
Pancreatic Cancer and Gut Microbiome-Related Aspects: A Comprehensive Review and Dietary Recommendations

 ,
,  , ,
, ,
Abstract
1. Introduction
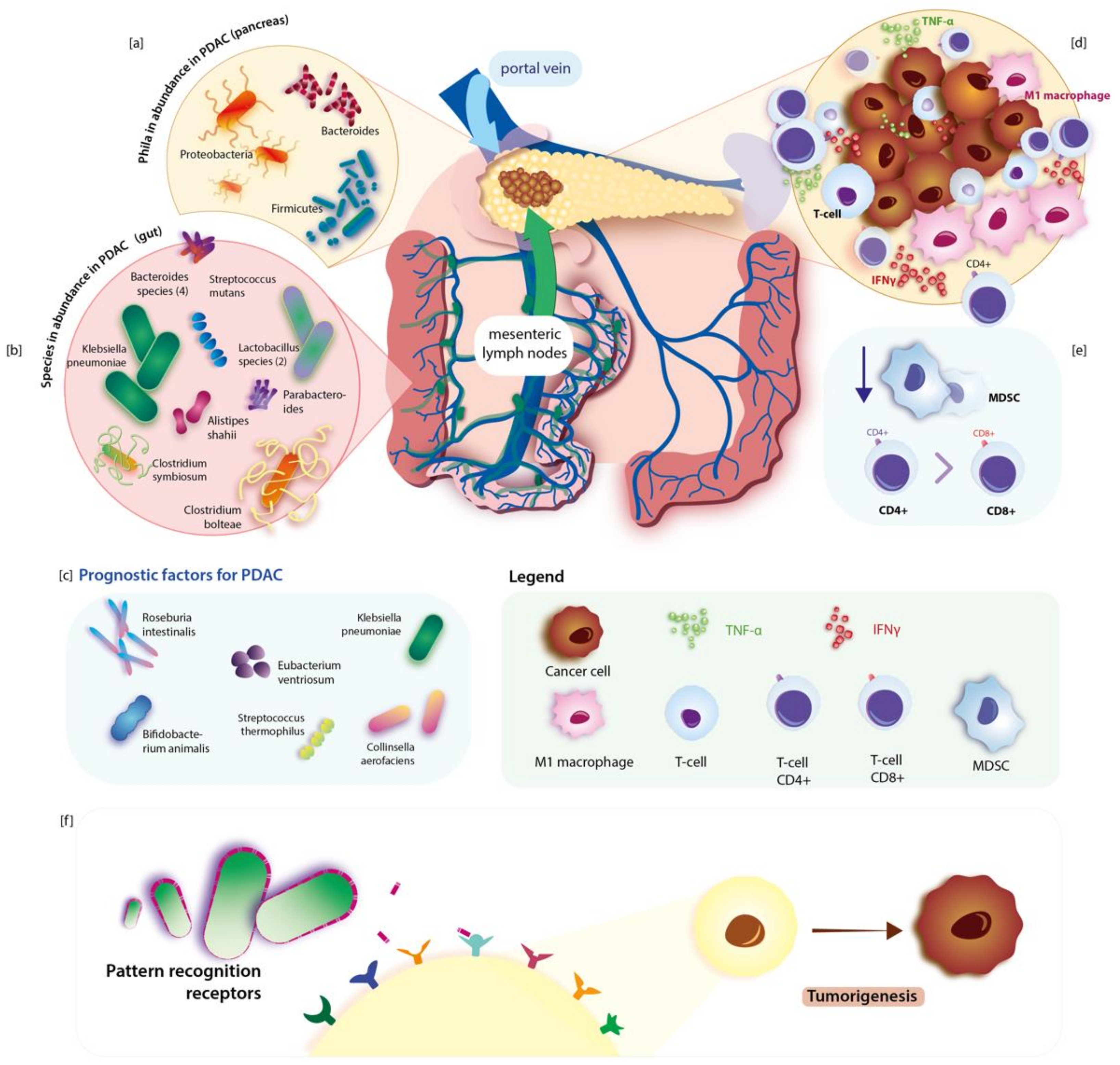
2. The Link between Gut Microbiota and Pancreatic Carcinogenesis. Intra-Tumour Microenvironment
2.1. Bacteria
2.2. Fungi
2.3. Viruses
3. Therapeutic Modulation of Gut Microbiota in Pancreatic Cancer
3.1. Prebiotics
3.2. Probiotics and Next-Generation Probiotics
3.3. Synbiotics
3.4. Postbiotics
3.5. Fecal Microbiota Transplantation
3.6. Short Summary
4. The Registration of Studies Regarding Pancreatic Cancer and Gut Microbiota-Related Aspects in the System
5. Clinical Nutrition in Pancreatic Cancer
5.1. Nutritional Assessment and Support
5.2. Nutrition
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Cancer Facts & Figures. 2019. Available online: https://www.cancer.org/content/dam/cancer-org/research/cancer-facts-and-statistics/annual-cancer-facts-and-figures/2019/cancer-facts-and-figures-2019.pdf?fbclid=IwAR24wyni_jp6BAwwQ0Xj7-1E2aSFSjHPq4BrG-KSMcF2Vuee5o-wJMuaalc (accessed on 15 May 2020).
