
Glucose as a Potential Key to Fuel Inflammation in Rheumatoid Arthritis


{kind=link}
Abstract
1. Introduction
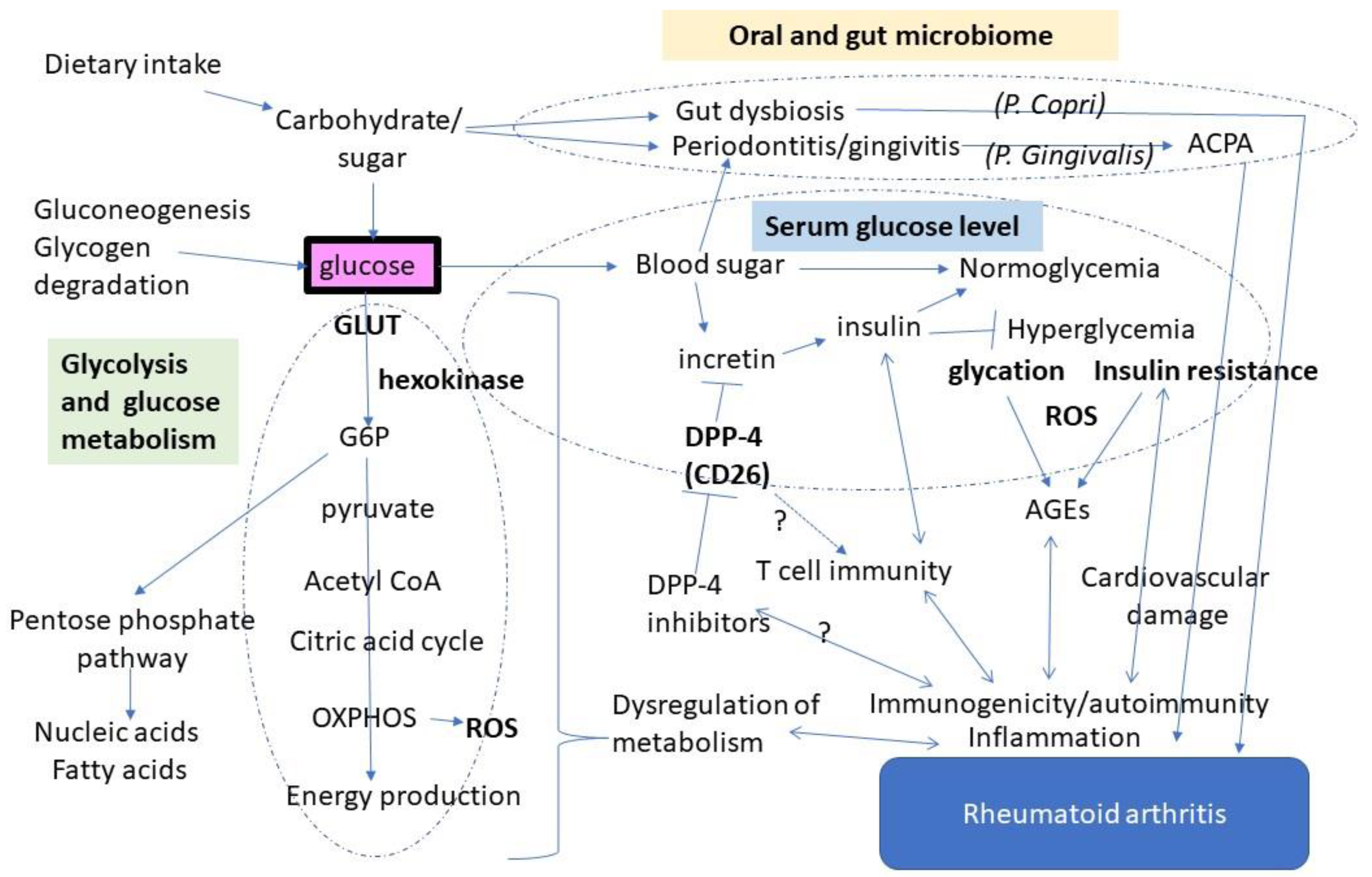
2. Glucose Metabolism Shifts to Glycolysis in Rheumatoid Synovium
3. Glucose May Enhance Autoimmune Responses via Antigen Modification
4. Diabetes and Autoimmunity: Is There a Link through DPP-4?
5. Modulation of the Gut and Oral Microbiome by Glucose
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Scheepers, A.; Joost, H.G.; Schürmann, A. The glucose transporter families SGLT and GLUT: Molecular basis of normal and aberrant function. JPEN J. Parenter. Enter. Nutr. 2004, 28, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Soto-Heredero, G.; Gómez de Las Heras, M.M.; Gabandé-Rodríguez, E.; Oller, J.; Mittelbrunn, M. Glycolysis—A key player in the inflammatory response. FEBS J. 2020, 287, 3350–3369. [Google Scholar] [CrossRef] [PubMed]
- Warmoes, M.O.; Locasale, J.W. Heterogeneity of glycolysis in cancers and therapeutic opportunities. Biochem. Pharm. 2014, 92, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Cairns, R.A. Drivers of the Warburg phenotype. Cancer J. 2015, 21, 56–61. [Google Scholar] [CrossRef]
- Gill, K.S.; Fernandes, P.; O’Donovan, T.R.; McKenna, S.L.; Doddakula, K.K.; Power, D.G.; Soden, D.M.; Forde, P.F. Glycolysis inhibition as a cancer treatment and its role in an anti-tumour immune response. Biochim. Biophys. Acta. 2016, 1866, 87–105. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, M.F.; Garcia-Carbonell, R.; Whisenant, K.D.; Guma, M. Fibroblast-like synoviocyte metabolism in the pathogenesis of rheumatoid arthritis. Arthritis Res. 2017, 19, 110. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Carbonell, R.; Divakaruni, A.S.; Lodi, A.; Vicente-Suarez, I.; Saha, A.; Cheroutre, H.; Boss, G.R.; Tiziani, S.; Murphy, A.N.; Guma, M. Critical role of glucose metabolism in rheumatoid arthritis fibroblast-like synoviocytes. Arthritis Rheumatol. 2016, 68, 1614–1626. [Google Scholar] [CrossRef]
- Bustamante, M.F.; Oliveira, P.G.; Garcia-Carbonell, R.; Croft, A.P.; Smith, J.M.; Serrano, R.L.; Sanchez-Lopez, E.; Liu, X.; Kisseleva, T.; Hay, N.; et al. Hexokinase 2 as a novel selective metabolic target for rheumatoid arthritis. Ann. Rheum. Dis. 2018, 77, 1636–1643. [Google Scholar] [CrossRef]
- Biniecka, M.; Canavan, M.; McGarry, T.; Gao, W.; McCormick, J.; Cregan, S.; Gallagher, L.; Smith, T.; Phelan, J.J.; Ryan, J.; et al. Dysregulated bioenergetics: A key regulator of joint inflammation. Ann. Rheum. Dis. 2016, 75, 2192–2200. [Google Scholar] [CrossRef]
- Koedderitzsch, K.; Zezina, E.; Li, L.; Herrmann, M.; Biesemann, N. TNF induces glycolytic shift in fibroblast like synoviocytes via GLUT1 and HIF1A. Sci. Rep. 2021, 11, 19385. [Google Scholar] [CrossRef]
