraldo, P.R.; Kurti, A.; Miller, M.E.; Cook, M.D.; Whitlock, K.; Goldenfeld, N.; Woods, J.A.; White, B.A.; Chia, N.; et al. Diet and exercise orthogonally alter the gut microbiome and reveal independent associations with anxiety and cognition. Mol. Neurodegener. 2014, 9, 36. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.C.; LePard, K.J.; Kwak, J.W.; Stancukas, M.C.; Laskowski, S.; Dougherty, J.; Moulton, L.; Glawe, A.; Wang, Y.; Leone, V.; et al. Exercise prevents weight gain and alters the gut microbiota in a mouse model of high fat diet-induced obesity. PLoS ONE 2014, 9, e92193. [Google Scholar] [CrossRef] [PubMed]
- Petriz, B.A.; Castro, A.P.; Almeida, J.A.; Gomes, C.P.; Fernandes, G.R.; Kruger, R.H.; Pereira, R.W.; Franco, O.L. Exercise induction of gut microbiota modifications in obese, non-obese and hypertensive rats. BMC Genom. 2014, 15, 511. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.M.; Berg Miller, M.E.; Pence, B.D.; Whitlock, K.; Nehra, V.; Gaskins, H.R.; White, B.A.; Fryer, J.D.; Woods, J.A. Voluntary and forced exercise differentially alters the gut microbiome in C57BL/6J mice. J. Appl. Physiol. 2015, 118, 1059–1066. [Google Scholar] [CrossRef]
- Hsu, Y.J.; Chiu, C.C.; Li, Y.P.; Huang, W.C.; Huang, Y.T.; Huang, C.C.; Chuang, H.L. Effect of intestinal microbiota on exercise performance in mice. J. Strength Cond. Res. 2015, 29, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.W.; Park, Y.M.; Holscher, H.D.; Padilla, J.; Scroggins, R.J.; Welly, R.; Britton, S.L.; Koch, L.G.; Vieira-Potter, V.J.; Swanson, K.S. Physical Activity Differentially Affects the Cecal Microbiota of Ovariectomized Female Rats Selectively Bred for High and Low Aerobic Capacity. PLoS ONE 2015, 10, e0136150. [Google Scholar] [CrossRef]
- Mika, A.; Van Treuren, W.; González, A.; Herrera, J.J.; Knight, R.; Fleshner, M. Exercise is More Effective at Altering Gut Microbial Composition and Producing Stable Changes in Lean Mass in Juvenile versus Adult Male F344 Rats. PLoS ONE 2015, 10, e0125889. [Google Scholar]
- Campbell, S.C.; Wisniewski, P.J.; Noji, M.; McGuinness, L.R.; Häggblom, M.M.; Lightfoot, S.A.; Joseph, L.B.; Kerkhof, L.J. The Effect of Diet and Exercise on Intestinal Integrity and Microbial Diversity in Mice. PLoS ONE 2016, 11, e0150502. [Google Scholar] [CrossRef]
- Denou, E.; Marcinko, K.; Surette, M.G.; Steinberg, G.R.; Schertzer, J.D. High-intensity exercise training increases the diversity and metabolic capacity of the mouse distal gut microbiota during diet-induced obesity. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E982–E993. [Google Scholar] [CrossRef] [PubMed]
- Lamoureux, E.V.; Grandy, S.A.; Langille, M. Moderate Exercise Has Limited but Distinguishable Effects on the Mouse Microbiome. mSystems 2017, 2, e00006-17. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Uchida, Y.; Koch, L.; Britton, S.; Hu, J.; Lutrin, D.; Maze, M. Exercise Prevents Enhanced Postoperative Neuroinflammation and Cognitive Decline and Rectifies the Gut Microbiome in a Rat Model of Metabolic Syndrome. Front. Immunol. 2017, 8, 1768. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Xu, S.; Huang, H.; Liang, J.; Wu, Y.; Li, C.; Yuan, H.; Zhao, X.; Lai, X.; Hou, S. Influence of excessive exercise on immunity, metabolism, and gut microbial diversity in an overtraining mice model. Scand. J. Med. Sci. Sports 2018, 28, 1541–1551. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, F.M.; Ribeiro, C.; Ana Claudia, M.G.; Castro, A.P.; Almeida, J.A.; Franco, O.L.; Petriz, B.A. Limited Effects of Low-to-Moderate Aerobic Exercise on the Gut Microbiota of Mice Subjected to a High-Fat Diet. Nutrients 2019, 11, 149. [Google Scholar] [CrossRef] [PubMed]
- Abraham, D.; Feher, J.; Scuderi, G.L.; Szabo, D.; Dobolyi, A.; Cservenak, M.; Juhasz, J.; Ligeti, B.; Pongor, S.; Gomez-Cabrera, M.C.; et al. Exercise and probiotics attenuate the development of Alzheimer’s disease in transgenic mice: Role of microbiome. Exp. Gerontol. 2019, 115, 122–131. [Google Scholar] [CrossRef]
- Leigh, S.J.; Kaakoush, N.O.; Escorihuela, R.M.; Westbrook, R.F.; Morris, M.J. Treadmill exercise has minimal impact on obesogenic diet-related gut microbiome changes but alters adipose and hypothalamic gene expression in rats. Nutr. Metab. 2020, 17, 71. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Liu, A.; Zong, W.; Dai, L.; Liu, Y.; Luo, R.; Ge, S.; Dong, G. Moderate exercise ameliorates osteoarthritis by reducing lipopolysaccharides from gut microbiota in mice. Saudi J. Biol. Sci. 2021, 28, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Sender, R.; Fuchs, S.; Milo, R. Revised estimates for the number of human and bacteria cells in the body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.W.; Hwang, I. Understanding the evolution of endosymbiotic organelles based on the targeting sequences of organellar proteins. New Phytol. 2021, 230, 924–930. [Google Scholar] [CrossRef]
- Zachar, I.; Boza, G. Endosymbiosis before eukaryotes: Mitochondrial establishment in protoeukaryotes. Cell. Mol. Life Sci. CMLS 2020, 77, 3503–3523. [Google Scholar] [CrossRef] [PubMed]
- Gabaldón, T. Relative timing of mitochondrial endosymbiosis and the “pre-mitochondrial symbioses” hypothesis. IUBMB Life 2018, 70, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Ursell, L.K.; Metcalf, J.L.; Parfrey, L.W.; Knight, R. Defining the human microbiome. Nutr. Rev. 2012, 70 (Suppl. S1), S38–S44. [Google Scholar] [CrossRef]
- Gill, S.R.; Pop, M.; Deboy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic analysis of the human distal gut microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef] [PubMed]