- Kaźmierczak-Siedlecka, K.; Dvořák, A.; Folwarski, M.; Daca, A.; Przewłócka, K.; Makarewicz, W. Fungal Gut Microbiota Dysbiosis and Its Role in Colorectal, Oral, and Pancreatic Carcinogenesis. Cancers 2020, 12, 1326. [Google Scholar] [CrossRef] [PubMed]
- Mendez, R.; Kesh, K.; Arora, N.; Di Martino, L.; McAllister, F.; Merchant, N.; Banerjee, S.; Banerjee, S. Microbial Dysbiosis and Polyamine Metabolism as Predictive Markers for Early Detection of Pancreatic Cancer. Carcinogenesis 2020, 41, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Karpiński, T.M. The Microbiota and Pancreatic Cancer. Gastroenterol. Clin. N. Am. 2019, 48, 447–464. [Google Scholar]
- Wei, M.-Y.; Shi, S.; Liang, C.; Meng, Q.C.; Hua, J.; Zhang, Y.-Y.; Liu, J.; Bo, Z.; Xu, J.; Yu, X.J. The Microbiota and Microbiome in Pancreatic Cancer: More Influential than Expected. Mol. Cancer 2019, 18, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Sendid, B.; Hoarau, G.; Colombel, J.F.; Poulain, D.; Ghannoum, M.A. Mycobiota in gastrointestinal diseases. Nat. Rev. Gastroenterol. Hepatol. 2015, 2, 77–87. [Google Scholar] [CrossRef]
- Hoffmann, C.; Dollive, S.; Grunberg, S.; Chen, J.; Li, H.; Wu, G.D.; Lewis, J.D.; Bushman, F.D. Archaea and Fungi of the Human Gut Microbiome: Correlations with Diet and Bacterial Residents. PLoS ONE 2013, 6, e66019. [Google Scholar] [CrossRef] [PubMed]
- Aykut, B.; Pushalkar, S.; Chen, R.; Li, Q.; Abengozar, R.; Kim, J.I.; Shadaloey, S.A.; Wu, D.; Preiss, P.; Verma, N.; et al. The fungal mycobiome promotes pancreatic oncogenesis via activation of MBL. Nature 2019, 7777, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Luan, C.; Xie, L.; Yang, X.; Miao, H.; Lv, N.; Zhang, R.; Xiao, X.; Hu, Y.F.; Liu, Y.L.; Wu, N.; et al. Dysbiosis of Fungal Microbiota in the Intestinal Mucosa of Patients with Colorectal Adenomas. Sci. Rep. 2015, 5, 7980. [Google Scholar] [CrossRef]
- Zambirinis, C.; Levie, E.; Nguy, S.; Avanzi, A.; Barilla, R.; Xu, Y.; Seifert, L.; Daley, D.; Greco, S.H.; Deutsch, M.; et al. TLR9 Ligation in Pancreatic Stellate Cells Promotes Tumorigenesis. J. Exp. Med. 2015, 212, 2077–2094. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, J.; Nguyen, A.H.; Rehman, A.; Ochi, A.; Jamal, M.; Graffeo, C.S.; Henning, J.R.; Zambirinis, C.; Fallon, N.C.; Barilla, R.; et al. Dendritic Cell Populations with Different Concentrations of Lipid Regulate Tolerance and Immunity in Mouse and Human Liver. Gastroenterology 2012, 143, 1061–1072. [Google Scholar] [CrossRef]
- Ochi, A.; Nguyen, A.H.; Bedrosian, A.S.; Mushlin, H.M.; Zarbakhsh, S.; Barilla, R.; Zambirinis, C.P.; Fallon, N.C.; Rehman, A.; Pylayeva-Gupta, Y.; et al. MyD88 Inhibition Amplifies Dendritic Cell Capacity to Promote Pancreatic Carcinogenesis via Th2 Cells. J. Exp. Med. 2012, 209, 1671–1687. [Google Scholar] [CrossRef] [PubMed]
- Zambirinis, C.P.; Ochi, A.; Barilla, R.; Greco, S.; Deutsch, M.; Miller, G. Induction of TRIF- or MYD88-Dependent Pathways Perturbs Cell Cycle Regulation in Pancreatic Cancer. Cell Cycle 2013, 12, 1153–1154. [Google Scholar]
- Seifert, L.; Werba, G.; Tiwari, S.; Ngoc Giao Ly, N.; Alothman, S.; Alqunaibit, D.; Avanzi, A.; Barilla, R.; Daley, D.; Greco, S.H.; et al. The Necrosome Promotes Pancreatic Oncogenesis via CXCL1 and Mincle-Induced Immune Suppression. Nature 2016, 532, 245–249. [Google Scholar] [CrossRef]
- Pushalkar, S.; Hundeyin, M.; Daley, D.; Zambirinis, C.P.; Kurz, E.; Mishra, A.; Mohan, N.; Aykut, B.; Usyk, M.; Torres, L.E.; et al. The Pancreatic Cancer Microbiome Promotes Oncogenesis by Induction of Innate and Adaptive Immune Suppression. Cancer Discov. 2018, 8, 403–416. [Google Scholar] [CrossRef]
- Fan, X.; Alekseyenko, A.V.; Wu, J.; Peters, B.A.; Jacobs, E.J.; Gapstur, S.M.; Purdue, M.P.; Abnet, C.C.; Stolzenberg-Solomon, R.; Miller, G.; et al. Human Oral Microbiome and Prospective Risk for Pancreatic Cancer: A Population-Based Nested Case-Control Study. Gut 2018, 67, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.M.; Jobin, C. Microbiota in Pancreatic Health and Disease: The next Frontier in Microbiome Research. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Farrell, J.J.; Zhang, L.; Zhou, H.; Chia, D.; Elashoff, D.; Akin, D.; Paster, B.J.; Joshipura, K.; Wong, D.T.W. Variations of oral microbiota are associated with pancreatic diseases including pancreatic cancer. Gut 2012, 61, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Del Castillo, E.; Meier, R.; Chung, M.; Koestler, D.C.; Chen, T.; Paster, B.J.; Charpentier, K.P.; Kelsey, K.T.; Izard, J.; Michaud, D.S. The Microbiomes of Pancreatic and Duodenum Tissue Overlap and Are Highly Subject Specific but Differ between Pancreatic Cancer and Noncancer Subjects. Cancer Epidemiol. Biomark. Prev. 2019, 28, 370–383. [Google Scholar] [CrossRef]
