
Exercise Performance Upregulatory Effect of R-α-Lipoic Acid with γ-Cyclodextrin

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
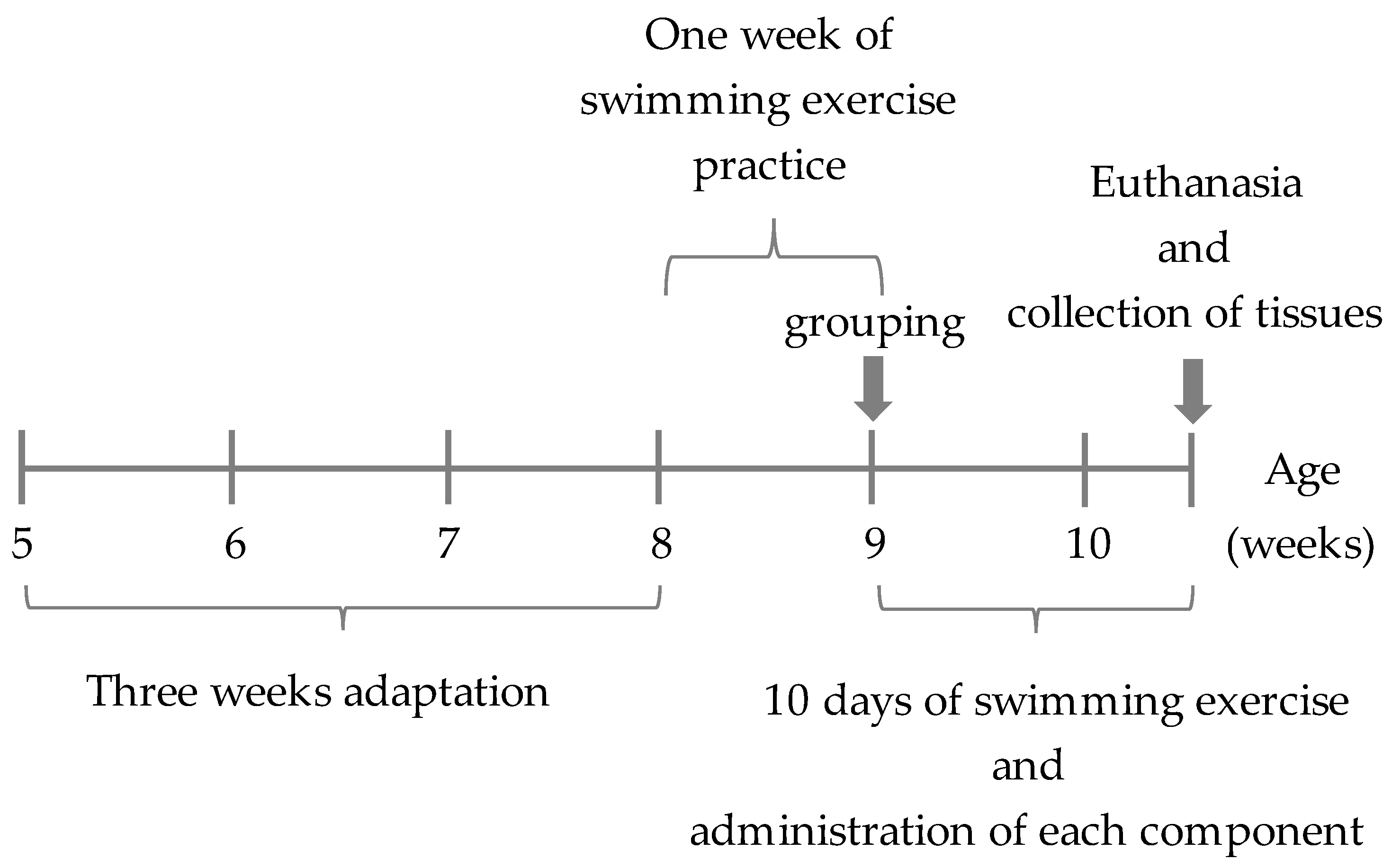
2.1. Animals and Experimental Design
2.2. Blood Biochemical Analyses
2.3. Measurement of Artificial Superoxide Anion Production
2.4. Trace Element Analysis
2.5. Exercise Time to Exhaustion in Loading Swimming Test
2.6. Lactic Acid Measurement
2.7. Histopathological and Immunohistochemical Examination of the Kidney
2.8. Statistical Analysis
3. Results
3.1. General Characteristics
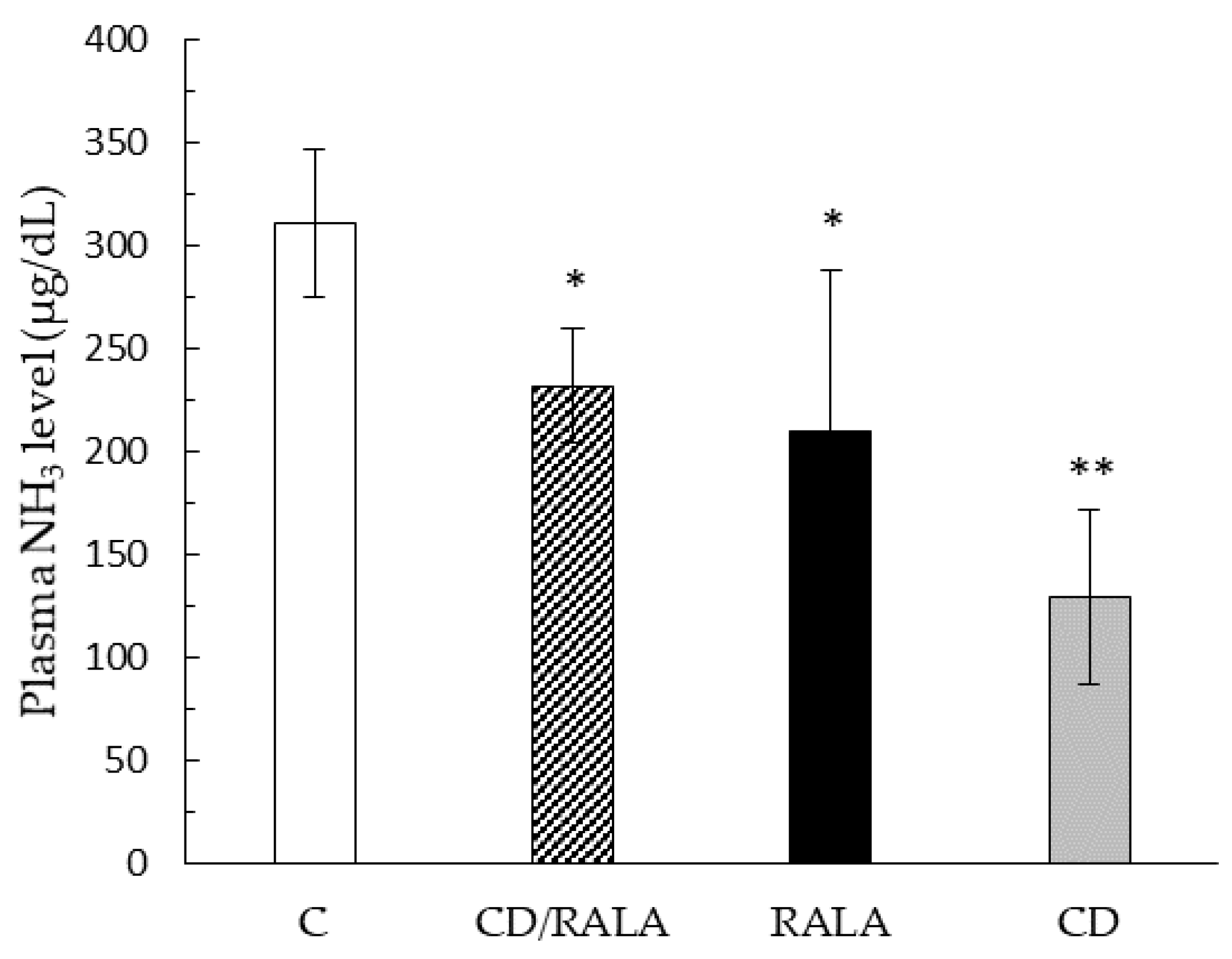
3.2. Blood Biochemical Analyses
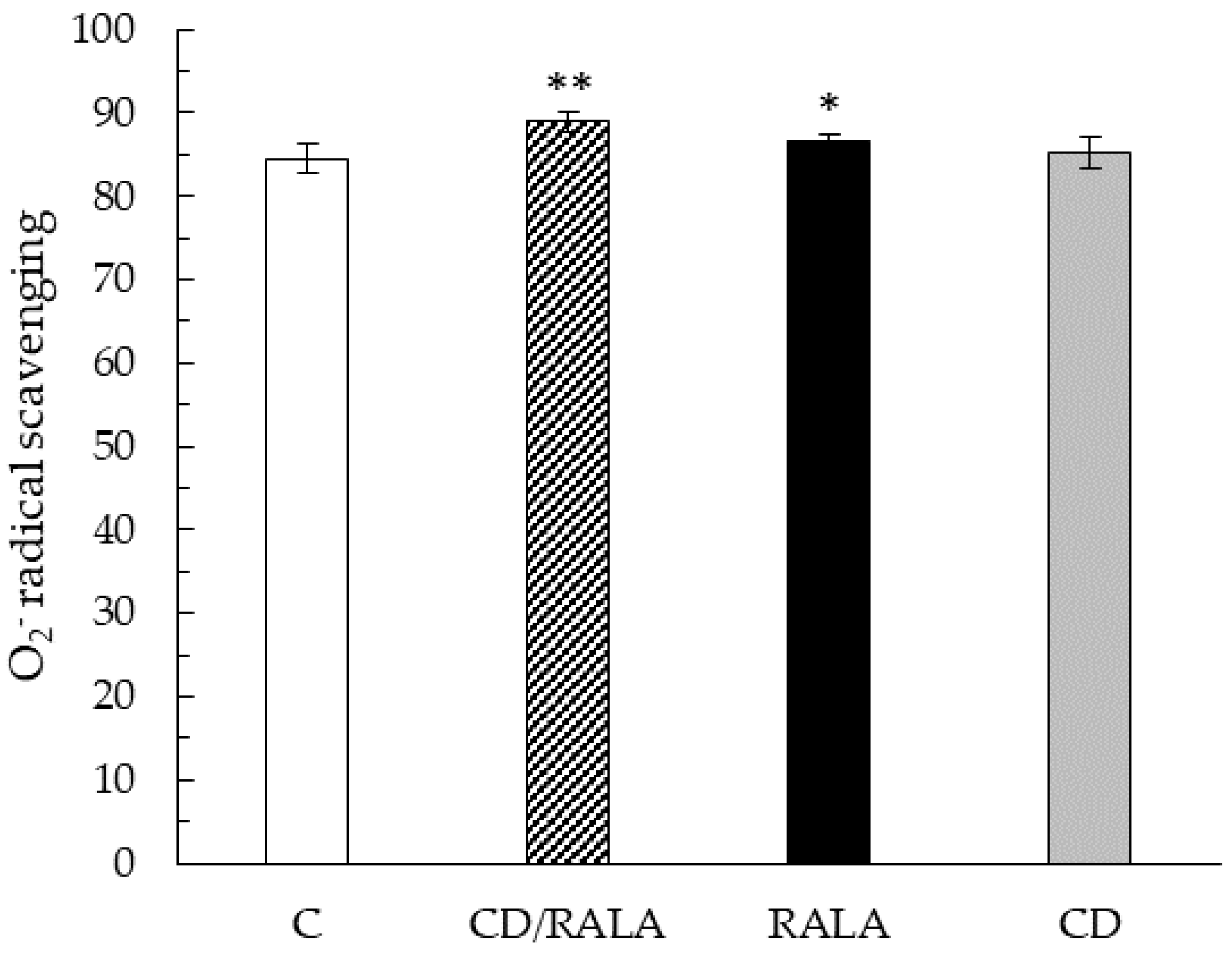
3.3. O2− Radical Scavenging
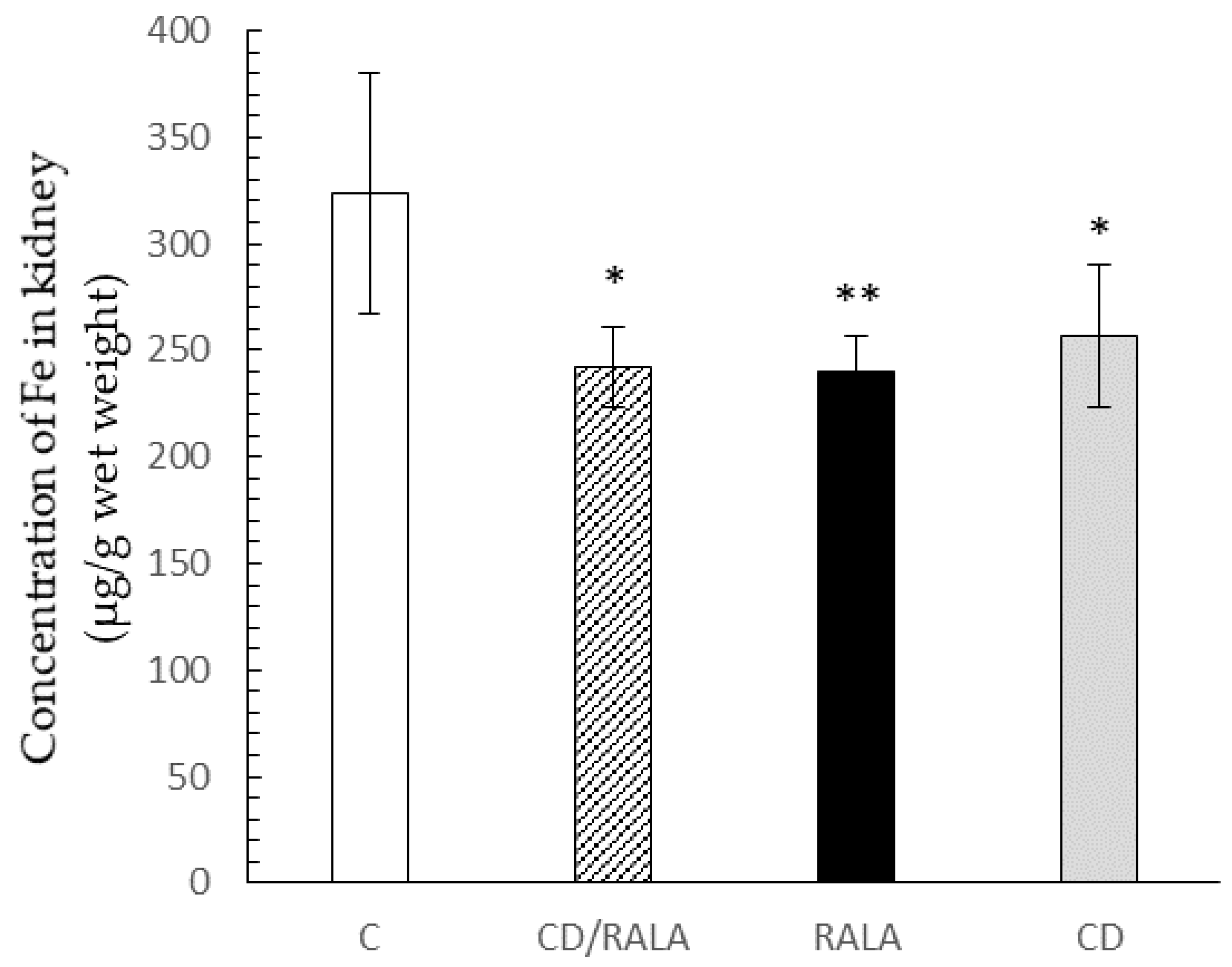
3.4. Trace Element Analysis
3.5. Other Experiments
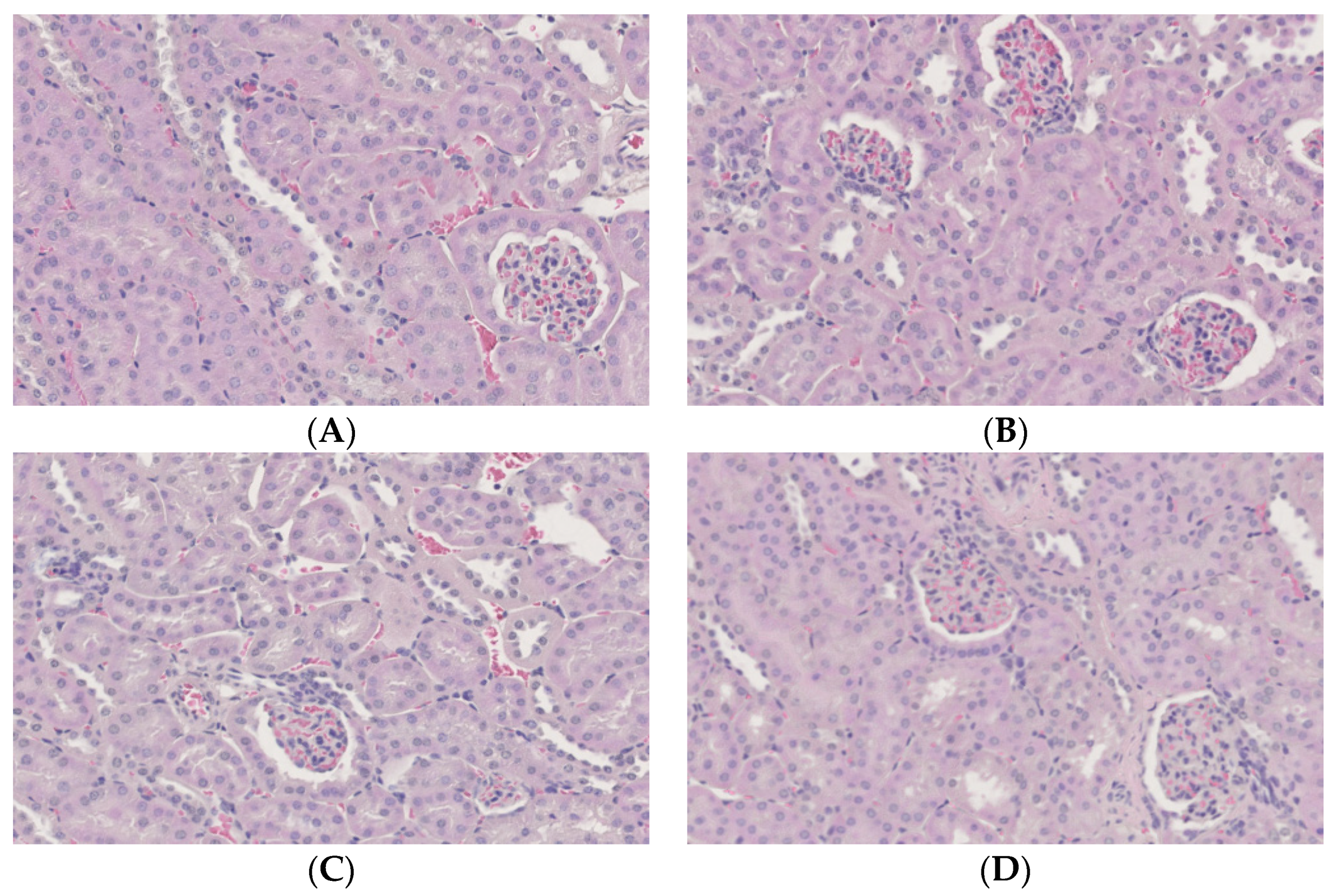
3.6. Histopathological and Immunohistochemical Examination of the Kidney
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reed, L.J. A trail of research from lipoic acid to α-keto acid dehydrogenase complexes. J. Biol. Chem. 2001, 276, 38329–38336. [Google Scholar] [CrossRef] [PubMed]
- Itabe, Y. Lifestyle-related disease and oxidative stress. Yakugaku Zasshi 2001, 37, 805–809. [Google Scholar]
- Perham, R. Swinging arms and swinging domains in multifunctional enzymes: Catalytic machines for multistep reactions. Annu. Rev. Biochem. 2000, 69, 961–1004. [Google Scholar] [CrossRef] [PubMed]
- Goraca, A.; Huk-Kolega, H.; Piechota, A.; Kleniwska, P.; Ciejka, E.; Skibska, B. Lipoic acid—Biological activity and therapeutic potential. Pharmacol. Rep. 2011, 63, 849–858. [Google Scholar] [CrossRef]