- Piccini, F. Alla Scoperta del Microbioma Umano: Flora Batterica, Nutrizione e Malattie del Progresso; Amazon: London, UK, 2017; ISBN 9781521884195. [Google Scholar]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef]
- Sovran, B.; Hugenholtz, F.; Elderman, M.; Van Beek, A.A.; Graversen, K.; Huijskes, M.; Boekschoten, M.V.; Savelkoul, H.; De Vos, P.; Dekker, J.; et al. Age-associated Impairment of the Mucus Barrier Function is Associated with Profound Changes in Microbiota and Immunity. Sci. Rep. 2019, 9, 1437. [Google Scholar] [CrossRef] [PubMed]
- Ballini, A.; Scacco, S.; Boccellino, M.; Santacroce, L.; Arrigoni, R. Microbiota and Obesity: Where Are We Now? Biology 2020, 9, 415. [Google Scholar] [CrossRef]
- Asadi, A.; Shadab Mehr, N.; Mohamadi, M.H.; Shokri, F.; Heidary, M.; Sadeghifard, N.; Khoshnood, S. Obesity and gut-microbiota-brain axis: A narrative review. J. Clin. Lab. Anal. 2022, 36, e24420. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Musso, G.; Gambino, R.; Cassader, M. Obesity, diabetes, and gut microbiota: The hygiene hypothesis expanded? Diabetes Care 2010, 33, 2277–2284. [Google Scholar] [CrossRef] [PubMed]
- Ludewig, G.; Robertson, L.W. Polychlorinated biphenyls (PCBs) as initiating agents in hepatocellular carcinoma. Cancer Lett. 2013, 334, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Rude, K.M.; Pusceddu, M.M.; Keogh, C.E.; Sladek, J.A.; Rabasa, G.; Miller, E.N.; Sethi, S.; Keil, K.P.; Pessah, I.N.; Lein, P.J.; et al. Developmental exposure to polychlorinated biphenyls (PCBs) in the maternal diet causes host-microbe defects in weanling offspring mice. Environ. Pollut. 2019, 253, 708–721. [Google Scholar] [CrossRef] [PubMed]
- Norström, K.; Czub, G.; McLachlan, M.S.; Hu, D.; Thorne, P.S.; Hornbuckle, K.C. External exposure and bioaccumulation of PCBs in humans living in a contaminated urban environment. Environ. Int. 2010, 36, 855–861. [Google Scholar] [CrossRef]
- Boesten, R.J.; de Vos, W.M. Interactomics in the human intestine: Lactobacilli and Bifidobacteria make a difference. J. Clin. Gastroenterol. 2008, 42 Pt 2 (Suppl. S3), S163–S167. [Google Scholar] [CrossRef]
- Cummings, J.H.; Macfarlane, G.T. Role of intestinal bacteria in nutrient metabolism. J. Parenter. Enter. Nutr. 1997, 21, 357–365. [Google Scholar] [CrossRef]
- Zakostelska, Z.; Kverka, M.; Klimesova, K.; Rossmann, P.; Mrazek, J.; Kopecny, J.; Hornova, M.; Srutkova, D.; Hudcovic, T.; Ridl, J.; et al. Lysate of probiotic Lactobacillus casei DN-114 001 ameliorates colitis by strengthening the gut barrier function and changing the gut microenvironment. PLoS ONE 2011, 6, e27961. [Google Scholar] [CrossRef]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.J.; Wells, J.M. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. American journal of physiology. Gastrointest. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef]
- Claus, S.P.; Ellero, S.L.; Berger, B.; Krause, L.; Bruttin, A.; Molina, J.; Paris, A.; Want, E.J.; de Waziers, I.; Cloarec, O.; et al. Colonization-induced host-gut microbial metabolic interaction. mBio 2011, 2, e00271-10. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef]
- Lay, C.; Sutren, M.; Rochet, V.; Saunier, K.; Doré, J.; Rigottier-Gois, L. Design and validation of 16S rRNA probes to enumerate members of the Clostridium leptum subgroup in human faecal microbiota. Environ. Microbiol. 2005, 7, 933–946. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Tailford, L.E.; Owen, C.D.; Walshaw, J.; Crost, E.H.; Hardy-Goddard, J.; Le Gall, G.; de Vos, W.M.; Taylor, G.L.; Juge, N. Discovery of intramolecular trans-sialidases in human gut microbiota suggests novel mechanisms of mucosal adaptation. Nat. Commun. 2015, 6, 7624. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Bäckhed, F.; Fulton, L.; Gordon, J.I. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef]
- Kellermayer, R.; Dowd, S.E.; Harris, R.A.; Balasa, A.; Schaible, T.D.; Wolcott, R.D.; Tatevian, N.; Szigeti, R.; Li, Z.; Versalovic, J.; et al. Colonic mucosal DNA methylation, immune response, and microbiome patterns in Toll-like receptor 2-knockout mice. FASEB J. 2011, 25, 1449–1460. [Google Scholar] [CrossRef]
- Sudo, N.; Chida, Y.; Aiba, Y.; Sonoda, J.; Oyama, N.; Yu, X.N.; Kubo, C.; Koga, Y. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J. Physiol. 2004, 558 Pt 1, 263–275. [Google Scholar] [CrossRef]
- Cox, L.M.; Yamanishi, S.; Sohn, J.; Alekseyenko, A.V.; Leung, J.M.; Cho, I.; Kim, S.G.; Li, H.; Gao, Z.; Mahana, D.; et al. Altering the intestinal microbiota during a critical developmental window has lasting metabolic consequences. Cell 2014, 158, 705–721. [Google Scholar] [CrossRef]
- Wall, R.; Ross, R.P.; Ryan, C.A.; Hussey, S.; Murphy, B.; Fitzgerald, G.F.; Stanton, C. Role of gut microbiota in early infant development. Clinical medicine. Pediatrics 2009, 3, 45–54. [Google Scholar]
- Koenig, J.E.; Spor, A.; Scalfone, N.; Fricker, A.D.; Stombaugh, J.; Knight, R.; Angenent, L.T.; Ley, R.E. Succession of microbial consortia in the developing infant gut microbiome. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4578–4585. [Google Scholar] [CrossRef]
- McNamara, M.P.; Singleton, J.M.; Cadney, M.D.; Ruegger, P.M.; Borneman, J.; Garland, T. Early-life effects of juvenile Western diet and exercise on adult gut microbiome composition in mice. J. Exp. Biol. 2021, 224 Pt 4, jeb239699. [Google Scholar] [CrossRef]
- Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Cheng, J.; Duncan, A.E.; Kau, A.L.; Griffin, N.W.; Lombard, V.; Henrissat, B.; Bain, J.R.; et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 2013, 341, 1241214. [Google Scholar] [CrossRef]