- Matsukawa, H.; Iida, N.; Kitamura, K.; Terashima, T.; Seishima, J.; Makino, I.; Kannon, T.; Hosomichi, K.; Yamashita, T.; Sakai, Y.; et al. Dysbiotic Gut Microbiota in Pancreatic Cancer Patients Form Correlation Networks with the Oral Microbiota and Prognostic Factors. Am. J. Cancer Res. 2021, 11, 3163. [Google Scholar] [PubMed]
- Zhou, W.; Zhang, D.; Li, Z.; Jiang, H.; Li, J.; Ren, R.; Gao, X.; Li, J.; Wang, X.; Wang, W.; et al. The Fecal Microbiota of Patients with Pancreatic Ductal Adenocarcinoma and Autoimmune Pancreatitis Characterized by Metagenomic Sequencing. J. Transl. Med. 2021, 19, 215. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, E.; Zhang, Y.; Zhang, L.; Montiel, M.; Zoltan, M.; Dong, W.; Quesada, P.; Sahin, I.; Chandra, V.; Lucas, A.S.; et al. Tumor Microbiome Diversity and Composition Influence Pancreatic Cancer Outcomes. Cell 2019, 178, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Sethi, V.; Kurtom, S.; Tarique, M.; Lavania, S.; Malchiodi, Z.; Hellmund, L.; Zhang, L.; Sharma, U.; Giri, B.; Garg, B.; et al. Gut Microbiota Promotes Tumor Growth in Mice by Modulating Immune Response. Gastroenterology 2018, 155, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Cao, Z.; Qiu, J.; Liu, Y.; Zheng, L.; Zhang, T. Novel Discoveries Targeting Pathogenic Gut Microbes and New Therapies in Pancreatic Cancer: Does Pathogenic E. coli Infection Cause Pancreatic Cancer Progression Modulated by TUBB/Rho/ROCK Signaling Pathway? A Bioinformatic Analysis. Biomed. Res. Int. 2020, 2020, 2340124. [Google Scholar] [CrossRef] [PubMed]
- Shrader, H.R.; Miller, A.M.; Tomanek-Chalkley, A.; McCarthy, A.; Coleman, K.L.; Ear, P.H.; Mangalam, A.K.; Salem, A.K.; Chan, C.H.F. Effect of Bacterial Contamination in Bile on Pancreatic Cancer Cell Survival. Surgery 2021, 169, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Di Carlo, P.; Serra, N.; D’Arpa, F.; Agrusa, A.; Gulotta, G.; Fasciana, T.; Rodolico, V.; Giammanco, A.; Sergi, C. The Microbiota of the Bilio-Pancreatic System: A Cohort, STROBE-Compliant Study. Infect. Drug Resist. 2019, 12, 1513–1527. [Google Scholar] [CrossRef]
- Geller, L.T.; Barzily-Rokni, M.; Danino, T.; Jonas, O.H.; Shental, N.; Nejman, D.; Gavert, N.; Zwang, Y.; Cooper, Z.A.; Shee, K.; et al. Potential role of intratumor bacteria in mediating tumor resistance to the chemotherapeutic drug gemcitabine. Science 2017, 357, 1156–1160. [Google Scholar] [CrossRef]
- Panebianco, C.; Adamberg, K.; Jaagura, M.; Copetti, M.; Fontana, A.; Adamberg, S.; Kolk, K.; Vilu, R.; Andriulli, A.; Pazienza, V. Influence of Gemcitabine Chemotherapy on the Microbiota of Pancreatic Cancer Xenografted Mice. Cancer Chemother. Pharmacol. 2018, 81, 773–782. [Google Scholar] [CrossRef]
- Iida, N.; Dzutsev, A.; Stewart, C.A.; Smith, L.; Bouladoux, N.; Weingarten, R.A.; Molina, D.A.; Salcedo, R.; Back, T.; Cramer, S.; et al. Commensal Bacteria Control Cancer Response to Therapy by Modulating the Tumor Microenvironment. Science 2013, 342, 967–970. [Google Scholar] [CrossRef]
- Oar, A.; Lee, M.; Le, H.; Wilson, K.; Aiken, C.; Chantrill, L.; Simes, J.; Nguyen, N.; Barbour, A.; Samra, J.; et al. AGITG MASTERPLAN: A Randomised Phase II Study of Modified FOLFIRINOX Alone or in Combination with Stereotactic Body Radiotherapy for Patients with High-Risk and Locally Advanced Pancreatic Cancer. BMC Cancer 2021, 21, 936. [Google Scholar] [CrossRef] [PubMed]
- Dong, T.S.; Chang, H.-H.; Hauer, M.; Lagishetty, V.; Katzka, W.; Rozengurt, E.; Jacobs, J.P.; Eibl, G. Metformin alters the duodenal microbiome and decreases the incidence of pancreatic ductal adenocarcinoma promoted by diet-induced obesity. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G763–G772. [Google Scholar] [CrossRef] [PubMed]