- Lee, W.J.; Song, K.H.; Kohh, E.H.; Won, J.C.; Kim, H.S.; Park, H.S.; Kim, M.S.; Kim, S.W.; Lee, K.U.; Park, J.Y. α-Lipoic acid increases insulin sensitivity by activating AMPK in skeletal muscle. Biochem. Biophys. Res. Commun. 2005, 332, 885–891. [Google Scholar] [CrossRef]
- Wollin, S.D.; Jones, P.J.H. α-Lipoic acid and cardiovascular disease. J. Nutr. 2003, 133, 3327–3330. [Google Scholar] [CrossRef] [PubMed]
- Song, K.-H.; Lee, W.J.; Koh, J.-M.; Kim, H.S.; Youn, J.-Y.; Park, H.-S.; Koh, E.H.; Kim, M.-S.; Youn, J.H.; Lee, K.-U.; et al. α-Lipoic acid prevents diabetes mellitus in diabetes prone obese rats. Biochem. Biophys. Res. Commun. 2004, 326, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Packer, L.; Witt, E.H.; Tristchler, H.J. Alpha-lipoic acid as a biological antioxidant. Free Radic. Biol. Med. 1995, 19, 227–250. [Google Scholar] [CrossRef]
- Biewenga, G.P.; Haenen, G.R.M.M.; Bast, A. The pharmacology of the antioxidant lipoic acid. Gen. Pharmacol.-Vasc. S 1997, 29, 315–331. [Google Scholar] [CrossRef]
- Steeper, R.S.; Henriksen, E.J.; Jacob, S.; Hokama, J.Y.; Fogt, D.L.; Tritschler, H.J. Differential effects of lipoic acid stereoisomers on glucose metabolism in insulin-resistant skeletal muscle. Am. J. Physiol. 1997, 273, 185–191. [Google Scholar] [CrossRef]
- Matsugo, S.; Konishi, T. Chemical and biochemical properties of natural occurring novel antioxidant, lipoic acid. Oleoscience 2001, 1, 47–54. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Saito, K.; Iwasaki, Y.; Ito, R.; Nakazawa, Y. Enantiomeric determination of α-lipoic acid in dietary supplements by liquid chromatography/mass spectrometry. Bunseki Kagaku 2012, 61, 109–114. [Google Scholar] [CrossRef]