- Kvacskay, P.; Yao, N.; Schnotz, J.H.; Scarpone, R.; Carvalho, R.A.; Klika, K.D.; Merkt, W.; Tretter, T.; Lorenz, H.M.; Tykocinski, L.O. Increase of aerobic glycolysis mediated by activated T helper cells drives synovial fibroblasts towards an inflammatory phenotype: New targets for therapy? Arthritis Res. 2021, 23, 56. [Google Scholar] [CrossRef] [PubMed]
- Papachristoforou, E.; Lambadiari, V.; Maratou, E.; Makrilakis, K. Association of glycemic indices (hyperglycemia, glucose variability, and hypoglycemia) with oxodative stress and diabetic compliations. J. Diabetee Res. 2020, 7489795. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Collier, B.; Dossett, L.A.; May, A.K.; Diaz, J.J. Glucose control and the inflammatory response. Nutr. Clin. Pr. 2008, 23, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Luc, K.; Schramm-Luc, A.; Guzik, T.J.; Mikolajczyk, T.P. Oxidative stress and inflammatory markers in prediabetes and diabetes. J. Physiol. Pharm. 2019, 70, 809–824. [Google Scholar]
- Chu, C.Q. Highlights of strategies targeting fibroblasts for novel therapies for rheumatoid arthritis. Front. Med. 2022, 9, 846300. [Google Scholar] [CrossRef]
- Monu, P.; Agnihotri, P.; Biswas, S. AGE/non-AGE glycation: An important event in rheumatoid arthritis pathophysiology. Inflammation 2022, 45, 477–496. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, L.; Regueiro, C.; Amhaz-Escanlar, S.; Pena, C.; Herbello-Hermelo, P.; Moreda-Pineiro, A.; Rodriguez-Garcia, J.; Mera-Varela, A.; Perez-Pampin, E.; Gonzalez, A. Antibodies against 4 atypical post-translational protein modifications in patients with rheumatoid arthritis. Diagnostics 2022, 12, 352. [Google Scholar] [CrossRef]
- Kwon, E.J.; Ju, J.H. Impact of posttranslational modification in pathogenesis of rheumatoid arthritis: Focusing on citrullination, carbamylation, and acetylation. Int. J. Mol. Sci. 2021, 22, 10576. [Google Scholar] [CrossRef]
- Trouw, L.A.; Rispens, T.; Toes, R.E.M. Beyond citrullination: Other post-translational protein modifications in rheumatoid arthritis. Nat. Rev. Rheumatol. 2017, 13, 331–339. [Google Scholar] [CrossRef]
- Rantapää-Dahlqvist, S.; de Jong, B.A.; Berglin, E.; Hallmans, G.; Wadell, G.; Stenlund, H.; Sundin, U.; van Venrooij, W.J. Antibodies against cyclic citrullinated peptide and IgA rheumatoid factor predict the development of rheumatoid arthritis. Arthritis Rheum. 2003, 48, 2741–2749. [Google Scholar] [CrossRef]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Mel’nikova, T.I.; Porozov, Y.B.; Terentiev, A.A. Oxidative stress and advanced lipoxidation and glycation end products (ALEs and AGEs) in aging and age-related diseases. Oxid. Med. Cell. Longev. 2019, 2019, 3085756. [Google Scholar] [CrossRef] [PubMed]