- Maletzki, C.; Linnebacher, M.; Kreikemeyer, B.; Emmrich, J. Pancreatic Cancer Regression by Intratumoural Injection of Live Streptococcus Pyogenes in a Syngeneic Mouse Model. Gut 2008, 57, 483–491. [Google Scholar] [CrossRef]
- Kesh, K.; Mendez, R.; Abdelrahman, L.; Banerjee, S.; Banerjee, S. 2 Diabetes Induced Microbiome Dysbiosis Is Associated with Therapy Resistance in Pancreatic Adenocarcinoma. Microb. Cell Fact. 2020, 19, 75. [Google Scholar] [CrossRef] [PubMed]
- Elaskandrany, M.; Patel, R.; Patel, M.; Miller, G.; Saxena, D.; Saxena, A. Fungi, Host Immune Response, and Tumorigenesis. Am. J. Physiol. Gastrointest. Liver Physiol. 2021, 321, G213–G222. [Google Scholar] [CrossRef] [PubMed]
- Dambuza, I.M.; Brown, G.D. Fungi Accelerate Pancreatic Cancer. Nature 2019, 574, 184–185. [Google Scholar] [CrossRef] [PubMed]
- Conche, C.; Greten, F.R. Fungi Enter the Stage of Colon Carcinogenesis. Immunity 2018, 49, 384–386. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, B.; Delgado, S.; Blanco-Míguez, A.; Lourenço, A.; Gueimonde, M.; Margolles, A. Probiotics, Gut Microbiota, and Their Influence on Host Health and Disease. Mol. Nutr. Food Res. 2017, 61, 1600240. [Google Scholar] [CrossRef]
- Arafa, A.; Eshak, E.S.; Rahman, T.A.A.; Anwar, M.M. Hepatitis C Virus Infection and Risk of Pancreatic Cancer: A Meta-Analysis. Cancer Epidemiol. 2020, 65, 101691. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wu, B.; Yang, L.-B.; Yin, G.-C.; Liu, J.-Y. Chronic Hepatitis B Virus Infection and Risk of Pancreatic Cancer: A Meta-Analysis. Asian Pac. J. Cancer Prev. 2013, 14, 275–279. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liu, X.; Zhang, Z.-H.; Jiang, F. Hepatitis B Virus Infection Increases the Risk of Pancreatic Cancer: A Meta-Analysis. Scand. J. Gastroenterol. 2021, 56, 252–258. [Google Scholar] [CrossRef]
- Jin, Y.; Gao, H.; Chen, H.; Chen, H.; Wang, J.; Chen, M.; Li, G.; Wang, L.; Gu, J.; Tu, H. Identification and Impact of Hepatitis B Virus DNA and Antigens in Pancreatic Cancer Tissues and Adjacent Non-Cancerous Tissues. Cancer Lett. 2013, 335, 447–454. [Google Scholar] [CrossRef]
- Chen, Y.; Bai, X.; Zhang, Q.; Wen, L.; Su, W.; Fu, Q.; Sun, X.; Lou, Y.; Yang, J.; Zhang, J.; et al. The Hepatitis B Virus X Protein Promotes Pancreatic Cancer through Modulation of the PI3K/AKT Signaling Pathway. Cancer Lett. 2016, 380, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Walter, R.J.; Attar, B.M.; Rafiq, A.; Tejaswi, S.; Delimata, M. Newcastle Disease Virus Lasota Strain Kills Human Pancreatic Cancer Cells in Vitro with High Selectivity. J. Pancreas 2012, 13, 45–53. [Google Scholar] [CrossRef]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Jalil Masoumi, S.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Trivieri, N.; Panebianco, C.; Villani, A.; Pracella, R.; Latiano, T.P.; Perri, F.; Binda, E.; Pazienza, V. High Levels of Prebiotic Resistant Starch in Diet Modulate a Specific Pattern of miRNAs Expression Profile Associated to a Better Overall Survival in Pancreatic Cancer. Biomolecules 2021, 11, 26. [Google Scholar] [CrossRef] [PubMed]
- Panebianco, C.; Villani, A.; Pazienza, V. High Levels of Prebiotic Resistant Starch in Diet Modulate Gene Expression and Metabolomic Profile in Pancreatic Cancer Xenograft Mice. Nutrients 2019, 11, 709. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Ren, W.; Huang, X.; Deng, J.; Li, T.; Yin, Y. Potential Mechanisms Connecting Purine Metabolism and Cancer Therapy. Front. Immunol. 2018, 9, 1697. [Google Scholar] [CrossRef] [PubMed]
- Hotel, A.; Cordoba, A. Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria. Prevention 2001, 5, 1–34. [Google Scholar]
- Folwarski, M.; Dobosz, M.; Małgorzewicz, S.; Skonieczna-Żydecka, K.; Kaźmierczak-Siedlecka, K. Effects of Lactobacillus Rhamnosus Gg on Early Postoperative Outcome after Pylorus-Preserving Pancreatoduodenectomy: A Randomized Trial. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 397–405. [Google Scholar] [PubMed]
- Nomura, T.; Tsuchiya, Y.; Nashimoto, A.; Yabusaki, H.; Takii, Y.; Nakagawa, S.; Sato, N.; Kanbayashi, C.; Tanaka, O. Probiotics reduce infectious complications after pancreaticoduodenectomy. Hepatogastroenterology 2007, 54, 661–663. [Google Scholar] [PubMed]