- Arima, H. New development of cyclodextrin in pharmacology. Yakuzaigaku 2017, 77, 54–60. [Google Scholar]
- Sato, K.; Ishida, Y.; Ikuta, N.; Uekaji, Y.; Nakata, D.; Terao, K. Stabilization of unstable functional food ingredients by complexation with cyclodextrin. Oleo Sci. 2013, 13, 123–130. [Google Scholar] [CrossRef]
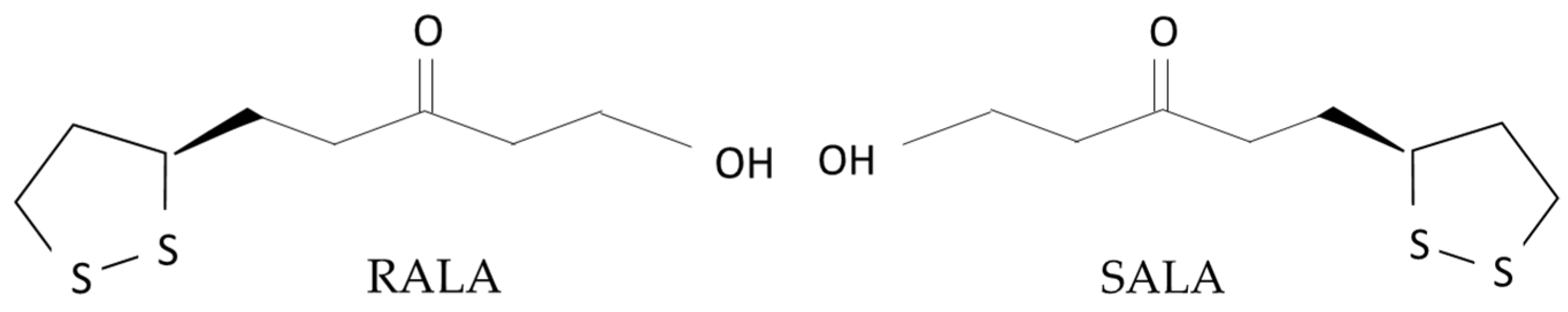
- Ikuta, N.; Shiga, H.; Uekaji, Y.; Jo, A.; Ishida, Y.; Nakata, D.; Kamoi, K.; Nakano, M.; Terao, K. Stabilization of R (+) alpha lipoic acid by complexation with γ-cyclodextrin. J. Jpn Soc. Clin. Nutr. 2011, 33, 53–60. [Google Scholar]
- Nelson, M.E.; Rejeski, W.J.; Blair, S.N.; Duncan, P.W.; Judge, J.O.; King, A.C.; Macera, C.A.; Castaneda-Sceppa, C. Physical activity and public health in older adults: Recommendation from the American College of Sports Medicine and the American Heart Association. Circulation 2007, 116, 1094–1105. [Google Scholar] [CrossRef] [PubMed]
- Powell, K.E.; Palch, A.E.; Blair, S.N. Physical activity for health: What kind? How Much? How Intense? On top of what? Annu. Rev. Public Health 2011, 32, 349–365. [Google Scholar] [CrossRef]
- Arem, H.; Moore, S.C.; Patel, A.; Patel, A.; Hartge, P.; Gonzalez, A.B.; Visvanathan, K.; Campbell, P.; Freedman, M.; Weiderpass, E. Leisure time physical activity and mortality: A detailed pooled analysis of the dose-response relationship. JAMA Intern. Med. 2015, 175, 959–967. [Google Scholar] [CrossRef]
- Stessman, J.; Hammerman-Rozenberg, R.; Cohen, A.; Ein-Mor, E.; Jacobs, J.M. Physical activity, function, and longevity among the very old. Arch. Intern. Med. 2009, 169, 1476–1483. [Google Scholar] [CrossRef]
