
The Complex Role of Lactic Acid Bacteria in Food Detoxification

 ,
,  , , , ,
, , , ,  and
and
Abstract
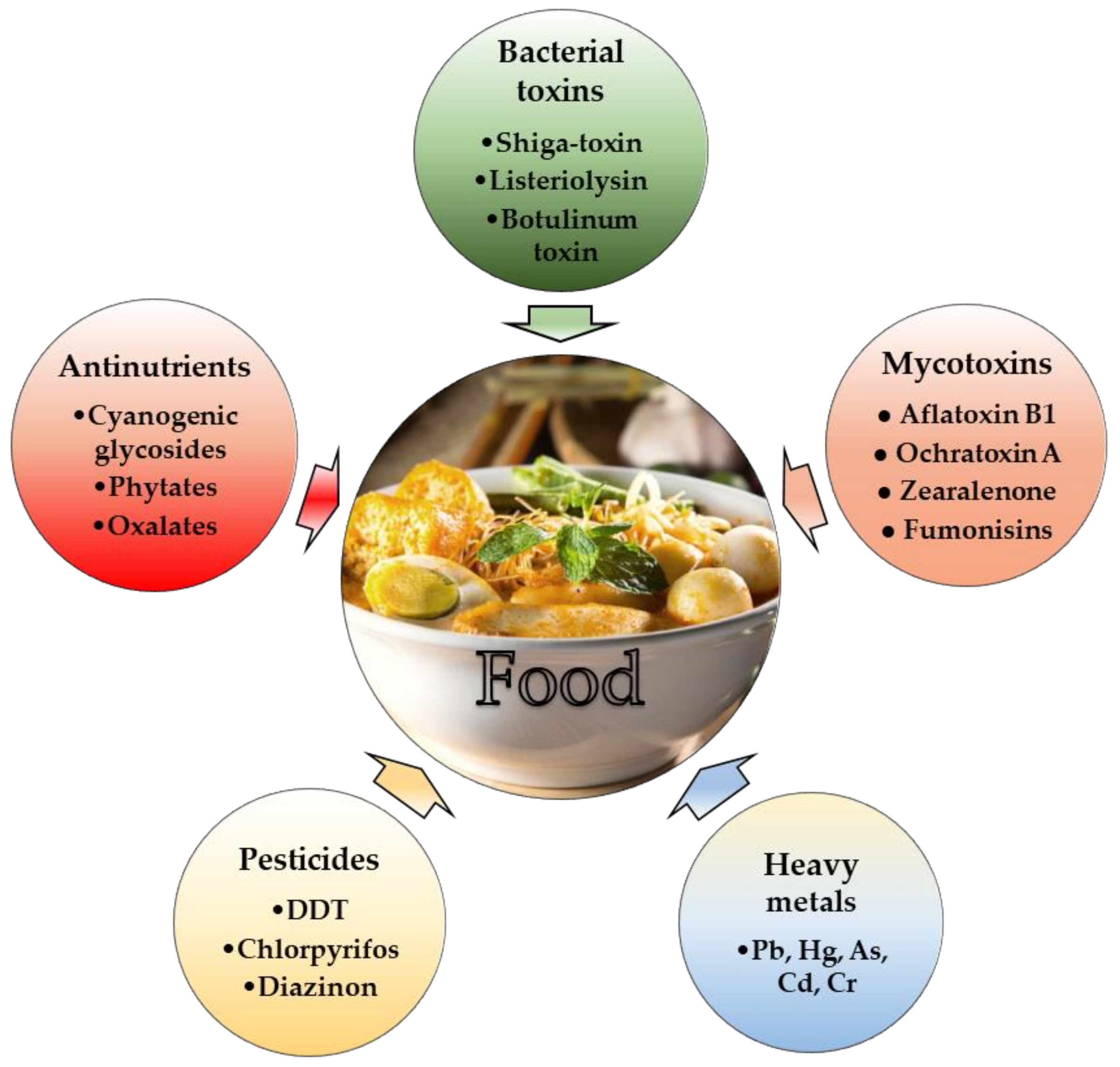
1. Introduction
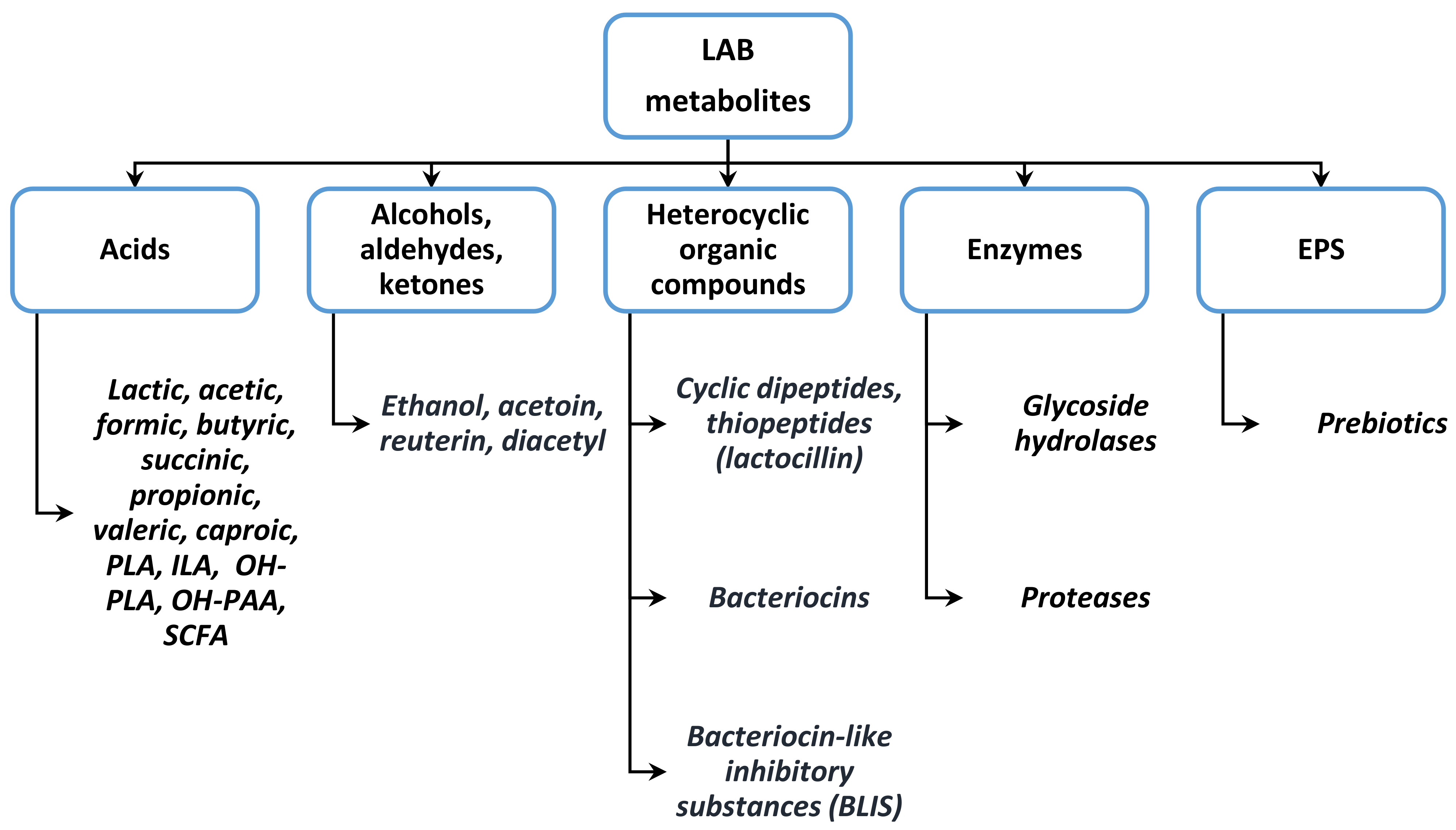
2. Lactic Acid Bacteria as Probiotics
3. LAB against Bacterial Toxins and Their Producers
3.1. LAB against Toxigenic Escherichia coli
3.2. LAB against Listeria Monocytogenes
3.3. LAB Preventing the Growth and Toxin Production by Clostridium botulinum
3.4. LAB Preventing the Growth and Toxin Production by Other Pathogenic Bacteria
4. LAB against Mycotoxins and Their Producers
4.1. Mycotoxins—Overview and Medical Relevance
4.2. LAB Detoxification of Mycotoxins
4.2.1. LAB against Aflatoxin B1 (AFB1)
4.2.2. LAB against Ochratoxin A (OTA)
4.2.3. LAB against Patulin
4.2.4. LAB against Deoxynivalenol (DON), Fumonisins, and Zearalenone (ZEA)
5. Lactic Acid Bacteria for Reducing Pesticide Levels in Food
6. LAB against Heavy Metals Intoxication

{kind=link}
{kind=link}
| Heavy Metal | Biosorbent | Initial Metal Concentration (mg/L) | Metal Removal (%) | Metal Removal Capacity (mg/g Dry Biomass) | References |
|---|---|---|---|---|---|
| Hg | Weissella viridescens MY 205 | 1 | 79.6 | [300] | |
| Cd | Propionibacterium freudenreichii shermanii JS | 50 | 49.1 | [305] | |
| L. acidophilus ATCC 20552 | 50 | 65.5 | [304] | ||
| Lc. rhamnosus, L. acidophilus, Bifidobacterium longum | 43 a,* | 48.0 | [310] | ||
| Bifidobacterium longum 46 | 10 | 54.7 | [314] | ||
| Ent. faecium EF031 | 10 | 97.5 | [315] | ||
| Lp. plantarum PTCC 1896 | 10 | 90.9 | 122.7 b,c | [299] | |
| Lp. plantarum CCFM8610 | 5 | 77.0 | 3.85 | [311] | |
| Li. reuteri Cd70-13 | 1 | 25.0 | [302] | ||
| P. acidilactici As105-7 | 1 | 0.13 d | [306] | ||
| W. viridescens MY 205 | 1 | 54.1 | [286] | ||
| Pb | Lp. plantarum LAB-32 | 200 | 82.25 | 57.31 b | [255] |
| Lp. plantarum PTCC 1896 | 50 | 65.4 | 34.5 b,c | [299] | |
| L. acidophilus ATCC 20552 | 50 | 72.6 | [304] | ||
| Propionibacterium freudenreichii shermanii JS | 50 | 69.9 | [305] | ||
| Li. reuteri Pb71-1 | 6 | 59.0 | [302] | ||
| P. acidilactici As105-7 | 6 | 0.76 d | [306] | ||
| As | L. acidophilus | 1 | 60.0 | [316] | |
| L. acidophilus ATCC 20552 | 0.5 | 49.8 | [304] | ||
| Lc. casei DSM20011 | 0.1 | 38.1 | 0.312 c | [289] | |
| P. acidilactici As102-4 | 0.1 | 0.006 d | [306] | ||
| Al | Lp. plantarum CCFM639 | 50 | 26.83 | [312] | |
| Cu | Ent. faecium | 250 | 106.38 c | [308] | |
| Lentilactobacillus buchneri DSM 20057 | 40 | 46.17 c | [317] | ||
| Lev. brevis | 20 | 26.5 c | [318] | ||
| Fe | L. bulgaricus Lb-12 | 100 | 99.3 | [307] | |
| Str. thermophilus STM-7 | 100 | 100.0 | [307] | ||
| Zn | L. bulgaricus Lb-12 | 100 | 90.2 | [307] | |
| Str. thermophilus STM-7 | 100 | 92.8 | [307] | ||
| Leuc. mesenteroides | 20 | 27.10 c | [319] | ||
| W. viridescens MY 205 | 1 | 20.0 | [300] |
7. LAB in Detoxication of Food from Natural Antinutrients
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- E. Coli: Rapid Response in a Crisis. Available online: http://www.efsa.europa.eu/en/press/news/120711 (accessed on 3 March 2022).
- Investigations of Foodborne Illness Outbreaks. US Food and Drugs Administration. Available online: https://www.fda.gov/food/outbreaks-foodborne-illness/investigations-foodborne-illness-outbreaks (accessed on 30 March 2022).
- Gossner, C.M.; Schlundt, J.; Ben Embarek, P.; Hird, S.; Lo-Fo-Wong, D.; Beltran, J.J.; Teoh, K.N.; Tritscher, A. The melamine incident: Implications for international food and feed safety. Environ. Health Perspect. 2009, 117, 1803–1808. [Google Scholar] [CrossRef] [PubMed]
- INFOSAN Quarterly Summary, 2021 #4. Available online: https://www.who.int/news/item/04-02-2022-infosan-quarterly-summary-2021-4 (accessed on 25 February 2022).
- Regueiro, J.; López-Fernández, O.; Rial-Otero, R.; Cancho-Grande, B.; Simal-Gándara, J. A Review on the Fermentation of Foods and the Residues of Pesticides—Biotransformation of Pesticides and Effects on Fermentation and Food Quality. Crit. Rev. Food Sci. Nutr. 2015, 55, 839–863. [Google Scholar] [CrossRef] [PubMed]
- Pesticide Residues in Food—2017: Toxicological Evaluations. Available online: https://www.who.int/publications/i/item/9789240006775 (accessed on 28 February 2022).