- Wilson, M. Microbial Inhabitants of Humans: Their Ecology and Role in Health and Disease; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Shindo, D.; Matsuura, T.; Suzuki, M. Effects of prepubertal-onset exercise on body weight changes up to middle age in rats. J. Appl. Physiol. 2014, 116, 674–682. [Google Scholar] [CrossRef][Green Version]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef]
- Martarelli, D.; Verdenelli, M.C.; Scuri, S.; Cocchioni, M.; Silvi, S.; Cecchini, C.; Pompei, P. Effect of a probiotic intake on oxidant and antioxidant parameters in plasma of athletes during intense exercise training. Curr. Microbiol. 2011, 62, 1689–1696. [Google Scholar] [CrossRef]
- Lamprecht, M.; Bogner, S.; Schippinger, G.; Steinbauer, K.; Fankhauser, F.; Hallstroem, S.; Schuetz, B.; Greilberger, J.F. Probiotic supplementation affects markers of intestinal barrier, oxidation, and inflammation in trained men; a randomized, double-blinded, placebo-controlled trial. J. Int. Soc. Sports Nutr. 2012, 9, 45. [Google Scholar] [CrossRef]
- Klaenhammer, T.; Altermann, E.; Arigoni, F.; Bolotin, A.; Breidt, F.; Broadbent, J.; Cano, R.; Chaillou, S.; Deutscher, J.; Gasson, M.; et al. Discovering lactic acid bacteria by genomics. Antonie Van Leeuwenhoek 2002, 82, 29–58. [Google Scholar]
- Bouchard, C.; Lesage, R.; Lortie, G.; Simoneau, J.A.; Hamel, P.; Boulay, M.R.; Pérusse, L.; Thériault, G.; Leblanc, C. Aerobic performance in brothers, dizygotic and monozygotic twins. Med. Sci. Sports Exerc. 1986, 18, 639–646. [Google Scholar] [CrossRef]
- Huang, Z.; Kraus, V.B. Does lipopolysaccharide-mediated inflammation have a role in OA? Nature reviews. Rheumatology 2016, 12, 123–129. [Google Scholar]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef]
- World Health Organization. Global Recommendations on Physical Activity for Health; WHO Library Cataloguing-in-Publication Data: Geneva, Switzerland, 2010. [Google Scholar]
- Mahizir, D.; Briffa, J.F.; Wood, J.L.; Anevska, K.; Hill-Yardin, E.L.; Jefferies, A.J.; Gravina, S.; Mazzarino, G.; Franks, A.E.; Moritz, K.M.; et al. Exercise improves metabolic function and alters the microbiome in rats with gestational diabetes. FASEB J. 2020, 34, 1728–1744. [Google Scholar] [CrossRef]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humarán, L.G.; Gratadoux, J.J.; Blugeon, S.; Bridonneau, C.; Furet, J.P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef]
- Canani, R.B.; Costanzo, M.D.; Leone, L.; Pedata, M.; Meli, R.; Calignano, A. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J. Gastroenterol. 2011, 17, 1519–1528. [Google Scholar] [CrossRef]
- Machiels, K.; Joossens, M.; Sabino, J.; De Preter, V.; Arijs, I.; Eeckhaut, V.; Ballet, V.; Claes, K.; Van Immerseel, F.; Verbeke, K.; et al. A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 2014, 63, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Myhrstad, M.; Tunsjø, H.; Charnock, C.; Telle-Hansen, V.H. Dietary Fiber, Gut Microbiota, and Metabolic Regulation-Current Status in Human Randomized Trials. Nutrients 2020, 12, 859. [Google Scholar] [CrossRef]
- Castellanos, N.; Diez, G.G.; Antúnez-Almagro, C.; Bailén, M.; Bressa, C.; González Soltero, R.; Pérez, M.; Larrosa, M. A Critical Mutualism—Competition Interplay Underlies the Loss of Microbial Diversity in Sedentary Lifestyle. Front. Microbiol. 2020, 10, 3142. [Google Scholar] [CrossRef]
- Guinane, C.M.; Cotter, P.D. Role of the gut microbiota in health and chronic gastrointestinal disease: Understanding a hidden metabolic organ. Ther. Adv. Gastroenterol. 2013, 6, 295–308. [Google Scholar] [CrossRef]
- Cella, V.; Migliaccio, S.; Paoli, A. Microbiota intestinale ed esercizio fisico: Nuova possibile area di intervento? L’Endocrinologo 2020, 21, 338–343. [Google Scholar] [CrossRef]
- Samuel, B.S.; Gordon, J.I. A humanized gnotobiotic mouse model of host-archaeal-bacterial mutualism. Proc. Natl. Acad. Sci. USA 2006, 103, 10011–10016. [Google Scholar] [CrossRef] [PubMed]
- Samuel, B.S.; Hansen, E.E.; Manchester, J.K.; Coutinho, P.M.; Henrissat, B.; Fulton, R.; Latreille, P.; Kim, K.; Wilson, R.K.; Gordon, J.I. Genomic and metabolic adaptations of Methanobrevibacter smithii to the human gut. Proc. Natl. Acad. Sci. USA 2007, 104, 10643–10648. [Google Scholar] [CrossRef]
- Kimura, I.; Inoue, D.; Maeda, T.; Hara, T.; Ichimura, A.; Miyauchi, S.; Kobayashi, M.; Hirasawa, A.; Tsujimoto, G. Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41). Proc. Natl. Acad. Sci. USA 2011, 108, 8030–8035. [Google Scholar] [CrossRef]
- Pluznick, J.L.; Protzko, R.J.; Gevorgyan, H.; Peterlin, Z.; Sipos, A.; Han, J.; Brunet, I.; Wan, L.X.; Rey, F.; Wang, T.; et al. Olfactory receptor responding to gut microbiota-derived signals plays a role in renin secretion and blood pressure regulation. Proc. Natl. Acad. Sci. USA 2013, 110, 4410–4415. [Google Scholar] [CrossRef]
- Chambers, E.S.; Byrne, C.S.; Aspey, K.; Chen, Y.; Khan, S.; Morrison, D.J.; Frost, G. Acute oral sodium propionate supplementation raises resting energy expenditure and lipid oxidation in fasted humans. Diabetes Obes. Metab. 2018, 20, 1034–1039. [Google Scholar] [CrossRef]
- Clavel, T.; Lepage, P.; Charrier, C. The family coriobacteriaceae. In The Prokaryotes, 4th ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 201–238. [Google Scholar]
- Tsukahara, T.; Ushida, K. Succinate accumulation in pig large intestine during antibiotic-associated diarrhea and the constitution of succinate-producing flora. J. Gen. Appl. Microbiol. 2002, 48, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Santacruz, A.; Collado, M.C.; García-Valdés, L.; Segura, M.T.; Martín-Lagos, J.A.; Anjos, T.; Martí-Romero, M.; Lopez, R.M.; Florido, J.; Campoy, C.; et al. Gut microbiota composition is associated with body weight, weight gain and biochemical parameters in pregnant women. Br. J. Nutr. 2010, 104, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Tims, S.; Derom, C.; Jonkers, D.M.; Vlietinck, R.; Saris, W.H.; Kleerebezem, M.; de Vos, W.M.; Zoetendal, E.G. Microbiota conservation and BMI signatures in adult monozygotic twins. ISME J. 2013, 7, 707–717. [Google Scholar] [CrossRef]
