
Anti-Aging Effect of Dietary Fiber Compound Mediated by Guangxi Longevity Dietary Pattern on Natural Aging Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of CDFC
2.2. Animals and Experimental Design
2.3. Behavioral Tests
2.4. Sample Collection and Processing
2.5. Measurement of Oxidation-Related Biomarkers and Inflammatory Cytokines
2.6. Brain Tissue Section Observation
2.7. Investigation of Gut Microbiota
2.7.1. Extraction of Mouse Fecal DNA
2.7.2. Mouse Fecal DNA PCR Reaction
2.8. Statistical Analysis of Date
3. Results
3.1. Effect of CDFC on Body Weight and Organ Coefficients in Aging Mice
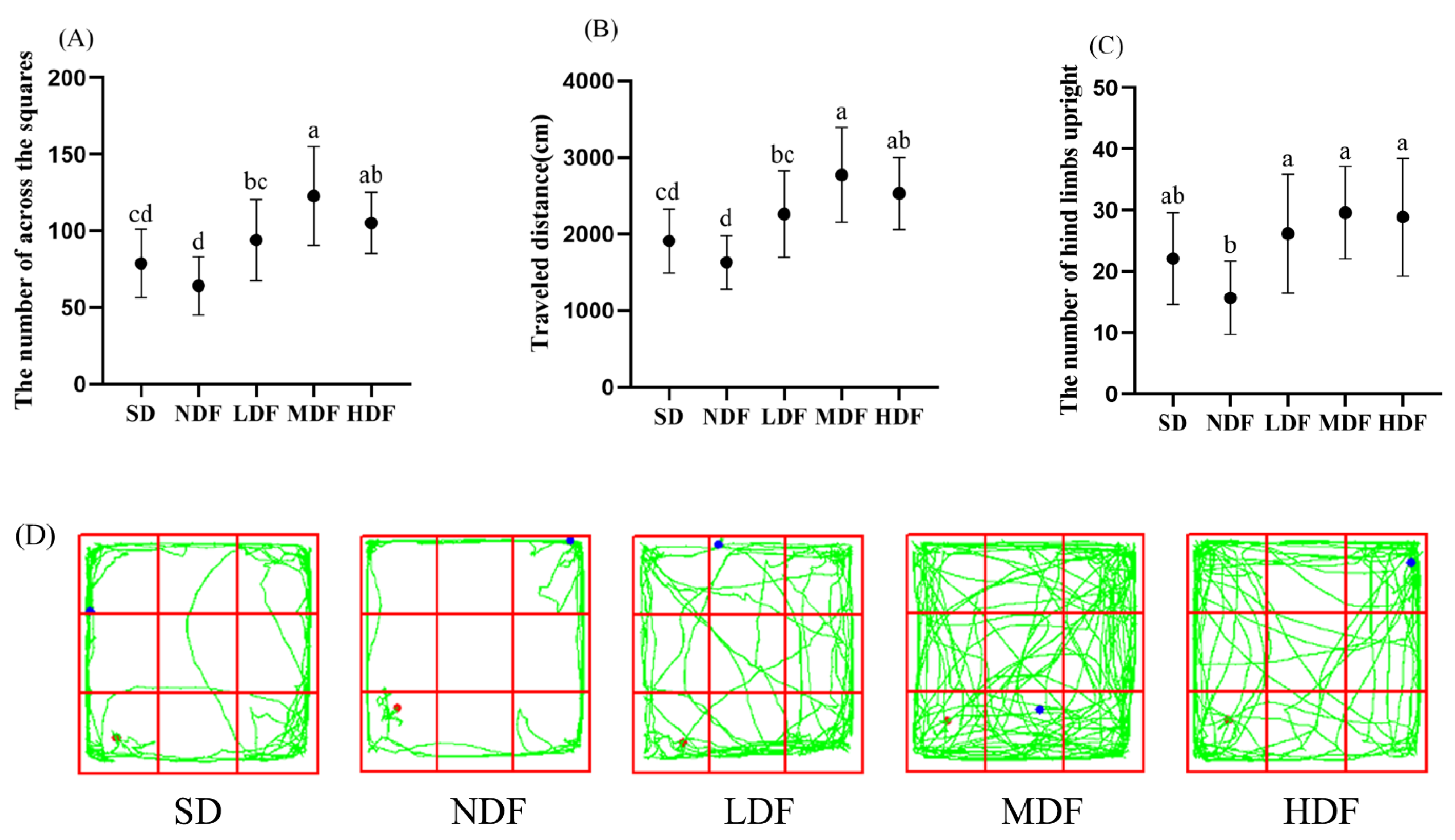
3.2. Effects of CDFC on Anxiety-like Behavior in Field Tests
3.3. Effect of CDFC on the Morris Water Maze Test
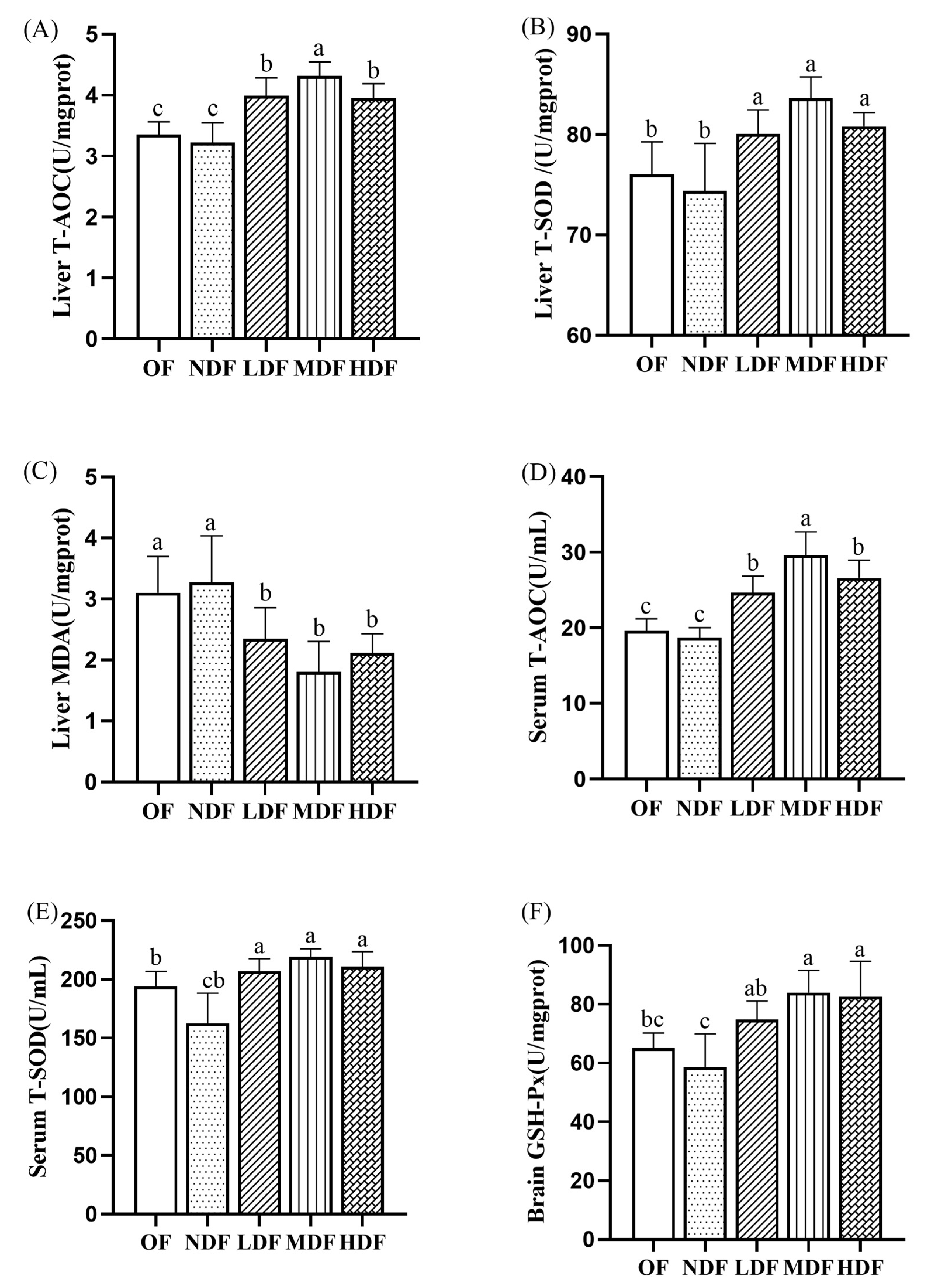
3.4. Effect of CDFC on Antioxidant Capacity
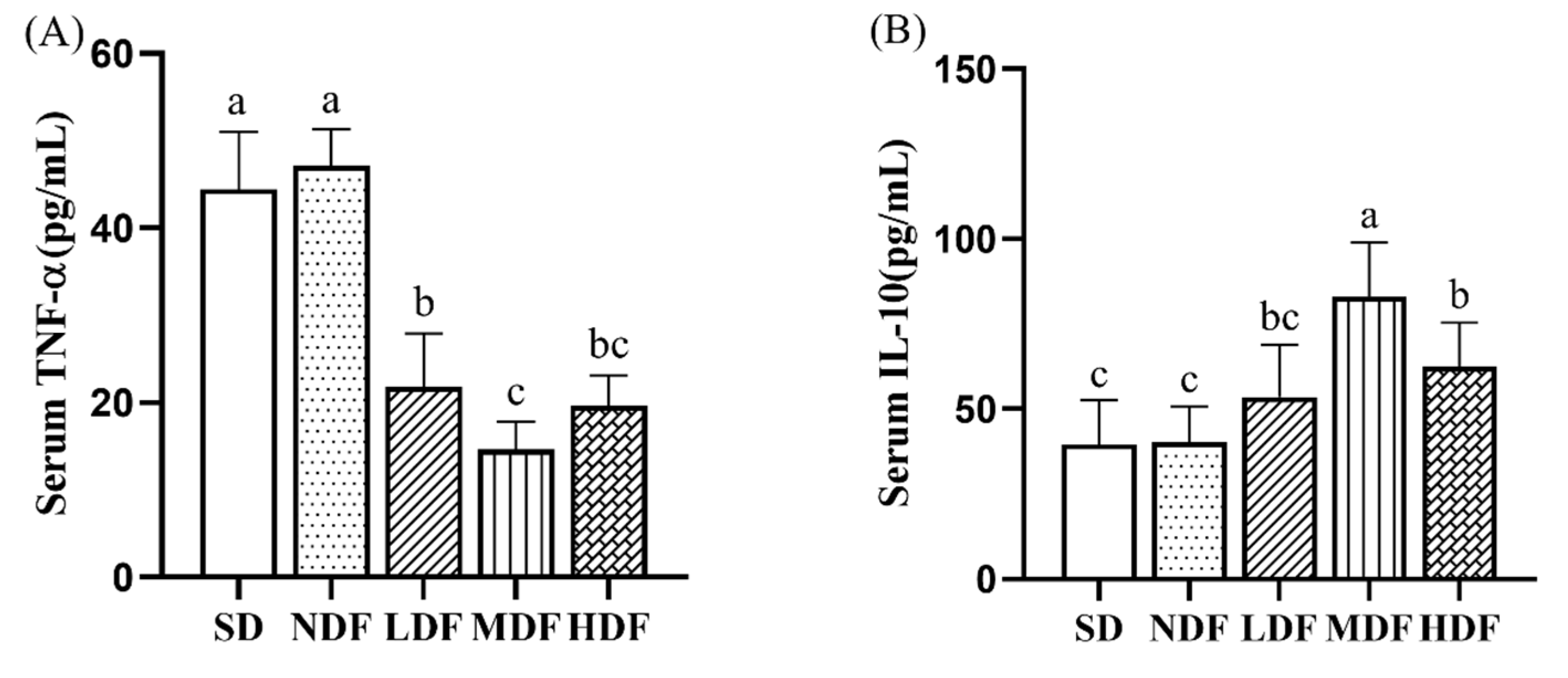
3.5. Effect of CDFC on Serum Inflammatory Markers in Aging Mice
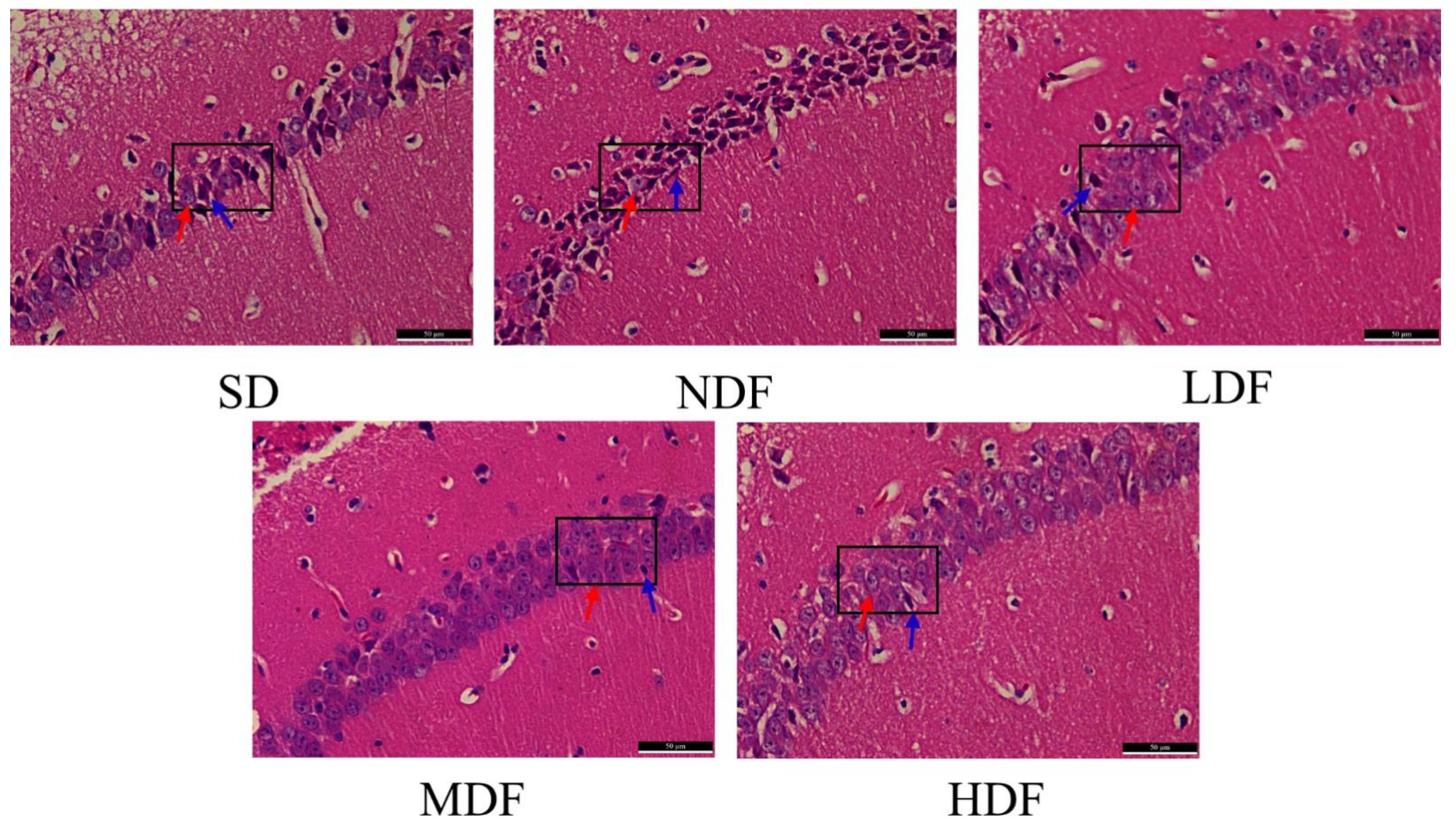
3.6. Protective Effect of CDFC on the Hippocampal Region of Aging Mice
3.7. Perturbation of Fecal Typical Microorganisms by CDFC Intervention
3.8. Interaction of Fecal Microbes with Behavioral, Antioxidant Indicators and Inflammatory Markers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dominguez, L.J.; Nicola, V. Healthy Aging and Dietary Patterns. Nutrients 2022, 14, 889. [Google Scholar] [CrossRef]
- Foundation, B.N. Healthy Ageing: The Role of Nutrition and Lifestyle. Nutr. Food Sci. 2010, 40, 142–143. [Google Scholar] [CrossRef]
- Kaye, D.M.; Shihata, W.A. Deficiency of Prebiotic Fiber and Insufficient Signaling Through Gut Metabolite-Sensing Receptors Leads to Cardiovascular Disease. Circulation 2020, 141, 1393–1403. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.K.; Deehan, E.C. Gut microbiota modulation with long-chain corn bran arabinoxylan in adults with overweight and obesity is linked to an individualized temporal increase in fecal propionate. Microbiome 2020, 8, 118. [Google Scholar] [CrossRef] [PubMed]
