
Effects of Anthocyanin on Intestinal Health: A Systematic Review

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Eligibility Criteria
2.2. Information Source
2.3. Search Strategy
2.4. Selection, Data Collection Process and Data Items
- -
- Gut microbiota: short chain fatty acids (caecal, fecal or in the serum); intraluminal pH (ileal, caecal or feces); microbial quantification; secretory immunoglobulin A (sIgA);
- -
- Epithelial physical barrier: tight junction proteins; proteins of intestinal brush border membrane; intestinal permeability; plasm endotoxin;
- -
- Intestinal morphology: number of goblet cells; length, height and depth of villi and crypts; mucin secretion; antimicrobial peptides.
2.5. Study Risk-of-Bias Assessment
3. Results
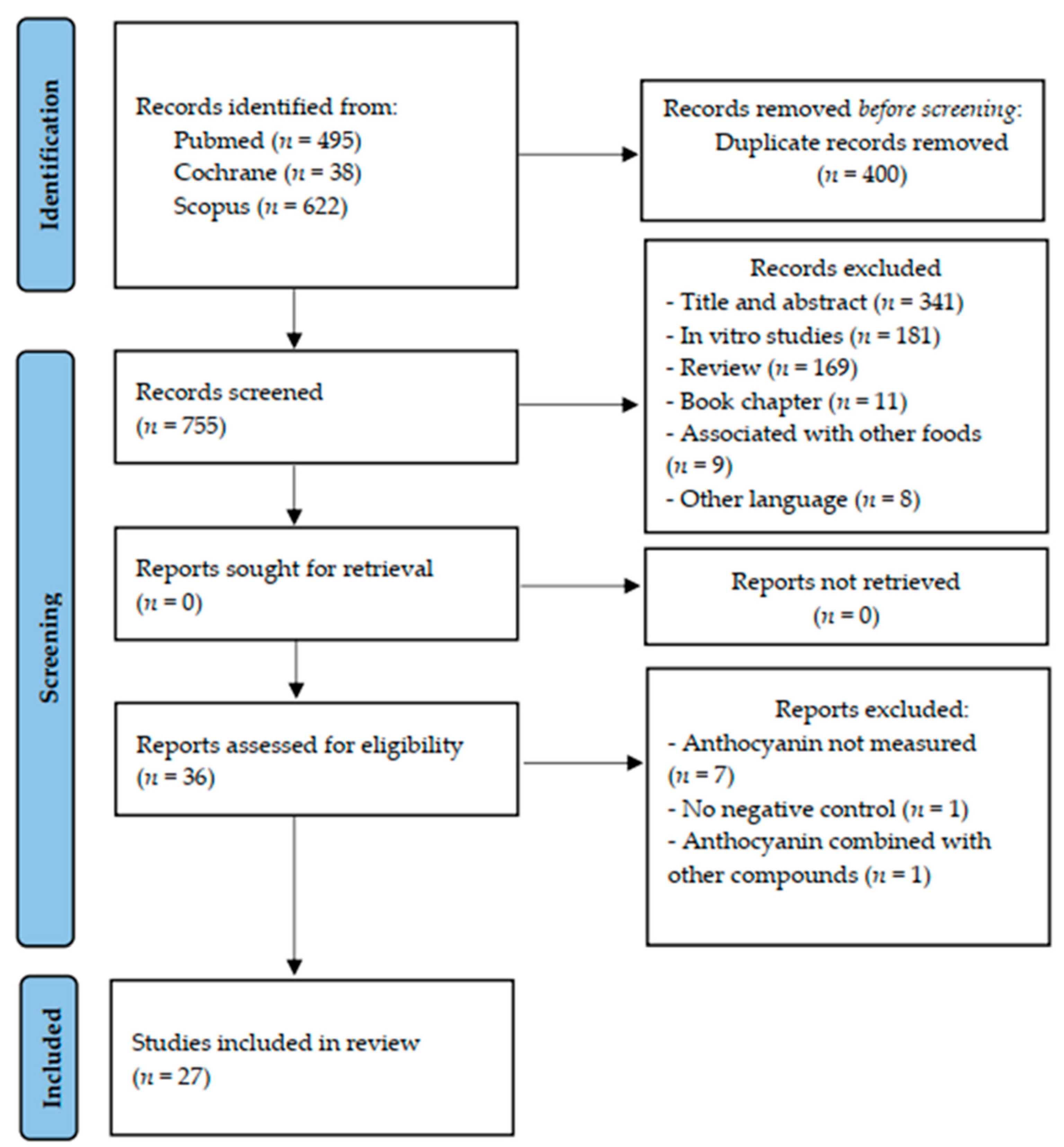
3.1. Study Selection
3.2. Study Characteristics
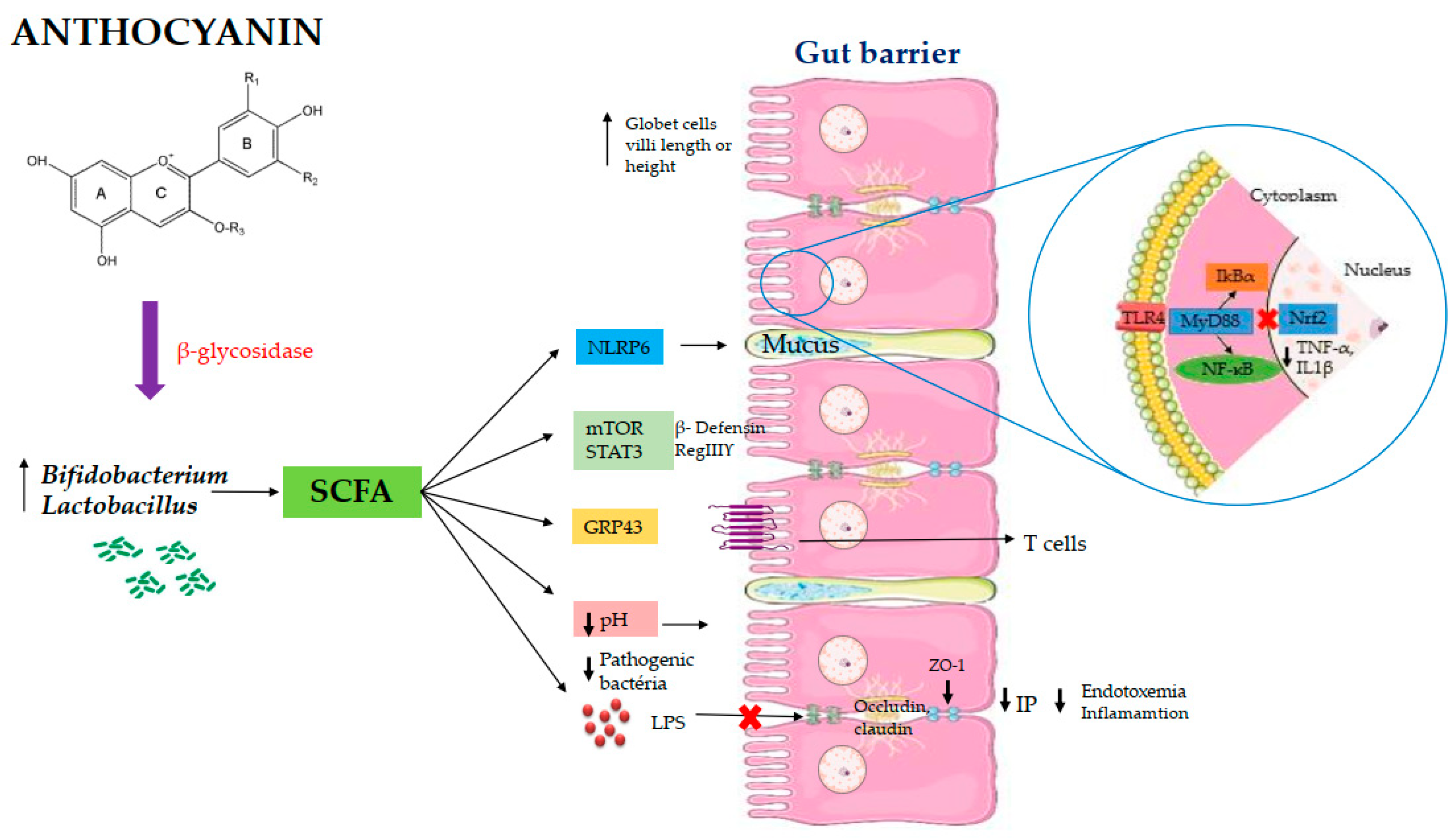
3.3. Main Findings
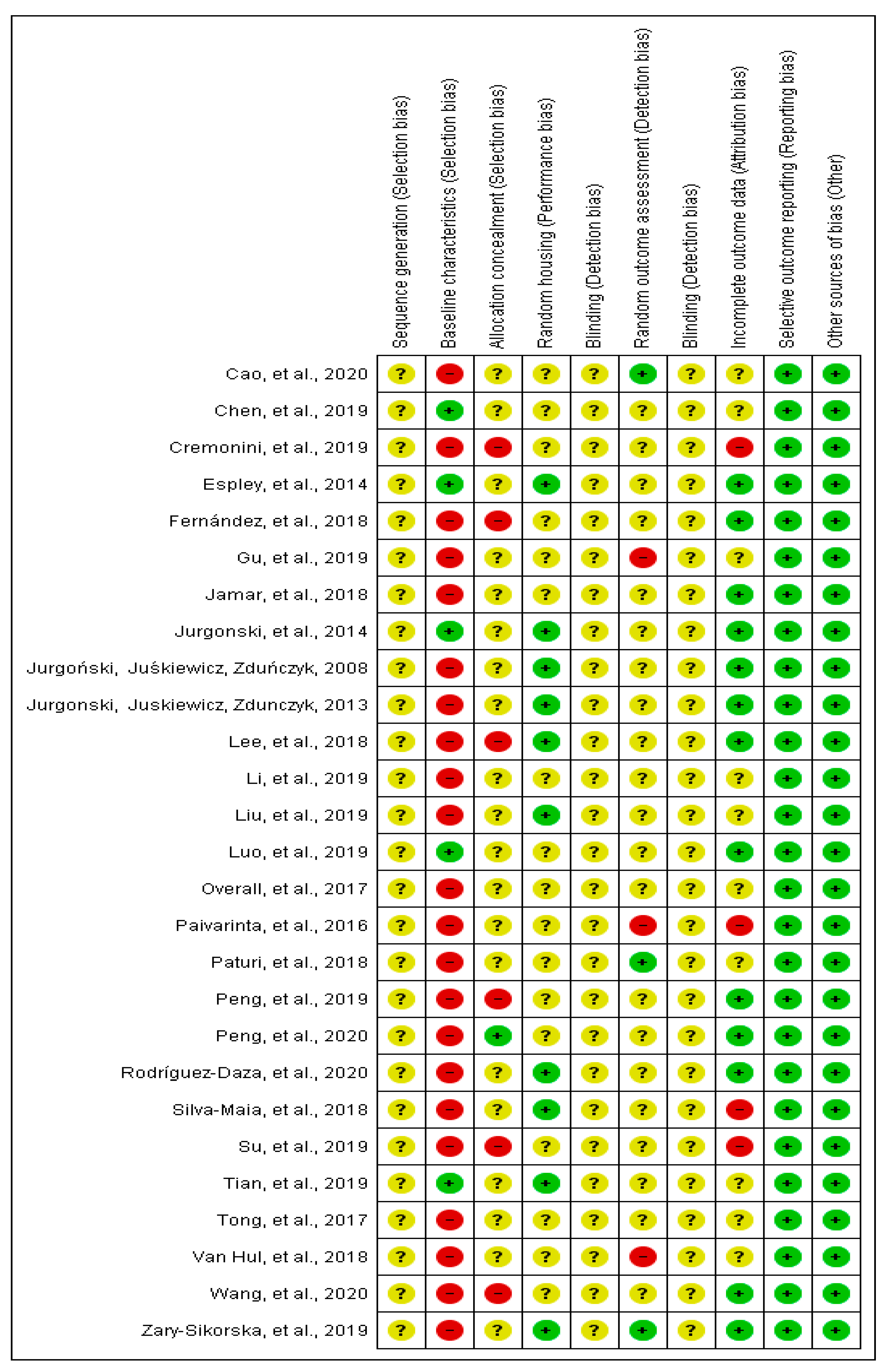
3.4. Risk of Bias
4. Discussion
Dosage and Reporting Quality
5. Conclusions
Registration and Protocol
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bischoff, S.C. “Gut health”: A new objective in medicine? BMC Med. 2011, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, K. The intestinal microbiota and its role in human health and disease. J. Med. Investig. 2016, 63, 27–37. [Google Scholar] [CrossRef]
- Natividad, J.M.M.; Verdu, E.F. Modulation of intestinal barrier by intestinal microbiota: Pathological and therapeutic implications. Pharm. Res. 2013, 69, 42–51. [Google Scholar] [CrossRef]
- Sui, X.; Zhang, Y.; Jiang, L.; Zhou, W. Anthocyanins in food. Encycl. Food Chem. 2019, 2, 10–17. [Google Scholar]
- Curtis, P.J.; van der Velpen, V.; Berends, L.; Jennings, A.; Feelisch, M.; Umpleby, A.M.; Evans, M.; Fernandez, B.O.; Meiss, M.S.; Minnion, M.; et al. Blueberries improve biomarkers of cardiometabolic function in participants with metabolic syndrome—Results from a 6-month, double-blind, randomized controlled trial. Am. J. Clin. Nutr. 2019, 109, 1535–1545. [Google Scholar] [CrossRef]
- Li, D.; Zhang, Y.; Liu, Y.; Sun, R.; Xia, M. Purified anthocyanin supplementation reduces dyslipidemia, enhances antioxidant capacity, and prevents insulin resistance in diabetic. J. Nutr. Dis. 2015, 145, 742–748. [Google Scholar] [CrossRef]
- Jayarathne, S.; Stull, A.J.; Park, O.; Kim, J.H.; Thompson, L.; Moustaid-Moussa, N. Protective effects of anthocyanins in obesity-associated inflammation and changes in gut microbiome. Mol. Nutr. Food Res. 2019, 63, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Edirisinghe, I.; Wei, H.; Vijayakumar, L.P.; Banaszewski, K.; Cappozzo, J.C.; Burton-Freeman, B. A dose-response evaluation of freeze-dried strawberries independent of fiber content on metabolic indices in abdominally obese individuals with insulin resistance in a randomized, single-blinded, diet-controlled crossover trial. Mol. Nutr. Food Res. 2016, 60, 1099–1109. [Google Scholar] [CrossRef]
- Tian, L.; Tan, Y.; Chen, G.; Wang, G.; Sun, J.; Ou, S.; Chen, W.; Bai, W. Metabolism of anthocyanins and consequent effects on the gut microbiota. Crit. Rev. Food Sci. Nutr. 2018, 59, 982–991. [Google Scholar] [CrossRef] [PubMed]
- Mcghie, T.K.; Walton, M.C. Review The bioavailability and absorption of anthocyanins: Towards a better understanding. Mol. Nutr. Food Res. 2007, 51, 702–713. [Google Scholar] [CrossRef] [PubMed]
- Fang, J. Bioavailability of anthocyanins. Drug Metab. Rev. 2014, 46, 508–520. [Google Scholar] [CrossRef]
- Morais, C.A.; Rosso, V.V.; Estadella, D.; Pisani, L.P. Anthocyanins as inflammatory modulators and the role of the gut microbiota. J. Nutr. Biochem. 2016, 33, 1–7. [Google Scholar] [CrossRef]
- Braga, A.R.C.; Murador, D.C.; Mesquita, L.M.D.S.; de Rosso, V.V. Bioavailability of anthocyanins: Gaps in knowledge, challenges and future research. J. Food Compos. Anal. 2018, 68, 31–40. [Google Scholar] [CrossRef]
- Igwe, E.O.; Charlton, K.E.; Probst, Y.C.; Kent, K.; Netzel, M.E. A systematic literature review of the effect of anthocyanins on gut microbiota populations. J. Hum. Nutr. Diet. 2019, 32, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Hooijmans, C.R.; Rovers, M.M.; De Vries, R.B.M.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.T.; Altman, D.G.; Gøtzsche, P.C.; Jüni, P.; Moher, D.; Oxman, A.D.; Savovic, J.; Schulz, K.F.; Weeks, L.; Sterne, J.A.C.; et al. The Cochrane Collaboration’s tool for assessing risk of bias in randomised trials. BMJ 2011, 343, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tong, T.; Niu, Y.H.; Yue, Y.; Wu, S.-C.; Ding, H. Beneficial effects of anthocyanins from red cabbage (Brassica oleracea L. var. capitata L.) administration to prevent irinotecan-induced mucositis. J. Funct. Foods 2017, 32, 9–17. [Google Scholar] [CrossRef]