- Egawa, T.; Hayashi, T. Association of glycative stress with motor and muscle function. Front. Physiol. 2022, 13, 855358. [Google Scholar] [CrossRef] [PubMed]
- Vytásek, R.; Sedová, L.; Vilím, V. Increased concentration of two different advanced glycation end-products detected by enzyme immunoassays with new monoclonal antibodies in sera of patients with rheumatoid arthritis. BMC Musculoskelet Disord. 2010, 11, 83. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.W.A.; Al Otaibi, A.; Sherwani, S.; Khan, W.A.; Alshammari, E.M.; Al-Zahrani, S.A.; Saleem, M.; Khan, S.N.; Alouffi, S. Glycation and oxidative stress increase autoantibodies in the elderly. Molecules 2020, 25, 3675. [Google Scholar] [CrossRef]
- Ahmad, S.; Moinuddin, A.A. Immunological studies on glycated human IgG. Life Sci. 2012, 90, 980–987. [Google Scholar] [CrossRef]
- Albano, S.A.; Santana-Sahagun, E.; Weisman, M.H. Cigarette smoking and rheumatoid arthritis. Semin. Arthritis Rheum. 2001, 31, 146–159. [Google Scholar] [CrossRef]
- Sugiyama, D.; Nishimura, K.; Tamaki, K.; Tsuji, G.; Nakazawa, T.; Morinobu, A.; Kumagai, S. Impact of smoking as a risk factor for developing rheumatoid arthritis: A meta-analysis of observational studies. Ann. Rheum. Dis. 2010, 69, 70–81. [Google Scholar] [CrossRef]
- Yoshida, K.; Wang, J.; Malspeis, S.; Marchand, N.; Lu, B.; Prisco, L.C.; Martin, L.W.; Ford, J.A.; Costenbader, K.H.; Karlson, E.W.; et al. Passive smoking throughout the life course and the risk of incident rheumatoid arthritis in adulthood among women. Arthritis Rheumatol. 2021, 73, 2219–2228. [Google Scholar] [CrossRef]
- Sharma, H.; Sahlot, R.; Purwar, N.; Garg, U.; Saran, S.; Sharma, B.; Mathur, S.K. Co-existence of type 1 diabetes and other autoimmune ailments in subjects with autoimmune thyroid disorders. Diabetes Metab. Syndr. 2022, 16, 102405. [Google Scholar] [CrossRef]
- Li, J.; Chen, Y.; Liu, Q.; Tian, Z.; Zhang, Y. Mechanistic and therapeutic links between rheumatoid arthritis and diabetes mellitus. Clin. Exp. Med. 2022. Epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, L.; Cregan, S.; Biniecka, M.; Cunningham, C.; Veale, D.J.; Kane, D.J.; Fearon, U.; Mullan, R.H. Insulin-resistant pathways are associated with disease activity in rheumatoid arthritis and are subject to disease modification through metabolic reprogramming: A potential novel therapeutic approach. Arthritis Rheumatol. 2020, 72, 896–902. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Tang, X.; Pang, M. Prevalence of metabolic syndrome in patients with rheumatoid arthritis: An updated systematic review and meta-analysis. Front. Med. 2022, 9, 855141. [Google Scholar] [CrossRef] [PubMed]
- Aussel, C.; Desmoulins, D.; Agneray, J.; Ekindjian, O.G. Effect of insulin on aminoisobutyric acid uptake by human non-rheumatoid and rheumatoid synovial cells. FEBS Lett. 1987, 214, 327–330. [Google Scholar] [CrossRef]