- Petrova, P.; Ivanov, I.; Tsigoriyna, L.; Valcheva, N.; Vasileva, E.; Parvanova-Mancheva, T.; Arsov, A.; Petrov, K. Traditional Bulgarian Dairy Products: Ethnic Foods with Health Benefits. Microorganisms 2021, 9, 480. [Google Scholar] [CrossRef] [PubMed]
- Petrova, P.; Petrov, K. Lactic Acid Fermentation of Cereals and Pseudocereals: Ancient Nutritional Biotechnologies with Modern Applications. Nutrients 2020, 12, 1118. [Google Scholar] [CrossRef] [PubMed]
- Petrova, P.; Petrov, K. Prebiotic–probiotic relationship: The genetic fundamentals of polysaccharides conversion by Bifidobacterium and Lactobacillus genera. In Handbook of Food Bioengineering, 1st ed.; Grumezescu, A.M., Holban, A.M., Eds.; Elsevier Inc.: San Diego, CA, USA, 2017; Volume 2, pp. 237–278. [Google Scholar]
- Ranjha, M.M.A.N.; Shafique, B.; Batool, M.; Kowalczewski, P.Ł.; Shehzad, Q.; Usman, M.; Manzoor, M.F.; Zahra, S.M.; Yaqub, S.; Aadil, R.M. Nutritional and Health Potential of Probiotics: A Review. Appl. Sci. 2021, 11, 11204. [Google Scholar] [CrossRef]
- Grumet, L.; Tromp, Y.; Stiegelbauer, V. The Development of High-Quality Multispecies Probiotic Formulations: From Bench to Market. Nutrients 2020, 12, 2453. [Google Scholar] [CrossRef] [PubMed]
- Żółkiewicz, J.; Marzec, A.; Ruszczyński, M.; Feleszko, W. Postbiotics—A Step Beyond Pre- and Probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Bertin, Y.; Habouzit, C.; Duniere, L.; Laurier, M.; Durand, A.; Duchez, D.; Segura, A.; Thevenot-Sergentet, D.; Baruzzi, F.; Chaucheyras-Durand, F. Lactobacillus reuteri suppresses E. coli O157:H7 in bovine ruminal fluid: Toward a pre-slaughter strategy to improve food safety? PLoS ONE 2017, 12, e0187229. [Google Scholar] [CrossRef]
- Trinder, M.; Bisanz, J.E.; Burton, J.P.; Reid, G. Probiotic lactobacilli: A potential prophylactic treatment for reducing pesticide absorption in humans and wildlife. Benef. Microbes 2015, 6, 841–847. [Google Scholar] [CrossRef]
- Grzywacz, J.G.; Belden, J.B.; Robertson, A.M.; Hernandez, D.C.; Carlos Chavez, F.L.; Merten, M.J. Parenting, Pesticides and Adolescent Psychological Adjustment: A Brief Report. Int. J. Environ. Res. Public Health 2022, 19, 540. [Google Scholar] [CrossRef] [PubMed]
- Goh, Y.J.; Klaenhammer, T.R. Genetic Mechanisms of Prebiotic Oligosaccharide Metabolism in Probiotic Microbes. Ann. Rev. Food Sci. Technol. 2015, 6, 137–156. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhou, T.; Tang, H.; Li, X.; Chen, Y.; Zhang, L.; Zhang, J. Probiotic potential and amylolytic properties of lactic acid bacteria isolated from Chinese fermented cereal foods. Food Control 2020, 111, 107057. [Google Scholar] [CrossRef]
- Velikova, P.; Stoyanov, A.; Blagoeva, G.; Popova, L.; Petrov, K.; Gotcheva, V.; Angelov, A.; Petrova, P. Starch utilization routes in lactic acid bacteria: New insight by gene expression assay. Starch-Stärke 2016, 68, 953–960. [Google Scholar] [CrossRef]
- Gibson, G.R. From probiotics to prebiotics and a healthy digestive system. J. Food Sci. 2004, 69, 141–143. [Google Scholar] [CrossRef]
- Fujiya, M.; Kohgo, Y. Novel perspectives in probiotic treatment: The efficacy and unveiled mechanisms of the physiological functions; Clin. J. Gastroenterol. 2010, 3, 117–127. [Google Scholar] [CrossRef][Green Version]
- Bendali, F.; Madi, N.; Sadoun, D. Beneficial effects of a strain of Lactobacillus paracasei subsp. paracasei in Staphylococcus aureus-induced intestinal and colonic injury. Int. J. Infect. Dis. 2011, 15, e787–e794. [Google Scholar] [CrossRef]
- Makino, S.; Ikegami, S.; Kano, H.; Sashihara, T.; Sugano, H.; Horiuchi, H.; Saito, T.; Oda, M. Immunomodulatory Effects of Polysaccharides Produced by Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1. J. Dairy Sci. 2006, 89, 2873–2881. [Google Scholar] [CrossRef]
- Kalliomaki, M.; Salminen, S.; Arvilommi, H.; Kero, P.; Koskinen, P.; Isolauri, E. Probiotics in primary prevention of atopic disease, a randomised placebo-controlled trial. Lancet 2001, 357, 1076–1079. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, C.; Ai, L.; Zhou, F.; Zhou, Z.; Wang, L.; Zhang, H.; Chen, W.; Guo, B. Complete genome sequence of the probiotic Lactobacillus plantarum ST-III. J. Bacteriol. 2011, 193, 313–314. [Google Scholar] [CrossRef]
- Van Hoorde, K.; Verstraete, T.; Vandamme, P.; Huys, G. Diversity of lactic acid bacteria in two Flemish artisan raw milk Gouda-type cheeses. Food Microbiol. 2008, 25, 929–935. [Google Scholar] [CrossRef] [PubMed]
- Arena, M.P.; Capozzi, V.; Russo, P.; Drider, D.; Spano, G.; Fiocco, D. Immunobiosis and probiosis: Antimicrobial activity of lactic acid bacteria with a focus on their antiviral and antifungal properties. Appl. Microbiol. Biotechnol. 2018, 102, 9949–9958. [Google Scholar] [CrossRef] [PubMed]
- Birt, D.F.; Boylston, T.; Hendrich, S.; Jane, J.L.; Hollis, J.; Li, L.; McClelland, J.; Moore, S.; Phillips, G.J.; Rowling, M.; et al. Resistant starch: Promise for improving human health. Adv. Nutr. 2013, 4, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Fazeli, M.R.; Shahverdi, A.R.; Sedaghat, B.; Jamalifar, H.; Samadi, N. Sourdough-isolated Lactobacillus fermentum as a potent anti-mould preservative of a traditional Iranian bread. Eur. Food Res. Technol. 2004, 218, 554–556. [Google Scholar] [CrossRef]
- Van der Meulen, R.; Scheirlinck, I.; Van Schoor, A.; Huys, G.; Vancanneyt, M.; Vandamme, P.; De Vuyst, L. Population dynamics and metabolite target analysis of lactic acid bacteria during laboratory fermentations of wheat and spelt sourdoughs. Appl. Environ. Microbiol. 2007, 73, 4741–4750. [Google Scholar] [CrossRef]
- Negatu, D.A.; Gengenbacher, M.; Dartois, V.; Dick, T. Indole Propionic Acid, an Unusual Antibiotic Produced by the Gut Microbiota, With Anti-inflammatory and Antioxidant Properties. Front. Microbiol. 2020, 11, 575586. [Google Scholar] [CrossRef]
- Zhao, Z.H.; Xin, F.Z.; Xue, Y.; Hu, Z.; Han, Y.; Ma, F.; Zhou, D.; Liu, X.-L.; Cui, A.; Liu, Z.; et al. Indole-3-propionic acid inhibits gut dysbiosis and endotoxin leakage to attenuate steatohepatitis in rats. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef]
- Lanciotti, R.; Patrignani, F.; Bagnolini, F.; Guerzoni, M.E.; Gardini, F. Evaluation of diacetyl antimicrobial activity against Escherichia coli, Listeria monocytogenes and Staphylococcus aureus. Food Microbiol. 2003, 20, 537–543. [Google Scholar] [CrossRef]
- Reid, G. Probiotic lactobacilli for urogenital health in women. J. Clin. Gastroenterol. 2008, 42 (Suppl. 3), 234–236. [Google Scholar] [CrossRef]
- Kumariya, R.; Garsa, A.K.; Rajput, Y.S.; Sood, S.K.; Akhtar, N.; Patel, S. Bacteriocins: Classification, synthesis, mechanism of action and resistance development in food spoilage causing bacteria. Microb. Pathog. 2019, 128, 171–177. [Google Scholar] [CrossRef]
- Demirbaş, F.; İspirli, H.; Kurnaz, A.A.; Tahsin Yilmaz, M.T.; Dertli, E. Antimicrobial and functional properties of lactic acid bacteria isolated from sourdoughs. LWT Food Sci. Technol. 2017, 79, 361–366. [Google Scholar] [CrossRef]
- Lo, S.; Thiam, I.; Fall, B.; Ba-Diallo, A.; Diallo, O.F.; Diagne, R.; Dia, M.L.; Ka, R.; Sarr, A.M.; Sow, A.I. Urinary tract infection with Corynebacterium aurimucosum after urethroplasty stricture of the urethra: A case report. J. Med. Case Rep. 2015, 9, 156. [Google Scholar] [CrossRef] [PubMed]
- Axel, C.; Zannini, E.; Arendt, E.K. Mold spoilage of bread and its biopreservation: A review of current strategies for bread shelf life extension. Crit. Rev. Food Sci. Nutr. 2017, 57, 3528–3542. [Google Scholar] [CrossRef] [PubMed]
- Strom, K.; Sjogren, J.; Broberg, A.; Schnurer, J. Lactobacillus plantarum MiLAB 393 produces the antifungal cyclic dipeptides cyclo(L-Phe-L-Pro) and cyclo(L-Phe-trans-4-OH-L-Pro) and 3-phenyllactic acid. Appl. Environ. Microbiol. 2002, 68, 4322–4327. [Google Scholar] [CrossRef] [PubMed]
- Dal Bello, F.; Clarke, C.I.; Ryan, L.A.M.; Ulmer, H.; Schober, T.J.; Strom, K.; Sjogren, J.; van Sinderen, D.; Schnurer, J.; Arendt, E.K.; et al. Improvement of the quality and shelf life of wheat bread by fermentation with the antifungal strain Lactobacillus plantarum FST 1.7. J. Cereal Sci. 2007, 45, 309–318. [Google Scholar] [CrossRef]
- Axelsson, L.T.; Chung, T.C.; Dobrogosz, W.J.; Lindgren, S.E. Production of a Broad Spectrum Antimicrobial Substance by Lactobacillus reuteri. Microb. Ecol. Health Dis. 1989, 2, 131–136. [Google Scholar] [CrossRef]
- Broberg, A.; Jacobsson, K.; Strom, K.; Schnurer, J. Metabolite Profiles of Lactic Acid Bacteria in Grass Silage. Appl. Environ. Microbiol. 2007, 73, 5547–5552. [Google Scholar] [CrossRef]
- Black, B.A.; Zannini, E.; Curtis, J.M.; Gänzle, M.G. Antifungal Hydroxy Fatty Acids Produced during Sourdough Fermentation: Microbial and Enzymatic Pathways, and Antifungal Activity in Bread. Appl. Environ. Microbiol. 2013, 79, 1866–1873. [Google Scholar] [CrossRef]
- Valerio, F.; Di Biase, M.; Lattanzio, V.M.T.; Lavermicocca, P. Improvement of the antifungal activity of lactic acid bacteria by addition to the growth medium of phenylpyruvic acid, a precursor of phenyllactic acid. Int. J. Food Microbiol. 2016, 222, 1–7. [Google Scholar] [CrossRef]
- Mieszkin, S.; Hymery, N.; Debaets, S.; Coton, E.; Le Blay, G.; Valence, F.; Mounier, J. Action mechanisms involved in the bioprotective effect of Lactobacillus harbinensis K.V9.3.1.Np against Yarrowia lipolytica in fermented milk. Int. J. Food Microbiol. 2017, 248, 47–55. [Google Scholar] [CrossRef]
- Allocati, N.; Masulli, M.; Alexeyev, M.; Di Ilio, C. Escherichia coli in Europe: An Overview. Int. J. Environ. Res. Public Health 2013, 10, 6235–6254. [Google Scholar] [CrossRef] [PubMed]
- Nakao, H.; Takeda, T. Escherichia coli Shiga toxin. J. Nat. Toxins 2000, 9, 299–313. [Google Scholar] [PubMed]
- Melton-Celsa, A.; Mohawk, K.; Teel, L.; O’Brien, A. Pathogenesis of Shiga-Toxin Producing Escherichia coli, Ricin and Shiga. Toxins 2011, 357, 67–103. [Google Scholar]
- Padhye, N.; Doyle, M. Escherichia coli O157:H7: Epidemiology, pathogenesis, and methods for detection in food. J. Food Prot. 1992, 55, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Griffin, P.; Tauxe, R. The epidemiology of infections caused by Escherichia coli O157:H7, other enterohemorrhagic E. coli, and the associated hemolytic uremic syndrome. Epidemiol. Rev. 1991, 13, 60–98. [Google Scholar] [CrossRef] [PubMed]
- Tarr, P. Escherichia coli O157:H7: Clinical, diagnostic, and epidemiological aspects of human infection. Clin. Infect. Dis. 1995, 20, 1–10. [Google Scholar] [CrossRef]
- Besser, R.; Lett, S.; Weber, J.; Doyle, M.; Barrett, T.; Wells, J.; Griffin, P. An outbreak of diarrhea and hemolytic uremic syndrome from Escherichia coli O157:H7 in fresh-pressed apple cider. JAMA 1993, 269, 2217–2220. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Update: Multistate outbreak of Escherichia coli O157:H7 infections from hamburgers—western United States 1992–1993. Morbid. Mortal. Weekly Rep. 1993, 42, 258–263.
- Steele, B.; Murphy, N.; Rance, C. An outbreak of hemolytic uremic syndrome associated with ingestion of fresh apple juice. J. Pediatr. 1982, 101, 963–965. [Google Scholar] [CrossRef]
- Leyer, G.; Wang, L.; Johnson, E. Acid Adaptation of Escherichia coli O157:H7 Increases Survival in Acidic Foods. Appl. Environ. Microbiol. 1995, 61, 3752–3755. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC167674/ (accessed on 12 March 2022). [CrossRef]
- Centers for Disease Control and Prevention. Escherichia coli O157:H7 outbreak linked to commercially distributed dry-cured salami—Washington and California. Morbid. Mortal. Wkly. Rep. 1995, 44, 157–160.
- Riley, L.; Remis, R.; Helgerson, S.; McGee, H.; Wells, J.; Davis, B.; Herbert, R.; Olcott, G.; Johnson, L.; Blake, N.; et al. Hemorrhagic colitis associated with a rare Escherichia coli serotype 0157:H7. N. Engl. J. Med. 1983, 308, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Ackers, M.; Mahon, B.; Leahy, E.; Goode, B.; Damrow, T.; Hayes, P.; Bibb, W.; Rice, D.; Barrett, T.; Hutwagner, L.; et al. An outbreak of Escherichia coli O157:H7 infections associated with leaf lettuce consumption. J. Infect. Dis. 1998, 177, 1588–1593. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Janda, J.; Blahova, K.; Minarikova, H.; Jikova, E.; Karmali, M.; Laubova, J.; Sikulova, J.; Preston, M.; Khakhria, R.; et al. Human Escherichia coli O157:H7 infection associated with the consumption of unpasteurized goat’s milk. Epidemiol. Infect. 1997, 119, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Outbreaks of Escherichia coli O157:H7 infection associated with eating alfalfa sprouts—Michigan and Virginia. Morbid. Mortal. Wkly. Rep. 1997, 46, 741–744. [Google Scholar]
- Doyle, M.; Zhao, T.; Meng, J.; Zhao, S. Escherichia coli O157:H7. In Food Microbiology, Fundamentals and Frontiers; Doyle, M.P., Beuchat, L.R., Montville, T.J., Eds.; ASM Press: Washington, DC, USA, 1997; pp. 171–191. Available online: https://www.worldcat.org/title/food-microbiology-fundamentals-and-frontiers/oclc/46642017 (accessed on 9 March 2022).