- Chen, G.; Wang, G.; Zhu, C.; Jiang, X.; Sun, J.; Tian, L.; Bai, W. Effects of cyanidin-3-O-glucoside on 3-chloro-1,2-propanediol induced intestinal microbiota dysbiosis in rats. Food Chem. Toxicol. 2019, 133, 1–9. [Google Scholar] [CrossRef]
- Li, J.; Wu, T.; Li, N.; Wang, X.; Chen, G.; Lyu, X. Bilberry anthocyanin extract promotes intestinal barrier function and inhibits digestive enzyme activity by regulating the gut microbiota in aging rats. Food Funct. 2019, 10, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yan, Y.; Wan, P.; Chen, D.; Ding, Y.; Ran, L.; Mi, J.; Lu, L.; Zhang, Z.; Li, X. Gut microbiota modulation and anti-inflammatory properties of anthocyanins from the fruits of Lycium ruthenicum Murray in dextran sodium sulfate-induced colitis in mice. Free Radic. Biol. Med. 2019, 136, 96–108. [Google Scholar] [CrossRef]
- Su, H.; Xie, L.; Xu, Y.; Ke, H.; Bao, T.; Li, Y.; Chen, W. Pelargonidin-3- O-glucoside derived from wild raspberry exerts antihyperglycemic effect by inducing autophagy and modulating gut microbiota. J. Agric. Food Chem. 2019, 68, 13025–13037. [Google Scholar] [CrossRef]
- Tian, B.; Zhao, J.; An, W.; Zhang, J.; Cao, X.; Mi, J. Lycium ruthenicum diet alters the gut microbiota and partially enhances gut barrier function in male C57BL/6 mice. J. Funct. Foods 2019, 52, 516–528. [Google Scholar] [CrossRef]
- Luo, Y.; Fang, J.L.; Yuan, K.; Jin, S.H.; Guo, Y. Ameliorative effect of purified anthocyanin from Lycium ruthenicum on atherosclerosis in rats through synergistic modulation of the gut microbiota and NF-κB/SREBP-2 pathways. J. Funct. Foods 2019, 59, 223–233. [Google Scholar] [CrossRef]
- Peng, Y.; Yan, Y.; Wan, P.; Dong, W.; Huang, K.; Ran, L.; Mi, J.; Lu, L.; Zheng, X.; Cao, Y. Effects of long-term intake of anthocyanins from Lycium ruthenicum Murray on the organism health and gut microbiota in vivo. Food Res. Int. 2020, 130, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, D.; Ji, Y.; Liu, Y.; Xu, L.; Guo, Y. Dietary supplementation of black rice anthocyanin extract regulates cholesterol metabolism and improves gut microbiota dysbiosis in C57BL/6J mice fed a high-fat and cholesterol diet. Mol. Nutr. Food Res. 2020, 64, e1900876. [Google Scholar] [CrossRef] [PubMed]
- Overall, J.; Bonney, S.A.; Wilson, M.; Beermann, A.; Grace, M.H.; Esposito, D.; Lila, M.A.; Komarntysky, S. Metabolic effects of berries with structurally diverse anthocyanins. Int. J. Mol. Sci. 2017, 18, 422. [Google Scholar] [CrossRef] [PubMed]
- Cremonini, E.; Daveri, E.; Mastaloudis, A.; Adamo, A.M.; Mills, D.; Kalanetra, K.; Hester, S.N.; Wood, S.M.; Fraga, C.G.; Oteiza, P.I. Anthocyanins protect the gastrointestinal tract from high fat diet-induced alterations in redox signaling, barrier integrity and dysbiosis. Redox. Biol. 2019, 26, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Thomas-Ahner, J.M.; Riedl, K.M.; Bailey, M.T.; Vodovotz, Y.; Schwartz, S.J.; Clinton, S.K. Dietary black raspberries impact the colonic microbiome and phytochemical metabolites in mice. Mol. Nutr. Food Res. 2019, 63, 1–9. [Google Scholar] [CrossRef]
- Liu, F.; Wang, T.T.Y.; Tang, Q.; Xue, C.; Li, R.W.; Wu, V.C.H. Malvidin 3-Glucoside modulated gut microbial dysbiosis and global metabolome disrupted in a murine colitis model induced by dextran sulfate sodium. Mol. Nutr. Food Res. 2019, 63, 1–14. [Google Scholar] [CrossRef]
- Cao, L.; Lee, S.G.; Melough, M.M.; Sakaki, J.R.; Maas, K.R.; Koo, S.I.; Chun, O.K. Long-term blackcurrant supplementation modified gut microbiome profiles in mice in an age- dependent manner: An exploratory study. Nutrients 2020, 12, 290. [Google Scholar] [CrossRef]
- Espley, R.V.; Butts, C.A.; Laing, W.A.; Martell, S.; Smith, H.; McGhie, T.K.; Zhang, J.; Paturi, G.; Hedderley, D.; Bovy, A.; et al. Dietary flavonoids from modified apple reduce inflammation markers and modulate gut microbiota in mice. J. Nutr. 2014, 144, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Päivärinta, E.; Niku, M.; Maukonen, J.; Storvik, M.; Heiman-Lindh, A.; Saarela, M.; Pajari, A.M.; Mutanen, M. Changes in intestinal immunity, gut microbiota, and expression of energy metabolism–related genes explain adenoma growth in bilberry and cloudberry-fed ApcMin mice. Nutr. Res. 2016, 36, 1285–1297. [Google Scholar] [CrossRef]