- Goodrich, J.K.; Waters, J.L.; Poole, A.C.; Sutter, J.L.; Koren, O.; Blekhman, R.; Beaumont, M.; Van Treuren, W.; Knight, R.; Bell, J.T.; et al. Human genetics shape the gut microbiome. Cell 2014, 159, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Schwiertz, A.; Hold, G.L.; Duncan, S.H.; Gruhl, B.; Collins, M.D.; Lawson, P.A.; Flint, H.J.; Blaut, M. Anaerostipes caccae gen. nov., sp. nov., a new saccharolytic, acetate-utilising, butyrate-producing bacterium from human faeces. Syst. Appl. Microbiol. 2002, 25, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Mazidi, M.; Shekoohi, N.; Covic, A.; Mikhailidis, D.P.; Banach, M. Adverse Impact of Desulfovibrio spp. and Beneficial Role of Anaerostipes spp. on Renal Function: Insights from a Mendelian Randomization Analysis. Nutrients 2020, 12, 2216. [Google Scholar] [CrossRef]
- Chen, Y.J.; Wu, H.; Wu, S.D.; Lu, N.; Wang, Y.T.; Liu, H.N.; Dong, L.; Liu, T.T.; Shen, X.Z. Parasutterella, in association with irritable bowel syndrome and intestinal chronic inflammation. J. Gastroenterol. Hepatol. 2018, 33, 1844–1852. [Google Scholar] [CrossRef]
- Feng, Z.; Long, W.; Hao, B.; Ding, D.; Ma, X.; Zhao, L.; Pang, X. A human stool-derived Bilophila wadsworthia strain caused systemic inflammation in specific-pathogen-free mice. Gut Pathog. 2017, 9, 59. [Google Scholar] [CrossRef]
- Baron, E.J. Bilophila wadsworthia: A unique Gram-negative anaerobic rod. Anaerobe 1997, 3, 83–86. [Google Scholar] [CrossRef]
- American College of Sports Medicine; Chodzko-Zajko, W.J.; Proctor, D.N.; Fiatarone Singh, M.A.; Minson, C.T.; Nigg, C.R.; Salem, G.J.; Skinner, J.S. American College of Sports Medicine position stand. Exercise and physical activity for older adults. Med. Sci. Sports Exerc. 2009, 41, 1510–1530. [Google Scholar] [CrossRef]
- Kreznar, J.H.; Keller, M.P.; Traeger, L.L.; Rabaglia, M.E.; Schueler, K.L.; Stapleton, D.S.; Zhao, W.; Vivas, E.I.; Yandell, B.S.; Broman, A.T.; et al. Host Genotype and Gut Microbiome Modulate Insulin Secretion and Diet-Induced Metabolic Phenotypes. Cell Rep. 2017, 18, 1739–1750. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.R.; Pike, C.M.; Parsons, R.J.; Rivera, A.J.; Foley, M.H.; McLaren, M.R.; Montgomery, S.A.; Theriot, C.M. Clostridioides difficile exploits toxin-mediated inflammation to alter the host nutritional landscape and exclude competitors from the gut microbiota. Nat. Commun. 2021, 12, 462. [Google Scholar] [CrossRef] [PubMed]
- Scheepers, L.E.; Penders, J.; Mbakwa, C.A.; Thijs, C.; Mommers, M.; Arts, I.C. The intestinal microbiota composition and weight development in children: The KOALA Birth Cohort Study. Int. J. Obes. 2015, 39, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Baker, S.S.; Gill, C.; Liu, W.; Alkhouri, R.; Baker, R.D.; Gill, S.R. Characterization of gut microbiomes in nonalcoholic steatohepatitis (NASH) patients: A connection between endogenous alcohol and NASH. Hepatology 2013, 57, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Rosenkilde, M.; Petersen, M.B.; Gram, A.S.; Quist, J.S.; Winther, J.; Kamronn, S.D.; Milling, D.H.; Larsen, J.E.; Jespersen, A.P.; Stallknecht, B. The GO-ACTIWE randomized controlled trial—An interdisciplinary study designed to investigate the health effects of active commuting and leisure time physical activity. Contemp. Clin. Trials 2017, 53, 122–129. [Google Scholar] [CrossRef]
- Wang, Y.; Kasper, L.H. The role of microbiome in central nervous system disorders. Brain Behav. Immun. 2014, 38, 1–12. [Google Scholar] [CrossRef]
- Bercik, P.; Park, A.J.; Sinclair, D.; Khoshdel, A.; Lu, J.; Huang, X.; Deng, Y.; Blennerhassett, P.A.; Fahnestock, M.; Moine, D.; et al. The anxiolytic effect of Bifidobacterium longum NCC3001 involves vagal pathways for gut-brain communication. Neurogastroenterol. Motil. 2011, 23, 1132–1139. [Google Scholar] [CrossRef]
- Bercik, P.; Denou, E.; Collins, J.; Jackson, W.; Lu, J.; Jury, J.; Deng, Y.; Blennerhassett, P.; Macri, J.; McCoy, K.D.; et al. The intestinal microbiota affect central levels of brain-derived neurotropic factor and behavior in mice. Gastroenterology 2011, 141, 599–609.e6093. [Google Scholar] [CrossRef]
- Erny, D.; Hrabě de Angelis, A.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef]
- Cowan, C.; Hoban, A.E.; Ventura-Silva, A.P.; Dinan, T.G.; Clarke, G.; Cryan, J.F. Gutsy Moves: The Amygdala as a Critical Node in Microbiota to Brain Signaling. BioEssays News Rev. Mol. Cell. Dev. Biol. 2018, 40, 1700172. [Google Scholar] [CrossRef]
- Sharon, G.; Sampson, T.R.; Geschwind, D.H.; Mazmanian, S.K. The Central Nervous System and the Gut Microbiome. Cell 2016, 167, 915–932. [Google Scholar] [CrossRef] [PubMed]
- Junges, V.M.; Closs, V.E.; Nogueira, G.M.; Gottlieb, M. Crosstalk between Gut Microbiota and Central Nervous System: A Focus on Alzheimer’s Disease. Curr. Alzheimer Res. 2018, 15, 1179–1190. [Google Scholar] [CrossRef] [PubMed]
- Brookes, S.J.; Spencer, N.J.; Costa, M.; Zagorodnyuk, V.P. Extrinsic primary afferent signalling in the gut. Nature reviews. Gastroenterol. Hepatol. 2013, 10, 286–296. [Google Scholar]
- Royes, L. Cross-talk between gut and brain elicited by physical exercise. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165877. [Google Scholar] [CrossRef]
- Grenham, S.; Clarke, G.; Cryan, J.F.; Dinan, T.G. Brain-gut-microbe communication in health and disease. Front. Physiol. 2011, 2, 94. [Google Scholar] [CrossRef]