- Kita, A.; Fujiya, M.; Konishi, H.; Tanaka, H.; Kashima, S.; Iwama, T.; Ijiri, M.; Murakami, Y.; Takauji, S.; Goto, T.; et al. Probiotic-Derived Ferrichrome Inhibits the Growth of Refractory Pancreatic Cancer Cells. Int. J. Oncol. 2020, 57, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Ijiri, M.; Fujiya, M.; Konishi, H.; Tanaka, H.; Ueno, N.; Kashima, S.; Moriichi, K.; Sasajima, J.; Ikuta, K.; Okumura, T. Ferrichrome Identified from Lactobacillus Casei ATCC334 Induces Apoptosis through Its Iron-Binding Site in Gastric Cancer Cells. Tumor Biol. 2017, 39, 1010428317711311. [Google Scholar] [CrossRef]
- Konishi, H.; Fujiya, M.; Tanaka, H.; Ueno, N.; Moriichi, K.; Sasajima, J.; Ikuta, H.K.M.F.H.T.N.U.K.M.J.S.K.; Akutsu, H.; Tanabe, H.; Kohgo, H.T.Y. Probiotic-Derived Ferrichrome Inhibits Colon Cancer Progression via JNK-Mediated Apoptosis. Nat. Commun. 2016, 7, 12365. [Google Scholar] [CrossRef]
- Satokari, R. Modulation of Gut Microbiota for Health by Current and Next-Generation Probiotics. Nutrients 2019, 11, 1921. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-J.; Lin, T.-L.; Tsai, Y.-L.; Wu, T.-R.; Lai, W.-F.; Lu, C.-C.; Lai, H.-C. Next generation probiotics in disease amelioration. J. Food Drug Anal. 2019, 27, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Liu, Z.; Zhu, C.; Mou, H.; Kong, G. Nondigestible carbohydrates, butyrate, and butyrate-producing bacteria. Crit. Rev. Food Sci. Nutr. 2019, 59 (Suppl. 1), S130–S152. [Google Scholar] [CrossRef] [PubMed]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; De Vos, W.M. Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 1469–1476. [Google Scholar] [CrossRef] [PubMed]
- Jayachandran, M.; Chung, S.S.M.; Xu, B. A critical review of the relationship between dietary components, the gut microbe Akkermansia muciniphila, and human health. Crit. Rev. Food Sci. Nutr. 2020, 60, 2265–2276. [Google Scholar] [CrossRef]
- Liu, M.N.; Zhang, L.; Dong, X.Y.; Liu, M.; Cheng, G.; Zhang, X.L.; He, F.; Wang, G.Q. Effect of Akkermansia muciniphila on the Proliferation, Apoptosis and Insulin Secretion of Rat Islet Cell Tumor Cells. Sichuan Da Xue Xue Bao Yi Xue Ban 2020, 51, 13–17. [Google Scholar] [PubMed]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Oláh, A.; Belágyi, T.; Pótó, L.; Romics, L., Jr.; Bengmark, S. Synbiotic control of inflammation and infection in severe acute pancreatitis: A prospective, randomized, double blind study. Hepatogastroenterology 2007, 54, 590–594. [Google Scholar]
- Derwa, Y.; Gracie, D.J.; Hamlin, P.J.; Ford, A.C. Systematic Review with Meta-Analysis: The Efficacy of Probiotics in Inflammatory Bowel Disease. Aliment. Pharmacol. Ther. 2017, 46, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Szajewska, H.; Kołodziej, M. Systematic Review with Meta-Analysis: Saccharomyces Boulardii in the Prevention of Antibiotic-Associated Diarrhoea. Aliment. Pharmacol. Ther. 2015, 42, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Kothari, D.; Patel, S.; Kim, S.K. Probiotic Supplements Might Not Be Universally-Effective and Safe: A Review. Biomed. Pharmacother. 2019, 111, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Fong, W.; Li, Q.; Yu, J. Gut Microbiota Modulation: A Novel Strategy for Prevention and Treatment of Colorectal Cancer. Oncogene 2020, 39, 4925–4943. [Google Scholar] [CrossRef] [PubMed]
- Żółkiewicz, J.; Marzec, A.; Ruszczyński, M.; Feleszko, W. Postbiotics—A Step beyond Pre-and Probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef] [PubMed]
- Tsilingiri, K.; Rescigno, M. Postbiotics: What Else? Benef. Microbes 2013, 4, 101–107. [Google Scholar] [CrossRef]
- Wang, L.; Cao, H.; Liu, L.; Wang, B.; Walker, W.; Acra, S.A.; Yan, F. Activation of Epidermal Growth Factor Receptor Mediates Mucin Production Stimulated by P40, a Lactobacillus Rhamnosus GG-Derived Protein. J. Biol. Chem. 2014, 289, 20234–20244. [Google Scholar] [CrossRef]
- Gao, J.; Li, Y.; Wan, Y.; Hu, T.; Liu, L.; Yang, S.; Gong, Z.; Zeng, Q.; Wei, Y.; Yang, W.; et al. A Novel Postbiotic from Lactobacillus Rhamnosus GG with a Beneficial Effect on Intestinal Barrier Function. Front. Microbiol. 2019, 10, 477. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Muñoz-Quezada, S.; Gomez-Llorente, C.; Matencio, E.; Bernal, M.J.; Romero, F.; Gil, A. Cell-Free Culture Supernatant of Bifidobacterium Breve CNCM I-4035 Decreases Pro-Inflammatory Cytokines in Human Dendritic Cells Challenged with Salmonella Typhi through TLR Activation. PLoS ONE 2013, 8, e59370. [Google Scholar] [CrossRef]