- Chodzko-Zajiko, W.J.; Proctor, D.N.; Singh, M.A.F.; Minson, C.T.; Nigg, C.R.; Salem, G.J.; Skinner, J.S. American College of Sports Medicine. Exercise and physical activity for older adults. Med. Sci. Sports Exerc. 2009, 41, 1510–1530. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.C.; Lee, I.-M.; Weiderpass, E.; Campbell, P.T.; Sampson, J.N.; Kitahara, C.M.; Keadle, S.K.; Arem, H.; Gonzalez, A.B.; Hartge, P. Association of leisure-time physical activity with risk of 26 type of cancer in 1.44 million adults. JAMA Intern. Med. 2016, 176, 816–825. [Google Scholar] [CrossRef]
- Franco, O.H.; de Laet, C.; Peeters, A.; Jonker, J.; Mackenbach, J.; Nusselder, W. Effects of physical activity on life expectancy with cardiovascular disease. Arch. Intern. Med. 2005, 165, 2355–2360. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.P.; Wai, J.P.M.; Tsai, M.K.; Yang, Y.C.; Cheng, T.Y.D.; Lee, M.-C.; Chan, H.T.; Tsao, C.K.; Tsai, S.P.; Wu, X. Minimum amount of physical activity for reduced mortality and extended life expectancy: A prospective cohort study. Lancet 2011, 378, 1244–1253. [Google Scholar] [CrossRef]
- da Silva Borqes, L.; Dermarqos, A.; da Silva, E., Jr. P.; Weimann, E.; Lambertucci, R.H.; Hatanaka, E. Melatonin decreases muscular oxidative stress and inflammation induced by strenuous exercise and stimulates growth factor synthesis. J. Pineal Res. 2015, 58, 166–172. [Google Scholar] [CrossRef]
- Jablan, J.; Inic, S.; Stosnach, H.; Hadziabdic, M.O.; Vujic, L.; Domijan, A.-M. Level of minerals and trace elements in the urine of the participants of mountain ultra-marathon race. J. Trace Elem. Med. Biol. 2017, 41, 54–59. [Google Scholar] [CrossRef]