- Van Hul, M.; Geurts, L.; Plovier, H.; Druart, C.; Everard, A.; Ståhlman, M.; Rhimi, M.; Chira, K.; Teissedre, P.L.; Delzenne, N.M.; et al. Reduced obesity, diabetes, and steatosis upon cinnamon and grape pomace are associated with changes in gut microbiota and markers of gut barrier. Am. J. Physiol. Endocrinol. Metab. 2018, 314, E334–E352. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Daza, M.C.; Daoust, L.; Boutkrabt, L.; Pilon, G.; Varin, T.; Dudonné, S.; Levy, E.; Marette, A.; Roy, D.; Desjardins, Y. Wild blueberry proanthocyanidins shape distinct gut microbiota profile and influence glucose homeostasis and intestinal phenotypes in high-fat high-sucrose fed mice. Sci. Rep. 2020, 10, 1–16. [Google Scholar]
- Jurgoński, A.; Juśkiewicz, J.; Zduńczyk, Z. Ingestion of black chokeberry fruit extract leads to intestinal and systemic changes in a rat model of prediabetes and hyperlipidemia. Plant. Foods Hum. Nutr. 2008, 63, 176–182. [Google Scholar] [CrossRef]
- Jurgoński, A.; Juśkiewicz, J.; Zduńczyk, Z. An anthocyanin-rich extract from Kamchatka honeysuckle increases enzymatic activity within the gut and ameliorates abnormal lipid and glucose metabolism in rats. Nutrition 2013, 29, 898–902. [Google Scholar] [CrossRef]
- Fernández, J.; García, L.; Monte, J.; Villar, C.J.; Lombó, F. Functional anthocyanin-rich sausages diminish colorectal cancer in an animal model and reduce pro-inflammatory bacteria in the intestinal microbiota. Genes 2018, 9, 133. [Google Scholar] [CrossRef]
- Jamar, G.; Santamarina, A.B.; Mennitti, L.V.; de Cesar, H.C.; Oyama, L.M.; de Rosso, V.V.; Pisani, L.P. Bifidobacterium spp. reshaping in the gut microbiota by low dose of juçara supplementation and hypothalamic insulin resistance in Wistar rats. J Funct. Foods 2018, 46, 212–219. [Google Scholar] [CrossRef]
- Lee, S.; Keirsey, K.I.; Kirkland, R.; Grunewald, Z.I.; Fischer, J.G.; de La Serre, C.B. Blueberry supplementation influences the gut microbiota, inflammation, and insulin resistance in high-fat-diet-fed rats. J. Nutr. 2018, 148, 209–219. [Google Scholar] [CrossRef]
- Paturi, G.; Butts, C.A.; Monro, J.A.; Hedderley, D. Effects of blackcurrant and dietary fibers on large intestinal health biomarkers in rats. Plant. Foods Hum. Nutr. 2018, 73, 54–60. [Google Scholar] [CrossRef]
- da Silva-Maia, J.K.; Batista, A.G.; Correa, L.C.; Lima, G.C.; Bogusz Junior, S.; Maróstica Junior, M.R. Aqueous extract of berry (Plinia jaboticaba) byproduct modulates gut microbiota and maintains the balance on antioxidant defense system in rats. J. Food Biochem. 2018, 43, 1–11. [Google Scholar] [CrossRef]
- Żary-Sikorska, E.; Fotschki, B.; Fotschki, J.; Wiczkowski, W.; Juśkiewicz, J. Preparations from purple carrots containing anthocyanins improved intestine microbial activity, serum lipid profile and antioxidant status in rats. J. Funct. Foods 2019, 60, 103442. [Google Scholar] [CrossRef]
- Jurgoński, A.; Juśkiewicz, J.; Zduńczyk, Z.; Matusevicius, P.; Kołodziejczyk, K. Polyphenol-rich extract from blackcurrant pomace attenuates the intestinal tract and serum lipid changes induced by a high-fat diet in rabbits. Eur. J. Nutr. 2014, 53, 1603–1613. [Google Scholar] [CrossRef]
- Hugenholtz, F.; Zhang, J.; O’Toole, P.W.; Smidt, H. Studying the mammalian intestinal microbiome using animal models. In Manual of Environmental Microbiology, 4th ed.; Yates, M.V., Nakatsu, C.H., Miller, R.V., Pillai, S.D., Eds.; American Society for Microbiolog: Washington, DC, USA, 2015. [Google Scholar]
- Faria, A.; Fernandes, I.; Norberto, S.; Mateus, N.; Calhau, C. Interplay between anthocyanins and gut microbiota. J. Agric. Food Chem. 2014, 62, 6898–6902. [Google Scholar] [CrossRef] [PubMed]
- Crovesy, L.; Masterson, D.; Rosado, E.L. Profile of the gut microbiota of adults with obesity: A systematic review. Eur J Clin Nutr. 2020, 74, 1251–1262. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Reddy, D.N. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787–8809. [Google Scholar] [CrossRef]
- Ibrahim, M.; Anishetty, S. A meta-metabolome network of carbohydrate metabolism: Interactions between gut microbiota and host. Biochem. Biophys. Res. Commun. 2012, 428, 278–284. [Google Scholar] [CrossRef]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut. Microbes. 2018, 9, 308–325. [Google Scholar] [CrossRef]
- Markowiak, P.; Ślizewska, K. Effects of probiotics, prebiotics, and synbiotics on human health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Boto-Ordóñez, M.; Urpi-Sarda, M.; Queipo-Ortuño, M.I.; Tulipani, S.; Tinahones, F.J.; Andres-Lacueva, C. High levels of Bifidobacteria are associated with increased levels of anthocyanin microbial metabolites: A randomized clinical trial. Food Funct. 2014, 5, 1932–1938. [Google Scholar] [CrossRef]
- Hidalgo-Cantabrana, C.; Delgado, S.; Ruiz, L.; Ruas-Madiedo, P.; Sánchez, B.; Margolles, A. Bifidobacteria and their health-promoting effects. Microbiol. Spectr. 2017, 5, 1–19. [Google Scholar]