- Han, J.M.; Patterson, S.J.; Speck, M.; Ehses, J.A.; Levings, M.K. Insulin inhibits IL-10-mediated regulatory T cell function: Implications for obesity. J. Immunol. 2014, 192, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lu, Y.; Lin, S.H.; Li, N.; Han, Y.; Huang, Q.; Zhao, Y.; Xie, F.; Guo, Y.; Deng, B.; et al. Insulin signaling establishes a developmental trajectory of adipose regulatory T. cells. Nat. Immunol. 2021, 22, 1175–1185. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, P.G.; Farinon, M.; Sanchez-Lopez, E.; Miyamoto, S.; Guma, M. Fibroblast-like synoviocytes glucose metabolism as a therapeutic target in rheumatoid arthritis. Front. Immunol. 2019, 10, 1743. [Google Scholar] [CrossRef]
- Masoumi, M.; Mehrabzadeh, M.; Mahmoudzehi, S.; Mousavi, M.J.; Jamalzehi, S.; Sahebkar, A.; Karami, J. Role of glucose metabolism in aggressive phenotype of fibroblast-like synoviocytes: Latest evidence and therapeutic approaches in rheumatoid arthritis. Int. Immunopharmacol 2020, 89, 107064. [Google Scholar] [CrossRef]
- Tripolino, C.; Ciaffi, J.; Pucino, V.; Ruscitti, P.; van Leeuwen, N.; Borghi, C.; Giacomelli, R.; Meliconi, R.; Ursini, F. Insulin signaling in arthritis. Front. Immunol. 2021, 12, 672519. [Google Scholar] [CrossRef]
- Holst, J.J. The incretin system in healthy humans: The role of GIP and GLP-1. Metabolism 2019, 96, 46–55. [Google Scholar] [CrossRef]
- Deacon, C.F. Physiology and pharmacology of DPP-4 in glucose homeostasis and the treatment of Type 2 diabetes. Front. Endocrinol. 2019, 10, 80. [Google Scholar] [CrossRef]
- Shao, S.; Xu, Q.; Yu, X.; Pan, R.; Chen, Y. Dipeptidyl peptidase 4 inhibitors and their potential immune modulatory functions. Pharm. Ther. 2020, 209, 107503. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yang, L.; Wang, X.; Zhou, Z. The new insights from DPP-4 inhibitors: Their potential immune modulatory function in autoimmune diabetes. Diabetes Metab. Res. Rev. 2014, 30, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Cordero, O.J.; Varela-Calvino, R.; Lopez-Gonzalez, T.; Calvino-Sampedro, C.; Vinuela, J.E.; Mourino, C.; Hernandez-Rodriguez, I.; Rodriguez-Lopez, M.; Aspe de la Iglesia, B.; Pego-Reigosa, J.M. CD26 expression on T helper populations and sCD26 serum levels in patients with rheumatoid arthritis. PLoS ONE 2015, 1, e0131992. [Google Scholar] [CrossRef] [PubMed]
- Yeganeh, F.; Mousavi, S.M.J.; Hosseinzadeh-Sarband, S.; Ahmadzadeh, A.; Bahrami-Motlagh, H.; Hoseini, M.H.M.; Sattari, M.; Sohrabi, M.R.; Pouriran, R.; Dehghan, P. Association of CD26/dipeptidyl peptidase IV mRNA level in peripheral blood mononuclear cells with disease activity and bone erosion in rheumatoid arthritis. Clin. Rheumatol. Assoc. CD26 2018, 37, 3183–3190. [Google Scholar] [CrossRef]