- Akihito, N.; Naoko, K. Maternal High Fiber Diet during Pregnancy and Lactation Influences Regulatory T Cell Differentiation in Offspring in Mice. J. Immunol. 2017, 199, 3516–3524. [Google Scholar]
- Daniel, S.; Kevin, W. Dietary fiber intervention on gut microbiota composition in healthy adults: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2018, 107, 965–983. [Google Scholar]
- Assmann, K.E.; Lassale, C. A Healthy Dietary Pattern at Midlife, Combined with a Regulated Energy Intake, Is Related to Increased Odds for Healthy Aging. J. Nutr. 2015, 145, 2139–2145. [Google Scholar] [CrossRef] [Green Version]
- Gunaranjan, P.; Suman, M. Gut microbiota responses to dietary fibre sources in rats fed starch-based or quasi-human background diets. J. Funct. Foods 2021, 83, 104565. [Google Scholar]
- Pontifex, M.G.; Aleena, M. Differential Influence of Soluble Dietary Fibres on Intestinal and Hepatic Carbohydrate Response. Nutrients 2021, 13, 4278. [Google Scholar] [CrossRef]
- Noye Tuplin, E.W.; Erna, A. Dietary fiber combinations to mitigate the metabolic, microbial, and cognitive imbalances resulting from diet-induced obesity in rats. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2022, 36, e22269. [Google Scholar] [CrossRef]
- Tian, T.; Xiaobo, Z. Effects of Short-Term Dietary Fiber Intervention on Gut Microbiota in Young Healthy People. Diabetes Metab. Syndr. Obes. Targets Ther. 2021, 14, 3507–3516. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Zhao, S. Nutrient Intake Is Associated with Longevity Characterization by Metabolites and Element Profiles of Healthy Centenarians. Nutrients 2016, 8, 564. [Google Scholar] [CrossRef] [Green Version]