- Farzi, A.; Fröhlich, E.E.; Holzer, P. Gut Microbiota and the Neuroendocrine System. Neurother. J. Am. Soc. Exp. NeuroTherapeutics 2018, 15, 5–22. [Google Scholar] [CrossRef]
- Jenkins, T.A.; Nguyen, J.C.; Polglaze, K.E.; Bertrand, P.P. Influence of Tryptophan and Serotonin on Mood and Cognition with a Possible Role of the Gut-Brain Axis. Nutrients 2016, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Gubert, C.; Kong, G.; Renoir, T.; Hannan, A.J. Exercise, diet and stress as modulators of gut microbiota: Implications for neurodegenerative diseases. Neurobiol. Dis. 2020, 134, 104621. [Google Scholar] [CrossRef]
- Huang, T.; Larsen, K.T.; Ried-Larsen, M.; Møller, N.C.; Andersen, L.B. The effects of physical activity and exercise on brain-derived neurotrophic factor in healthy humans: A review. Scand. J. Med. Sci. Sports 2014, 24, 1–10. [Google Scholar] [CrossRef]
- Kotekar, N.; Shenkar, A.; Nagaraj, R. Postoperative cognitive dysfunction—Current preventive strategies. Clin. Interv. Aging 2018, 13, 2267–2273. [Google Scholar] [CrossRef]
- Cerovic, M.; Forloni, G.; Balducci, C. Neuroinflammation and the Gut Microbiota: Possible Alternative Therapeutic Targets to Counteract Alzheimer’s Disease? Front. Aging Neurosci. 2019, 11, 284. [Google Scholar] [CrossRef] [PubMed]
- Micheli, L.; Ceccarelli, M.; D’Andrea, G.; Tirone, F. Depression and adult neurogenesis: Positive effects of the antidepressant fluoxetine and of physical exercise. Brain Res. Bull. 2018, 143, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P.; LeVine, H., 3rd. Alzheimer’s disease and the amyloid-beta peptide. J. Alzheimer Dis. 2010, 19, 311–323. [Google Scholar] [CrossRef]
- Lin, T.W.; Shih, Y.H.; Chen, S.J.; Lien, C.H.; Chang, C.Y.; Huang, T.Y.; Chen, S.H.; Jen, C.J.; Kuo, Y.M. Running exercise delays neurodegeneration in amygdala and hippocampus of Alzheimer’s disease (APP/PS1) transgenic mice. Neurobiol. Learn. Mem. 2015, 118, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Barres, B.A. The mystery and magic of glia: A perspective on their roles in health and disease. Neuron 2008, 60, 430–440. [Google Scholar] [CrossRef]
- Shen, T.; Yue, Y.; He, T.; Huang, C.; Qu, B.; Lv, W.; Lai, H.Y. The Association Between the Gut Microbiota and Parkinson’s Disease, a Meta-Analysis. Front. Aging Neurosci. 2021, 13, 636545. [Google Scholar] [CrossRef]
- Zapanta, K.; Schroeder, T.; Fisher, B. Rethinking Parkinson Disease: Exploring Gut-Brain Interactions and the Potential Role of Exercise. Phys. Ther. 2022, 102, pzac022. [Google Scholar] [CrossRef]


{kind=link}
| Authors | Study Design | Sample | Subjects Age (Years) | Type PA | Protocol | Diet Assessment | Duration Intervention | Main Outcomes |
|---|---|---|---|---|---|---|---|---|
| Clarke et al., 2014 [22] | Cross-sectional | n = 86 (M) elite professional rugby players (n = 40) (BMI 29.1 ± 2.9), healthy control (n = 46) (23: BMI ≤ 25—23: BMI > 28) | Elite: 29 (±4) Control: 29 (±6) | Rugby | / | 187-food items FFQ. Macronutrients, fiber, and supplement intake | / | Athletes: ↑ α-diversity, ↑ diversity Firmicutes (phylum), ↑ Prevotella, ↓ Bacteroides, ↓ Lactobacillus Athletes/Low BMI: ↑ Akkermansia (genus) |
| Estaki et al., 2016 [23] | Cross-sectional | n = 39 (M/F) healthy subjects, stratified by CRF (Low; Average; High) | L: 25.5 (±3.3) A: 24.3 (±3.7) H: 26.2 (±5.5) | Aerobic (Mixed activities) | / | 24 h dietary recall interview. Macronutrients, fiber, saturated fat, and PUFA intake | / | VO2 peak positively associated with ↑ GM diversity; ↑ CRF = ↑ taxa producers SCFAs. No differences in α and β-diversity |
| Bressa et al., 2017 [24] | Cross-sectional | n = 40 (F) active (ACT) (n = 19) and sedentary (SED) (n = 21) subjects, defined by WHO recommendations | ACT: 30.7 (±5.9) SED: 32.2 (±8.7) | Aerobic (Mixed activities) | / | 97-food items FFQ. Macronutrients, fiber, and main food intake | / | ACT: PA ↑ health-promoting bacteria (F.prausnitzii, R.hominis, A.muciniphila) SED: ↑ Barnesiellaceae, ↑ Turicibacter, ↓ Cropococcus No differences in α/β-diversity and at phylum level between groups. |
| Mörkl et al., 2017 [25] | Cross-sectional | n = 106 (F) Anorexia nervosa (AN) patients (n = 18), normal weight (NW) (n = 26), overweight (OW) (n = 22), obese (O) (n = 20) and athletes (AT) (n = 20) | 24.5 (±4.6) | Ball sports | / | Two 24 h recalls. Macronutrients, fiber, Vit D, and magnesium intake | / | ↓ GM α-diversity in obese and AN groups compared to athletes. |
| Yang et al., 2017 [26] | Cross-sectional | n = 71 (F) premenopausal with low (L), moderate (M), high (H) CRF | L: 40.4 (36.9–44.0) M: 39.7 (35.5–43.8) H: 30.6 (25.6–35.6) | Aerobic (Mixed activities) | / | 3-days food records (2 weekdays, 1 weekend day). Macronutrients and total energy intake. | / | ↓ Bacteroides and ↑ Eubacterium rectale–clostridium coccoides in Low VO2max compared to High VO2max group. |
| Petersen et al., 2017 [27] | Cross-sectional | n = 33 (M/F) professional (n = 22) and amateur (n = 11) level competitive cyclists | 19–49 (Median age 33) | Cycling | / | Food questionnaire. Macronutrients and alcohol intake. | / | No significant correlations between taxonomic cluster and professional or amateur level. ↑ Prevotella relative abundance in cyclists training >11 h/wk |
| Paulsen et al., 2017 [28] | Pilot study | n = 12 (F) BCS subjects engaging less than 30′ of vigorous or 60′ of moderate-intensity PA per wk in previous 6 months | 55 (±13) | Aerobic (Mixed activities) | Gradually increase participants to ≥150 weekly minutes of moderate intensity. | 3-day diet record | 3 months | Significant differences in β-diversity were found for CRF suggesting changes in specific taxa present (↑ Roseburia ↑ SMB53 subset of family Clostridiaceae) |
