
Preventive Effect and Molecular Mechanism of Lactobacillus rhamnosus JL1 on Food-Borne Obesity in Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cholesterol-Lowering Ability of L. rhamnosus JL1 In Vitro
2.3. Animal Groups and Feeding
2.4. Determination of Blood Lipid Levels of Mice Serum
2.5. Determination of Inflammatory Cytokines of Mice Serum
2.6. Liver Histological Analysis
2.7. Determination of the mRNA Expression in Mouse Liver by RT-PCR
2.8. Determination of Protein Expression in Mouse Liver by Western Blotting
2.9. Analysis of SCFAs in the Faeces
2.10. Statistical Methods
3. Results
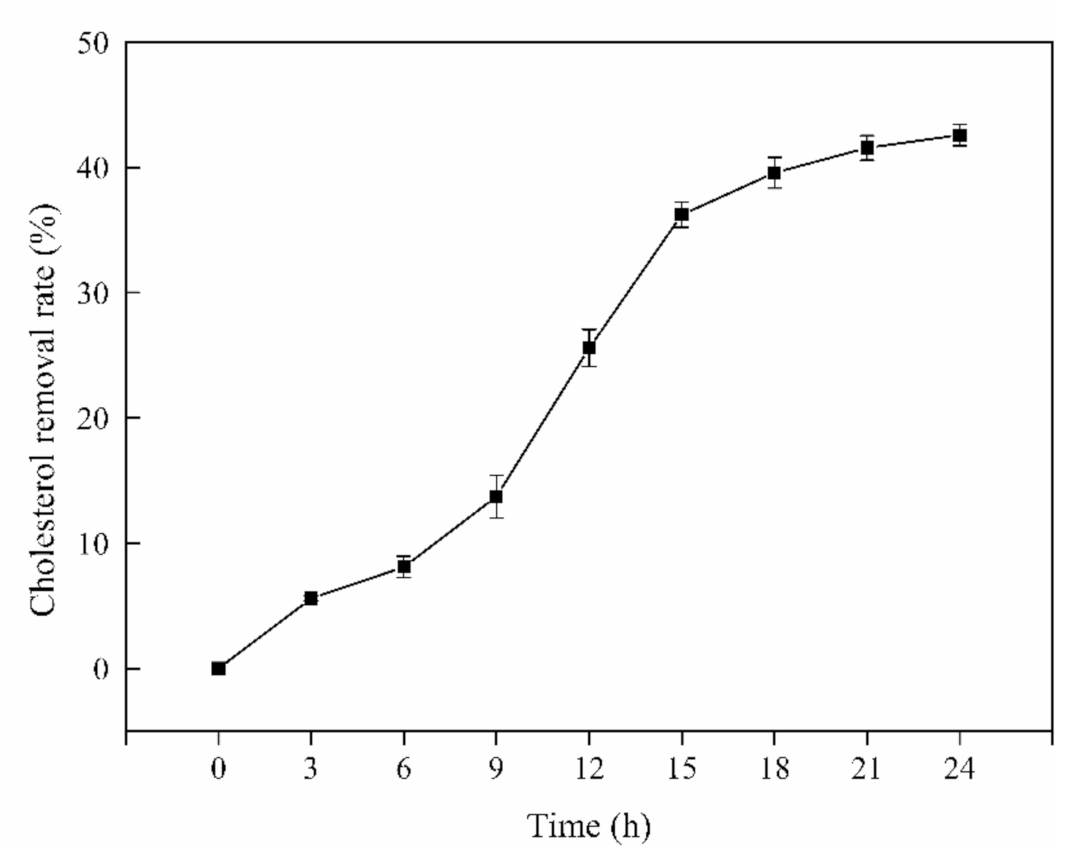
3.1. Cholesterol-Lowing Ability of L. Rhamnosus JL1 In Vitro
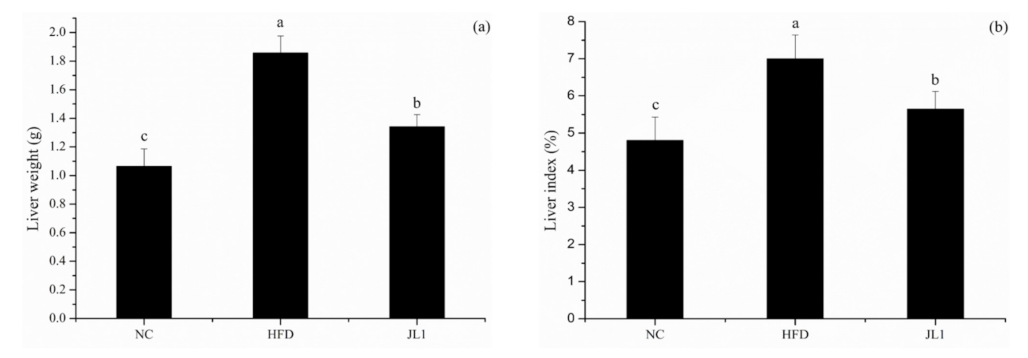
3.2. Effect of L. rhamnosus JL1 on Body Weight and the Liver Index of Mice
3.3. Effect of L. rhamnosus JL1 on the Levels of Serum Lipids in Mice
3.4. Cholesterol-Lowering Ability of L. rhamnosus JL1 In Vitro
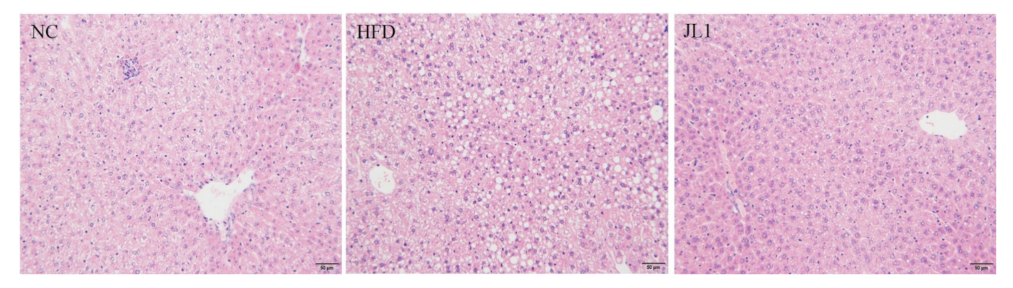
3.5. Liver Histological Image of Mice
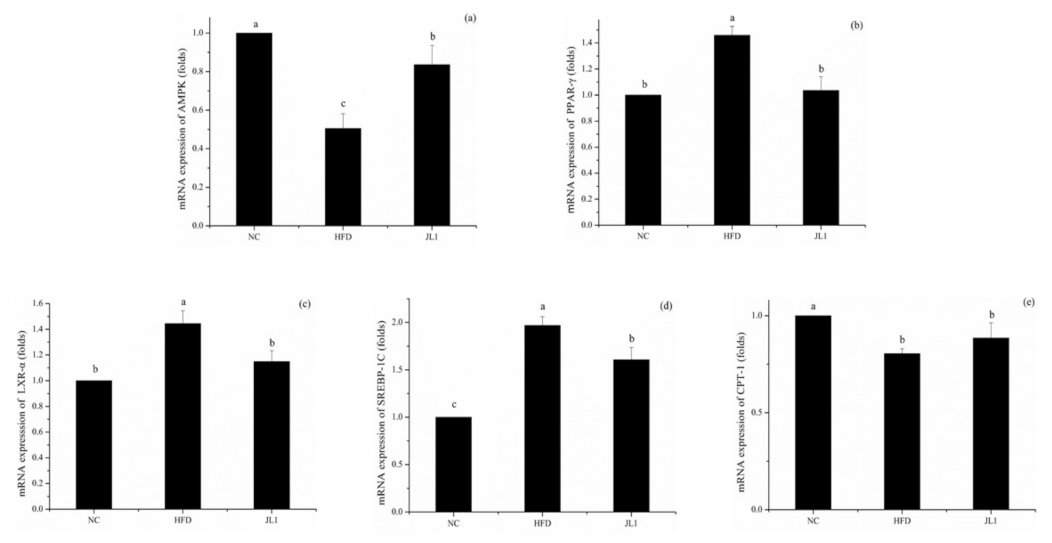
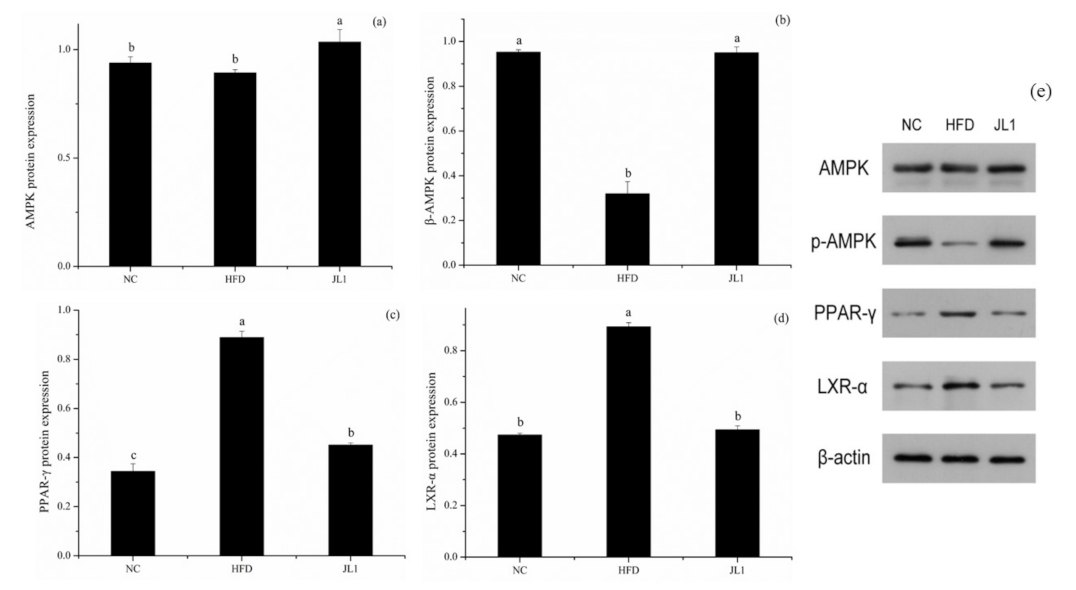
3.6. Effect of L. rhamnosus JL1 on the Expression of Genes and Proteins in Mice
3.7. Effect of L. rhamnosus JL1 on the Concentration of SCFAs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gregg, E.W.; Shaw, J.E. Global Health Effects of Overweight and Obesity. N. Engl. J. Med. 2017, 377, 80–81. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, Z.; Song, Y.; Liu, Y.; Zhao, H.; Liu, Y.; Zhang, T.; Yuan, Y.; Cai, X.; Wang, S.; et al. Study on urine metabolic profiling and pathogenesis of hyperlipidemia. Clin. Chim. Acta 2019, 495, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Kabagambe, E.K.; Tsai, M.Y.; Hopkins, P.N.; Ordovas, J.M.; Peacock, J.M.; Borecki, I.B.; Arnett, D.K. Erythrocyte fatty acid composition and the metabolic syndrome: A national heart, lung, and blood institute GOLDN study. Clin. Chem. 2008, 54, 154–162. [Google Scholar] [CrossRef]