- De Marco, S.; Sichetti, M.; Muradyan, D.; Piccioni, M.; Traina, G.; Pagiotti, R.; Pietrella, D. Probiotic Cell-Free Supernatants Exhibited Anti-Inflammatory and Antioxidant Activity on Human Gut Epithelial Cells and Macrophages Stimulated with LPS. Evid.-Based Complement Altern. Med. 2018, 2018, 1756308. [Google Scholar] [CrossRef]
- Zhang, Z.; Mocanu, V.; Cai, C.; Dang, J.; Slater, L.; Deehan, E.C.; Walter, J.; Madsen, K.L. Impact of Fecal Microbiota Transplantation on Obesity and Metabolic Syndrome—A Systematic Review. Nutrients 2019, 11, 2291. [Google Scholar] [CrossRef] [PubMed]
- Szczyrek, M.; Bitkowska, P.; Chunowski, P.; Czuchryta, P.; Krawczyk, P.; Milanowski, J. Diet, Microbiome, and Cancer Immunotherapy—A Comprehensive Review. Nutrients 2021, 13, 2217. [Google Scholar] [CrossRef] [PubMed]
- Heo, G.; Lee, Y.; Im, E. Interplay between the Gut Microbiota and Inflammatory Mediators in the Development of Colorectal Cancer. Cancers 2021, 13, 734. [Google Scholar] [CrossRef] [PubMed]
- Kaźmierczak-Siedlecka, K.; Daca, A.; Fic, M.; van de Wetering, T.; Folwarski, M.; Makarewicz, W. Therapeutic Methods of Gut Microbiota Modification in Colorectal Cancer Management—Fecal Microbiota Transplantation, Prebiotics, Probiotics, and Synbiotics. Gut Microbes 2020, 11, 1518–1530. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, C.V.; de Camargo, M.R.; Russo, E.; Amedei, A. Role of Diet and Gut Microbiota on Colorectal Cancer Immunomodulation. World J. Gastroenterol. 2019, 25, 151–162. [Google Scholar] [CrossRef]
- Moffat, G.T.; Epstein, A.S.; O’Reilly, E.M. Pancreatic Cancer—A Disease in Need: Optimizing and Integrating Supportive Care. Cancer 2019, 125, 3927–3935. [Google Scholar] [CrossRef]
- Vanbutsele, G.; Pardon, K.; Van Belle, S.; Surmont, V.; De Laat, M.; Colman, R.; Eecloo, K.; Cocquyt, V.; Geboes, K.; Deliens, L. Effect of Early and Systematic Integration of Palliative Care in Patients with Advanced Cancer: A Randomised Controlled Trial. Lancet Oncol. 2018, 19, 394–404. [Google Scholar] [CrossRef]
- Bakitas, M.A.; Tosteson, T.D.; Li, Z.; Lyons, K.D.; Hull, J.G.; Li, Z.; Dionne-Odom, J.N.; Frost, J.; Dragnev, K.H.; Hegel, M.T.; et al. Early Versus Delayed Initiation of Concurrent Palliative Oncology Care: Patient Outcomes in the ENABLE III Randomized Controlled Trial. J. Clin. Oncol. 2015, 33, 1438–1445. [Google Scholar] [CrossRef]
- Bachmann, J.; Büchler, M.W.; Friess, H.; Martignoni, M.E. Cachexia in Patients with Chronic Pancreatitis and Pancreatic Cancer: Impact on Survival and Outcome. Nutr. Cancer 2013, 65, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Mueller, T.C.; Burmeister, M.A.; Bachmann, J.; Martignoni, M.E. Cachexia and Pancreatic Cancer: Are There Treatment Options? World J. Gastroenterol. 2014, 20, 9361–9373. [Google Scholar]
- Basile, D.; Corvaja, C.; Caccialanza, R.; Aprile, G. Sarcopenia: Looking to Muscle Mass to Better Manage Pancreatic Cancer Patients. Curr. Opin. Support Palliat Care 2019, 13, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Gilliland, T.M.; Villafane-Ferriol, N.; Shah, K.P.; Shah, R.M.; Tran Cao, H.S.; Massarweh, N.N.; Silberfein, E.J.; Choi, E.A.; Hsu, C.; McElhany, A.L.; et al. Nutritional and Metabolic Derangements in Pancreatic Cancer and Pancreatic Resection. Nutrients 2017, 9, 243. [Google Scholar] [CrossRef] [PubMed]
- Witvliet-van Nierop, J.E.; Lochtenberg-Potjes, C.M.; Wierdsma, N.J.; Scheffer, H.J.; Kazemier, G.; Ottens-Oussoren, K.; Meijerink, M.R.; de van der Schueren, M.A.E. Assessment of Nutritional Status, Digestion and Absorption, and Quality of Life in Patients with Locally Advanced Pancreatic Cancer. Gastroenterol. Res. Pract. 2017, 2017, 6193765. [Google Scholar] [CrossRef] [PubMed]