| Zhao et al., 2018 [29] | Cross-sectional | n = 20 (M/F) health amateur runner | 31.3 (±6.1) | Endurance | Half-marathon | Dietary questionnaire. Macronutrient intake | / | After running no changes in α-diversity. ↑ Coriobacteriaceae and Succinivibrionaceae families. ↓ Ezakiella and Romboutsia genus, ↑ Coprococcus, Actinobacillus and Ruminococcus genus |
| Barton et al., 2018 [30] | Cross-sectional | n = 86 (M) elite professional athletes (n = 40), healthy control (n = 46) (22: BMI ≤ 25.2–24: BMI ≥ 26.5) | Elite: 29 (±4) Control: 29 (±6) | Rugby | / | 187-food items FFQ. Macronutrients and total energy intake. | / | ↑ Pathways (↑ AA biosynthesis, ↑ carbohydrate metabolism) and ↑ fecal metabolites (microbial produced SCFAs) in athletes |
| Allen et al., 2018 [31] | Longitudinal | n = 32 (M/F) previously sedentary subjects, lean (n = 18) and obese (n = 14) | Lean: 25.1 (±6.52) Obese: 31.14 (±8.57) | Aerobic (cycling or running) | 30′ to 60′ 3 × wk moderate-to-vigorous intensity (60–75% HRR) exercises | 7-days dietary records, 3-days food menu before each fecal collection. Macronutrient, micronutrient, and total energy intake | 6 weeks | No β-diversity differences among groups. ↑ SCFAs producing taxa related to BMI (Fecalibacterium: ↑ lean ↓ obese, Bacteroides: ↓ lean ↑ obese). Changes largely reversed after 6 wk of inactivity. |
| Munukka et al., 2018 [32] | Non-randomized trial | n = 17 (F) sedentary subjects BMI > 27.5 kg/m2 | 36.8 (±3.9) | Endurance (cycling) | 40′ to 60′ 3 × wk exercises, low to moderate intensity | 3-days food records (2 weekdays and 1 weekend day). Macronutrients, fiber, and total energy intake | 6 weeks | ↑ Akkermansia and ↓ Proteobacteria (exercise-responsive taxa). Changes in GM do not affect systemic metabolites. No differences in α-diversity, slight ↑ β-diversity |
| Taniguchi et al., 2018 [33] | Randomized crossover trial | n = 33 (M) elderly Japanese subjects | 62–76 | Endurance (cycling) | 3 × wk ce, 30′ (wk 1/2)—45′ (wk 3/5), with incremental intensity | Self-administered FFQ, semi-weighted 16-days dietary records. Macronutrients and total energy intake. | 5 weeks | No differences in α and β-diversity. ↓ C.difficile, ↑ Oscillospira. Minor changes in GM associated with cardiometabolic risk factors. |
| Durk et al., 2019 [34] | Cross-sectional | n = 37 (M/F) healthy subjects | 25.7 (±2.2) | Aerobic (running) | / | Instructed to follow their normal diet for 7-days and MyFitnessPal app tracking. Macronutrients, fiber, coffee, alcohol, and total energy intake. | / | VO2max positively associated to ↑ Firmicutes:Bacteroidetes ratio. No differences in α and β-diversity. |
| Scheiman et al., 2019 [35] | Cross-sectional | n = 25 (M) subjects. Athletes from the Boston Marathon (n = 15), sedentary controls (n = 10) | / | Endurance | Marathon | questionnaire and daily annotation sheet | / | ↑ Veillonella relative abundance, in marathon runners post marathon, which can positively influence running performance through the conversion of lactate. |
| Keohane et al., 2019 [36] | Observational | n = 4 (M) ultra-endurance athletes | 26.5 (±1.3) | Endurance | Trans-oceanic rowing | FFQ and MyFitnsessPal mobile application | 33-day event and 3 months follow-up | ↑ α-diversity throughout event. ↑ abundance of butyrate producing species (i.e., Roseburia) and species associated with improved metabolic health (Dorea longicatena). Many of the adaptions in GM structure and metaproteomics persisted at 3 months follow-up. |
| Morita et al., 2019 [37] | Non-randomized comparative trial | n = 32 (F) healthy sedentary elderly subjects, trunk muscle (TM) (n = 14) and aerobic exercise (AE) (n = 18) intervention | 70 (66–75) | Aerobic or anaerobic | TM: 1 h weekly resistance training AE: 1 h daily brisk walking ≥ 3 METs | 138-food and beverage items FFQ. Macronutrients, fiber, saturated fat and total energy intake. | 12 weeks | ↑ Bacteroides relative abundance only in the AE group. |
| Kern et al., 2020 [38] | Randomized controlled trial | n = 88 (M/F) overweight/obese subjects, moderate intensity (n = 31) (MOD), vigorous intensity (n = 24) (VIG), bicycling (n = 18) (BIKE), control (n = 14) (CON) | 36 (30; 41) Median (25th percentile; 75th percentile) | Aerobic (MOD&VIG: walking/running, cycling, stepping. BIKE: cycling) | MOD: 5 × wk LTPA at 50% VO2peak VIG: 5 × wk LTPA at 70% VO2peak BIKE: 5 × wk active bicycle commuting to and from work (F: 9–15 km/M: 11–17 km daily), self-selected intensity | Food registrations (3 weekdays—1 weekend day), participants were asked to weigh and register intake of food and beverages. Macronutrients, fiber, and total energy itnake. | 6 months | β-diversity changed in all groups compared to CON, ↑ α-diversity in VIG compared to CON. Decreased heterogeneity in VIG. No genera changed significantly. |
| Catellanos et al., 2020 [39] | Cross-sectional | n = 109 (M/F) healthy subjects, active (n = 64) (ACT) and sedentary (n = 45) (SED), described by WHO recommendations | ACT: 32.17 (±7.40) SED: 33.69 (±7.96) | Aerobic (Mixed activities) | / | 93-food items FFQ. Macronutrients, fiber, ethanol, and total energy intake. | / | GM network of active people has higher efficiency and transmissibility rate. Key bacteria reorganization from ACT to SED: Roseburia fecis, unclassified roseburia spp. Key bacteria reorganization from SED to ACT: unclassified Sutterella spp. |
| Quiroga et al., 2020 [40] | Randomized controlled trial | n = 39 obese pediatric children (n = 25) and healthy control (n = 14) | 7–12 | Endurance plus strength | 2 × wk combined endurance (sprint of 30″ max cadence at 3′30″, 4′30″, 5′30″, and 6′30″) and strength training (30–70% 1 RM) | Nutritional advice for a healthy and balanced diet. | 12 weeks | ↓ Proteobacteria phylum and Gammaproteobacteria class, ↑ Blautia, Dialister and Roseburia genera lead to a GM profile like that of healthy children. |