- Rohr, M.W.; Narasimhulu, C.A.; Rudeski-Rohr, T.A.; Parthasarathy, S. Negative Effects of a High-Fat Diet on Intestinal Permeability: A Review. Adv. Nutr. 2020, 11, 77–91. [Google Scholar] [CrossRef] [Green Version]
- Guth, E. Counting Calories as an Approach to Achieve Weight Control. JAMA J. Am. Med. Assoc. 2018, 319, 225–226. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, W.P.; Wei, Z.H.; Yin, B.X.; Man, C.X.; Jiang, Y.J. Enhancement of functional characteristics of blueberry juice fermented by Lactobacillus plantarum. LWT-Food Sci. Technol. 2021, 139, 110590. [Google Scholar] [CrossRef]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 642. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Pontes, K.S.; Guedes, M.R.; da Cunha, M.R.; de Souza Mattos, S.; Silva, M.I.B.; Neves, M.F.; Marques, B.C.A.A.; Klein, M.R.S.T. Effects of probiotics on body adiposity and cardiovascular risk markers in individuals with overweight and obesity: A systematic review and meta-analysis of randomized controlled trials. Clin. Nutr. 2021, 40, 4915–4931. [Google Scholar] [CrossRef]
- Rittiphairoj, T.; Pongpirul, K.; Janchot, K.; Mueller, N.T.; Li, T. Probiotics Contribute to Glycemic Control in Patients with Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis. Adv. Nutr. 2021, 12, 722–734. [Google Scholar] [CrossRef]
- Tunapong, W.; Apaijai, N.; Yasom, S.; Tanajak, P.; Wanchai, K.; Chunchai, T.; Kerdphoo, S.; Eaimworawuthikul, S.; Thiennimitr, P.; Pongchaidecha, A.; et al. Chronic treatment with prebiotics, probiotics and synbiotics attenuated cardiac dysfunction by improving cardiac mitochondrial dysfunction in male obese insulin-resistant rats. Eur. J. Nutr. 2018, 57, 2091–2104. [Google Scholar] [CrossRef]
- De Filippis, F.; Pasolli, E.; Ercolini, D. The food-gut axis: Lactic acid bacteria and their link to food, the gut microbiome and human health. FEMS Microbiol. Rev. 2020, 44, 454–489. [Google Scholar] [CrossRef] [PubMed]