- Ryu, S.H.; Lee, J.H.; Park, S.H.; Song, M.O.; Park, S.H.; Jung, H.W.; Park, G.Y.; Choi, S.M.; Kim, M.S.; Chae, Y.Z.; et al. Antimicrobial resistance profiles among Escherichia coli strains isolated from commercial and cooked foods. Int. J. Food Microbiol. 2012, 159, 263–266. [Google Scholar] [CrossRef]
- Abdul-Raouf, U.; Ammar, M.; Beuchat, L. Isolation of Escherichia coli 0157:H7 from some Egyptian foods. Int. J. Food Microbiol. 1996, 29, 423–426. [Google Scholar] [CrossRef]
- Dontorou, C.; Papadopoulou, C.; Filioussis, G.; Economou, V.; Apostolou, I.; Zakkas, G.; Salamoura, A.; Kansouzidou, A.; Levidiotou, S. Isolation of Escherichia coli O157:H7 from foods in Greece. Int. J. Food Microbiol. 2003, 82, 273–279. [Google Scholar] [CrossRef]
- Lukásová, J.; Abraham, B.; Cupáková, S. Occurrence of Escherichia coli O157 in raw material and food in Czech Republic. J. Vet. Med. B Infect. Dis. Vet. Public Health 2004, 51, 77–81. [Google Scholar] [CrossRef]
- Mora, A.; Blanco, J.E.; Blanco, M.; Alonso, M.P.; Dhabi, G.; Echeita, A.; González, E.A.; Bernárdez, M.I.; Blanco, J. Antimicrobial resistance of Shiga toxin (verotoxin)-producing Escherichia coli O157:H7 and non-O157 strains isolated from humans, cattle, sheep and food in Spain. Res. Microbiol. 2005, 156, 793–806. [Google Scholar] [CrossRef]
- Lee, K.-S.; Jeong, Y.-J.; Lee, M.-S. Escherichia coli Shiga Toxins and Gut Microbiota Interactions. Toxins 2021, 13, 416. [Google Scholar] [CrossRef] [PubMed]
- Orihuel, A.; Terán, L.; Renaut, J.; Vignolo, G.; De Almeida, A.; Saavedra, M.; Fadda, S. Differential Proteomic Analysis of Lactic Acid Bacteria—Escherichia coli O157:H7 Interaction and Its Contribution to Bioprotection Strategies in Meat. Front. Microbiol. 2018, 9, 1083. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Shimizu, K.; Nomoto, K.; Tanaka, R.; Hamabata, T.; Yamasaki, S.; Takeda, T.; Takeda, Y. Inhibition of in vitro growth of Shiga toxin-producing Escherichia coli O157:H7 by probiotic Lactobacillus strains due to production of lactic acid. Int. J. Food Microbiol. 2001, 68, 135–140. [Google Scholar] [CrossRef]
- Ogawa, M.; Shimizu, K.; Nomoto, K.; Takahashi, M.; Watanuki, M.; Tanaka, R.; Tanaka, T.; Hamabata, T.; Yamasaki, S.; Takeda, Y. Protective Effect of Lactobacillus casei Strain Shirota on Shiga Toxin-Producing Escherichia coli O157:H7 Infection in Infant Rabbits. Infect. Immun. 2001, 69, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Byakika, S.; Mukisa, I.; Mugabi, R.; Muyanja, C. Antimicrobial Activity of Lactic Acid Bacteria Starters against Acid Tolerant, Antibiotic Resistant, and Potentially Virulent E. coli Isolated from a Fermented Sorghum-Millet Beverage. Int. J. Microbiol. 2019, 2019, 2013539. [Google Scholar] [CrossRef]
- Brashears, M.M.; Reilly, S.S.; Gilliland, S.E. Antagonistic action of cells of Lactobacillus lactis toward Escherichia coli O157:H7 on refrigerated raw chicken meat. J. Food Prot. 1998, 61, 166–170. [Google Scholar] [CrossRef]
- Walsham, A.D.S.; MacKenzie, D.A.; Cook, V.; Wemyss-Holden, S.; Hews, C.L.; Juge, N.; Schüller, S. Lactobacillus reuteri Inhibition of Enteropathogenic Escherichia coli Adherence to Human Intestinal Epithelium. Front. Microbiol. 2016, 7, 00244. [Google Scholar] [CrossRef]
- Eaton, K.A.; Honkala, A.; Auchtung, T.A.; Britton, R.A. Probiotic Lactobacillus reuteri ameliorates disease due to enterohemorrhagic Escherichia coli in germfree mice. Infect. Immun. 2011, 79, 185–191. [Google Scholar] [CrossRef]
- Peterson, R.E.; Klopfenstein, T.J.; Erickson, G.E.; Folmer, J.; Hinkley, S.; Moxley, R.A.; Smith, D.R. Effect of Lactobacillus acidophilus strain NP51 on Escherichia coli O157:H7 fecal shedding and finishing performance in beef feedlot cattle. J. Food Prot. 2007, 70, 287–291. [Google Scholar] [CrossRef]
- Aditya, A.; Li, Y.; Biswas, D. Antagonistic Effects of Conjugated Linoleic Acids of Lactobacillus casei Against Foodborne Enterohemorrhagic Escherichia coli. J. Food Prot. 2022, 85, 712–719. [Google Scholar] [CrossRef]
- Zeinhom, M.; Tellez, A.M.; Delcenserie, V.; El-Kholy, A.M.; El-Shinawy, S.H.; Griffiths, M.W. Yogurt Containing Bioactive Molecules Produced by Lactobacillus acidophilus La-5 Exerts a Protective Effect against Enterohemorrhagic Escherichia coli in Mice. J. Food Prot. 2012, 75, 1796–1805. [Google Scholar] [CrossRef] [PubMed]
- Medellin-Peña, M.; Griffiths, M. Effect of Molecules Secreted by Lactobacillus acidophilus Strain La-5 on Escherichia coli O157:H7 Colonization. Appl. Environ. Microbiol. 2009, 75, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Henry, K.C.; Hagen, K.E.; Gordonpour, M.; Tompkins, T.A.; Sherman, P.M. Surface-layer protein extracts from Lactobacillus helveticus inhibit enterohaemorrhagic Escherichia coli O157:H7 adhesion to epithelial cells. Cell Microbiol. 2007, 9, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Hirano, J.; Yoshida, T.; Sugiyama, T.; Koide, N.; Mori, I.; Yokochi, T. The Effect of Lactobacillus rhamnosus on Enterohemorrhagic Escherichia coli Infection of Human Intestinal Cells In Vitro. Microbiol. Immunol. 2003, 47, 405–409. [Google Scholar] [CrossRef]
- Cadieux, P.; Burton, J.; Devillard, E.; Reid, G. Lactobacillus By-Products Inhibit The Growth And Virulence Of Uropathogenic Escherichia coli. J. Phys. Pharmacol. 2009, 60, 13–18. Available online: https://pubmed.ncbi.nlm.nih.gov/20224146/ (accessed on 11 March 2022).
- Caridi, A. Selection of Escherichia coli-inhibiting strains of Lactobacillus paracasei subsp. paracasei. J. Ind. Microbiol. Biotechnol. 2002, 29, 303–308. [Google Scholar] [CrossRef]
- Fijan, S.; Šulc, D.; Steyer, A. Study of the In Vitro Antagonistic Activity of Various Single-Strain and Multi-Strain Probiotics against Escherichia coli. Int. J. Environ. Res. Public Health 2018, 15, 1539. [Google Scholar] [CrossRef]
- Maury, M.M.; Tsai, Y.-H.; Charlier, C.; Touchon, M.; Chenal-Francisque, V.; Leclercq, A.; Criscuolo, A.; Gaultier, C.; Roussel, S.; Brisabois, A.; et al. Uncovering Listeria monocytogenes hypervirulence by harnessing its biodiversity. Nat. Gen. 2016, 48, 308–313. [Google Scholar] [CrossRef]
- Farber, J.M.; Peterkin, P.I. Listeria monocytogenes: A food borne pathogen. Microbiol. Rev. 1991, 5, 476–511. [Google Scholar] [CrossRef]
- Kessler, S.L.; Dajani, A.S. Listeria meningitis in infants and children. Pediatr. Infect. Dis. J. 1990, 9, 6–63. [Google Scholar] [CrossRef]
- Buchanan, R.; Lindqvist, R.; Ross, T.; Smith, M.; Todd, E.; Whiting, R. Risk Assessment of Listeria Monocytogenes in Ready-to-Eat Foods. Microbiol. Risk Assess. Ser. 2004, 4. Food and Agriculture Organization of the United Nations, Rome (Italy). Available online: https://www.fao.org/3/y5394e/Y5394E.pdf (accessed on 15 February 2022).
- Vazquez-Boland, J.-A.; Kuhn, M.; Berche, P.; Chakraborty, T.; Domínguez-Bernal, G.; Goebel, W.; González-Zorn, B.; Wehland, J.; Kreft, J. Listeria pathogenesis and molecular virulence determinants. Clin. Microbiol. Rev. 2001, 14, 584–640. [Google Scholar] [CrossRef] [PubMed]
- Yap, P.C.; MatRahim, N.A.; AbuBakar, S.; Lee, H.Y. Antilisterial potential of lactic acid bacteria in eliminating Listeria monocytogenes in host and ready-to-eat food application. Microbiol. Res. 2021, 12, 234–257. [Google Scholar] [CrossRef]
- Dramsi, S.; Cossart, P. Listeriolysin O: A genuine cytolysin optimized for an intracellular parasite. J. Cell Biol. 2002, 156, 943–946. [Google Scholar] [CrossRef]
- Glass, K.A.; Doyle, M.P. Fate of Listeria monocytogenes in processed meat products during refrigerated storage. Appl. Environ. Microbiol. 1989, 55, 1565–1569. [Google Scholar] [CrossRef]
- Leasor, S.B.; Foegeding, P.M. Listeria spp. in commercially broken raw liquid whole egg. J. Food Prot. 1989, 52, 777–780. [Google Scholar] [CrossRef]
- Yin, H.B.; Chi-Hung Chen, C.H.; Boomer, A.; Byun, S.; Venkitanarayanan, K.; Macarisin, D.; Patel, J. Biocontrol of Listeria on cantaloupes with lactic acid bacteria. J. Food Process Preserv. 2020, 44, 14465. [Google Scholar] [CrossRef]
- Hoelzer, K.; Sauders, B.D.; Sanchez, M.D.; Olsen, P.T.; Pickett, M.M.; Mangione, K.J.; Rice, D.H.; Corby, J.; Stich, S.; Fortes, E.D.; et al. Prevalence, distribution, and diversity of Listeria monocytogenes in retail environments, focusing on small establishments and establishments with a history of failed inspections. J. Food Prot. 2011, 74, 1083–1095. [Google Scholar] [CrossRef]
- Gandhi, M.; Chikindas, M.L. Listeria: A foodborne pathogen that knows how to survive. Int. J. Food Microbiol. 2007, 113, 1–15. [Google Scholar] [CrossRef]
- McClure, P.J.; Roberts, T.A.; Otto Oguru, P. Comparison of the effects of sodium chloride, pH and temperature on the growth of Listeria monocytogenes on gradient plates and in liquid medium. Lett. Appl. Microbiol. 1989, 9, 95–99. [Google Scholar] [CrossRef]
- Lomonaco, S.; Decastelli, L.; Nucera, D.; Gallina, S.; Bianchi, D.M.; Civera, T. Listeria monocytogenes in Gorgonzola: Subtypes diversity and persistence over time. Int. J. Food Microbiol. 2009, 128, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Miladi, H.; Bakhrouf, A.; Ammar, E. Cellular lipid fatty acid profiles of reference and food isolates Listeria monocytogenes as a response to refrigeration and freezing stress. J. Food Biochem. 2013, 37, 136–143. [Google Scholar] [CrossRef]
- Bayles, D.; Wilkinson, B. Osmoprotectants and cryoprotectants for Listeria monocytogenes. Lett. Appl. Microbiol. 2000, 30, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Amit, S.K.; Uddin, M.M.; Rahman, R.; Islam, S.M.R.; Khan, M.S. A review on mechanisms and commercial aspects of food preservation and processing. Agric. Food Secur. 2017, 6, 51. [Google Scholar] [CrossRef]
- Scatassa, M.L.; Gaglio, R.; Cardamone, C.; Macaluso, G.; Arcuri, L.; Todaro, M.; Mancuso, I. Anti-listeria activity of lactic acid bacteria in two traditional Sicilian cheeses. Ital. J. Food Safety 2017, 6, 13–17. [Google Scholar] [CrossRef][Green Version]
- Linares-Morales, J.R.; Gutierrez-Mendez, N.; Rivera-Chavira, B.E.; Perez-Vega, S.B.; Nevarez-Moorillon, G.V. Biocontrol processes in fruits and fresh produce, the use of lactic acid bacteria as a sustainable option. Front. Sustain. Food Syst. 2018, 2, 50. [Google Scholar] [CrossRef]
- Kasra-Kermanshahi, R.; Mobarak-Qamsari, E. Inhibition effect of lactic acid bacteria against food born pathogen Listeria monocytogenes. Appl. Food Biotechnol. 2015, 2, 11–19. [Google Scholar] [CrossRef]
- Panebianco, F.; Giarratana, F.; Caridi, A.; Sidari, R.; De Bruno, A.; Giuffrida, A. Lactic acid bacteria isolated from traditional Italian dairy products: Activity against Listeria monocytogenes and modelling of microbial competition in soft cheese. LWT 2020, 137, 110446. [Google Scholar] [CrossRef]
- Mishra, C.; Lambert, J. Production of antimicrobial substances by probiotics. Asia Pac. J. Clin. Nut. 1996, 5, 20–24. Available online: https://apjcn.nhri.org.tw/server/APJCN/5/1/20.pdf (accessed on 18 February 2022).
- Leroy, F.; Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Jay, J. Antimicrobial properties of diacetyl. Appl. Environ. Mirobiol. 1982, 44, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Mulet-Powell, N.; Lacoste-Armynot, A.M.; Vicas, M.; De Buochberg, M.S. Interactions between pairs of bacteriocins from lactic acid bacteria. J. Food Prot. 1998, 61, 1210–1212. [Google Scholar] [CrossRef] [PubMed]
- Amezquita, A.; Brashears, M.M. Competitive inhibition of Listeria monocytogenes in ready-to-eat meat products by lactic acid bacteria. J. Food Prot. 2001, 65, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Saraoui, T.; Fall, P.A.; Leroi, F.; Antignac, J.P.; Chereau, S.; Pilet, M.F. Inhibition mechanism of Listeria monocytogenes by a bioprotective bacteria Lactococcus piscium CNCM I-4031. Food Microbiol. 2016, 53, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Plaza-Diaz, J.; Munoz-Quezada, S.; Gomez-Llorente, C.; Gil, A. Probiotic mechanisms of action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef]
- Wang, L.; He, Z.; Tian, P.; Wang, G. Lactic Acid Bacteria and Host Immunity. In Lactic Acid Bacteria: Omics and Functional Evaluation; Chen, W., Ed.; Springer: New York, NY, USA, 2019; pp. 261–296. [Google Scholar]
- Upadhyay, A.; Upadhyaya, I.; Mooyottu, S.; Venkitanarayanan, K. Eugenol in combination with lactic acid bacteria attenuates Listeria monocytogenes virulence in vitro and in invertebrate model Galleria mellonella. J. Med. Microbiol. 2016, 65, 443–455. [Google Scholar] [CrossRef]
- Winkelstroter, L.K.; De Martinis, E.C. Effect of bacteriocins and conditions that mimic food and digestive tract on biofilm formation, in vitro invasion of eukaryotic cells and internalin gene expression by Listeria monocytogenes. Probiotics Antimicrob. Proteins 2013, 5, 153–164. [Google Scholar] [CrossRef]
- Rios-Covian, D.; Nogacka, A.; Salazar, N.; Hernandez-Barranco, A.M.; Cuesta, I.; Gueimonde, M.; de Los Reyes Gavilan, C.G. Bifidobacterium breve IPLA20005 affects in vitro the expression of hly and luxS genes, related to the virulence of Listeria monocytogenes Lm23. Can. J. Microbiol. 2018, 64, 215–221. [Google Scholar] [CrossRef]
- Fernandez, N.; Wrzosek, L.; Radziwill-Bienkowska, J.M.; Ringot-Destrez, B.; Duviau, M.P.; Noordine, M.L.; Laroute, V.; Robert, V.; Cherbuy, C.; Daveran-Mingot, M.L.; et al. Characterization of mucus-related properties of Streptococcus thermophilus: From adhesion to induction. Front. Physiol. 2018, 9, 980. [Google Scholar] [CrossRef]
- Ren, C.; Zhang, Q.; de Haan, B.J.; Zhang, H.; Faas, M.M.; de Vos, P. Identification of TLR2/TLR6 signalling lactic acid bacteria for supporting immune regulation. Sci. Rep. 2016, 6, 34561. [Google Scholar] [CrossRef]
- Azizpour, M.; Hosseini, S.D.; Jafari, P.; Akbary, N. Lactococcus lactis: A New Strategy for Vaccination. Avicenna J. Med. Biotechnol. 2017, 9, 163–168. [Google Scholar] [PubMed]
- Sullivan, L.O.; Ross, R.P.; Hill, C. Potential of bacteriocin-producing lactic acid bacteria for improvements in food safety and quality. Biochimie 2002, 84, 593–604. [Google Scholar] [CrossRef]
- Pinilla, C.M.B.; Brandelli, A. Antimicrobial activity of nanoliposomes co-encapsulating nisin and garlic extract against Gram-positive and Gram-negative bacteria in milk. Innov. Food Sci. Emerg. Technol. 2016, 36, 287–293. [Google Scholar] [CrossRef]
- Malheiros, S.; Sant’Anna, V.; Barbosa, S.; Brandelli, A.; Franco, B.D. Effect of liposome-encapsulated nisin and bacteriocin-like substance P34 on Listeria monocytogenes growth in Minas frescal cheese. Int. J. Food Microbiol. 2012, 156, 272–277. [Google Scholar] [CrossRef]
- Malheiros, S.; Daroit, D.J.; Brandelli, A. Inhibition of Listeria monocytogenes in minas frescal cheese by free and nanovesicle-encapsulated nisin. Braz. J. Microbiol. 2012, 43, 1414–1418. [Google Scholar] [CrossRef]
- Thomas, L.; Wimpenny, J. Investigation of the effect of combined variations in temperature, pH, and NaCl concentration on nisin inhibition of Listeria monocytogenes and Staphylococcus aureus. Appl. Environ. Microbiol. 1996, 62, 2006–2012. [Google Scholar] [CrossRef]
- Martín, I.; Rodríguez, A.; Alía, A.; Martínez-Blanco, M.; Ojalvo, D.L.; Córdoba, J.J. Control of Listeria monocytogenes growth and virulence in a traditional soft cheese model system based on lactic acid bacteria and a whey protein hydrolysate with antimicrobial activity. Int. J. Food Microbiol. 2022, 361, 109444. [Google Scholar] [CrossRef]
- Morandi, S.; Silvetti, T.; Vezzini, V.; Morozzo, E.; Brasca, M. How we can improve the antimicrobial performances of lactic acid bacteria? A new strategy to control Listeria monocytogenes in Gorgonzola cheese. Food Microbiol. 2020, 90, 103488. [Google Scholar] [CrossRef]
- Koster, S.; van Pee, K.; Hudel, M.; Leustik, M.; Rhinow, D.; Kuhlbrandt, W.; Chakraborty, T.; Yildiz, O. Crystal structure of listeriolysin O reveals molecular details of oligomerization and pore formation. Nat. Commun. 2014, 5, 3690. [Google Scholar] [CrossRef]
- Hernandez-Flores, K.G.; Vivanco-Cid, H. Biological effects of listeriolysin O: Implications for vaccination. BioMed Res. Int. 2015, 2015, 360741. [Google Scholar] [CrossRef]
- Szatraj, K.; Szczepankowska, A.K.; Chmielewska-Jeznach, M. Lactic acid bacteria promising vaccine vectors: Possibilities, limitations, doubts. J. Appl. Microbiol. 2017, 123, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Mercenier, A.; Muller-Alouf, H.; Grangette, C. Lactic acid bacteria as live vaccines. Curr. Issues Mol. Biol. 2000, 2, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Wyszynska, A.; Kobierecka, P.; Bardowski, J.; Jagusztyn-Krynicka, E.K. Lactic acid bacteria-20 years exploring their potential as live vectors for mucosal vaccination. Appl. Microbiol. Biotechnol. 2015, 99, 2967–2977. [Google Scholar] [CrossRef] [PubMed]
- Marcotte, H.; Koll-Klais, P.; Hultberg, A.; Zhao, Y.; Gmur, R.; Mandar, R.; Mikelsaar, M.; Hammarstrom, L. Expression of single-chain antibody against RgpA protease of Porphyromonas gingivalis in Lactobacillus. J. Appl. Microbiol. 2006, 100, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Dhaked, R.K.; Singh, M.K.; Singh, P.; Gupta, P. Botulinum toxin: Bioweapon & magic drug. Indian J. Med. Res. 2010, 132, 489–503. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3028942/ (accessed on 26 March 2022).