- Ajiboye, T.O.; Habibu, R.S.; Saidu, K.; Haliru, F.Z.; Ajiboye, H.O.; Aliyu, N.O.; Ibitoye, O.B.; Uwazie, J.N.; Muritala, H.F.; Bello, S.A.; et al. Involvement of oxidative stress in protocatechuic acid-mediated bacterial lethality. Microbiologyopen 2017, 6, 1–10. [Google Scholar] [CrossRef]
- Li, Y.; Xie, Z.; Gao, T.; Li, L.; Chen, Y.; Xiao, D.; Liu, W.; Zou, B.; Lu, B.; Tian, X.; et al. A holistic view of gallic acid-induced attenuation in colitis based on microbiome-metabolomics analysis. Food Funct. 2019, 10, 4046–4061. [Google Scholar] [CrossRef]
- Wang, C.; Yang, S.; Gao, L.; Wang, L.; Cao, L. Carboxymethyl pachyman (CMP) reduces intestinal mucositis and regulates the intestinal microflora in 5-fluorouracil-treated CT26 tumour-bearing mice. Food Funct. 2018, 9, 2695–2704. [Google Scholar] [CrossRef]
- Xiao, S.; Jiang, S.; Qian, D.; Duan, J. Modulation of microbially derived short-chain fatty acids on intestinal homeostasis, metabolism, and neuropsychiatric disorder. Appl. Microbiol. Biotechnol. 2020, 104, 589–601. [Google Scholar] [CrossRef]
- Luu, M.; Visekruna, A. Short-chain fatty acids: Bacterial messengers modulating the immunometabolism of T cells. Eur. J. Immunol. 2019, 49, 842–848. [Google Scholar] [CrossRef]
- Ratajczak, W.; Rył, A.; Mizerski, A.; Walczakiewicz, K.; Sipak, O.; Laszczyńska, M. Immunomodulatory potential of gut microbiome-derived shortchain fatty acids (SCFAs). Acta. Biochim. Pol. 2019, 66, 1–12. [Google Scholar]
- Zhao, Y.; Chen, F.; Wu, W.; Sun, M.; Bilotta, A.J.; Yao, S.; Xiao, Y.; Huang, X.; Eaves-Pyles, T.D.; Golovko, G.; et al. GPR43 mediates microbiota metabolite SCFA regulation of antimicrobial peptide expression in intestinal epithelial cells via activation of mTOR and STAT3. Mucosal. Immunol. 2018, 11, 752–762. [Google Scholar] [CrossRef]
- Prasad, K.N.; Bondy, S.C. Dietary fibers and their fermented short-chain fatty acids in prevention of human diseases. Bioact. Carbohydr. Diet. Fibre 2019, 17, 100170. [Google Scholar] [CrossRef]
- Fachi, J.L.; de Felipe, J.S.; Pral, L.P.; da Silva, B.K.; Corrêa, R.O.; de Andrade, M.C.P.; da Fonseca, D.M.; Basso, P.J.; Camara, N.O.S.; de Sales E Souza, E.L.; et al. Butyrate protects mice from clostridium difficile-induced colitis through an HIF-1-dependent mechanism. Cell. Rep. 2019, 27, 750–761. [Google Scholar] [CrossRef]
- Witten, J.; Samad, T.; Ribbeck, K. Selective permeability of mucus barriers. Curr. Opin. Biotechnol. 2018, 52, 124–133. [Google Scholar] [CrossRef]
- Suzuki, T. Regulation of intestinal epithelial permeability by tight junctions. Cell. Mol. Life Sci. 2013, 70, 631–659. [Google Scholar] [CrossRef]
- Vendrame, S.; Klimis-Zacas, D. Anti-inflammatory effect of anthocyanins via modulation of nuclear factor- kB and mitogen-activated protein kinase signaling cascades. Nutr. Rev. 2015, 73, 348–358. [Google Scholar] [CrossRef]
- Zhang, Q.; Luna-vital, D.; Gonzalez, E.; Mejia, D. Anthocyanins from colored maize ameliorated the inflammatory paracrine interplay between macrophages and adipocytes through regulation of NF-κB and JNK-dependent MAPK pathways. J. Funct. Foods 2019, 54, 175–186. [Google Scholar] [CrossRef]
- Xiao, Y.T.; Yan, W.H.; Cao, Y.; Yan, J.K.; Cai, W. Neutralization of IL-6 and TNF-α ameliorates intestinal permeability in DSS-induced colitis. Cytokine 2016, 83, 189–192. [Google Scholar] [CrossRef]
- Fries, W.; Muja, C.; Crisafulli, C.; Cuzzocrea, S.; Mazzon, E. Dynamics of enterocyte tight junctions: Effect of experimental colitis and two different anti-TNF strategies. Am. J. Physiol. Gastrointest. Liver. Physiol. 2008, 294, 938–947. [Google Scholar] [CrossRef]
- Ferrari, D.; Speciale, A.; Cristani, M.; Fratantonio, D.; Molonia, M.S.; Ranaldi, G.; Cimino, F. Cyanidin-3-O-glucoside inhibits NF-kB signalling in intestinal epithelial cells exposed to TNF-α and exerts protective effects via Nrf2 pathway activation. Toxicol. Lett. 2016, 264, 51–58. [Google Scholar] [CrossRef]
- Tarabova, L.; Makova, Z.; Piesova, E.; Szaboova, R.; Faixova, Z. Intestinal Mucus Layer and Mucins (A Review). Folia. Vet. 2016, 60, 21–25. [Google Scholar] [CrossRef]
- Soderholm, A.T.; Pedicord, V.A. Intestinal epithelial cells: At the interface of the microbiota and mucosal immunity. Immunology 2019, 158, 267–280. [Google Scholar] [CrossRef]
- Furter, M.; Sellin, M.E.; Hansson, G.C.; Hardt, W.D. Mucus architecture and near-surface swimming affect distinct Salmonella Typhimurium infection patterns along the murine intestinal tract. Cell. Rep. 2019, 27, 2665–2678. [Google Scholar] [CrossRef]
- Ghattamaneni, N.K.; Sharma, A.; Panchal, S.K.; Brown, L. Pelargonidin 3-glucoside-enriched strawberry attenuates symptoms of DSS-induced inflammatory bowel disease and diet-induced metabolic syndrome in rats. Eur. J. Nutr. 2020, 59, 2905–2918. [Google Scholar] [CrossRef]