- Basturk, A.; Isik, I.; Atalay, A.; Yilmaz, A. Investigation of the Efficacy of Lactobacillus rhamnosus GG in Infants With Cow’s Milk Protein Allergy: A Randomised Double-Blind Placebo-Controlled Trial. Probiotics Antimicrob. Proteins 2020, 12, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Mian, M.F.; Neufeld, K.-A.M.; Forsythe, P. CD4 (+) CD25(+) T Cells are Essential for Behavioral Effects of Lactobacillus rhamnosus JB-1 in Male BALB/c mice. Brain Behav. Immun. 2020, 88, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.R.; Park, H.-J.; Kang, D.; Chung, H.; Nam, M.H.; Lee, Y.; Park, J.-H.; Lee, H.-Y. A protective mechanism of probiotic Lactobacillus against hepatic steatosis via reducing host intestinal fatty acid absorption. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Veiga, P.; Suez, J.; Derrien, M.; Elinav, E. Moving from probiotics to precision probiotics. Nat. Microbiol. 2020, 5, 878–880. [Google Scholar] [CrossRef]
- Li, X.; Hu, D.; Tian, Y.; Song, Y.; Hou, Y.; Sun, L.; Zhang, Y.; Man, C.; Zhang, W.; Jiang, Y. Protective effects of a novel Lactobacillus rhamnosus strain with probiotic characteristics against lipopolysaccharide-induced intestinal inflammation in vitro and in vivo. Food Funct. 2020, 11, 5799–5814. [Google Scholar] [CrossRef]
- Kuerman, M.; Bao, Y.H.; Guo, Y.; Guo, M. Effects of prebiotic carbohydrates on the growth promotion and cholesterol-lowering abilities of compound probiotics in vitro. LWT-Food Sci. Technol. 2019, 118, 108703. [Google Scholar] [CrossRef]
- Bagchi, D.P.; MacDougald, O.A. Identification and Dissection of Diverse Mouse Adipose Depots. JoVE-J. Vis. Exp. 2019, 149, 59499. [Google Scholar] [CrossRef]