- McClung, J.P.; Gaffney-Stomberg, E.; Lee, J.J. Female athletes: A population at risk of vitamin and mineral deficies affecting health and performance. J. Trace Elem. Med. Biol. 2014, 28, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, J.; Oki, T.; Takebayashi, J.; Yamasaki, K.; Tsushida, T. Toward the unification if methods for measuring the antioxidant capacity of foods usefulness of ORAC method and its correlation with other measurement methods. Sci. Biol. 2009, 47, 237–243. [Google Scholar]
- Nakata, D.; Uekaji, Y.; Shiga, K.; Jo, A.; Terao, K.; Nakano, M.; Nakanishi, K. Effect of γ-cyclodextrin inclusion of α-lipoic acid and coenzyme Q10 on swimming time in mice. J. Jpn Soc. Clin. Nutr. 2011, 32, 263–266. [Google Scholar]
- Kadowaki, S.; Kaburugi, T. Effect of a low carbohydrate-high protein diet in non-obese mice. Jpn. J. Nutr. Diet. 2016, 74, 51–59. [Google Scholar] [CrossRef][Green Version]
- Yuki, N.; Yutaka, Y.; Katsuhiko, Y.; Akiko, T.; Hiroyuki, Y. Beneficial effect of bis (hinokitiolato) Zn complex on high-fat diet-induced lipid accumulation in mouse liver and kidney. In Vivo 2017, 31, 1145–1151. [Google Scholar]
- Matsumoto, K.; Ishihara, K.; Tanaka, K.; Inoue, K.; Fushiki, T. An adjustable-current swimming pool for the evaluation of endurance capacity of mice. J. Appl. Physiol. 1996, 81, 1843–1849. [Google Scholar] [CrossRef]
- Kim, K.-M.; Kawada, T.; Ishihara, K.; Inoue, K.; Fushiki, T. Increase in swimming endurance capacity of mice by capsaicin-induced adrenal catecholamine secretion. Biosci. Biotechnol. Biochem. 1997, 61, 1718–1723. [Google Scholar] [CrossRef]
- Suzanne, M.S.; Oscar, A.B. Cytoskeletal protein carbonylation and degradation in experimental autoimmune encephalomyelitis. J. Neurochem. 2007, 105, 763–772. [Google Scholar] [CrossRef]