| Rettedal et al., 2020 [41] | Non-randomized trial | n = 29 (M) overweight (n = 15) and lean (n = 14) subjects | Overweight: 31 (±2) Lean: 29 (±2) | Aerobic (cycling) | 3 × wk ce HIIT, 60″ cycling intervals at VO2peak workload interspersed with 75″ rest, 8 to 12 intervals | FFQ for baseline intake. Instructed to maintain normal dietary pattern. Macronutrients, fiber, saturated fat, PUFA, and total energy intake. | 3 weeks | No differences in α and β-diversity. Significant association between the abundance of bacterial spp. (Coprococcus_3, Blautia, Lachnospiraceae_ge, Dorea) and insulin sensitivity marker in the overweight group. |
| Fart et al., 2020 [42] | Cross-sectional | n = 98 (M/F) older adults. community-dwelling older adults (CDO) (n = 70) and senior orienteers (SO) (n = 28) | CDO: 72 SO: 68.5 | Orienteering | / | FFQ | / | In SO group compared to CDO group: ↑ F.prausnitzii. No enhanced microbial diversity. ↓ Parasutterella excrementihominis and Bilophila wadsworthia, associated with decreased intestinal health. |
| Bycura et al., 2021 [43] | Non-randomized trial | n = 56 (M/F) healthy students, cardiorespiratory exercise (n = 28) (CRE), resistance exercise (n = 28) (RTE) | CRE: 20.54 (1.93) RTE: 21.28 (3.85) | Aerobic or anaerobic | CRE: 1 h, 3 × wk (2-days group cycling, 1-day rotating CRE activity) 60–90% HRmax RTE: 1 h 3 × wk full/lower/upper body at 70–85% 1 RM | Not controlled or recorded. Instructed to maintain their typical dietary practice and report major deviations. | 8 weeks | CRE: initial changes to GM (wk 2, 3) not sustained through or after the intervention. RTE: no changes in microbiome composition. |
| Zhong et al., 2021 [44] | Randomized controlled trial | n = 12 (F) previously inactive older healthy subjects, exercise (n = 6) and control (n = 6) | Exercise: 69.83 (±4.50) Control: 67.50 (±4.28) | Aerobic (stepping) and anaerobic | 1 h 4 × wk combined aerobic and resistance exercises (progressive overload) | Not controlled or recorded | 8 weeks | No changes in α-diversity. ↑ Prevotella, ↑ Verrucomicrobia, ↓ Proteobacteria abundance in the exercise group. |
| Moitinho-Silva et al., 2021 [45] | Randomized controlled trial | n = 36 (M/F) healthy physical inactive subjects, endurance (n = 12) and strength exercises (n = 13) with control (n = 11). Elite athletes for comparison (n = 13) | Endurance: 31.4 (±8.3) Strength: 29.9 (±7.9) Control: 33.4 (±7.9) Elite: 30 (±9.9) | Aerobic or anaerobic | Endurance: 30′ (at least) 3 × wk running Strength: 30′ 3 × wk whole-body hypertrophy strength training | Food questionnaire Elite: no data. Macronutrients, fiber, and total energy intake. | 6 weeks | No specific bacteria changes. GM change patterns largely varied among individuals of the same group. No differences in α-diversity between elite and physical inactive subjects. |
| Morishima et al., 2021 [46] | Cross-sectional | n = 29 (F) subjects. Endurance runner (n = 15) and healthy non-athletic (n = 14) | Runners(R): 20.5 (±1.2) Control(C): 20.9 (±0.3) | Endurance (running) | / | / | / | In ER group: ↑ Haemophilus, Rothia and Ruminococcus gnavus genus, associated with gut inflammation. |
| Erlandson et al., 2021 [47] | Pilot study | n = 15 (M/F) sedentary older adults | 58 (±8.0) | Aerobic (walking) and anaerobic | 20/30′ aerobic exercise + 3 sets × 8 reps of resistance exercise at low intensity: 3 × wk ~50′ session | 3-day diet record. Macronutrient intake | 24 weeks | ↑ Bifidobacterium, Oscillospira and Anaerostipes, associatet to gut health benefits. ↓ Prevotella and Succinivibrio, associated to inflammatory states. |
| Šoltys et al., 2021 [48] | Cross-sectional | n = 22 (M) elderly subjects. Lifetime endurance athletes (n = 13) and healthy control who met ACSM PA recommendation (n = 9) | LA: 63.5 CTRL: 64.9 | Endurance (cycling) | / | 24 h dietary recording over five consecutive days | / | In LA group comparet to CTRL group: no differences in α-diversity. ↓ Bacteroidetes genus ↑ Prevotella genus. |
| Authors | Study Design | Sample | Species | Type PA | Protocol | Duration Intervention | Main Outcomes |
|---|---|---|---|---|---|---|---|
| Matsumoto et al., 2008 [49] | Randomized block design | n = 14 (M) exercise (n = 7) and sedentary control (n = 7) | Wistar rats (6 wk old) | Aerobic | VWR | 5 wk | VWR group: ↑ Butyrate (SCFA); ↑ Butyrate-producing bacteria, phylum Firmicutes (SM/11, T2-87) |
| Choi et al., 2013 [50] | Randomized controlled trial | n = 12 (M) exercise (n = 6) and sedentary control (n = 6) | C57BL/6 mice (11–13 months old) | Aerobic | VWR | 5 wk | VWR group: ↑ phylum Firmicutes (i.e., lactobacillales order), ↓ phyla Tenericutes and Bacteroidetes. Changes in microbiota induced by PCBs exposure were attenuated. |
| Queipo-Ortuño et al., 2013 [51] | Case-control study | n = 40 (M) ABA exercise (n = 10) ABA sed (n = 10) AL exercise (n = 10) AL sed (n = 10) | Sprague-Dawley rats (5 wk old) | Aerobic | VWR | 6 days | AL exercise group: ↑ Lactobacillus, Bifidobacterium and Blautia. ↑ Organic acid lactate converted in butyrate (SCFA) ↓ Clostridium and Enterococcus. |
| Kang et al., 2014 [52] | Randomized controlled trial | n = 40 (M) ND (n = 10) ND exercise (n = 10) HFD (n = 10) and HFD exercise (n = 10) | Wild-type mice (8 wk old) | Aerobic | FWR 1 h at 7 m/min × 5 days/wk | 16 wk | Exercise alone caused great changes in gut microbiota: ↑ Firmicutes, Proteobacteria and Actinobacteria phyla. ↓ Bateroidetes phylum. |
| Evans et al., 2014 [53] | Randomized controlled trial | n = 48 (M) LF sed (n = 12) LF exercise (n = 12) HF sed (n = 12) HF exercise (n = 12) | Wild-type mice (5 wk old) | Aerobic | VWR | 12 wk | Exercise induced unique change in gut microbiota: ↑ Bacteroidetes and ↓ Firmicutes phylum; ↓ Actinobacteria preventing DIO |