- Lund, B.M.; Peck, M.W. Clostridium botulinum. In Guide to Foodborne Pathogens, 2nd ed.; Labbé, R.G., García, S., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2013; pp. 91–111. [Google Scholar]
- Nowakowska, M.B.; Selby, K.; Przykopanski, A.; Krüger, M.; Krez, N.; Dorner, B.G.; Dorner, M.B.; Jin, R.; Minton, N.P.; Rummel, A.; et al. Construction and validation of safe Clostridium botulinum Group II surrogate strain producing inactive botulinum neurotoxin type E toxoid. Sci. Rep. 2022, 12, 1790. [Google Scholar] [CrossRef]
- Alizadeh, A.M.; Hashempour-Baltork, F.; Alizadeh-Sani, M.; Maleki, M.; Azizi-Lalabad, M.; Khosravi-Darani, K. Inhibition of Clostridium botulinum and its toxins by probiotic bacteria and their metabolites. Qual. Assur. Saf. Crops Foods 2020, 12, 59–68. [Google Scholar] [CrossRef]
- Lynt, R.K.; Kautter, D.A.; Solomon, H. Differences and Similarities among Proteolytic and Nonproteolytic Strains of Clostridium botulinum Types A, B, E and F: A Review. J. Food Prot. 1982, 45, 466–474. [Google Scholar] [CrossRef]
- Eskandari, M.H.; Hosseinpour, S.; Mesbahi, G.R.; Shekarforoush, S. New composite nitrite-free and low-nitrite meat-curing systems using natural colorants. Food Sci. Nutr. 2013, 1, 392–401. [Google Scholar] [CrossRef]
- Lucumi-Banguero, R.S.; Ramírez-Toro, C.; Bolivar, G.A. Potential Use of Lactic Acid Bacteria with Pathogen Inhibitory Capacity as a Biopreservative Agent for Chorizo. Processes 2021, 9, 1582. [Google Scholar] [CrossRef]
- Okereke, A.; Montville, T.J. Bacteriocin Inhibition of Clostridium botulinum Spores by Lactic Acid Bacteria. J. Food Prot. 1991, 54, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Okereke, A.; Montville, T.J. Bacteriocin-mediated inhibition of Clostridium botulinum spores by lactic acid bacteria at refrigeration and abuse temperatures. Appl. Environ. Microbiol. 1991, 57, 3423–3428. [Google Scholar] [CrossRef] [PubMed]
- Dobson, A.; Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocin Production: A Probiotic Trait? Appl. Environ. Microbiol. 2012, 78, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, S. Preserving non-fermented refrigerated foods with microbial cultures—A review. Trends Food Sci. Technol. 2002, 12, 276–284. [Google Scholar] [CrossRef]
- Lewus, C.B.; Kaiser, A.; Montville, T.J. Inhibition of food-borne bacterial pathogens by bacteriocins from lactic acid bacteria isolated from meat. Appl. Environ. Microbiol. 1991, 57, 1683–1688. [Google Scholar] [CrossRef] [PubMed]
- Roces, C.; Rodríguez, A.; Martínez, B. Cell Wall-active Bacteriocins and Their Applications beyond Antibiotic Activity. Probiotics Antimicrob. Proteins 2012, 4, 259–272. [Google Scholar] [CrossRef]
- De Arauz, L.J.; Jozala, A.F.; Mazzola, P.G.; Penna, T.C.V. Nisin biotechnological production and application: A review. Trends Food Sci. Technol. 2009, 20, 146–154. [Google Scholar] [CrossRef]
- Khan, I.; Oh, D.H. Integration of nisin into nanoparticles for application in foods. Innov. Food Sci. Emerg. Technol. 2016, 34, 376–384. [Google Scholar] [CrossRef]
- Wiedemann, I.; Breukink, E.; Kraaij, C.v.; Kuipers, O.P.; Bierbaum, G.; Kruijff, B.D.; Sahl, H.G. Specific binding of nisin to the peptidoglycan precursor lipid II combines pore formation and inhibition of cell wall biosynthesis for potent antibiotic activity. J. Biol. Chem. 2001, 276, 1772–1779. [Google Scholar] [CrossRef]
- Hernandez-González, J.C.; Martinez-Tapia, A.; Lazcano-Hernández, G.; Garcia-Pérez, B.; Castrejon-Jiménez, N.S. Bacteriocins from Lactic Acid Bacteria. A Powerful Alternative as Antimicrobials, Probiotics, and Immunomodulators in Veterinary Medicine. Animals 2021, 11, 979. [Google Scholar] [CrossRef]
- Biscola, V.; Todorov, S.D.; Capuano, V.S.C.; Abriouel, H.; Gálvez, A.; Franco, B.D.G.M. Isolation and characterization of a nisin-like bacteriocin produced by a Lactococcus lactis strain isolated from charqui, a Brazilian fermented, salted and dried meat product. Meat. Sci. 2013, 93, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Field, D.; Daly, K.; O′Connor, P.; Cotter, P.D.; Hill, C.; Ross, R. Efficacies of Nisin A and Nisin V Semipurified Preparations Alone and in Combination with Plant Essential Oils for Controlling Listeria monocytogenes. Appl. Environ. Microbiol. 2015, 81, 2762–2769. [Google Scholar] [CrossRef] [PubMed]
- Jabés, D.; Brunati, C.; Candiani, G.; Riva, S.; Romano, G.; Donadio, S. Efficacy of the new lantibiotic NAI-107 in experimental infections induced by multidrug-resistant Gram-positive pathogens. Antimicrob. Agents Chemother. 2011, 55, 1671–1676. [Google Scholar] [CrossRef] [PubMed]
- Field, D.; O’Conner, R.; Cotter, P.D.; Ross, R.P.; Hill, C. In Vitro Activities of Nisin and Nisin Derivatives Alone and In Combination with Antibiotics against Staphylococcus Biofilms. Front. Microbiol. 2016, 7, 508. [Google Scholar] [CrossRef]
- Petit, L.; Gibert, M.; Popoff, M.R. Clostridium perfringens: Toxinotype and genotype. Trends Microbiol. 1999, 7, 104–110. [Google Scholar] [CrossRef]
- Keyburn, A.L.; Sheedy, S.A.; Ford, M.E.; Williamson, M.M.; Awad, M.M.; Rood, J.I. Alpha-toxin of Clostridium perfringens is not an essential virulence factor in necrotic enteritis in chickens. Infect. Immun. 2006, 74, 6496–6500. [Google Scholar] [CrossRef]
- Guo, S.; Liu, D.; Zhang, B.; Li, Z.; Li, Y.; Ding, B.; Guo, Y. Two Lactobacillus Species Inhibit the Growth and α-Toxin Production of Clostridium perfringens and Induced Proinflammatory Factors in Chicken Intestinal Epithelial Cells in Vitro. Front Microbiol. 2017, 8, 2081. [Google Scholar] [CrossRef]
- Banykó, J.; Vyletelová, M. Determining the source of Bacillus cereus and Bacillus licheniformis isolated from raw milk, pasteurized milk and yoghurt. Lett. Appl. Microbiol. 2009, 48, 318–323. [Google Scholar] [CrossRef]
- Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernández Escámez, P.S.; Gironés, R.; Wahlström, H. Risks for public health related to the presence of Bacillus cereus and other Bacillus spp. including Bacillus thuringiensis in foodstuffs. EFSA J. 2016, 14, 4524–4550. [Google Scholar] [CrossRef]
- Halverson, L.J.; Clayton, M.K.; Handelsman, J. Variable stability of antibiotic-resistance markers in Bacillus cereus UW85 in the soybean rhizosphere in the field. Mol. Ecol. 1993, 2, 65–78. [Google Scholar] [CrossRef]
- Jensen, G.B.; Hansen, B.M.; Eilenberg, J.; Mahillon, J. The hidden lifestyles of Bacillus cereus and relatives. Environ. Microbiol. 2003, 5, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Vilain, S.; Luo, Y.; Hildreth, M.B.; Brozel, V.S. Analysis of the life cycle of the soil saprophyte Bacillus cereus in liquid soil extract and in soil. Appl. Environ. Microbiol. 2006, 72, 4970–4977. [Google Scholar] [CrossRef] [PubMed]
- Guinebretière, M.H.; Thompson, F.L.; Sorokin, A.; Normand, P.; Dawyndt, P.; Ehling-Schulz, M.; Svensson, B.; Sanchis, V.; Nguyen-The, C.; Heyndrickx, M.; et al. Ecological diversification in the Bacillus cereus group. Environ. Microbiol. 2008, 10, 851–865. [Google Scholar] [CrossRef] [PubMed]
- Bintsis, T. Foodborne pathogens. AIMS Microbiol. 2017, 3, 529–563. [Google Scholar] [CrossRef]
- Senesi, S.; Ghelardi, E. Production, secretion and biological activity of Bacillus cereus enterotoxins. Toxins 2010, 2, 1690–1703. [Google Scholar] [CrossRef]
- Dierick, K.; Van Coillie, E.; Swiecicka, I.; Meyfroidt, G.; Devlieger, H.; Meulemans, A.; Hoedemaekers, G.; Fourie, L.; Heyndrickx, M.; Mahillon, J. Fatal family outbreak of Bacillus cereus-associated food poisoning. J. Clin. Microbiol. 2005, 43, 4277–4279. [Google Scholar] [CrossRef]
- Gemechu, T. Review on lactic acid bacteria function in milk fermentation and preservation. Afr. J. Food Sci. 2015, 9, 170–175. [Google Scholar] [CrossRef]
- Wang, C.; Chang, T.; Yang, H.; Cui, M. Antibacterial mechanism of lactic acid on physiological and morphological properties of Salmonella Enteritidis, Escherichia coli and Listeria monocytogenes. Food Control 2015, 47, 231–236. [Google Scholar] [CrossRef]
- Tirloni, E.; Ghelardi, E.; Celandroni, F.; Bernardi, C.; Stella, S. Effect of dairy product environment on the growth of Bacillus cereus. J. Dairy Sci. 2017, 100, 7026–7034. [Google Scholar] [CrossRef]
- Røssland, E.; Borge, G.I.A.; Langsrud, T.; Sørhaug, T. Inhibition of Bacillus cereus by strains of Lactobacillus and Lactococcus in milk. Int. J. Food Microbiol. 2003, 89, 205–212. [Google Scholar] [CrossRef]
- Bogovič-Matijašić, B.; Rogelj, I.; Nes, I.F.; Holo, H. Isolation and characterization of two bacteriocins of Lactobacillus acidophilus LF221. J. Microbiol. Biotechnol. 1998, 49, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, S.; Martínez-Blanco, H.; Rodríguez-Aparicio, L.B.; Ferrero, M.A. Effect of fermented broth from lactic acid bacteria on pathogenic bacteria proliferation. Int. J. Dairy Sci. 2016, 99, 2654–2665. [Google Scholar] [CrossRef] [PubMed]
- Olaniyi, O.I.; Adeniran, H.A.; Abiose, S.H. Antimicrobial characteristics of lactic acid bacteria in African yam bean-based drink. Int. Food Res. J. 2019, 26, 1733–1740. Available online: http://www.ifrj.upm.edu.my/26%20(06)%202019/09%20-%20IFRJ16216.R1-Final.pdf (accessed on 18 March 2022).
- Caldera, L.; Franzetti, L. Effect of storage temperature on the microbial composition of ready-to-use vegetables. Curr. Microbiol. 2014, 68, 133–139. [Google Scholar] [CrossRef]
- Miller, A., III; Scanlan, R.A.; Lee, J.S.; Libbey, L.M. Volatile compounds produced in sterile fish muscle (Sebastes melanops) by Pseudomonas putrefaciens, Pseudomonas fluorescens, and an Achromobacter species. Appl. Microbiol. 1973, 26, 18–21. [Google Scholar] [CrossRef]
- Mellor, G.E.; Bentley, J.A.; Dykes, G.A. Evidence for a role of biosurfactants produced by Pseudomonas fluorescens in the spoilage of fresh aerobically stored chicken meat. Food Microbiol. 2011, 28, 1101–1104. [Google Scholar] [CrossRef]
- Edwards, R.A.; Dainty, R.H.; Hibbard, C.M. Volatile compounds produced by meat pseudomonads and related reference strains during growth on beef stored in air at chill temperatures. J. Appl. Bacteriol. 1987, 62, 403–412. [Google Scholar] [CrossRef]
- Reichler, S.J.; Trmčić, A.; Martin, N.H.; Boor, K.J.; Wiedmann, M. Pseudomonas fluorescens group bacterial strains are responsible for repeat and sporadic post pasteurization contamination and reduced fluid milk shelf life. Int. J. Dairy Sci. 2018, 101, 7780–7800. [Google Scholar] [CrossRef]
- Scatamburlo, T.M.; Yamazi, A.K.; Cavicchioli, V.Q.; Pieri, F.; Nero, L.A. Spoilage potential of Pseudomonas species isolated from goat milk. Int. J. Dairy Sci. 2015, 98, 759–764. [Google Scholar] [CrossRef]
- Decimo, M.; Morandi, S.; Silvetti, T.; Brasca, M. Characterization of gram-negative psychrotrophic bacteria isolated from Italian bulk tank milk. J. Food Sci. 2014, 79, M2081–M2090. [Google Scholar] [CrossRef]
- Samet-Bali, O.; Felfoul, I.; Lajnaf, R.; Attia, H.; Ayadi, M.A.; Route de Soukra, B.P.W. Study of proteolytic and lipolytic activities of Pseudomonas spp. isolated from pasteurized milk in Tunisia. J. Agric. Sci. 2013, 5, 7. [Google Scholar] [CrossRef]
- Vercet, A.; Lopez, P.; Burgos, J. Inactivation of heat-resistant lipase and protease from Pseudomonas fluorescens by manothermosonication. Int. J. Dairy Sci. 1997, 80, 29–36. [Google Scholar] [CrossRef]
- Caputo, L.; Quintieri, L.; Bianchi, D.M.; Decastelli, L.; Monaci, L.; Visconti, A.; Baruzzi, F. Pepsin-digested bovine lactoferrin prevents Mozzarella cheese blue discoloration caused by Pseudomonas fluorescens. Food Microbiol. 2015, 46, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.V. Extracellular toxins of Pseudomonas aeruginosa. J. Infect. Dis. 1974, 130, S94–S99. [Google Scholar] [CrossRef] [PubMed]
- Okorhi, B.F. Anti-pseudomonas activity of organic acids produced by lactic acid bacteria. Issues Bio. Sci. Pharma. Res. 2014, 2, 106–114. [Google Scholar]
- Duan, X.; Chen, S.; Duan, S.; Lan, C.; Yang, Z.; Cao, Y.; Miao, J. Antibiotic activities of the natural antimicrobial substance produced by Lactobacillus paracasei FX-6 against Pseudomonas putida. LWT 2020, 123, 109096. [Google Scholar] [CrossRef]
- Felicio, B.A.; Pinto, M.S.; Oliveira, F.S.; Lempk, M.W.; Pires, A.C.S.; Lelis, C.A. Effects of nisin on Staphylococcus aureus count and physicochemical properties of Minas Frescal cheese. J. Dairy Sci. 2015, 98, 4364–4369. [Google Scholar] [CrossRef]
- Pinchuk, I.V.; Beswick, E.J.; Reyes, V.E. Staphylococcal enterotoxins. Toxins 2010, 2, 2177–2197. [Google Scholar] [CrossRef]
- Sears, P.M.; McCarthy, K.K. Management and treatment of staphylococcal mastitis. Vet. Clin. Nutr. Am. Food Anim. Pract. 2003, 19, 171–185. [Google Scholar] [CrossRef]
- Le Loir, Y.; Baron, F.; Gautier, M. Staphylococcus aureus and food poisoning. Genet. Mol. Res. GMR 2003, 2, 63–76. Available online: https://hal.archives-ouvertes.fr/hal-01123026 (accessed on 15 February 2022).