- Mutch, B.J.; Banister, E.W. Ammonia metabolism in exercise and fatigue: A review. Med. Sci. Sports Exerc. 1983, 15, 41–50. [Google Scholar] [CrossRef]
- Graham, T.E.; MacLean, D.A. Ammonia and amino acid metabolism in human skeletal muscle during exercise. Can. J. Physiol. Pharmacol. 1992, 70, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Wagenmakers, A.J.; Coakley, J.H.; Edwards, R.H.T. Metabolism of branched-chain amino acids and ammonia during exercise: Clues from McArdle’s disease. Int. J. Sports Med. 1990, 11, S101–S113. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, S.; Venditti, P. Mitochondria in exercise-induced oxidative stress. Biol. Signals Recept 2001, 10, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Davison, G.W.; Morgan, R.M.; Hiscock, N.; Garcia, J.M.; Grace, F.; Boisseau, N.; Davies, B.; Castell, L.; McEneny, J.; Young, I.S.; et al. Manipulation of systemic oxygen flux by acute exercise and normobaric hypoxia: Implications for reactive oxygen species generation. Clin. Sci. 2006, 110, 133–141. [Google Scholar] [CrossRef]
- Qaisar, R.; Bhaskaran, S.; Remmen, H.V. Muscle fiber type diversification during exercise and regeneration. Free Radic. Biol. Med. 2016, 98, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Merry, T.L.; Ristow, M. Do antioxidant supplements interfere with skeletal muscle adaptation to exercise training? J. Physiol. 2016, 594, 5135–5147. [Google Scholar] [CrossRef]
- Wada, M.; Sakamoto, M.; Sugiyama, M.; Matsunaga, S. Possible factors contributing to muscle fatigue during intense exercise: Effects of inorganic phosphate, glycogen and reactive oxygen species. Taiikugaku Kenkyu 2006, 51, 399–408. [Google Scholar] [CrossRef][Green Version]