- Ozola Zalite, I.; Zykus, R.; Francisco Gonzalez, M.; Saygili, F.; Pukitis, A.; Gaujoux, S.; Charnley, R.M.; Lyadov, V. Influence of Cachexia and Sarcopenia on Survival in Pancreatic Ductal Adenocarcinoma: A Systematic Review. Pancreatology 2015, 15, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Hébuterne, X.; Lemarié, E.; Michallet, M.; de Montreuil, C.B.; Schneider, S.M.; Goldwasser, F. Prevalence of Malnutrition and Current Use of Nutrition Support in Patients with Cancer. JPEN J. Parenter. Enteral Nutr. 2014, 38, 196–204. [Google Scholar] [CrossRef]
- Muscaritoli, M.; Lucia, S.; Farcomeni, A.; Lorusso, V.; Saracino, V.; Barone, C.; Plastino, F.; Gori, S.; Magarotto, R.; Carteni, G.; et al. Prevalence of Malnutrition in Patients at First Medical Oncology Visit: The PreMiO Study. Oncotarget 2017, 8, 79884–79896. [Google Scholar] [CrossRef] [PubMed]
- Álvaro Sanz, E.; Garrido Siles, M.; Rey Fernández, L.; Villatoro Roldán, R.; Rueda Domínguez, A.; Abilés, J. Nutritional Risk and Malnutrition Rates at Diagnosis of Cancer in Patients Treated in Outpatient Settings: Early Intervention Protocol. Nutrition 2019, 57, 148–153. [Google Scholar] [CrossRef]
- Caccialanza, R.; Pedrazzoli, P.; Cereda, E.; Gavazzi, C.; Pinto, C.; Paccagnella, A.; Beretta, G.D.; Nardi, M.; Laviano, A.; Zagonel, V. Nutritional Support in Cancer Patients: A Position Paper from the Italian Society of Medical Oncology (AIOM) and the Italian Society of Artificial Nutrition and Metabolism (SINPE). J. Cancer 2016, 7, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Caccialanza, R.; Lobascio, F.; Brugnatelli, S.; Pedrazzoli, P. Nutritional Support in Pancreatic Cancer. Cancer 2020, 126, 1810–1811. [Google Scholar] [CrossRef] [PubMed]
- Deftereos, I.; Yeung, J.M.-C.; Arslan, J.; Carter, V.M.; Isenring, E.; Kiss, N.; On Behalf of The Nourish Point Prevalence Study Group. Preoperative Nutrition Intervention in Patients Undergoing Resection for Upper Gastrointestinal Cancer: Results from the Multi-Centre NOURISH Point Prevalence Study. Nutrients 2021, 13, 3205. [Google Scholar] [CrossRef]
- Bozzetti, F.; Arends, J.; Lundholm, K.; Micklewright, A.; Zurcher, G.; Muscaritoli, M. ESPEN ESPEN Guidelines on Parenteral Nutrition: Non-Surgical Oncology. Clin. Nutr. 2009, 28, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Arends, J.; Bodoky, G.; Bozzetti, F.; Fearon, K.; Muscaritoli, M.; Selga, G.; van Bokhorst-de van der Schueren, M.A.E.; von Meyenfeldt, M.; DGEM (German Society for Nutritional Medicine); Zürcher, G.; et al. ESPEN Guidelines on Enteral Nutrition: Non-Surgical Oncology. Clin. Nutr. 2006, 25, 245–259. [Google Scholar] [CrossRef] [PubMed]
- Weimann, A.; Braga, M.; Harsanyi, L.; Laviano, A.; Ljungqvist, O.; Soeters, P.; DGEM (German Society for Nutritional Medicine); Jauch, K.W.; Kemen, M.; Hiesmayr, J.M.; et al. ESPEN Guidelines on Enteral Nutrition: Surgery Including Organ Transplantation. Clin. Nutr. 2006, 25, 224–244. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Lee, S.M.; Jeung, H.C.; Lee, I.J.; Park, J.S.; Song, M.; Lee, D.K.; Lee, S.-M. The Effect of Nutrition Intervention with Oral Nutritional Supplements on Pancreatic and Bile Duct Cancer Patients Undergoing Chemotherapy. Nutrients 2019, 11, 1145. [Google Scholar] [CrossRef] [PubMed]
- Bozzetti, F. Nutritional Support of the Oncology Patient. Crit. Rev. Oncol. Hematol. 2013, 87, 172–200. [Google Scholar] [CrossRef] [PubMed]
- August, D.A.; Huhmann, M.B. American Society for Parenteral and Enteral Nutrition (A.S.P.E.N.) Board of Directors A.S.P.E.N. Clinical Guidelines: Nutrition Support Therapy during Adult Anticancer Treatment and in Hematopoietic Cell Transplantation. JPEN J. Parenter. Enteral Nutr. 2009, 33, 472–500. [Google Scholar] [CrossRef] [PubMed]
- Arends, J.; Bachmann, P.; Baracos, V.; Barthelemy, N.; Bertz, H.; Bozzetti, F.; Fearon, K.; Hütterer, E.; Isenring, E.; Kaasa, S.; et al. ESPEN Guidelines on Nutrition in Cancer Patients. Clin. Nutr. 2017, 36, 11–48. [Google Scholar] [CrossRef]


{kind=link}
| Title of Project | ClinicalTrials.gov | Participants (n) | Conditions | Treatment | Study Type | Locations | Current Status |
|---|---|---|---|---|---|---|---|
| The Mechanism of Enhancing the Anti-tumor Effects of CAR-T on PC by Gut Microbiota Regulation | NCT04203459 | 80 | Pancreatic cancer | nd | Observational | The first affiliated hospital of Harbin medical university Harbin, Heilongjiang, China | Recruiting |