- Park, J.H.; Kotani, T.; Konno, T.; Setiawan, J.; Kitamura, Y.; Imada, S.; Usui, Y.; Hatano, N.; Shinohara, M.; Saito, Y.; et al. Promotion of intestinal epithelial cell turnover by commensal bacteria: Role of short-chain fatty acids. PLoS ONE 2016, 11, e0156334. [Google Scholar] [CrossRef]
- Wlodarska, M.; Thaiss, C.A.; Nowarski, R.; Henao-Mejia, J.; Zhang, J.P.; Brown, E.M.; Frankel, G.; Levy, M.; Katz, M.N.; Philbrick, W.M.; et al. NLRP6 inflammasome orchestrates the colonic host-microbial interface by regulating goblet cell mucus secretion. Cell 2014, 156, 1045–1059. [Google Scholar] [CrossRef]
- Li, J.M.; Yu, R.; Zhang, L.P.; Wen, S.Y.; Wang, S.J.; Zhang, X.Y.; Xu, Q.; Kong, L.D. Dietary fructose-induced gut dysbiosis promotes mouse hippocampal neuroinflammation: A benefit of short-chain fatty acids. Microbiome 2019, 7, 1–14. [Google Scholar] [CrossRef]
- Wang, Y.; Hatabu, T. Mulberry juice freeze-dried powder attenuates the disease severity by the maintaining of colon mucosa in mice with DSS-induced acute colitis. Biosci. Biotechnol. Biochem. 2019, 83, 914–922. [Google Scholar] [CrossRef]
- Crawley, S.W.; Mooseker, M.S.; Tyska, M.J. Shaping the intestinal brush border. J. Cell. Biol. 2014, 207, 441–451. [Google Scholar] [CrossRef]
- Csernus, B.; Biró, S.; Babinszky, L.; Komlosi, I.; Jávor, A.; Stundl, L.; Remenyik, J.; Bai, P.; Olah, J.; Pesti-Asboth, G.; et al. Effect of carotenoids, oligosaccharides and anthocyanins on growth performance, immunological parameters and intestinal morphology in broiler chickens challenged with escherichia coli lipopolysaccharide. Animals 2020, 10, 347. [Google Scholar] [CrossRef]
- Gessner, D.K.; Fiesel, A.; Most, E.; Dinges, J.; Wen, G.; Ringseis, R.; Eder, K. Supplementation of a grape seed and grape marc meal extract decreases activities of the oxidative stress-responsive transcription factors NF-κB and Nrf2 in the duodenal mucosa of pigs. Acta. Vet. Scand. 2013, 55, 1–10. [Google Scholar] [CrossRef]
- Ghosh, S.S.; Wang, J.; Yannie, P.J.; Ghosh, S. Intestinal barrier dysfunction, LPS translocation, and disease development. J. Endocr. Soc. 2020, 4, 1–15. [Google Scholar] [CrossRef]
- Cochet, F.; Peri, F. The role of carbohydrates in the lipopolysaccharide (LPS)/toll-like receptor 4 (TLR4) Signalling. Int. J. Mol. Sci. 2017, 18, 2318. [Google Scholar] [CrossRef]
- Santamarina, A.B.; Jamar, G.; Mennitti, L.V.; de Cesar, H.C.; Vasconcelos, J.R.; Oyama, L.M.; de Rosso, V.V.; Pisani, L.P. Obesity-related inflammatory modulation by juçara berry (Euterpe edulis Mart.) supplementation in Brazilian adults: A double-blind randomized controlled trial. Eur. J. Nutr. 2019, 59, 1693–1705. [Google Scholar] [CrossRef]
- Magro, D.O.; Kotze, P.G.; Martinez, C.A.R.; Camargo, M.G.; Guadagnini, D.; Calixto, A.R.; Vasques, A.C.J.; de Lordes Setsuko Ayrizono, M.; Geloneze, B.; Pareja, J.C.; et al. Changes in serum levels of lipopolysaccharides and CD26 in patients with Crohn’s disease. Intest. Res. 2017, 15, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Martini, E.; Krug, S.M.; Siegmund, B.; Neurath, M.F.; Becker, C. The epithelial barrier and its relationship with mucosal immunity in inflammatory bowel disease. Cell. Mol. Gastroenterol. Hepatol. 2017, 4, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Yan, D.; Xu, M.; Li, F.; Ren, M.; Zhang, J.; Wu, M. Interactive association of lipopolysaccharide and free fatty acid with the prevalence of type 2 diabetes: A community-based cross-sectional study. J. Diabetes Investig. 2019, 10, 1438–1446. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Postigo, M.; Oliva-Olivera, W.; Coin-Aragüez, L.; Ramos-Molina, B.; Giraldez-Perez, R.M.; Lhamyani, S.; Alcaide-Torres, J.; Perez-Martinez, P.; El Bekay, R.; Cardona, F.; et al. Metabolic endotoxemia promotes adipose dysfunction and inflammation in human obesity. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E319–E332. [Google Scholar] [CrossRef]
- Rodrigrez-Morató, J.; Matthan, N.R.; Liu, J.; Torre, R.; Chen, C.Y.O. Cranberries attenuate animal-based diet-induced changes in microbiota composition and functionality: A randomized crossover controlled feeding trial. J. Nutr. Biochem. 2018, 62, 76–86. [Google Scholar] [CrossRef]
- Thounaojam, M.C.; Jadeja, R.N.; Sankhari, J.M.; Devkar, R.V.; Ramachandran, A.V. Safety evaluations on ethanolic extract of red cabbage (Brassica oleracea L.) in Mice. J. Food Sci. 2011, 76, T35–T39. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 71, 372. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Inclusion Criteria | Exclusion Criteria |
|---|---|---|
| Population | In vivo animal studies | Clinical studies and in vitro studies |