- Arivazhagan, P.; Ramanathan, K.; Panneerselvam, C. Effect of DL-alpha-lipoic acid on mitochondrial enzymes in aged rats. Chem. Biol. Interact. 2001, 138, 189–198. [Google Scholar] [CrossRef]
- Reddy, M.B.; Clark, L. Iron, oxidative stress, and disease risk. Nutr. Rev. 2004, 62, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Rochette, L.; Chibu, S.; Muresan, A.; Vergely, C. Alpha-lipoic acid: Molecular mechanism and therapeutic potential in diabetes. Can. J. Pharmacol. 2015, 93, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, H.; Tanaka, J.; Seki, A.; Honda, H.; Akaogi, S.; Komatsubara, H.; Suzuki, N.; Kameyama, M.; Tamura, S.; Murakami, N. Safety and Structural Analysis of Polymers Produced in Manufacturing Process of α-Lipoic Acid. Food Hyg. Saf. Sci. 2007, 48, 125–131. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Compounds | Solvent (mL/g BW) | Dose (mg/kg BW) |
|---|---|---|---|
| C | Distilled Water | 0.01 | - |
| CD/RALA | RALA-γCD | 0.01 | 50 |
| RALA | RALA | 0.01 | 50 |
| CD | γCD | 0.01 | 50 |
| Organs | C | CD/RALA | RALA | CD |
|---|---|---|---|---|
| Body weight (g) | 22.5 ± 1.8 | 23.3 ± 1.4 | 22.1 ± 1.0 | 22.8 ± 1.0 |
| Food intake (g/day) | 3.0 ± 0.4 | 3.0 ± 0.5 | 2.8 ± 0.5 | 2.9 ± 0.3 |
| Liver (mg/kg BW) | 47.6 ± 1.9 | 47.4 ± 4.8 | 44.2 ± 1.5 | 45.0 ± 2.3 |
| Kidney (mg/kg BW) | 12.7 ± 0.8 | 13.0 ± 0.9 | 13.2 ± 0.7 | 12.5 ± 0.6 |
| Spleen (mg/kg BW) | 2.6 ± 0.2 | 2.6 ± 0.5 | 2.5 ± 0.2 | 2.5 ± 0.2 |
| Lung (mg/kg BW) | 6.8 ± 0.3 | 7.4 ± 1.6 | 7.4 ± 0.6 | 7.6 ± 0.4 |
| Heart (mg/kg BW) | 4.8 ± 0.3 | 4.9 ± 0.4 | 4.8 ± 0.4 | 4.6 ± 0.3 |
| Organs | C | CD/RALA | RALA | CD |
|---|---|---|---|---|