- Amin, M.; Jorfi, M.; Khosravi, A.D.; Samarbafzadeh, A.R.; Sheikh, A.F. Isolation and identification of Lactobacillus casei and Lactobacillus plantarum from Plants by PCR and detection of their antibacterial activity. Int. J. Biol. Sci. 2009, 9, 810–814. [Google Scholar] [CrossRef]
- Pal, V.; Jamuna, M.; Jeevaratnam, K. Isolation and characterization of bacteriocin producing lactic acid bacteria from a South Indian Special dosa (Appam) batter. J. Cult. Collect. 2005, 4, 53–60. Available online: http://www.bioline.org.br/pdf?cc05007 (accessed on 12 March 2022).
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Applegate, T.J. Modulation of Intestinal Functions Following Mycotoxin Ingestion: Meta-Analysis of Published Experiments in Animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [PubMed]
- Boonen, J.; Malysheva, S.V.; Taevernier, L.; Di Mavungu, J.D.; De Saeger, S.; De Spiegeleer, B. Human skin penetration of selected model mycotoxins. Toxicology 2012, 301, 21–32. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Risk assessment of aflatoxins in food. EFSA J. 2020, 18, 6040. [Google Scholar] [CrossRef]
- Awuchi, C.G.; Ondari, E.N.; Ogbonna, C.U.; Upadhyay, A.K.; Baran, K.; Okpala, C.O.R.; Korzeniowska, M.; Guiné, R.P.F. Mycotoxins Affecting Animals, Foods, Humans, and Plants: Types, Occurrence, Toxicities, Action Mechanisms, Prevention, and Detoxification Strategies—A Revisit. Foods 2021, 10, 1279. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Paterson, R.R.M.; Venâncio, A. Biodegradation of Ochratoxin A for Food and Feed Decontamination. Toxins 2010, 2, 1078–1099. [Google Scholar] [CrossRef]
- Ropejko, K.; Twarużek, M. Zearalenone and Its Metabolites—General Overview, Occurrence, and Toxicity. Toxins 2021, 13, 35. [Google Scholar] [CrossRef]
- Haschek, W.M.; Beasley, V.R. Handbook of Toxicology of Chemical Warfare Agents; Elsevier: Amsterdam, The Netherlands, 2009; pp. 353–369. [Google Scholar]
- De Walle, J.V.; Sergent, T.; Piront, N.; Toussaint, O.; Schneider, Y.J.; Larondelle, Y. Deoxynivalenol affects in vitro intestinal epithelial cell barrier integrity through inhibition of protein synthesis. Toxicol. Appl. Pharmacol. 2010, 245, 291–298. [Google Scholar] [CrossRef]
- Grenier, B.; Loureiro-Bracarense, A.P.; Lucioli, J.; Pacheco, G.D.; Cossalter, A.M.; Moll, W.D.; Schatzmayr, G.; Oswald, I.P. Individual and combined effects of subclinical doses of deoxynivalenol and fumonisins in piglets. Mol. Nutr. Food Res. 2011, 55, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Muhialdin, B.J.; Saari, N.; Meor Hussin, A.S. Review on the Biological Detoxification of Mycotoxins Using Lactic Acid Bacteria to Enhance the Sustainability of Foods Supply. Molecules 2020, 25, 2655. [Google Scholar] [CrossRef] [PubMed]
- Nasrollahzadeh, A.; Mokhtari, S.; Khomeiri, M.; Saris, P.E.J. Antifungal Preservation of Food by Lactic Acid Bacteria. Foods 2022, 11, 395. [Google Scholar] [CrossRef]
- Zoghi, A.; Massoud, R.; Todorov, S.D.; Chikindas, M.L.; Popov, I.; Smith, S.; Khosravi-Darani, K. Role of the lactobacilli in food bio-decontamination: Friends with benefits. Enzyme Microb. Technol. 2021, 150, 109861. [Google Scholar] [CrossRef]
- Zhao, S.; Hao, X.; Yang, F.; Wang, Y.; Fan, X.; Wang, Y. Antifungal Activity of Lactobacillus plantarum ZZUA493 and Its Application to Extend the Shelf Life of Chinese Steamed Buns. Foods 2022, 11, 195. [Google Scholar] [CrossRef] [PubMed]
- Gerez, C.; Torino, M.; Rollán, G.; Font de Valdez, G. Prevention of bread mould spoilage using lactic acid bacteria with antifungal properties. Food Control 2009, 20, 144–148. [Google Scholar] [CrossRef]
- Lavermicocca, P.; Valerio, F.; Visconti, A. Antifungal activity of phenyllactic acid against moulds isolated from bakery products. Appl. Environ. Microbiol. 2003, 69, 634–640. [Google Scholar] [CrossRef]
- Guimarães, A.; Venancio, A.; Abrunhosa, L. Antifungal effect of organic acids from lactic acid bacteria on Penicillium nordicum. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2018, 35, 1803–1818. [Google Scholar] [CrossRef]
- Liu, A.; Zheng, Y.; Liu, L.; Chen, S.; He, L.; Ao, X.; Yang, Y.; Liu, S. Decontamination of Aflatoxins by Lactic Acid Bacteria. Curr. Microbiol. 2020, 77, 3821–3830. [Google Scholar] [CrossRef]
- Peltonen, K.; El-Nezami, H.; Haskard, C.; Ahokas, J.; Salminen, S. Aflatoxin B1 binding by dairy strains of lactic acid bacteria and bifidobacteria. J. Dairy Sci. 2001, 84, 2152–2156. [Google Scholar] [CrossRef]
- El-Nezami, H.; Kankaanpaa, P.; Salminen, S.; Ahokas, J. Ability of dairy strains of lactic acid bacteria to bind a common food carcinogen, aflatoxin B1. Food Chem. Toxicol. 1998, 36, 321–326. [Google Scholar] [CrossRef]
- Śliżewska, K.; Smulikowska, S. Detoxification of aflatoxin B1 and change in microflora pattern by probiotic in vitro fermentation of broiler feed. J. Anim. Feed Sci. 2011, 20, 300–309. [Google Scholar] [CrossRef]
- Sezer, Ç.; Güven, A.; Oral, N.B.; Vatansever, L. Detoxification of aflatoxin B1 by bacteriocins and bacteriocinogenic lactic acid bacteria. Turk. J. Vet. Anim. Sci. 2013, 37, 594–601. [Google Scholar] [CrossRef]
- Taheur, F.B.; Fedhila, K.; Chaieb, K.; Kouidhi, B.; Bakhrouf, A.; Abrunhosa, L. Adsorption of aflatoxin B1, zearalenone and ochratoxin A by microorganisms isolated from Kefir grains. Int. J. Food Microbiol. 2017, 251, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, E.Z.; Abdelall, M.F.; El-Mahdy, O.M. Detoxification of Aflatoxin B1 by Antifungal Compounds from Lactobacillus brevis and Lactobacillus paracasei, Isolated from Dairy Products. Probiotics Antimicrob. Proteins 2018, 10, 201–209. [Google Scholar] [CrossRef]
- Møller, C.O.A.; Freire, L.; Rosim, R.E.; Margalho, L.P.; Balthazar, C.F.; Franco, L.T.; Sant′Ana, A.S.; Corassin, C.H.; Rattray, F.P.; de Oliveira, C.A.F. Effect of Lactic Acid Bacteria Strains on the Growth and Aflatoxin Production Potential of Aspergillus parasiticus, and Their Ability to Bind Aflatoxin B1, Ochratoxin A, and Zearalenone in vitro. Front. Microbiol. 2021, 12, 655386. [Google Scholar] [CrossRef]
- Skrinjar, M.; Rasic, J.L.; Stojicic, V. Lowering of ochratoxin A level in milk by yoghurt bacteria and bifidobacteria. Folia Microbiol. 1996, 41, 26–28. [Google Scholar] [CrossRef]
- Böhm, J.; Grajewski, J.; Asperger, H.; Rabus, B.; Razzazi, E. Study on biodegradation of some trichothecenes (NIV, DON, DAS, T-2) and ochratoxin A by use of probiotic microorganisms. Mycot. Res. 2000, 16, 70–74. [Google Scholar] [CrossRef]
- Piotrowska, M.; Zakowska, Z. The elimination of ochratoxin A by lactic acid bacteria strains. Pol. J. Microbiol. 2005, 54, 279–286. Available online: https://pubmed.ncbi.nlm.nih.gov/16599298/ (accessed on 23 March 2022).
- Fuchs, S.; Sontag, G.; Stidl, R.; Ehrlich, V.; Kundi, M.; Knasmüller, S. Detoxification of patulin and ochratoxin A, two abundant mycotoxins, by lactic acid bacteria. Food Chem. Toxicol. 2008, 46, 1398–1407. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Inês, A.; Rodrigues, A.I.; Guimarães, A.; Pereira, V.L.; Parpot, P.; Venâncio, A. Biodegradation of ochratoxin A by Pediococcus parvulus isolated from Douro wines. Int. J. Food Microbiol. 2014, 188, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Luz, C.; Ferrer, J.; Mañes, J.; Meca, G. Toxicity reduction of ochratoxin A by lactic acid bacteria. Food. Chem. Toxicol. 2018, 112, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yue, T.; Yuan, Y.; Wang, Z.; Ye, M.; Cai, R. A new insight into the adsorption mechanism of patulin by the heat-inactive lactic acid bacteria cells. Food Control. 2015, 50, 104–110. [Google Scholar] [CrossRef]
- Zoghi, A.; Khosravi-Darani, K.; Sohrabvandi, S.; Attar, H. Patulin removal from synbiotic apple juice using Lactobacillus plantarum ATCC 8014. J. Appl. Microbiol. 2019, 126, 1149–1160. [Google Scholar] [CrossRef]
- Bahati, P.; Zeng, X.; Uzizerimana, F.; Tsoggerel, A.; Awais, M.; Qi, G.; Cai, R.; Yue, T.; Yuan, Y. Adsorption Mechanism of Patulin from Apple Juice by Inactivated Lactic Acid Bacteria Isolated from Kefir Grains. Toxins 2021, 13, 434. [Google Scholar] [CrossRef]
- Franco, T.S.; Garcia, S.; Hirooka, E.Y.; Ono, Y.S.; Dos Santos, J.S. Lactic acid bacteria in the inhibition of Fusarium graminearum and deoxynivalenol detoxification. J. Appl. Microbiol. 2011, 111, 739–748. [Google Scholar] [CrossRef]
- Zhai, Y.; Hu, S.; Zhong, L.; Lu, Z.; Bie, X.; Zhao, H.; Zhang, C.; Lu, F. Characterization of deoxynivalenol detoxification by Lactobacillus paracasei LHZ-1 isolated from Yogurt. J. Food Prot. 2019, 82, 1292–1299. [Google Scholar] [CrossRef]
- Mokoena, M.P.; Chelule, P.K.; Gqaleni, N. Reduction of fumonisin B1 and zearalenone by lactic acid bacteria in fermented maize meal. J. Food Prot. 2005, 68, 2095–2099. [Google Scholar] [CrossRef]
- Niderkorn, V.; Boudra, H.; Morgavi, D.P. Binding of Fusarium mycotoxins by fermentative bacteria in vitro. J. Appl. Microbiol. 2006, 101, 849–856. [Google Scholar] [CrossRef]
- Čvek, D.; Markov, K.; Frece, J.; Friganović, M.; Duraković, L.; Delaš, F. Adhesion of zearalenone to the surface of lactic acid bacteria cells. Croat. J. Food Technol. Biotechnol. Nutr. 2012, 7, 49–52. Available online: https://hrcak.srce.hr/file/123168 (accessed on 12 March 2022).