- Yamauchi, K.; Sato, Y.; Yamashita, K.; Funase, Y.; Kaneko, T.; Hashimoto, T.; Aizawa, T. RS3PE in association with dipeptidyl peptidase-4 inhibitor: Report of two cases. Diabetes Care 2012, 35, e7. [Google Scholar] [CrossRef]
- Yokota, K.; Igaki, N. Sitagliptin (DPP-4 inhibitor)-induced rheumatoid arthritis in type 2 diabetes mellitus: A case report. Intern. Med. 2012, 51, 2041–2044. [Google Scholar] [CrossRef]
- Padron, S.; Rogers, E.; Demory Beckler, M.; Kesselman, M. DPP-4 inhibitor (sitagliptin)-induced seronegative rheumatoid arthritis. BMJ Case Rep. 2019, 12, e228981. [Google Scholar] [CrossRef]
- Douros, A.; Abrahami, D.; Yin, H.; Yu, O.H.Y.; Renoux, C.; Hudson, M.; Azoulay, L. Use of dipeptidyl peptidase-4 inhibitors and new-onset rheumatoid arthritis in patients with Type 2 diabetes. Epidemiology 2018, 29, 904–912. [Google Scholar] [CrossRef]
- Chen, Y.C.; Chen, T.H.; Sun, C.C.; Chen, J.Y.; Chang, S.S.; Yeung, L.; Tsai, Y.W. Dipeptidyl peptidase-4 inhibitors and the risks of autoimmune diseases in type 2 diabetes mellitus patients in Taiwan: A nationwide population-based cohort study. Acta. Diabetol. 2020, 57, 1181–1192. [Google Scholar] [CrossRef]
- Charoenngam, N.; Rittiphairoj, T.; Ponvilawan, B.; Ungprasert, P. Use of dipeptidyl peptidase-4 inhibitors is associated with a lower risk of rheumatoid arthritis in patients with type 2 diabetes mellitus: A systematic review and meta-analysis of cohort studies. Diabetes Metab. Syndr. 2021, 15, 249–255. [Google Scholar] [CrossRef]
- Suezawa, M.; Dainichi, T.; Kaku, Y.; Izumi, M.; Kataoka, K.; Ishii, N.; Koga, H.; Izumi, K.; Nishie, W. Dipeptidyl peptidase 4 inhibitor-associated mucous membrane pemphigoid. J. Dermatol. 2021, 48, 1584–1587. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Sahoo, J.; Narayanan, N.; Merugu, C.; Kamalanathan, S.; Naik, D. Dipeptidyl peptidase-4 inhibitor-induced autoimmune diseases: Current evidence. World J. Diabetes 2021, 12, 1426–1441. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Liu, X.; Wei, Y.; Li, X.; Gao, S.; Dong, L.; Rao, X.; Zhong, J. Emerging role of dipeptidyl peptidase-4 in autoimmune disease. Front. Immunol. 2022, 13, 830863. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Takeda, K. Role of gut microbiota in rheumatoid arthritis. J. Clin. Med. 2017, 6, 60. [Google Scholar] [CrossRef]
- Rosser, E.C.; Mauri, C. A clinical update on the significance of the gut microbiota in systemic autoimmunity. J. Autoimmun. 2016, 74, 85–93. [Google Scholar] [CrossRef]
- Bergot, A.S.; Giri, R.; Thomas, R. The microbiome and rheumatoid arthritis. Best Pr. Res. Clin. Rheumatol. 2019, 33, 101497. [Google Scholar] [CrossRef]