- Da, C.; Danlei, L. A correlation between diet and longevity characterization by means of element profiles in healthy people over 80 years from a Chinese longevous region. Biol. Trace Elem. Res. 2015, 165, 18–29. [Google Scholar]
- Shivali, P.; Narinder, K. Utilization of fruits and vegetable by-products for isolation of dietary fibres and its potential application as functional ingredients. Bioact. Carbohydr. Diet. Fibre 2022, 27, 100295. [Google Scholar]
- Chuanyan, R.; Xiaohan, Y. Antioxidant Effect of Dietary Fiber in the Diet of Longevity Population on C57BL/6 Mice. Sci. Technol. Food Ind. 2021, 42, 342–349. [Google Scholar]
- Yanting, H.; Lihua, M. Anti-aging Effect of Bama Longevity Characteristic Dietary Patterns in Naturally Aging Mice. Food Sci. 2021, 42, 137–144. [Google Scholar]
- Qi, S.; Lianzhong, A. Effect of Bama Longevity Dietary Patterns on Antioxidant Stress in a Mouse Model of Aging. Food Sci. 2018, 39, 147–153. [Google Scholar]
- Hélène, A.; Valérie, B. Synergistic Effects of Licorice Root and Walnut Leaf Extracts on Gastrointestinal Candidiasis, Inflammation and Gut Microbiota Composition in Mice. Microbiol. Spectr. 2022, 10, e02355-21. [Google Scholar]
- Yanming, W.; Abduweli, N. Effects of composite probiotics isolated from fermented camel milk on intestinal microbiota in db/db diabetic mice. Chin. J. Microecol. 2019, 31, 1365–1371. [Google Scholar]
- Jing, W.; Yong, C. Assessing the Effects of Ginger Extract on Polyphenol Profiles and the Subsequent Impact on the Fecal Microbiota by Simulating Digestion and Fermentation In Vitro. Nutrients 2020, 12, 3194. [Google Scholar] [CrossRef]
- Tingting, S.; Rongbei, L. Altered Intestinal Microbiota with Increased Abundance of Prevotella Is Associated with High Risk of Diarrhea-Predominant Irritable Bowel Syndrome. Gastroent. Res. Pract. 2018, 2018, 6961783. [Google Scholar] [CrossRef] [Green Version]