| Intervention | Intake of foods’ source of anthocyanin or supplementation with extract of anthocyanin | Anthocyanin associated with other foods or not measured |
| Comparator | Negative control (without the intervention) | No control group |
| Outcomes | Changes in the gut microbiota composition, intraluminal pH, short chain fatty acids, histological parameters of small and large intestine, gene expression of tight junction’s proteins, gene expression of intestinal brush border membrane, integrity of intestinal barrier and intestinal permeability | |
| Study design | Experimental placebo-controlled studies | Review articles, clinical studies, theses, dissertations, book chapters, in vitro experiments and studies published in other languages than English. |
| Author, Year | Country | Animal Model/Age | Sex | Initial Weight (g) | Nº of Groups | Nº of Animals/Groups |
|---|---|---|---|---|---|---|
| Jurgonski, Juskiewicz, Zdunczyk, 2008 [35] | Poland | Wistar rats/NS | Male | 161 ± 8 | 3 | 8 |
| Jurgonski, Juskiewicz, Zdunczyk, 2013 [36] | Poland | Wistar rats/NS | Male | 548 ± 36 | 3 | 8 |
| Espley, et al., 2014 [31] | New Zealand | Swiss mice/6–7 wk | Male | 30 | 3 | 10 |
| Jurgonski, et al., 2014 [43] | Poland | White rabbits/34 days | Male | 631 ± 26 | 4 | 5 |
| Paivarinta, et al., 2016 [32] | Finland | C57BL/6J Apcmim mice/5 wk | Male and female | NS | 3 | 5–6 male and 4–6 female |
| Overall, et al., 2017 [26] | U.S.A. | C57BL/6J mice/6 wk | Male | 20–30 | 8 | 12 or 8 |
| Tong, et al., 2017 [17] | China | Kunming mice/NS | Male | 22 ± 5 | 5 | 10 |
| Fernández, et al., 2018 [37] | Spain | Fischer 344 rats/5wk | Male | 200–270 | 3 | 10 |
| Jamar, et al., 2018 [38] | Brazil | Wistar rats/90 days | Male | NS | 3 | 7 |
| Lee, et al., 2018 [39] | Georgia | Wistar rats/NS | Male | 200–220 | 3 | 8 |
| Paturi, et al., 2018 [40] | New Zealand | Sprague-Dawley rats/3 wk | Male | 256–265 | 8 | 16 |
| Silva-Maia, et al., 2018 [41] | Brazil | Wistar rats/3 wks | Male | 0–100 | 3 | 5 or 8 |
| Van Hul, et al., 2018 [33] | France | C57BL/6J mice/9 wk | Male | 25–30 | 4 | 14 |
| Chen, et al., 2019 [18] | China | Wistar rats/13 wks | Male | 403 ± 4 | 5 | 8 |
| Cremonini, et al., 2019 [27] | U.S.A. | C57BL/6J mice/NS | Male | 20–25 | 4 | 10 |
| Gu, et al., 2019 [28] | U.S.A. | C57BL/6J mice/4 wk | Male | NS | 2 | 12 or 14 |
| Li, et al., 2019 [19] | China | SD rats/4 and 12 months | Female | NS | 6 | 10 |
| Liu, et al., 2019 [29] | U.S.A. | C57BL/6J mice/4–5 wk | Male | 18–21 | 3 | 9 or 10 |
| Luo, et al., 2019 [23] | China | Sprague-Dawley rats/4 wk | Male | 100–120 | 6 | 8 |
| Peng, et al., 2019 [20] | China | C57BL/6J mice/5 wk | Male | 21–24 | 4 | 10 |
| Su, et al., 2019 [21] | China | db/db mice C57BL/6J/6 wk | Male | NS | 2 | 12 |
| Tian, et al., 2019 [22] | China | C57BL/6J mice/4 wk | Male | 15–18 | 6 | 11 |
| Zary-Sikorska, et al., 2019 [42] | Poland | Wistar rats/13 wk | Male | 146 ± 1.051 | 5 | 8 |
| Cao, et al., 2020 [30] | U.S.A. | C57BL/6J mice/3–18 months | Female | NS | 4 | 3 |
| Peng, et al., 2020 [24] | China | C57BL/6J mice/5 wk | Male | 20–24 | 2 | 10 |
| Rodríguez-Daza, et al., 2020 [34] | Canada | C57BL/6J mice/6 wk | Male | 20–25 | 6 | 12 |
| Wang, et al., 2020 [25] | China | C57BL/6J mice/6 wk | Male | 19–20 | 5 | 12 |
| Reference | Design (Intervention) | Control | Administration/Duration of Intervention (Weeks) | Method of Gut Microbiota Evaluation/Type of Sample | Anthocyanin Dosage (Total Anthocyanin) | Main Results (Intervention × Control) |
|---|---|---|---|---|---|---|
| Jurgonski, Juskiewicz, Zdunczyk, 2008 [35] | Chokeberry fruit extract (0.2%) + High fructose diet and streptozotocin | High fructose diet and streptozotocin | Oral (diet)/4 | NA | 80.9 mg/100 g diet | ↓ ileal pH; Mucosal disaccharidase activity: ↓ sucrase and maltase and ↑ lactase;  Total SCFA; cecum pH; α- and β- glucosidase, α- and β- galactosidase and β-glucuronidase on cecum. Total SCFA; cecum pH; α- and β- glucosidase, α- and β- galactosidase and β-glucuronidase on cecum. |
| Jurgonski, Juskiewicz, Zdunczyk, 2013 [36] | Kamchatka berry extract (2g/kg diet) + Diet with fructose replaced the corn starch | Diet with fructose replaced the corn starch | Oral (diet)/4 | NA | 65.4 mg/100 g diet | Mucosal disaccharidase activity: sucrase, maltase and lactase; cecum pH; ↑ α- and β- glucosidase on cecum; α- and β- galactosidase on cecum; ileal pH. |
| Espley, et al., 2014 [31] | Freeze-dried apple (20%) + Normal diet | Normal diet | Oral (diet)/3 | qPCR Colonic content | 397 µg/g diet * | ↑ Total bacteria; ↓ Lactobacillus spp.; Bifidobacterium spp.; Bacteroides-Prevotella-Porphyromonas group. |