- Król, A.; Pomastowski, P.; Rafińska, K.; Railean-Plugaru, V.; Walczak, J.; Buszewski, B. Microbiology neutralization of zearalenone using Lactococcus lactis and Bifidobacterium sp. Anal. Bioanal. Chem. 2018, 410, 943–952. [Google Scholar] [CrossRef]
- Piotrowska, M. Microbiological Decontamination of Mycotoxins: Opportunities and Limitations. Toxins 2021, 13, 819. [Google Scholar] [CrossRef] [PubMed]
- Monbaliu, S.; Van Poucke, C.; Detavernier, C.; Dumoulin, F.; Van De Velde, M.; Schoeters, E.; Van Dyck, S.; Averkieva, O.; Van Peteghem, C.; De Saeger, S. Occurrence of Mycotoxins in Feed as Analyzed by a Multi-Mycotoxin LC-MS/MS Method. J. Agric. Food Chem. 2010, 58, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Allam, A.; Abdeen, A.; Devkota, H.P.; Ibrahim, S.S.; Youssef, G.; Soliman, A.; Abdel-Daim, M.M.; Alzahrani, K.J.; Shoghy, K.; Ibrahim, S.F.; et al. N-Acetylcysteine Alleviated the Deltamethrin-Induced Oxidative Cascade and Apoptosis in Liver and Kidney Tissues. Int. J. Environ. Res. Public Health 2022, 19, 638. [Google Scholar] [CrossRef] [PubMed]
- D′Haenens, J.P.; McDonald, K.W.; Langley, R.L.; Higgins, S.A.; Scott, R.; Farquhar, P.N.; Meggs, W.J. Aldicarb: A Case Series of Watermelon-Borne Carbamate Toxicity. J. Agromed. 2013, 18, 174–177. [Google Scholar] [CrossRef]
- Abou-Arab, A.A.K. Effect of Ras cheese manufacturing on the stability of DDT and its metabolites. Food Chem. 1997, 64, 115–119. [Google Scholar] [CrossRef]
- Nasution, L.; Bakti, D.; Agusnar, H.; Harahap, E.M. Role of Lactobacillus sakei strain pro7 to reduce dichlorodiphenyl trichloroethane level. J. Phys. 2018, 1116, 042025. [Google Scholar] [CrossRef]
- Cho, K.M.; Math, R.K.; Islam, S.M.; Lim, W.J.; Hong, S.Y.; Kim, J.M.; Yun, M.G.; Cho, J.J.; Yun, H.D. Biodegradation of chlorpyrifos by lactic acid bacteria during kimchi fermentation. J. Agric. Food Chem. 2009, 57, 1882–1889. [Google Scholar] [CrossRef]
- Islam, S.M.A.; Math, R.K.; Cho, K.M.; Lim, W.J.; Hong, S.Y.; Kim, J.M.; Yun, M.G.; Cho, J.J.; Yun, H.D. Organophosphorus hydrolase (OpdB) of Lactobacillus brevis WCP902 from kimchi is able to degrade organophosphorus pesticides. J. Agric. Food Chem. 2010, 58, 5380–5386. [Google Scholar] [CrossRef]
- Maden, B.; Kumral, A.Y. Degradation Trends of Some Insecticides and Microbial Changes during Sauerkraut Fermentation under Laboratory Conditions. J. Agric. Food Chem. 2020, 68, 14988–14995. [Google Scholar] [CrossRef]
- Kumral, Y.A.; Kumral, N.A.; Kolcu, A.; Maden, B.; Artik, B. Simulation Study for the Degradation of Some Insecticides during Different Black Table Olive Processes. ACS Omega 2020, 5, 14164–14172. [Google Scholar] [CrossRef] [PubMed]
- Dorđević, T.M.; Siler-Marinkovic, S.S.; Durovic, R.D.; Dimitrijevic-Brankovic, S.I.; Gajic Umiljendic, J.S. Stability of the pyrethroid pesticide bifenthrin in milled wheat during thermal processing, yeast and lactic acid fermentation, and storage. J. Sci. Food Agric. 2013, 93, 3377–3383. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Bai, J.; Huang, W.; Li, F.; Ke, W.; Zhang, Y.; Xie, D.; Zhang, B.; Guo, X. Characterization of a novel beta-cypermethrin-degrading strain of Lactobacillus pentosus 3-27 and its effects on bioremediation and the bacterial community of contaminated alfalfa silage. J. Hazard. Mater. 2022, 423, 127101. [Google Scholar] [CrossRef]
- Dorđević, T.M.; Siler-Marinković, S.S.; Durović-Pejčev, R.D.; Dimitrijević-Branković, S.I.; Gajić Umiljendić, J.S. Dissipation of pirimiphos-methyl during wheat fermentation by Lactobacillus plantarum. Lett. Appl. Microbiol. 2013, 57, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.W.; Liu, H.F.; Zhao, X.H. The potencies of three microorganisms to dissipate four organophosphorus pesticides in three food materials during traditional fermentation. J. Food Sci. Technol. 2015, 52, 7353–7360. [Google Scholar] [CrossRef]
- Yuan, S.; Li, C.; Yu, H.; Xie, Y.; Guo, Y.; Yao, W. Screening of lactic acid bacteria for degrading organophosphorus pesticides and their potential protective effects against pesticide toxicity. LWT 2021, 147, 111672. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Xu, D.; Zhao, X.H.; Song, Y.; Liu, Y.L.; Li, H.N. Biodegradation of two organophosphorus pesticides in whole corn silage as affected by the cultured Lactobacillus plantarum. 3 Biotech 2016, 6, 73. [Google Scholar] [CrossRef]
- Rezaei, F.; Nejati, R.; Sayadi, M.; Nematollahi, A. Diazinon reduction in apple juice using probiotic bacteria during fermentation and storage under refrigeration. Environ. Sci. Pollut. Res. 2021, 28, 61213–61224. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Xu, D.; Liu, J.Q.; Zhao, X.H. Enhanced degradation of five organophosphorus pesticides in skimmed milk by lactic acid bacteria and its potential relationship with phosphatase production. Food Chem. 2014, 164, 173–178. [Google Scholar] [CrossRef]
- Mohammadi, M.; Shadnoush, M.; Sohrabvandi, S.; Yousefi, M.; Khorshidian, N.; Mortazavian, A.M. Probiotics as Potential Detoxification Tools for Mitigation of Pesticides: A Mini Review. Int. J. Food Sci. Technol. 2020, 56, 2078–2087. [Google Scholar] [CrossRef]
- Jänsch, A.; Korakli, M.; Vogel, R.F.; Gänzle, M.G. Glutathione reductase from Lactobacillus sanfranciscensis DSM 20451T: Contribution to oxygen tolerance and thiol exchange reactions in wheat sourdoughs. Appl. Environ. Microbiol. 2007, 73, 4469–4476. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.N.; Cai, C.J.; Zeng, X.F.; Zhang, F.R.; Zhang, G.L.; Thacker, P.A.; Wang, J.J.; Qiao, S.Y. Dietary supplementation with Lactobacillus fermentum I5007 improves the anti-oxidative activity of weanling piglets challenged with diquat. J. Appl. Microbiol. 2013, 114, 1582–1591. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhai, Q. Applications of Lactic Acid Bacteria in Heavy Metal Pollution Environment. In Lactic Acid Bacteria in Foodborne Hazards Reduction, 1st ed.; Chen, W., Narbad, A., Eds.; Springer: Singapore, 2018; pp. 213–248. [Google Scholar] [CrossRef]
- Balali-Mood, M.; Naseri, K.; Tahergorabi, Z.; Khazdair, M.R.; Sadeghi, M. Toxic Mechanisms of Five Heavy Metals: Mercury, Lead, Chromium, Cadmium, and Arsenic. Front. Pharmacol. 2021, 12, 643972. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Kumari, V.; Ram, C.; Thakur, K.; Tomar, S. Bio-prospectus of cadmium bioadsorption by lactic acid bacteria to mitigate health and environmental impacts. Appl. Microbiol. Biotechnol. 2018, 102, 1599–1615. [Google Scholar] [CrossRef]
- Lin, D.; Cao, H.; Zhong, Y.; Huang, Y.; Zou, J.; He, Q.; Ji, R.; Qin, T.; Chen, Y.; Wang, D.; et al. Screening and identification of Lactic acid bacteria from Ya’an pickle water to effectively remove Pb2. AMB Express 2019, 9, 10. [Google Scholar] [CrossRef]
- Mrvčić, J.; Stanzer, D.; Šolić, E.; Stehlik-Tomas, V. Interaction of lactic acid bacteria with metal ions: Opportunities for improving food safety and quality. World J. Microbiol. Biotechnol. 2012, 28, 2771–2782. [Google Scholar] [CrossRef]
- Cheng, W.W.L.; Gobas, F.A.P.C. Assessment of Human Health Risks of Consumption of Cadmium Contaminated Cultured Oysters. Hum. Ecol. Risk Assess. 2007, 13, 370–382. [Google Scholar] [CrossRef]
- Witkowska, D.; Słowik, J.; Chilicka, K. Heavy Metals and Human Health: Possible Exposure Pathways and the Competition for Protein Binding Sites. Molecules 2021, 26, 6060. [Google Scholar] [CrossRef]
- Solon, O.; Riddell, T.J.; Quimbo, S.A.; Butrick, E.; Aylward, G.P.; Lou Bacate, M.; Peabody, J.W. Associations between cognitive function, blood lead concentration, and nutrition among children in the central Philippines. J. Pediatr. 2008, 152, 237–243. [Google Scholar] [CrossRef]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2017, 7, 60–72. [Google Scholar] [CrossRef]
- Orisakwe, O.E. Lead and cadmium in public health in Nigeria: Physicians neglect and pitfall in patient management. N. Am. J. Med. Sci. 2014, 6, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Koyu, A.; Gokcimen, A.; Ozguner, F.; Bayram, D.S.; Kocak, A. Evaluation of the effects of cadmium on rat liver. Mol. Cell Biochem. 2006, 284, 81–85. [Google Scholar] [CrossRef]
- Nordberg, G.F.; Fowler, B.A.; Nordberg, M.; Friberg, L.T. Introduction—General Considerations and International Perspectives. In Handbook on the Toxicology of Metals, 4th ed.; Nordberg, G.F., Fowler, B.A., Nordberg, M., Friberg, L.T., Eds.; Academic Press: Burlington, MA, USA, 2014; pp. 1–9. [Google Scholar] [CrossRef]
- Cuevas-González, P.F.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Aguilar-Toalá, J.E.; Hall, F.G.; Urbizo-Reyes, U.C.; Liceaga, A.M.; Hernandez-Mendoza, A.; García, H.S. Protective role of lactic acid bacteria and yeasts as dietary carcinogen-binding agents—A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 160–180. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, G.F. Historical perspectives on cadmium toxicology. Toxicol. Appl. Pharmacol. 2009, 238, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Jihen, E.H.; Fatima, H.; Nouha, A.; Baati, T.; Imed, M.; Abdelhamid, K. Cadmium retention increase: A probable key mechanism of the protective effect of zinc on cadmium-induced toxicity in the kidney. Toxicol. Lett. 2010, 196, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Ahamed, M.; Siddiqui, M.K. Environmental lead toxicity and nutritional factors. Clin. Nutr. 2007, 26, 400–408. [Google Scholar] [CrossRef]
- Farmand, F.; Ehdaie, A.; Roberts, C.K.; Sindhu, R.K. Lead-induced dysregulation of superoxide dismutases, catalase, glutathione peroxidase, and guanylate cyclase. Environ. Res. 2005, 98, 33–39. [Google Scholar] [CrossRef]
- Valko, M.; Morris, H.; Cronin, M.T. Metals, toxicity and oxidative stress. Curr. Med. Chem. 2005, 12, 1161–1208. [Google Scholar] [CrossRef] [PubMed]
- Gomes-Caminero, A.; Howe, P.; Hughes, M.; Kenyon, E.; Lewis, D.R.; Moore, M.; Ng, J.C.; Aitio, A.; Becking, G. Environmental Health Criteria 224 Arsenic and Arsenic Compounds, 2nd ed.; World Health Organization: Geneva, Switzerland, 2001; Available online: http://apps.who.int/iris/bitstream/handle/10665/42366/WHO_EHC_224.pdf;jsessionid=BEED054DABAF62A47901D273F2A2531A?sequence=1 (accessed on 14 March 2022).
- Patel, A.; Sv, A.; Shah, N.; Verma, D.K. Lactic acid bacteria as metal quenchers to improve food safety and quality. AgroLife Sci. J. 2017, 6, 146–154. Available online: https://www.cabdirect.org/cabdirect/abstract/20183009341 (accessed on 14 March 2022).
- Babel, S.; Kurniawan, T.A. Low-cost adsorbents for heavy metals uptake from contaminated water: A review. J. Hazard. Mater. 2003, 97, 219–243. [Google Scholar] [CrossRef]
- Rahman, N.; Haseen, U.; Rashid, M. Synthesis and characterization of polyacrylamide zirconium (IV) iodate ion-exchanger: Its application for selective removal of lead (II) from wastewater. Arab. J. Chem. 2007, 10, S1765–S1773. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, Y.; Zhang, P.; Yang, L.; Xu, H.; Xi, G. Adsorption characteristics of a novel ceramsite for heavy metal removal from stormwater runoff. Chin. J. Chem. Eng. 2018, 26, 96–103. [Google Scholar] [CrossRef]
- Nezamzadeh-Ejhieh, A.; Khorsandi, S. Photocatalytic degradation of 4-nitrophenol with ZnO supported nano-clinoptilolite zeolite. J. Ind. Eng. Chem. 2014, 20, 937–946. [Google Scholar] [CrossRef]
- Kobya, M.; Demirbas, E.; Senturk, E.; Ince, M. Adsorption of heavy metal ions from aqueous solutions by activated carbon prepared from apricot stone. Bioresour. Technol. 2005, 96, 1518–1521. [Google Scholar] [CrossRef]
- Wang, J.; Chen, C. Biosorbents for heavy metals removal and their future. Biotechnol. Adv. 2009, 27, 195–226. [Google Scholar] [CrossRef]
- Bisanz, J.E.; Enos, M.K.; Mwanga, J.R.; Changalucha, J.; Burton, J.P.; Gloor, G.B.; Reid, G. Randomized open-label pilot study of the influence of probiotics and the gut microbiome on toxic metal levels in Tanzanian pregnant women and school children. mBio 2014, 5, e01580-14. [Google Scholar] [CrossRef]
- Yan, H.; Carter, C.E.; Xu, C.; Singh, P.K.; Jones, M.M.; Johnson, J.E.; Dietrich, M.S. Cadmium-induced apoptosis in the urogenital organs of the male rat and its suppression by chelation. J. Toxicol. Environ. Health 1997, 52, 149–168. [Google Scholar] [CrossRef]