- Nelson, E.A.; Palombo, E.A. Comparison of methods for the extraction of bacterial DNA from human faecal samples for analysis by real-time PCR. Curr. Res. Technol. Educ. Top. Appl. Microbiol. Microb. Biotechnol. 2010, 2, 1479–1485. [Google Scholar]
- Tatem, K.S.; Quinn, J.L. Behavioral and Locomotor Measurements Using an Open Field Activity Monitoring System for Skeletal Muscle Diseases. J. Vis. Exp. Jove 2014, 163, 51785. [Google Scholar] [CrossRef] [Green Version]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Open Field Test for Measuring Locomotor Activity and Anxiety-like Behavior; Humana Press: New York, NY, USA, 2019; pp. 99–103. [Google Scholar]
- Jie, Z.; Zhewen, C. Anti-aging effects of a functional food via the action of gut microbiota and metabolites in aging mice. Aging 2021, 13, 17880–17900. [Google Scholar]
- Yuka, M.; Yasuhiko, S. The Role of Underweight in Active Life Expectancy among Older Adults in Japan. J. Gerontol. Ser. B Psychol. Sci. Soc. Sci. 2020, 76, 756–765. [Google Scholar]
- Leigh, L.; Byles, J.E. BMI and healthy life expectancy in old and very old women. Brit. J. Nutr. 2016, 116, 692–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.; Ying, Z.P. Dihydromyricetin Improves Cognitive Impairments in D-galactose-Induced Aging Mice through Regulating Oxidative Stress and Inhibition of Acetylcholinesterase. Mol. Nutr. Food Res. 2021, 66, e2101002. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Chen, F. Antioxidative, anti-inflammatory and anti-apoptotic effects of ellagic acid in liver and brain of rats treated by D-galactose. Sci. Rep. 2018, 8, 1465. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Tang, Y. Changes in hippocampal synapses and learning-memory abilities in age-increasing rats and effects of tetrahydroxystilbene glucoside in aged rats. Neuroscience 2007, 149, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Hongli, S.; Xing, G. A fiber-deprived diet causes cognitive impairment and hippocampal microglia-mediated synaptic loss through the gut microbiota and metabolites. Microbiome 2021, 9, 223. [Google Scholar]