- Maeda, Y.; Kurakawa, T.; Umemoto, E.; Motooka, D.; Ito, Y.; Gotoh, K.; Hirota, K.; Matsushita, M.; Furuta, Y.; Narazaki, M.; et al. Dysbiosis contributes to arthritis development via activation of autoreactive T cells in the intestine. Arthritis Rheumatol. 2016, 68, 2646–2661. [Google Scholar] [CrossRef]
- Scher, J.U.; Sczesnak, A.; Longman, R.S.; Segata, N.; Ubeda, C.; Bielski, C.; Rostron, T.; Cerundolo, V.; Pamer, E.G.; Abramson, S.B.; et al. Expansion of intestinal Prevotella copri correlates with enhanced susceptibility to arthritis. eLife 2013, 2, e01202. [Google Scholar] [CrossRef]
- Alpizar-Rodriguez, D.; Lesker, T.R.; Gronow, A.; Gilbert, B.; Raemy, E.; Lamacchia, C.; Gabay, C.; Finckh, A.; Strowig, T. Prevotella copri in individuals at risk for rheumatoid arthritis. Ann. Rheum. Dis. 2019, 78, 590–593. [Google Scholar] [CrossRef]
- Leite, A.Z.; Rodrigues, N.C.; Gonzaga, M.I.; Paiolo, J.C.C.; de Souza, C.A.; Stefanutto, N.A.V.; Omori, W.P.; Pinheiro, D.G.; Brisotti, J.L.; Matheucci Junior, E.; et al. Detection of increased plasma interleukin-6 levels and prevalence of Prevotella copri and Bacteroides vulgatus in the feces of Type 2 diabetes patients. Front. Immunol. 2017, 8, 1107. [Google Scholar] [CrossRef]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature 2016, 535, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.D.; Nguyen, L.H.; Li, Y.; Yan, Y.; Ma, W.; Rinott, E.; Ivey, K.L.; Shai, I.; Willett, W.C.; Hu, F.B.; et al. The gut microbiome modulates the protective association between a Mediterranean diet and cardiometabolic disease risk. Nat. Med. 2021, 27, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Picchianti Diamanti, A.; Panebianco, C.; Salerno, G.; Di Rosa, R.; Salemi, S.; Sorgi, M.L.; Meneguzzi, G.; Mariani, M.B.; Rai, A.; Iacono, D.; et al. Impact of Mediterranean diet on disease activity and gut microbiota composition of rheumatoid arthritis patients. Microorganisms 2020, 8, 1989. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Ju, Z.; Zuo, T. Time for food: The impact of diet on gut microbiota and human health. Nutrition 2018, 51–52, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Liu, Q. Factors affecting gut microbiome in daily diet. Front. Nutr. 2021, 8, 644138. [Google Scholar] [CrossRef]
- Asnicar, F.; Berry, S.E.; Valdes, A.M.; Nguyen, L.H.; Piccinno, G.; Drew, D.A.; Leeming, E.; Gibson, R.; Le Roy, C.; Khatib, H.A.; et al. Microbiome connections with host metabolism and habitual diet from 1,098 deeply phenotyped individuals. Nat. Med. 2021, 27, 321–332. [Google Scholar] [CrossRef]
- Moszak, M.; Szulińska, M.; Bogdański, P. You are what you eat-the relationship between diet, microbiota, and metabolic disorders-A review. Nutrients 2020, 12, 1096. [Google Scholar] [CrossRef]