- Kojima, S.; Sugimura, Y.; Hirukawa, H.; Kiyozumi, M.; Shimada, H.; Funakoshi, T. Effects of Dithiocarbamates on Testicular Toxicity in Rats Caused by Acute Exposure to Cadmium. Toxicol. Appl. Pharmacol. 1992, 116, 24–29. [Google Scholar] [CrossRef]
- Jelenković, A.; Jovanović, M.D.; Stevanović, I.; Petronijević, N.; Bokonjić, D.; Zivković, J.; Igić, R. Influence of the green tea leaf extract on neurotoxicity of aluminium chloride in rats. Phytother Res. 2014, 28, 82–87. [Google Scholar] [CrossRef]
- Kratochvil, D.; Volesky, B. Advances in the biosorption of heavy metals. Trends Biotechnol. 1998, 16, 291–300. [Google Scholar] [CrossRef]
- Beveridge, T.J.; Fyfe, W.S. Metal fixation by bacterial cell walls. Can. J. Earth Sci. 1985, 22, 1893–1898. [Google Scholar] [CrossRef]
- Iskandar, I.; Koike, K.; Sendjaja, P. Identifying groundwater arsenic contamination mechanisms in relation to arsenic concentrations in water and host rocks. Environ. Earth Sci. 2012, 65, 2015–2026. [Google Scholar] [CrossRef]
- Tural, B.; Ertaş, E.; Enez, B.; Fincan, S.A.; Tural, S. Preparation and characterization of a novel magnetic biosorbent functionalized with biomass of Bacillus Subtilis: Kinetic and isotherm studies of biosorption processes in the removal of Methylene Blue. J. Environ. Chem. Eng. 2017, 5, 4795–4802. [Google Scholar] [CrossRef]
- Kinoshita, H.; Sohma, Y.; Ohtake, F.; Ishida, M.; Kawai, Y.; Kitazawa, H.; Saito, T.; Kimura, K. Biosorption of heavy metals by lactic acid bacteria and identification of mercury binding protein. Res. Microbiol. 2013, 164, 701–709. [Google Scholar] [CrossRef]
- Issazadeh, K.; Jahanpour, N.; Pourghorbanali, F.; Raeisi, G.; Faekhondeh, J. Heavy metals resistance by bacterial strains. Ann. Biol. Res. 2013, 4, 60–63. [Google Scholar]
- Monachese, M.; Burton, J.P.; Reid, G. Bioremediation and tolerance of humans to heavy metals through microbial processes: A potential role for probiotics? Appl. Environ. Microbiol. 2012, 78, 6397–6404. [Google Scholar] [CrossRef]
- Halttunen, T.; Finell, M.; Salminen, S. Arsenic removal by native and chemically modified lactic acid bacteria. Int. J. Food Microbiol. 2007, 120, 173–178. [Google Scholar] [CrossRef]
- Landersjö, C.; Yang, Z.; Huttunen, E.; Widmalm, G. Structural studies of the exopolysaccharide produced by Lactobacillus rhamnosus strain GG (ATCC 53103). Biomacromolecules 2002, 3, 880–884. [Google Scholar] [CrossRef]
- Hao, Z.; Reiske, H.R.; Wilson, D.B. Characterization of cadmium uptake in Lactobacillus plantarum and isolation of cadmium and manganese uptake mutants. Appl. Environ. Microbiol. 1999, 65, 4741–4745. [Google Scholar] [CrossRef]
- Teemu, H.; Seppo, S.; Jussi, M.; Raija, T.; Kalle, L. Reversible surface binding of cadmium and lead by lactic acid and bifidobacteria. Int. J. Food Microbiol. 2008, 125, 170–175. [Google Scholar] [CrossRef]
- Gerbino, E.; Mobili, P.; Tymczyszyn, E.; Fausto, R.; Gómez-Zavaglia, A. FTIR spectroscopy structural analysis of the interaction between Lactobacillus kefir S-layers and metal ions. J. Mol. Struct. 2011, 987, 186–192. [Google Scholar] [CrossRef]
- Boonyodying, K.; Watcharasupat, T.; Yotpanya, W.; Kitti, T.; Kawang, W.; Kunthalert, D.; Sitthisak, S. Factors Affecting the Binding of a Recombinant Heavy Metal-Binding Domain (CXXC motif) Protein to Heavy Metals. Environ. Asia 2012, 5, 70–75. [Google Scholar] [CrossRef]
- Sitthisak, S.; Knutsson, L.; Webb, J.W.; Jayaswal, R.K. Molecular characterization of the copper transport system in Staphylococcus aureus. Microbiology 2007, 153, 4274–4283. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Zhai, Q.; Zhao, J.; Liu, X.; Wang, G.; Zhang, H.; Zhang, H.; Chen, W. Lactobacillus plantarum CCFM8661 alleviates lead toxicity in mice. Biol. Trace Elem. Res. 2012, 150, 264–271. [Google Scholar] [CrossRef]
- Zhai, Q.; Wang, G.; Zhao, J.; Liu, X.; Tian, F.; Zhang, H.; Chen, W. Protective effects of Lactobacillus plantarum CCFM8610 against acute cadmium toxicity in mice. Appl. Environ. Microbiol. 2013, 79, 1508–1515. [Google Scholar] [CrossRef]
- Tian, F.; Xiao, Y.; Li, X.; Zhai, Q.; Wang, G.; Zhang, Q.; Zhang, H.; Chen, W. Protective Effects of Lactobacillus plantarum CCFM8246 against Copper Toxicity in Mice. PLoS ONE 2015, 10, e0143318. [Google Scholar] [CrossRef]
- Pakdel, M.; Soleimanian-Zad, S.; Akbari-Alavijeh, S. Screening of lactic acid bacteria to detect potent biosorbents of lead and cadmium. Food Control 2019, 100, 144–150. [Google Scholar] [CrossRef]
- Kinoshita, H.; Ohtake, F.; Ariga, Y.; Kimura, K. Comparison and characterization of biosorption by Weissella viridescens MYU 205 of periodic group 12 metal ions. Anim. Sci. J. 2016, 87, 271–276. [Google Scholar] [CrossRef]
- Abou-Shanab, R.A.; van Berkum, P.; Angle, J.S. Heavy metal resistance and genotypic analysis of metal resistance genes in gram-positive and gram-negative bacteria present in Ni-rich serpentine soil and in the rhizosphere of Alyssum murale. Chemosphere 2007, 68, 360–367. [Google Scholar] [CrossRef]
- Bhakta, J.N.; Ohnishi, K.; Munekage, Y.; Iwasaki, K.; Wei, M.Q. Characterization of lactic acid bacteria-based probiotics as potential heavy metal sorbents. J. Appl. Microbiol. 2012, 112, 1193–1206. [Google Scholar] [CrossRef]
- Akinbowale, O.L.; Peng, H.; Grant, P.; Barton, M.D. Antibiotic and heavy metal resistance in motile aeromonads and pseudomonads from rainbow trout (Oncorhynchus mykiss) farms in Australia. Int. J. Antimicrob. Agents 2007, 30, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Elsanhoty, R.M.; Al-Turki, I.A.; Ramadan, M.F. Application of lactic acid bacteria in removing heavy metals and aflatoxin B1 from contaminated water. Water Sci. Technol. 2016, 74, 625–638. [Google Scholar] [CrossRef] [PubMed]
- Halttunen, T.; Collado, M.C.; El-Nezami, H.; Meriluoto, J.; Salminen, S. Combining strains of lactic acid bacteria may reduce their toxin and heavy metal removal efficiency from aqueous solution. Lett. Appl. Microbiol. 2008, 46, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Bhakta, J.N.; Ohnishi, K.; Munekage, Y.; Iwasaki, K. Isolation and Probiotic Characterization of Arsenic-Resistant Lactic Acid Bacteria for Uptaking Arsenic. Int. J. Bioeng. Life Sci. 2010, 11, 831–838. [Google Scholar] [CrossRef]
- Sofu, A.; Sayilgan, E.; Guney, G. Experimental Design for Removal of Fe(II) and Zn(II) Ions by Different Lactic Acid Bacteria Biomasses. Int. J. Environ. Res. 2015, 9, 93–100. [Google Scholar] [CrossRef]
- Yilmaz, M.; Tay, T.; Kivanc, M.; Turk, H. Removal of copper ions from aqueous solution by a lactic acid bacterium. Braz. J. Chem. Eng. 2010, 27, 309–314. Available online: https://www.scielo.br/j/bjce/a/tt74Wx9drX6S3pV4×99Nhyb/?lang=en&format=pdf (accessed on 22 March 2022). [CrossRef]
- Zhai, Q.; Yue, X.; Fengwei, T.; Gang, W.; Jianxi, Z.; Xiaoming, L.; Yong, Q.; Hao, Z.; Wei, C. Protective effects of lactic acid bacteria-fermented soymilk against chronic cadmium toxicity in mice. RSC Adv. 2015, 5, 4648–4658. [Google Scholar] [CrossRef]
- Jama, A.M.; Mitić-Ćulafić, D.; Kolarević, S.; Đurašević, S.F.; Knežević-Vukčević, J. Protective effect of probiotic bacteria against cadmium-induced genotoxicity in rat hepatocytes in vivo and in vitro. Arch. Biol. Sci. 2012, 64, 1197–1206. [Google Scholar] [CrossRef]
- Zhai, Q.; Wang, G.; Zhao, J.; Liu, X.; Narbad, A.; Chen, Y.Q.; Zhang, H.; Tian, F.; Chen, W. Protective effects of Lactobacillus plantarum CCFM8610 against chronic cadmium toxicity in mice indicate routes of protection besides intestinal sequestration. Appl. Environ. Microbiol. 2014, 80, 4063–4071. [Google Scholar] [CrossRef]
- Zhai, Q.; Tian, F.; Zhao, J.; Zhang, H.; Narbad, A.; Chen, W. Oral Administration of Probiotics Inhibits Absorption of the Heavy Metal Cadmium by Protecting the Intestinal Barrier. Appl. Environ. Microbiol. 2016, 82, 4429–4440. [Google Scholar] [CrossRef]
- Halttunen, T.; Salminen, S.; Tahvonen, R. Rapid removal of lead and cadmium from water by specific lactic acid bacteria. Int. J. Food Microbiol. 2007, 114, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Topcu, A.; Bulat, T. Removal of cadmium and lead from aqueous solution by Enterococcus faecium strains. J. Food Sci. 2010, 75, T13–T17. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.L.; Sarma, P.N. Removal of Arsenic (III) from Waste Water Using Lactobacillus acidophilus. Bioremediat. J. 2010, 14, 92–97. [Google Scholar] [CrossRef]
- Yu, L.; Zhai, Q.; Liu, X.; Wang, G.; Zhang, Q.; Zhao, J.; Narbad, A.; Zhang, H.; Tian, F.; Chen, W. Lactobacillus plantarum CCFM639 alleviates aluminium toxicity. Appl. Microbiol. Biotechnol. 2016, 100, 1891–1900. [Google Scholar] [CrossRef] [PubMed]
- Schut, S.; Zauner, S.; Hampel, G.; König, H.; Claus, H. Biosorption of cooper by wine-relevant lactobacilli. Int. J. Food Microbiol. 2011, 145, 126–131. [Google Scholar] [CrossRef]
- Mrvcic, J.; Stanzer, D.; Bacun-Druzina, V.; Stehlik-Tomas, V. Copper binding by lactic acid bacteria (LAB). Biosci. Microflora 2009, 28, 1–6. [Google Scholar] [CrossRef][Green Version]
- Mrvčić, J.; Prebeg, T.; Barišić, L.; Stanzer, D.; Bačun-Družina, V.; Stehlik-Tomas, V. Zinc Binding by Lactic Acid Bacteria. Food Technol. Biotechnol. 2009, 47, 381–388. [Google Scholar]
- Zhai, Q.; Tian, F.; Wang, G.; Zhao, J.; Liu, X.; Cross, K.; Zhang, H.; Narbad, A.; Chen, W. The cadmium binding characteristics of a lactic acid bacterium in aqueous solutions and its application for removal of cadmium from fruit and vegetable juices. RSC Adv. 2016, 6, 5990–5998. [Google Scholar] [CrossRef]
- Petroski, W.; Minich, D.M. Is There Such a Thing as "Anti-Nutrients"? A Narrative Review of Perceived Problematic Plant Compounds. Nutrients 2020, 12, 2929. [Google Scholar] [CrossRef]
- Frontela, C.; García-Alonso, F.J.; Ros, G.; Martinez, C. Phytic acid and inositol phosphates in raw flours and infant cereals: The effect of processing. J. Food Comp. Anal. 2008, 21, 343–350. [Google Scholar] [CrossRef]
- Songré-Ouattara, L.T.; Mouquet-Rivier, C.; Icard-Vernière, C.; Humblot, C.; Diawara, B.; Guyot, J.P. Enzyme activities of lactic acid bacteria from a pearl millet fermented gruel (ben-saalga) of functional interest in nutrition. Int. J. Food Microbiol. 2008, 128, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Angural, S.; Rana, M.; Puri, N.; Kondepudi, K.K.; Gupta, N. Phytase producing lactic acid bacteria: Cell factories for enhancing micronutrient bioavailability of phytate rich foods. Trends Food Sci. Technol. 2020, 96, 1–12. [Google Scholar] [CrossRef]
- Castro-Alba, V.; Lazarte, C.E.; Perez-Rea, D.; Carlsson, N.; Almgren, A.; Bergenståhl, B.; Granfeldt, Y. Fermentation of pseudocereals quinoa, canihua, and amaranth to improve mineral accessibility through degradation of phytate. J. Sci. Food Agric. 2019, 99, 5239–5248. [Google Scholar] [CrossRef] [PubMed]
- Soto-Blanco, B. Herbal glycosides in healthcare. In Herbal Biomolecules in Healthcare Applications, 1st ed.; Mandal, S.C., Nayak, A.K., Dhara, A.K., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 239–282. [Google Scholar] [CrossRef]
- Menon, R.; Munjal, N.; Sturino, J.M. Characterization of amygdalin-degrading Lactobacillus species. J. Appl. Microbiol. 2015, 118, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Lei, V.; Amoa-Awua, W.K.; Brimer, L. Degradation of cyanogenic glycosides by Lactobacillus plantarum strains from spontaneous cassava fermentation and other microorganisms. Int. J. Food Microbiol. 1999, 53, 169–184. [Google Scholar] [CrossRef]
- Nivetha, N.; Suvarna, V.C.; Abhishek, R.U. Reduction of Phenolics, Tannins and Cyanogenic Glycosides Contents in Fermented Beverage of Linseed (Linum usitatissimum). Int. J. Food. Ferment. Technol. 2018, 8, 185–190. Available online: https://pdfs.semanticscholar.org/9a42/b58de8516f63e3292b071074e804679cbbbf.pdf (accessed on 27 March 2022). [CrossRef]
- Alphonce, S.; Kaale, L.D. Assessment of Biochemical Changes during Fermentation Process for Production of Traditional Fermented Cassava Meal “Mchuchume”. Tanz. J. Sci. 2020, 46, 228–240. Available online: https://www.ajol.info/index.php/tjs/article/view/196280 (accessed on 27 March 2022).