- Dorostkar, M.M.; Zou, C.; Blazquez-Llorca, L.; Herms, J. Analyzing dendritic spine pathology in Alzheimer’s disease: Problems and opportunities. Acta Neuropathol. 2015, 130, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Pietrelli, A.; Lopez-Costa, J. Aerobic exercise prevents age-dependent cognitive decline and reduces anxiety-related behaviors in middle-aged and old rats. Neuroscience 2012, 202, 252–266. [Google Scholar] [CrossRef] [PubMed]
- Bunce, D.; Batterham, P.J. Depression, anxiety and cognition in community-dwelling adults aged 70 years and over. J. Psychiatr. Res. 2012, 46, 1662–1666. [Google Scholar] [CrossRef] [PubMed]
- Yanying, W.; Yuying, L. Elucidating the Effects of a Cryptomeria fortunei Forest Environment on Overall Health Based on Open Field Testing of Mice. Sustainability 2021, 14, 59. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, J. Association between Dietary Fiber Intake and Incidence of Depression and Anxiety in Patients with Essential Hypertension. Nutrients 2021, 13, 4159. [Google Scholar] [CrossRef] [PubMed]
- Parladore, S.Y.; Andressa, B. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar]
- Xu, B.; Tu, J. Silybum marianum oil attenuates oxidative stress and ameliorates mitochondrial dysfunction in mice treated with D-galactose. Pharmacogn. Mag. 2014, 10, S92–S99. [Google Scholar]
- Ma, W.; Wei, S. Antioxidant Effect of Polygonatum sibiricum Polysaccharides in D-Galactose-Induced Heart Aging Mice. BioMed Res. Int. 2021, 2021, 6688855. [Google Scholar] [CrossRef]
- Pérez-Jiménez, J.; Díaz-Rubio, M.E. Non-extractable polyphenols, a major dietary antioxidant: Occurrence, metabolic fate and health effects. Nutr. Res. Rev. 2013, 26, 118–129. [Google Scholar] [CrossRef] [Green Version]