| Liver | 88 ± 1 | 88 ± 2 | 88 ± 1 | 84 ± 3 * |
| Kidney | 84 ± 2 | 89 ± 1 ** | 87 ± 1 * | 85 ± 2 |
| Spleen | 57 ± 9 | 51 ± 14 | 51 ± 11 | 62 ± 5 |
| Lung | 37 ± 8 | 38 ± 6 | 30 ± 5 | 24 ± 10 |
| Quadriceps | 44 ± 8 | 41 ± 10 | 34 ± 11 | 37 ± 4 |
| Gastrocnemius | 23 ± 9 | 20 ± 11 | 20 ± 14 | 8 ± 14 |
| Experimental item | C | CD/RALA | RALA | CD |
|---|---|---|---|---|
| Swimming exercise time to exhaustion (minutes) | 49.9 ± 6.1 | 57.0 ± 6.7 | 56.7 ± 4.6 | 52.1 ± 12.3 |
| Blood lactate level defference before and after training (nmol/L) | 1.5 ± 1.0 | 2.5 ± 1.3 | 1.3 ± 1.1 | 1.7 ± 1.5 |
| Blood CPK activity (U/L) | 101 ± 29 | 80 ± 22 | 69 ± 25 | 88 ± 22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hashimoto, Y.; Yoshizawa, K.; Kaido, Y.; Takenouchi, A.; Terao, K.; Yasui, H.; Yoshikawa, Y. Exercise Performance Upregulatory Effect of R-α-Lipoic Acid with γ-Cyclodextrin. Nutrients 2022, 14, 21. https://doi.org/10.3390/nu14010021
Hashimoto Y, Yoshizawa K, Kaido Y, Takenouchi A, Terao K, Yasui H, Yoshikawa Y. Exercise Performance Upregulatory Effect of R-α-Lipoic Acid with γ-Cyclodextrin. Nutrients. 2022; 14(1):21. https://doi.org/10.3390/nu14010021
Chicago/Turabian StyleHashimoto, Yuki, Katsuhiko Yoshizawa, Yuka Kaido, Akiko Takenouchi, Keiji Terao, Hiroyuki Yasui, and Yutaka Yoshikawa. 2022. "Exercise Performance Upregulatory Effect of R-α-Lipoic Acid with γ-Cyclodextrin" Nutrients 14, no. 1: 21. https://doi.org/10.3390/nu14010021
APA StyleHashimoto, Y., Yoshizawa, K., Kaido, Y., Takenouchi, A., Terao, K., Yasui, H., & Yoshikawa, Y. (2022). Exercise Performance Upregulatory Effect of R-α-Lipoic Acid with γ-Cyclodextrin. Nutrients, 14(1), 21. https://doi.org/10.3390/nu14010021