| Jurgonski, et al., 2014 [43] | Blackcurrant pomace extract (1.5%) + HFD | HFD | Oral (diet)/4 | NA | 733.5 mg/100 g diet | ↓ Small intestine pH; Caecum pH; ↓ β-glucuronidase; α- and β- glucosidase, α- and β- galactosidase; Total SCFA cecal. |
| Paivarinta, et al., 2016 [32] | Bilberry extract (10%) + HFD | HFD | Oral (diet)/10 | PCR-DGGE Cecum content | 553.2 mg/100 g diet | ↑ Bacterial diversity in cecal contents. |
| Overall, et al., 2017 [26] | Blueberry powder (400 µg/g total anth.) + HFD | HFD | Oral (diet)/12 | qPCR Fecal sample | 40 mg/100 g diet | ↑ Abundance of Bacteroidete and Actinobacteria. |
| Tong, et al., 2017 [17] | Anthocyanin from red cabbage extract (100mg/kg BW) + CPT-11 (to induce intestinal mucositis) | CPT-11 (to induce intestinal mucositis) | Oral (gavage)/1 | NA | 100 mg/kg BW | ↑ Goblet cell mucus; Preservation of the villi height and conserved epithelial cell surface in the ileum and colon. |
| Fernández, et al., 2018 [37] | Functional sausage (20g with 0.11% anth.) + AOM treatment (to induce CRC tumor) | AOM treatment + Control sausage (20g) | Oral (diet)/20 | NGS Caecal feces | 22 mg/20 g sausage | ↓ Hyperplastic payer patches in the small intestine mucosa; ↓ level of Desulfovibrionaceae and Enterobacteriaceae and ↑ of Clostridiaceae; ↓ Bilophila wadsworthia. |
| Jamar, et al., 2018 [38] | Juçara powder (0.25%) + HFD | HFD | Oral (diet)/1 | qPCR Colon content | 1.65 mg/kg/day | ↓ mRNA of TLR-4 in the colon; mRNA ZO-1; ↑ DNA levels of Bifidobacterium spp. |
| Lee, et al., 2018 [39] | Blueberry powder (10%) + HFD | HFD | Oral (diet)/8 | NGS Caecal content | 213.4 mg/100 g diet | ↓ Bacteroidetes and Firmicutes abundance; ↑ Abundance of Proteobacteria and Fusobacteria; ↑ Bacilli and Lactobacillales; ↑ mRNA Muc2 ileal; ↑ ileal villus length and goblet cell number; ↑ serum acetate; Serum propionate and butyrate; ↓ serum LBS (to assess LPS concentration); mRNA antimicrobial peptide Defb2. |
| Paturi, et al., 2018 [40] | Blackcurrant extract (40g/kg) + Control diet | Control diet | Oral (diet)/6 | qPCR Caecal content | 1280 mg/100g diet | ↓ cecal acetic and butyric and ↑ of propionic acid; ↑ Bacteroides-Provotella-Porphyromonas group and Lactobacillus spp.; ↓ Bifidobacterium spp. and Clostridium perfringens; crypt depth and goblet cells in the colon. |
| Silva-Maia, et al., 2018 [41] | Aqueous extract of berry (Plinia jaboticaba) peel (50g/L) + Normal diet | Normal diet | Oral (water)/7 | Colonies expressed as CFU Colon content | 75 mg/L | ↑ Enterobacteriaceae and Bifidobacterium, and Lactobacillus; total SCFA. |
| Van Hul, et al., 2018 [33] | Grape pomace extract (8.2 g/kg diet) + HFD | HFD | Oral (diet)/8 | NGS Caecal content | 35.59 mg/100 g diet | ↑ Abundance of Bacteroidetes; ↓ Desulfovibrionaceae and Spreptoccaceae; ↑ Prevotellaceae and Erysipelotrichaceae; mRNA of ZO-1, intectin, occludin, claudin3, Muc2, Reg3ϒ; ↑ mRNA Lyz1; Total SCFA cecal. |
| Chen, et al., 2019 [18] | Purified cyanidin-3-O-glucoside (1000mg/kg) + 3-MCPD | 3-MCPD (to damage the intestinal mucosa) | Oral (diet)/8 | NGS Colonic content | 1000 mg/kg diet ** | ↓ Bacteroidetes levels and ↑ Proteobacteria and Actinobacteria; ↑ Villus height, and number of epithelial cells. |
| Cremonini, et al., 2019 [27] | Anthocyanin rich mix (40mg/kg) + HFD | HFD | Oral (diet)/14 | NGS Caecal content | 40 mg/kg BW | ↓ Intestinal permeability; ↓ Plasm endotoxin; ↓ ratio Firmicutes/Bacteroidetes; ↑ Romansia abundance; ↑ Protein expression of occludin, ZO-1 and claudin-1; ↑ Muc2 secretion. |
| Gu, et al., 2019 [28] | Black rasberry powder (10%) + Control diet | Control diet | Oral (diet)/6 | NGS Luminal content | 290 mg/100 g diet | ↓ Abundance of Firmicutes and ↑ of Bacteroidetes; ↓ Clostridium ↑ Barnessiella |
| Li, et al., 2019 [19] | Bilberry anthocyanin extract (20 mg/kg) + Old rats | Old rats | Oral (gavage)/10 | NGS Caecal content | 20 mg/kg BW | ↓ Abundance of Verrucomicrobia and Euryarchaeota; ↓ Ratio Firmicutes/Bacteroidetes; ↑ Species of Weissella confuse and Aspergillus oryzae; ↑ Lactobacillus and Bacteroides; ↑ Total SCFA in cecal content; ↓ β-glucosidade and α-galactosidade and α-glucosidase, α-galactosidade, and β-glucoronidase; ↓ serum LPS. |
| Liu, et al., 2019 [29] | Malvindin 3-Glucoside (24mg/kg diet) + DSS | DSS | Oral (diet)/50 days | NGS Colon content | 24 mg/kg diet *** | ↓ Abundance of R. gnavus and ↑ Clostridium and Bacteroides ovatus; ↑ Firmicutes/Bacteroidetes ratio; ↑ crypt dilation. |
| Luo, et al., 2019 [23] | Purified anthocyanin from L. ruthenicum (200 mg/kg BW) + HFD + vit. D3 | HFD + vit. D3 (to induce atheroscherosis) | Oral (gavage)/6 | NGS Cecal content | 105.5 mg/kg BW | ↓ Abundance of Firmicutes and ↑ Bacteroidetes; ↑ Bifidobacterium and Lactobacillus; ↑ Abundance of Lria, Akkermansia and Lachnospiraceae; Improvement of structure and villi of the small intestine; |