- Krajka-Kuźniak, V.; Paluszczak, J. The Nrf2-ARE signaling pathway: An update on its regulation and possible role in cancer prevention and treatment. Pharmacol. Rep. 2017, 69, 393–402. [Google Scholar] [CrossRef]
- Kurek-Gorecka, A.; Rzepecka-Stojko, A. Structure and Antioxidant Activity of Polyphenols Derived from Propolis. Molecules 2014, 19, 78–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elisa, F.; Yang, A. Aging and the burden of multimorbidity: Associations with inflammatory and anabolic hormonal biomarkers. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2015, 70, 63–70. [Google Scholar]
- Michael, L.; Satya, J. Effect of whole grains on markers of subclinical inflammation. Nutr. Rev. 2012, 70, 387–396. [Google Scholar]
- Jiao, J.; Xu, J.Y.; Zhang, W.; Han, S.; Qin, L.Q. Effect of dietary fiber on circulating C-reactive protein in overweight and obese adults: A meta-analysis of randomized controlled trials. Int. J. Food Sci. Nutr. 2015, 66, 114–119. [Google Scholar] [CrossRef]
- Matt, S.M.; Allen, J.M. Butyrate and Dietary Soluble Fiber Improve Neuroinflammation Associated with Aging in Mice. Front. Immunol. 2018, 9, 1832. [Google Scholar] [CrossRef]
- Zhao, Y.; Jayachandran, M. In vivo antioxidant and anti-inflammatory effects of soluble dietary fiber Konjac glucomannan in type-2 diabetic rats. Int. J. Biol. Macromol. 2020, 159, 1186–1196. [Google Scholar] [CrossRef] [PubMed]
- Veiga, S.A.; Maria, T.C. Effect of resistance training on inflammatory markers of older adults: A meta-analysis. Exp. Gerontol. 2018, 111, 188–196. [Google Scholar]
- Liu, J.; Hefni, M.E. Effects of Whole Brown Bean and Its Isolated Fiber Fraction on Plasma Lipid Profile, Atherosclerosis, Gut Microbiota, and Microbiota-Dependent Metabolites in Apoe−/− Mice. Nutrients 2022, 14, 937. [Google Scholar] [CrossRef] [PubMed]