- Chongtham, N.; Bisht, M.S.; Premlata, T.; Bajwa, H.K.; Sharma, V.; Santosh, O. Quality improvement of bamboo shoots by removal of antinutrients using different processing techniques: A review. J. Food Sci. Technol. 2022, 59, 1–11. [Google Scholar] [CrossRef]
- Nionelli, L.; Montemurro, M.; Pontonio, E.; Verni, M.; Gobbetti, M.; Rizzello, C.G. Pro-technological and functional characterization of lactic acid bacteria to be used as starters for hemp (Cannabis sativa L.) sourdough fermentation and wheat bread fortification. Int. J. Food Microbiol. 2018, 279, 14–25. [Google Scholar] [CrossRef]
- Murru, N.; Blaiotta, G.; Peruzy, M.F.; Santonicola, S.; Mercogliano, R.; Aponte, M. Screening of Oxalate Degrading Lactic Acid Bacteria of Food Origin. Ital. J. Food Saf. 2017, 6, 6345. [Google Scholar] [CrossRef]
- Campieri, C.; Campieri, M.; Bertuzzi, V.; Swennen, E.; Matteuzzi, D.; Stefoni, S.; Pirovano, F.; Centi, C.; Ulisse, S.; Famularo, G.; et al. Reduction of oxaluria after an oral course of lactic acid bacteria at high concentration. Kidney Int. 2001, 60, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Azcarate-Peril, M.A.; Bruno-Barcena, J.M.; Hassan, H.M.; Klaenhammer, T.R. Transcriptional and functional analysis of oxalyl-coenzyme A (CoA) decarboxylase and formyl-CoA transferase genes from Lactobacillus acidophilus. Appl. Environ. Microbiol. 2006, 72, 1891–1899. [Google Scholar] [CrossRef] [PubMed]
- Gomathi, S.; Sasikumar, P.; Anbazhagan, K.; Sasikumar, S.; Kavitha, M.; Selvi, M.S.; Selvam, G.S. Screening of indigenous oxalate degrading lactic acid bacteria from human faeces and South Indian fermented foods: Assessment of probiotic potential. Sci. World J. 2014, 2014, 648059. [Google Scholar] [CrossRef] [PubMed]
- Hokama, S.; Honma, Y.; Toma, C.; Ogawa, Y. Oxalate-degrading Enterococcus faecalis. Microbiol. Immunol. 2000, 44, 235–240. [Google Scholar] [CrossRef]
- Wigner, P.; Bijak, M.; Saluk-Bijak, J. Probiotics in the Prevention of the Calcium Oxalate Urolithiasis. Cells 2022, 11, 284. [Google Scholar] [CrossRef]
- Schuppan, D.; Pickert, G.; Ashfaq-Khan, M.; Zevallos, V. Non-celiac wheat sensitivity: Differential diagnosis, triggers and implications. Best Pract. Res. Clin. Gastroenterol. 2015, 29, 469–476. [Google Scholar] [CrossRef]
- Caminero, A.; McCarville, J.L.; Zevallos, V.F.; Pigrau, M.; Yu, X.B.; Jury, J.; Galipeau, H.J.; Clarizio, A.V.; Casqueiro, J.; Murray, J.A.; et al. Lactobacilli degrade wheat amylase trypsin inhibitors to reduce intestinal dysfunction induced by immunogenic wheat proteins. Gastroenterology 2019, 156, 2266–2280. [Google Scholar] [CrossRef]
- Huang, X.; Schuppan, D.; Rojas Tovar, L.E.; Zevallos, V.F.; Loponen, J.; Gänzle, M. Sourdough fermentation degrades wheat alpha-amylase/trypsin inhibitor (ATI) and reduces pro-inflammatory Activity. Foods 2020, 9, 943. [Google Scholar] [CrossRef]


| E. coli Strain | LAB Species, Strain | Source/Model System | Agent/Bioactive Molecule | Mode of Action | References |
|---|---|---|---|---|---|
| O157:H7 | Lc. casei strain Shirota, L. acidophilus YIT 0070 | Yakult, Japan | Low pH, undissociated lactic acid | Growth inhibitory and bactericidal activities | [69] |
| O157:H7 | L. lactis | Raw chicken meat | H2O2 | Growth inhibition | [72] |
| O127:H6 | Li. reuteri ATCC PTA 6475, ATCC 53608 | Human, pig | Adhesins MUB, CmbA, MapA | Mucus layer binding and E. coli adherence decrease | [73] |
| O157:H7 | Li. reuteri ATCC PTA 6475 | Germfree mice | Reuterin | Decreased E. coli colonization, amended necrosis of the kidneys | [74] |
| O157:H7 | L. acidophilus NP51 | Cattle | Reuterin | Effective reduction of E. coli in cattle feces | [75] |
| EDL933 | Lc. casei LC wt, LC CLA | Batch fermentation | Conjugated linoleic acid | Downregulation of EHEC virulence genes | [76] |
| Mechanism | LAB Species/Strain | Source | Agent/Action | References |
|---|---|---|---|---|
| Organic acids production | Lactococcus lactis LM0230, Lp. plantarum, La. sakei | Calabrian cheeses | Intracellular pH acidification for unfavorable microenvironment for non-acidophiles | [103,104] |
| CO2 | Heterofermentative LAB | Foods | Anaerobic environment support; inhibition of enzyme decarboxylation; cell membrane disruption | [103] |
| H2O2 | Heterofermentative LAB | Foods | Inactivation of essential biomolecules by superoxide anion chain reaction; activation of the lactoperoxidase system | [105,106] |
| Diacetyl | Lactobacillus sp., Leuconostoc sp., P. aidilactici CC 8081, Streptococcus sp. | Foods | Affects the arginine-binding proteins | [105,106,107] |
| Bacteriocins production | Lactococcus lactis subsp. lactis, P. acidilactici, Ent. faecium, La. sakei, Li. reuteri INIA P572, Leu. gelidum UAL 187, Lc. rhamnosus CJNU 0519 | Drinks, Foods, Meats, Salads, Antimicrobial packaging | Bacteriocin synthesis: nisin, pediocin PA-1, enterocin A, sakacin A, reuterin, leucocin, rhamnocin 519 | [89,105,108] |
| Nutrients competition | Carnobacterium piscicola, Lactococcus piscium | Ready-to-eat meat products | Quick uptake of nutrients by LAB; bacteriocin synthesis | [89,109,110] |
| Niche competition | Li. reuteri, Li. fermentum, Lc. rhamnosus GC mutant | Foods, probiotics | Prevent the attachment on host cells through colonization and saturation of Lis. monocytogenes attachment receptor | [111,112] |
| Reduction of L. monocytogenes virulence | Li. reuteri, Li. fermentum, Lp. plantarum, Lactococcus lactis, Leu. mesenteroides, La. sakei | Human intestinal epithelial cells (Caco-2) | Competition for adhesion receptors expressed on host cells through downregulation of virulence gens (prfA, plcA, plcB, hly, actA, inlA, inlB, iap, luxS) | [113,114,115] |
| Protection of Gastrointestinal Tract from L. monocytogenes Invasion | Lc. casei, Li. reuteri, Lc. rhamnosus, Str. thermophilus | Human | MUC2 and TFF3 overexpression; mucus layer integrity conservation; serum cholesterol decrease | [89,116] |
| Host immune response modulation | L. bulgaricus, L. acidophilus, Lc. casei, L. salivarius, Lp. plantarum, Li. reuteri, Lc. rhamnosus, Lev. brevis, Str. thermophilus | Human | Reduction of the pro-inflammatory cytokines (IL-8) and anti-inflammatory cytokines (IL-10) increase | [89,117] |
| Vaccine vector | Lactococcus lactis | Human | Delivery and expression of listerial antigens | [118] |
| Strain | Metabolite | Food | References |
|---|---|---|---|
| P. pentosaceus 43200 | Bacteriocin | Meat | [139,140,141] |
| Lactococcus lactis 11454 | Nisin A | Beef | [142] |
| P. acidilactici LASC | Pediocin | Cured meat | [142] |
| P. acidilactici PO2 | Pediocin | Meat | [142] |
| Lp. plantarum BN | Bacteriocin | Meat | [143] |
| Streptococcus spp. | Nisin | Cheese | [142] |
| Inhibited Pathogen | LAB Species, Strain | Source | Agent | Mode of Action | References |
|---|---|---|---|---|---|
| C. perfringens | L. acidophilus CGMCC No. 1.1878, Li. fermentum CGMCC No. 1.2029 | Chicken | Lactic acid | Bacteriostatic effect on pathogen’s growth, repression of α-toxin synthesis, α-toxin degradation by lactobacilli, L. acidophilus inhibits C. perfringens adherence to GIT epithelium | [155] |
| B. cereus | Lactococcus lactis, Lactobacillus spp. | Skim milk, fresh cheese | Organic acids, H2O2, nisin | Bactericidal effect on pathogen’s growth by leakage of cytoplasmic content of the pathogen | [166,167,168] |
| B. cereus | L. acidophilus LF221 | Infant feces | Acidocin LF221 A and B | Bactericidal effect on pathogen’s growth | [169] |
| B. cereus | Lactococcus lactis C660, Lc. paracasei ATCC 27092 | Raw milk, human | Organic acids, H2O2, nisin | Reduced adhesion of the pathogen, prevention of biofilm formation | [170] |
| Pseudomonas spp. | Lp. plantarum, Li. fermentum, L. acidophilus, Str. thermophilus, Lactococcus lactis | Milk | Lactic, acetic, citric acids | Reduced growth | [183] |
| Ps. putida | Lc. paracasei FX-6, Lc. rhamnosus | Milk | Organic acids | Antibacterial activity, prevention of biofilm formation | [184] |
| S. aureus | Lactococcus lactis | Cheese | Lantibiotics | Reduced growth by cells disruption | [185] |
| Type * | Genus | Foods | Clinical Picture | Molecular Mechanisms | References |
|---|---|---|---|---|---|
| Aflatoxin B1 (AFB1) | Aspergillus | Nuts, peanuts, maize | Extremely potent carcinogen, strongly linked with liver cancer; immunosuppression; stunted growth | Mutagenic and genotoxic effects: binds N7 of guanine; GC to TA transversions; (–) transcription, (+) oxidative stress | [191,194,195] |
| Ochratoxin A (OTA) | Aspergillus | Cereals, coffee, figs, raisins, pork kidneys | Nephrotoxic effects in all species tested; liver damage, immune suppression, and teratogenic effects in animals | (–) Phe metabolism; (–) mitochondrial ATP production; (–) tumor-suppressor gene dmrt-1 in mice; (+) lipid peroxidation | [195,196] |
| Zearalenone (ZEA) | Fusarium | Maize, corn, other cereals | Reduced fertility, stillbirths in females; testicular atrophy and reduced spermatogenesis in males; hemato- and hepatoxic effects | ZEA-estrogen receptor complex is translocated into the nucleus which regulates the transcription of many genes | [195,197] |
| Fumonisins | Fusarium | Maize, rice, beans, beer, soybeans | Suppression of the immune response; pulmonary edema, esophageal cancer | (–) Sphingolipid synthesis; (–) mitochondrial ETC; (+) ROS generation; (+) cytotoxicity | [191,195] |
| Trichothecenes | Fusarium, Cephalosporium, Myrothecium, Stachybotrys, Trichothecium | Grains: rice, barley, oats, maize, eggs, milk, meat | Alimentary toxic aleukia (ATA): fever, diarrhea, nausea, vomiting, agranulocytosis, necrotic angina, bleeding; reduced serum levels of WBC and Ig in mice | (–) Translation; (–) mitochondrial ETC; (+) lipid peroxidation and membrane remodeling; (+) apoptosis | [191,196,198,199] |
| Patulin | Penicillium | Apples, pears, other fruits | Neurotoxic and immunotoxic effects reported in animals | As yet unknown | [195] |
| Citrinin | Penicillium, Aspergillus, Monascus | Cereals, Italian sausages | Nephrotoxic effects in all species tested; reproductive toxicity and chromosome aberrations in mice | (–) DNA and RNA synthesis; (–) microtubules assembly; (–) HSP90 multichaperone complex; (+) ROS generation | [191] |
| Ergot alkaloids | Claviceps | Various grasses and grains | Ergotism, convulsions, ataxia, gangrene, abortion | As yet unknown | [191,195] |
| Target Toxin | LAB Strain | Mechanism of Action | Maximum effectiveness | References |
|---|---|---|---|---|
| Aflatoxin B1 | ||||
| L. amylovorus CSCC 5197 and CSCC 5160, Lc. rhamnosus Lc1/3 | Probable adsorption on the cell surface | >50% AFB1 bound from solution, but reversibly | [209] | |
| Lc. rhamnosus LBGG and LC-705 | None proposed | 80% removal from liquid media, very rapidly | [210] | |
| Lc. paracasei LOCK 0920, Lev. brevis LOCK 9044, Lp. plantarum LOCK 0945 | None proposed | 39–55% decrease, depending on the initial concentration of AFB1 | [211] | |
| Lactococcus lactis, Lp. plantarum | Low-molecular proteins involved, possibly bacteriocins | 81% combined, 27–46% separately | [212] | |
| L. kefiri KFLM3 | Toxin-binding on the cell surface | 80% decrease in milk, 0% in MRS | [213] | |
| Lev. brevis NM101-1, Lc. paracasei ABRIINW.F58 | Antifungal compounds caused 52–80% transcriptional inhibition of the omt-A gene, a key player in the biosynthesis of AFB1 | 90–96% reduction of the AFB1 production by A. flavus and A. parasiticus | [214] | |
| Levilactobacillus spp. 2QB383, Lp. plantarum 1QB147, 1QB314 and 3QB350 | Toxin binding is assumed for the reduced amounts; no mechanism proposed for the reduced production | >50% reduced amount by inactivated strains in PPB *; >50% reduced production in YES broth at 25 °C | [215] | |
| Ochratoxin A | ||||
| Str. thermophilus T4, L. delbrueckii subsp. bulgaricus LB-51 | None proposed | Complete elimination of 0.5 mg/L in milk; 36 and 26% drop with 1.0 and 1.5 mg/L | [216] | |
| L. bulgaricus 259/2 and 171/2 | None proposed | Up to 94% detoxification, but very much strain-dependent | [217] | |
| Lc. rhamnosus GG, L. acidophilus CH-5, L. helveticus 8, Lactococcus lactis 202 | Toxin binding on the cell surface is assumed, another mechanism hypothesized | 60–87% decrease, rapid process but partially reversible | [218] | |
| L. acidophilus VM 20 | Toxin-binding on the cell surface | 96–97% decrease for 4 h | [219] | |
| P. parvulus UTAD 473 | Degradation by putative peptidase | 100% degradation in MRS for 7 days at 30 °C | [220] | |
| Lb. kefiri KFLM3 | Toxin-binding on the cell surface | 81% decrease in milk, 15% in MRS | [213] | |
| Lc. rhamnosus CECT 749, Lp. plantarum CECT 749 and CECT 288, Lc. casei CECT 4045, Lc. casei CECT 4040, L. bulgaricus CECT 4005 | >90% degradation by proteolytic activity; very little adsorption | 97–99% in MRS at pH 6.5 | [221] | |
| Lp. plantarum 3QB361 | Toxin-binding on cell surface assumed | ~60% reduced amount by inactivated strain in PPB | [215] | |
| Patulin | ||||
| Lev. brevis 20023 | Adsorption on the cell wall | 65% adsorption | [222] | |
| Lp. plantarum ATCC 8014 | Adsorption on the cell wall, proteins mediated | 96% decrease in apple juice during 6 weeks of cold storage | [223] | |
| L. kefiranofaciens JKSP109 | Adsorption on the cell wall | 93% removal at pH 4.6 and 15° Brix | [224] | |
| Deoxynivalenol | ||||
| Lp. plantarum GT III | Adsorption assumed; metabolic degradation suggested | 67% reduction by unviable cells (sterilized) | [225] | |
| Lc. paracasei LHZ-1 | Cell wall adsorption confirmed as the major mechanism | 40.7% reduction by the cell wall fraction, only 10.5 & 8.9% by SN or cellular lysate | [226] | |
| Fumonisins | ||||
| Lactococcus lactis, L. delbrueckii | Toxin-binding on the cell surface | 75% recovery from spiked maize meal after 4 days | [227] | |
| Lp. paraplantarum CNRZ 1885, Str. thermophilus RAR1 | Toxin binding was assumed; the role of peptidoglycan confirmed | 19–37% bound FB1, 65–76% FB2, both after TCA treatment | [228] | |
| Zearalenone | ||||
| Lactococcus lactis, L. delbrueckii | Toxin binding assumed | 68% recovery from spiked maize meal after 4 days | [227] | |
| Lp. plantarum A1 | Toxin-binding on the cell surface | 99% immediately, 77% after 72 h | [229] | |
| Lb. kefiri KFLM3 | Toxin-binding on the cell surface | 100% decrease in milk, 60% in MRS | [213] | |
| Lactococcus lactis | Surface adsorption assumed, interactions with surface proteins and intracellular uptake | 90% bound in the first 20 min | [230] | |
| Lp. plantarum 3QB361 | Toxin-binding on the cell surface | 70–80% amount reduction by inactivated strain in PPB | [215] |
| Pesticide | LAB Species/Strain | Sample/Food | Mode of Action | References |
|---|---|---|---|---|
| Organochlorine | ||||
| DDT | Lactobacillus spp. | Cereals | Phosphotriesterase | [13] |
| DDT | Streptococcus, Lactobacillus | Ras cheese | Biodegradation | [235] |
| DDT | La. sakei | Soil | Biodegradation | [236] |
| Organophosphorus | ||||
| Chlorpyrifos, coumaphos, diazinon, parathion, methyl parathion | Leuc. mesenteroides WCP907, Lev. brevis WCP902, Lp. plantarum WCP931, La. sakei WCP904 | Kimchi | Biodegradation | [237] |
| Chlorpyrifos, coumaphos, diazinon, parathion, methyl parathion | Lev. brevis WCP902 | Kimchi | Organophosphorus hydrolase OpdB | [238] |
| λ-Cyhalothrin, malathion, chlorpyrifos-methyl | Lp. plantarum 112 | Sauerkraut | Low pH | [239] |
| Deltamethrin, dimethoate, imidacloprid | Lp. plantarum 112,Lp. plantarum 123 | Black olives | Biodegradation | [240] |
| Pirimiphos-methyl | Lp. plantarum | Wheat | Organophosphorus hydrolase, low pH | [243] |
| Chlorpyrifos, dichlorvos, phorate, trichlorphon | Lp. plantarum | Wheat dough, Chinese cabbage, Tofu | Biodegradation | [244] |
| Dimethoate, parathion methyl, trichlorfon | Lp. plantarum subsp. plantarum CICC 20261 | Batch process | Phosphatase and Antioxydation | [245] |
| Phorate | Lp. plantarum | Corn silage | Enzyme hydrolysis | [246] |
| Diazinon | L. acidophilus | Apple juice | Enzyme hydrolysis | [247] |
| Diazinon, chlorpyrifos, fenitrothion, malathion | Lev. brevis 1.0209 | Milk | Enzyme hydrolysis | [248] |
| Pyrethroids | ||||
| Bifenthrin | Lp. plantarum | Wheat flour | Enzyme hydrolysis | [241] |
| Beta-cypermethrin | Lp. pentosus 3-27 | Alfalfa Silage | Enzyme hydrolysis | [242] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrova, P.; Arsov, A.; Tsvetanova, F.; Parvanova-Mancheva, T.; Vasileva, E.; Tsigoriyna, L.; Petrov, K. The Complex Role of Lactic Acid Bacteria in Food Detoxification. Nutrients 2022, 14, 2038. https://doi.org/10.3390/nu14102038
Petrova P, Arsov A, Tsvetanova F, Parvanova-Mancheva T, Vasileva E, Tsigoriyna L, Petrov K. The Complex Role of Lactic Acid Bacteria in Food Detoxification. Nutrients. 2022; 14(10):2038. https://doi.org/10.3390/nu14102038
Chicago/Turabian StylePetrova, Penka, Alexander Arsov, Flora Tsvetanova, Tsvetomila Parvanova-Mancheva, Evgenia Vasileva, Lidia Tsigoriyna, and Kaloyan Petrov. 2022. "The Complex Role of Lactic Acid Bacteria in Food Detoxification" Nutrients 14, no. 10: 2038. https://doi.org/10.3390/nu14102038
APA StylePetrova, P., Arsov, A., Tsvetanova, F., Parvanova-Mancheva, T., Vasileva, E., Tsigoriyna, L., & Petrov, K. (2022). The Complex Role of Lactic Acid Bacteria in Food Detoxification. Nutrients, 14(10), 2038. https://doi.org/10.3390/nu14102038