| Peng, et al., 2019 [20] | Purified anthocyanin from L. ruthenicum (200 mg/kg BW) + DSS | DSS | Oral (water)/7 days | NGS Feces samples | 200 mg/kg BW | ↑ mRNA of ZO-1, occludin, claudin-1; ↑ total SCFA in cecal content and feces; ↑ goblet cells ; ↑ abundance of Actinobacteria; Abundance of Firmicutes and Bacteroidete; Firmicutes/Bacteroidetes ratio. |
| Su, et al., 2019 [21] | Pelargonidin-3-O-glucoside (150 mg/kg BW) from raspberry + Diabetic db/db | Diabetc db/db | Oral (gavage)/8 | NGS Caecal content | 150 mg/kg BW **** | ↓ Abundance of Firmicutes and ↑ Bacteroidetes; ↓ serum LPS; ↑ Bacteroidetes/Firmicutes ratio; ↑ Total SCFA fecal; ↑ mRNA of occludin e ZO-1, Muc 2, and claudin; ↑ Pla2g2 and Lyz1 (antimicrobial peptides). |
| Tian, et al., 2019 [22] | L. ruthenicum dried (3%) + Normal diet | Normal diet | Oral (diet)/10 | NGS Fecal pellets | 104.2 mg/100 g diet | ↓ Abundance of Firmicutes; ↓ pH feces; ↓ Serum LPS; ↑ Serum and colon sIgA; ↑ Verrucomicrobia and Bacteroidetes; ↓ Proteobacteria and Deferribacteres; ↑ Total fecal SCFA; ↑ Ileal villus length and ratio of villus to crypt; ↑ mRNA of ZO-1, occludin, JAM-A and Muc2; Colon crypt length. |
| Zary-Sikorska, et al., 2019 [42] | Purple carrot root (dried) (10%) | Control (without carrot) | Oral (diet)/4 | NA | 12.9 mg/100 g diet | ↓ Cecal pH; ↑ α- and β-Glucosidase; α- and β-Galactosidade; β-glucuronidase; ↑ Total cecal SCFA. |
| Cao, et al., 2020 [30] | Blackcurrant extract (1%) + Old rats | Old rats | Oral (diet)/16 | NGS Feces samples | 17.41 mg/100 g diet | ↓ Firmicutes/Bacteroidetes ratio; ↓ Abundance of Verrucomicrobia, ↑ Bacteroidetes and Firmicutes and Proteobacteria. |
| Peng, et al., 2020 [24] | Anthocyanins from L. ruthenicum (200 mg/kg BW) | Control (without anth.) | Oral (diet)/12 | NGS Feces samples | 200 mg/kg BW | ↑ nº of intestinal villi, goblet cells and intestinal gland; ↑ mRNA of ZO-1, occludin, claudin and Muc1; ↑ total SCFA (cecal content and feces); ↑ Barnesiella, Alistipes, Eisenbergiella, Coprobacter and Odoribacter; pH in feces and cecal sIgA. |
| Rodríguez-Daza, et al., 2020 [34] | Blueberry extract (200 mg/kg BW) + High fat and high sucrose diet | High fat and high sucrose diet | Oral (gavage)/8 | NGS Feces samples | 1.68 mg/kg BW | ↑ Mucus layer thickness (colon); ↑ Adlercreutzia equolifaciens; Crypt depth and total goblet cells; Firmicutes/Bacteroidetes ratio; mRNA of ZO-1 and occludin. |
| Wang, et al., 2020 [25] | Black rice extract (0.48 g/kg diet) + High fat and cholesterol diet | High fat and cholesterol diet | Oral (diet)/12 | NGS Caecal content | 48 mg/100 g diet | ↓ Firmicutes/Bacteroidetes ratio; ↑ Abundance of Bifidobacterium and Lactobacillus; ↑ Cecal SCFA; ↑ Villus height (ileum and caecum); ↑ Goblet cell number per villus of the colon; ↑ mRNA of JAM-A, occludin and Muc-2. |
: no change; * cyanidin galactoside; ** cyanidin-3-O-glucoside; *** Malvindin 3-Glucoside; **** Pelargonidin-3-O-glucoside; Abbreviations: BW: Body weight; HFD: High fat diet; CPT-11: irinotecan; AOM: azoxymethane; DSS: dextan sodium sulfate; ZO-1: zonula occludentes–1; 3-MCPD: 3-Chloro-1,2-propanediol; SCFA: short chain fatty acids; JAM-A: junctional adhesion molecule-A; L. ruthenicum: Lycium ruthenicum; CRC: colorectal cancer; Pla2g2: phospholipase A2 group-II; Lyz1: Lysosome-1; LPS: lipopolysaccharides; anth.: anthocyanin; TLR-4: toll like receptor 4; sIgA: secretory Immunoglobulin A; mRNA: messenger ribonucleic acid; Muc: mucin; Defb2: beta-defensin 2; LBS: LPS-binding protein; qPCR: quantitative polymerase chain reaction; CFU: colony forming unit; NA: not applicable; NGS: next generation sequencing; DGGE: denaturation gradient gel electrophoresis.Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verediano, T.A.; Stampini Duarte Martino, H.; Dias Paes, M.C.; Tako, E. Effects of Anthocyanin on Intestinal Health: A Systematic Review. Nutrients 2021, 13, 1331. https://doi.org/10.3390/nu13041331
Verediano TA, Stampini Duarte Martino H, Dias Paes MC, Tako E. Effects of Anthocyanin on Intestinal Health: A Systematic Review. Nutrients. 2021; 13(4):1331. https://doi.org/10.3390/nu13041331
Chicago/Turabian StyleVerediano, Thaísa Agrizzi, Hércia Stampini Duarte Martino, Maria Cristina Dias Paes, and Elad Tako. 2021. "Effects of Anthocyanin on Intestinal Health: A Systematic Review" Nutrients 13, no. 4: 1331. https://doi.org/10.3390/nu13041331
APA StyleVerediano, T. A., Stampini Duarte Martino, H., Dias Paes, M. C., & Tako, E. (2021). Effects of Anthocyanin on Intestinal Health: A Systematic Review. Nutrients, 13(4), 1331. https://doi.org/10.3390/nu13041331