- Christin, H.; Sebastian, P. Effects of polydextrose supplementation on different faecal parameters in healthy volunteers. Int. J. Food Sci. Nutr. 2009, 60 (Suppl. S5), 96–105. [Google Scholar]
- Slavin, J. Fiber and Prebiotics: Mechanisms and Health Benefits. Nutrients 2013, 5, 1417–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ian, R.; Glenn, G. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar]
- Melo, C.B.; Abdalla, S.M.J. Influence of gut microbiota on subclinical inflammation and insulin resistance. Mediat. Inflamm. 2013, 2013, 986734. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Primer Sequence (5′–3′) | Annealing Temperature | Reference |
|---|---|---|---|
| Total intestinal flora | F: 5′-ACTCCTACGGGAGGCAGCAG-3′ R: 5′-ATTACCGCGGCTGCTGG-3′ | 60 °C | Hélène A.et al. [18] |
| Bacteroides | F: 5′-CTGAACCAGCCAAGTAGCG-3′ R: 5′-CCGCAAACTTTCACAACTGACTTA-3′ | 62 °C | Yanming W.et al. [19] |
| Bifidobacterium | F: 5′-TCGCGTCCGGTGTGAAAG-3′ R: 5′-CCACATCCAGCATCCAC-3′ | 58 °C | Jing, W.et al. [20] |
| Lactobacillus | F: 5′-AGCAGTAGGGAATCTTCCA-3′ R: 5′-CACCGCTACACATGGAG-3′ | 58 °C | Tingting, S.et al. [21] |
| Escherichia coli | F: 5′-GTTAATACCTTTGCTCATTGA-3′ R: 5′-ACCAGGGTATCTTAATCCTGTT-3′ | 60 °C | Nelson, E. A. et al. [22] |
| Group | Intervention Time/Week | ||||
|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | |
| SD | 31.18 ± 5.13 | 29.5 ± 3.98 | 28.76 ± 3.57 b | 29.24 ± 3.6 b | 28.78 ± 4.69 b |
| NDF | 30.9 ± 4.34 B | 32.85 ± 4.07 AB | 32.73 ± 3.83 aAB | 33.29 ± 4.06 aAB | 35.37 ± 5.08 abA |
| LDF | 30.79 ± 3.00 B | 31.93 ± 3.32 AB | 31.75 ± 2.76 abAB | 32.05 ± 3.14 abAB | 34.48 ± 3.51 aA |
| MDF | 31.47 ± 3.37 AB | 32.04 ± 3.94 AB | 31.36 ± 3.89 abB | 31.94 ± 3.75 abAB | 34.98 ± 3.41 aA |
| HDF | 31.39 ± 5.78 | 31.66 ± 5.48 | 31.05 ± 4.87 ab | 31.22 ± 4.89 ab | 32.03 ± 5.78 a |
| Group | Organ Index | ||||
|---|---|---|---|---|---|
| Heart | Liver | Spleen | Kidney | Brain | |