| Petriz et al., 2014 [54] | Prospective cohort study | n = 15 Obese (n = 5), hypertensive (n = 5) and high blood pressure (n = 5) | Wistar rats | Aerobic | Treadmill 30′/day × 5/days/wk. speed progressively increased | 5 wk | In hypertensive: ↑ Frimicutes ↓ Proteobacteria ↑ Lactobacillus ↑ Allobaculum. In obese: ↑ Pseudomonas and lactobacillus. In all groups: ↑ Firmicutes and ↓ Proteobacteria |
| Allen et al., 2015 [55] | Randomized controlled trial | n = 29 (M) VWR (n = 10) FTR (n = 10) sed control (n = 9) | C57BL/6J mice | Aerobic | VWR vs. FTR (40′ × 5 days/wk) | 6 wk | In VWR group: ↓ Turicibacter In both groups: ↔ Bacteroidetes and Firmicutes, ↓ bacterial richness. |
| Hsu et al., 2015 [56] | Prospective cohort study | n = 24 (M) SPF (n = 8) GF (n = 8) and BF (n = 8) gnotobiotic mice | C57BL/6JNarl mice (12 wk old) | Endurance | Swimming | / | In GF and BF groups: ↓ SCFAs ↓ Gpx and ↓ CAT. Gut microbial status can be crucial for physical performance linked to antioxidant enzyme systems. |
| Liu et al., 2015 [57] | Prospective cohort study | n = 30 (F) HCR-EX (n = 8) HCR-SED (n = 7) LCR-EX (n = 8) LCR-SED (n = 7) OVX rats | (26 wk old) | Aerobic | VWR | 11 wk | In HCR-ex group: ↑ Firmicutes In LCR-ex group: ↓ Firmicutes, ↔ Bacteroidetes. |
| Mika et al., 2015 [58] | Randomized controlled trial | n = 40 (M) juvenile (n = 20) and adults (n = 20) | F344 rats | Aerobic | VWR | 6 wk | In juvenile rats compared to adults: ↓ Firmicutes ↑ Bacteroidetes ↑ Bacterial genera. |
| Campbell et al., 2016 [59] | Case-control study | N = 36 (M) LS (n = 9), DIOS (n = 9) Lex (n = 9) and DIOex (n = 9) mice | C57BL/6NTac mice (6 wk old) | Aerobic | VWR | 12 wk | In DIOex and Lex groups: ↑ Fecalibacterium prausnitzii |
| Denou et al., 2016 [60] | Randomized controlled trial | n = 16 (M) exercise (n = 8) and untrained (n = 8) mice | C57 BL/6 mice (8 wk old) | Aerobic (HIIT) | 1 h of treadmill running 3 days/wk | 6 wk | In exercise group: ↓ Firmicutes:Bacteroidetes ratio. |
| Lamoureux et al., 2017 [61] | Prospective cohort study | n = 42 (M/F) voluntary exercise (n = 10), forced exercise (n = 11) and non-exercise control (n = 21) | C57BL/6 mice (6/10 wk old) | Aerobic | VWR vs. FTR | 8 wk | In all groups: ↑ Rikenellaceae and Lachnospiraceae; ↔ species richness. |
| Feng et al., 2017 [62] | Randomized controlled trial | n = 14 (M) Surgery (n = 6) and Sham (n = 8) group | HCR and LCR rats | Aerobic | Treadmill | 6 wk | In LCR: ↑ α-diversity In LCR and HCR: ↑ β-diversity In HCR: ↑ Firmicutes and ↓ Bacteroidetes. Exercise prevented POCD |
| Yuan et al., 2018 [63] | Randomized controlled trial | n = 20 (M) excessive swimming (ES) (n = 10) and non-swimming (NS) (n = 10) | Kunming (KM) mice (6 wk old) | Endurance | Swimming until exhaustion | 4 wk | In ES group: ↓ microbial diversity; ↓ Bacteroidales (S24-7) and Lachnospiraceae; ↑ Helicobacteraceae family. ↑ Helicobater and Bacteroides, ↓ Odoribater genus. |
| Ribeiro et al., 2019 [64] | Randomized controlled trial | n = 40 (M) SDC (n = 10) SDT (n = 10) HFC (n = 10) and HFT (n = 10) | C57BL6 mice | Aerobic | 30′ treadmill running 5 days/wk | 8 wk | In HFT group: ↑ Proteus genus In HFT and SDT: ↑ Vagococcus genera. No significant changes in gut microbiota structure. |
| Abraham et al., 2019 [65] | Prospective cohort study | n = 32 (M) exercise (n = 6) Fram (n = 6) and Combined (n = 6) and control (n = 14) | APP/PS1 transgenic mice | Aerobic | Treadmill 4 days/wk × 60′ | 20 wk | In exercise group: ↑ butyrate bacterial producing (Marvinbryantia formatexigens and Butyrivibrio pro teoclasticus) ↑ Clostridium Eubacterium and Roseburia; ↓ L. johnsonii. |
| Leigh et al., 2020 [66] | Case-control study | n = 48 (M) Csed (n = 12) Cex (n = 12) Cafsed (n = 12) Cafex (n = 12) | Sprague-Dawley rats (6/7 wk old) | Aerobic | FTR | 4 wk | In all groups: No significant change in the overall composition of gut microbiome |
| Li et al., 2021 [67] | Case-control study | n = 54 (M) SDsed (n = 6) SDex (n = 6) and HFsed (n = 6) HFex (n = 6) | C57BL/6J mice (12 wk old) | Aerobic | VWR | 4 wk | In SD groups: ↔ richness In HFex group: ↑ richness; ↓ Firmicutes:Bacteroidetes ratio; ↑ Bacteroidales S24-7; ↑ Prevotellaceae ↑ Bifidobacteriaceae. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cataldi, S.; Poli, L.; Şahin, F.N.; Patti, A.; Santacroce, L.; Bianco, A.; Greco, G.; Ghinassi, B.; Di Baldassarre, A.; Fischetti, F. The Effects of Physical Activity on the Gut Microbiota and the Gut–Brain Axis in Preclinical and Human Models: A Narrative Review. Nutrients 2022, 14, 3293. https://doi.org/10.3390/nu14163293
Cataldi S, Poli L, Şahin FN, Patti A, Santacroce L, Bianco A, Greco G, Ghinassi B, Di Baldassarre A, Fischetti F. The Effects of Physical Activity on the Gut Microbiota and the Gut–Brain Axis in Preclinical and Human Models: A Narrative Review. Nutrients. 2022; 14(16):3293. https://doi.org/10.3390/nu14163293
Chicago/Turabian StyleCataldi, Stefania, Luca Poli, Fatma Neşe Şahin, Antonino Patti, Luigi Santacroce, Antonino Bianco, Gianpiero Greco, Barbara Ghinassi, Angela Di Baldassarre, and Francesco Fischetti. 2022. "The Effects of Physical Activity on the Gut Microbiota and the Gut–Brain Axis in Preclinical and Human Models: A Narrative Review" Nutrients 14, no. 16: 3293. https://doi.org/10.3390/nu14163293
APA StyleCataldi, S., Poli, L., Şahin, F. N., Patti, A., Santacroce, L., Bianco, A., Greco, G., Ghinassi, B., Di Baldassarre, A., & Fischetti, F. (2022). The Effects of Physical Activity on the Gut Microbiota and the Gut–Brain Axis in Preclinical and Human Models: A Narrative Review. Nutrients, 14(16), 3293. https://doi.org/10.3390/nu14163293