| MS-20 on Gut Microbiota and Risk/Severity of Cachexia in Pancreatic Cancer Patients | NCT04600154 | 40 | Pancreatic cancer | Drug: MS-20 Other: Placebo | Interventional | Department of Internal Medicine, National Taiwan University Hospital Taipei, Taiwan | Recruiting |
| Gut Microbiome Modulation to Enable Efficacy of Checkpoint-based Immunotherapy in Pancreatic Adenocarcinoma | NCT03891979 | nd | Pancreatic cancer | Drug: Pembrolizumab Drug: Ciprofloxacin 500 mg PO BID days 1–29 Drug: Metronidazole 500 mg PO TID days 1–29 | Interventional | NYU Langone Health New York, NY, USA | Withdrawn |
| ARGONAUT: Stool and Blood Sample Bank for Cancer Patients | NCT04638751 | 4000 | Non-small Cell Lung Cancer Colorectal Cancer Triple NegTriple-Negativencer Pancreas Cancer | Drug: Immunotherapy Drug: Chemotherapeutic Agent | Observational | Persephone Biosciences, Inc. San Diego, CA, USA | Recruiting |
| Correlation Between Complications After Pancreaticoduodenectomy and Microbiota (COMPAMIC) | NCT04931069 | 30 | Pancreatic cancer | nd | Interventional | nd | Not yet recruiting |
| Microbial Diversity of Pancreatic Diseases | NCT03809247 | 330 | Pancreatic cancer Pancreatic diseases | nd | Observational | Ruijin Hospital Shanghai, Shanghai, China | Not yet recruiting |
| Microbiome Analysis in esoPhageal, PancreatIc and Colorectal Cancer Patients Undergoing Gastrointestinal Surgery (MA-PPING) | NCT04189393 | 60 | Gastrointestinal Cancer Colorectal Cancer Pancreatic Cancer Esophageal Cancer Rectum Neoplasm Esophageal Neoplasms Pancreatic Ductal Adenocarcinoma Colonic Neoplasms | nd | Observational | Nd | Not yet recruiting |
| The Microbiome of Pancreatic Cancer: “PANDEMIC” Study (PANDEMIC) | NCT04274972 | 20 | Pancreas Cancer Pancreas Infection Pancreas; Fistula | Diagnostic Test: Microbiome evaluation | Observational | AOUI Verona Verona, Italy | Recruiting |
| Volatiles in Breath and Headspace Analysis—Diagnostic Markers (Volatolome) | NCT03228095 | 3000 | Tuberculosis Gastric Cancer Peptic Ulcer Atrophic Gastritis Intestinal Metaplasia Gastric Dysplasia Colorectal Cancer Colorectal Polyp Colorectal Adenoma Pancreatic Cancer Pancreatitis, Chronic Liver Cancer Liver Cirrhosis Flu, Human Other Infectious Diseases Inflammatory Bowel Diseases | Diagnostic Test: VOC detection in breath and skin headspace Diagnostic Test: Breath sampling Procedure: Upper endoscopy with biopsies Procedure: Colonoscopy with biopsies Procedure: Whole blood/Plasma/serum sampling Diagnostic Test: Faecal sampling Procedure: Histological examination of the surgical specimen Diagnostic Test: Headspace analysis for biological material | Observational | University of Latvia Riga, Latvia | Enrolling by invitation |
| Nutrition in Gastrointestinal Tumors (NutriGIT) | NCT04476082 | 80 | Pancreatic Cancer Oesophageal Cancer Colon Cancer Liver Cancer Rectal Cancer Bile Duct Cancer GIST, Malignant Neuroendocrine Tumors Small Intestine Cancer Gastric Cancer | nd | Observational | University Medicine Greifswald Greifswald, Germany | Recruiting |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobocki, B.K.; Kaźmierczak-Siedlecka, K.; Folwarski, M.; Hawryłkowicz, V.; Makarewicz, W.; Stachowska, E. Pancreatic Cancer and Gut Microbiome-Related Aspects: A Comprehensive Review and Dietary Recommendations. Nutrients 2021, 13, 4425. https://doi.org/10.3390/nu13124425
Sobocki BK, Kaźmierczak-Siedlecka K, Folwarski M, Hawryłkowicz V, Makarewicz W, Stachowska E. Pancreatic Cancer and Gut Microbiome-Related Aspects: A Comprehensive Review and Dietary Recommendations. Nutrients. 2021; 13(12):4425. https://doi.org/10.3390/nu13124425
Chicago/Turabian StyleSobocki, Bartosz Kamil, Karolina Kaźmierczak-Siedlecka, Marcin Folwarski, Viktoria Hawryłkowicz, Wojciech Makarewicz, and Ewa Stachowska. 2021. "Pancreatic Cancer and Gut Microbiome-Related Aspects: A Comprehensive Review and Dietary Recommendations" Nutrients 13, no. 12: 4425. https://doi.org/10.3390/nu13124425
APA StyleSobocki, B. K., Kaźmierczak-Siedlecka, K., Folwarski, M., Hawryłkowicz, V., Makarewicz, W., & Stachowska, E. (2021). Pancreatic Cancer and Gut Microbiome-Related Aspects: A Comprehensive Review and Dietary Recommendations. Nutrients, 13(12), 4425. https://doi.org/10.3390/nu13124425