- Fajstova, A.; Galanova, N.; Coufal, S.; Malkova, J.; Kostovcik, M.; Cermakova, M.; Pelantova, H.; Kuzma, M.; Sediva, B.; Hudcovic, T.; et al. Diet rich in simple sugars promotes pro-inflammatory response via gut microbiota alteration and TLR4 signaling. Cells 2020, 9, 2701. [Google Scholar] [CrossRef]
- Correa-Rodríguez, M.; Pocovi-Gerardino, G.; Callejas-Rubio, J.L.; Ríos Fernández, R.; Martín-Amada, M.; Cruz-Caparros, M.G.; Medina-Martínez, I.; Ortego-Centeno, N.; Rueda-Medina, B. Dietary intake of free sugars is associated with disease activity and dyslipidemia in systemic lupus erythematosus patients. Nutrients 2020, 12, 1094. [Google Scholar] [CrossRef]
- Anhê, F.F.; Barra, N.G.; Schertzer, J.D. Glucose alters the symbiotic relationships between gut microbiota and host physiology. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E111–E116. [Google Scholar] [CrossRef]
- Silva, D.N.A.; Casarin, M.; Monajemzadeh, S.; Bezerra, B.B.; Lux, R.; Pirih, F.Q. The Microbiome in Periodontitis and Diabetes. Front Oral Health. 2022. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hung, S.-L.; Lee, N.G.; Chang, L.Y.; Chen, Y.T.; Lai, Y.L. Stimulatory effects of glucose and Porphyromonas gingivalis lipopolysaccharide on the secretion of inflammatory mediators from human macrophages. J. Periodontol. 2014, 85, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Moles, M.A.; Ramos-Garcia, P. State of evidence on oral health problems in diabetic patients: A critical review of the literature. J. Clin. Med. 2021, 10, 5383. [Google Scholar] [CrossRef] [PubMed]
- Bingham, C.O., III; Moni, M. Periodontal disease and rheumatoid arthritis: The evidence accumulates for complex pathobiologic interactions. Curr. Opin. Rheumatol. 2013, 25, 345–353. [Google Scholar] [CrossRef]
- Liu, J.; Gao, J.; Wu, Z.; Mi, L.; Li, N.; Wang, Y.; Peng, X.; Xu, K.; Wu, F.; Zhang, L. Anti-citrullinated Protein Antibody Generation, Pathogenesis, Clinical Application, and Prospects. Front Med 2022, 8, 802934. [Google Scholar]
- González-Febles, J.; Sanz, M. Periodontitis and rheumatoid arthritis: What have we learned about their connection and their treatment? Periodontol 2000 2021, 87, 181–203. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masuko, K. Glucose as a Potential Key to Fuel Inflammation in Rheumatoid Arthritis. Nutrients 2022, 14, 2349. https://doi.org/10.3390/nu14112349
Masuko K. Glucose as a Potential Key to Fuel Inflammation in Rheumatoid Arthritis. Nutrients. 2022; 14(11):2349. https://doi.org/10.3390/nu14112349
Chicago/Turabian StyleMasuko, Kayo. 2022. "Glucose as a Potential Key to Fuel Inflammation in Rheumatoid Arthritis" Nutrients 14, no. 11: 2349. https://doi.org/10.3390/nu14112349
APA StyleMasuko, K. (2022). Glucose as a Potential Key to Fuel Inflammation in Rheumatoid Arthritis. Nutrients, 14(11), 2349. https://doi.org/10.3390/nu14112349