| SD | 0.46 ± 0.03 b | 3.98 ± 0.39 | 0.34 ± 0.07 b | 1.20 ± 0.14 | 1.59 ± 0.16 b |
| NDF | 0.45 ± 0.03 ab | 3.92 ± 0.39 | 0.30 ± 0.10 a | 1.10 ± 0.11 | 1.48 ± 0.21 ab |
| LDF | 0.48 ± 0.03 a | 4.00 ± 0.39 | 0.37 ± 0.0.08 a | 1.22 ± 0.10 | 1.68 ± 0.22 a |
| MDF | 0.51 ± 0.05 a | 4.11 ± 0.40 | 0.40 ± 0.09 a | 1.35 ± 0.31 | 1.83 ± 0.16 a |
| HDF | 0.50 ± 0.03 ab | 4.19 ± 0.37 | 0.39 ± 0.08 ab | 1.28 ± 0.15 | 1.80 ± 0.22 b |
| Group | Escape Latency(s) | |||
|---|---|---|---|---|
| Day 1 | Day 2 | Day 3 | Day 4 | |
| SD | 57.70 ± 4.87 | 53.74 ± 7.12 | 51.11 ± 9.64 | 44.05 ± 14.32 a |
| NDF | 58.07 ± 3.83 | 54.38 ± 9.14 | 52.82 ± 9.43 | 46.90 ± 10.38 a |
| LDF | 57.87 ± 3.88 | 51.94 ± 9.82 | 46.95 ± 14.05 | 34.30 ± 9.86 b** |
| MDF | 57.48 ± 4.29 | 50.56 ± 6.22 * | 42.44 ± 9.96 * | 28.70 ± 10.85 b** |
| HDF | 58.05 ± 2.74 | 51.38 ± 9.58 * | 46.19 ± 7.55 * | 30.54 ± 9.01 b** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.; Liang, X.; Han, K.; Shi, F.; Meng, N.; Li, Q. Anti-Aging Effect of Dietary Fiber Compound Mediated by Guangxi Longevity Dietary Pattern on Natural Aging Mice. Nutrients 2022, 14, 3181. https://doi.org/10.3390/nu14153181
Yu X, Liang X, Han K, Shi F, Meng N, Li Q. Anti-Aging Effect of Dietary Fiber Compound Mediated by Guangxi Longevity Dietary Pattern on Natural Aging Mice. Nutrients. 2022; 14(15):3181. https://doi.org/10.3390/nu14153181
Chicago/Turabian StyleYu, Xiaohan, Xiaolin Liang, Kunchen Han, Fengcui Shi, Ning Meng, and Quanyang Li. 2022. "Anti-Aging Effect of Dietary Fiber Compound Mediated by Guangxi Longevity Dietary Pattern on Natural Aging Mice" Nutrients 14, no. 15: 3181. https://doi.org/10.3390/nu14153181
APA StyleYu, X., Liang, X., Han, K., Shi, F., Meng, N., & Li, Q. (2022). Anti-Aging Effect of Dietary Fiber Compound Mediated by Guangxi Longevity Dietary Pattern on Natural Aging Mice. Nutrients, 14(15), 3181. https://doi.org/10.3390/nu14153181