- Sun, L.; Tian, W.; Guo, X.; Zhang, Y.; Liu, X.; Li, X.; Tian, Y.; Man, C.; Jiang, Y. Lactobacillus gasseri JM1 with potential probiotic characteristics alleviates inflammatory response by activating the PI3K/Akt signaling pathway in vitro. J. Dairy Sci. 2020, 103, 7851–7864. [Google Scholar] [CrossRef]
- Wang, G.; Xu, Q.; Jin, X.; Hang, F.; Liu, Z.; Zhao, J.; Zhang, H.; Chen, W. Effects of lactobacilli with different regulatory behaviours on tight junctions in mice with dextran sodium sulphate-induced colitis. J. Funct. Foods 2018, 47, 107–115. [Google Scholar] [CrossRef]
- Farhat, T.; Iannotti, R.J.; Simons-Morton, B.G. Overweight, Obesity, Youth, and Health-Risk Behaviors. Am. J. Prev. Med. 2010, 38, 258–267. [Google Scholar] [CrossRef] [Green Version]
- Bibbins-Domingo, K.; Grossman, D.C.; Curry, S.J.; Davidson, K.W.; Epling, J.W., Jr.; Garcia, F.A.R.; Gillman, M.W.; Kemper, A.R.; Krist, A.H.; Kurth, A.E.; et al. Statin Use for the Primary Prevention of Cardiovascular Disease in Adults US Preventive Services Task Force Recommendation Statement. Jama J. Am. Med. Assoc. 2016, 323, 669. [Google Scholar]
- Capurso, L. Thirty Years of Lactobacillus rhamnosus GG A Review. J. Clin. Gastroenterol. 2019, 53, S1–S41. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Chen, L.; Zhao, Y.; Wang, C.; Duan, C.; Yang, G.; Niu, C.; Li, S. Lactobacillus plantarum NA136 ameliorates nonalcoholic fatty liver disease by modulating gut microbiota, improving intestinal barrier integrity, and attenuating inflammation. Appl. Microbiol. Biotechnol. 2020, 104, 5273–5282. [Google Scholar] [CrossRef]
- Woo, M.-N.; Bok, S.-H.; Lee, M.-K.; Kim, H.-J.; Jeon, S.-M.; Do, G.-M.; Shin, S.K.; Ha, T.Y.; Choi, M.-S. Anti-obesity and hypolipidemic effects of a proprietary herb and fiber combination (S&S PWH) in rats fed high-fat diets. J. Med. Food 2008, 11, 169–178. [Google Scholar]
- Irudayaraj, S.S.; Sunil, C.; Duraipandiyan, V.; Ignacimuthu, S. In vitro antioxidant and antihyperlipidemic activities of Toddalia asiatica (L) Lam. Leaves in Triton WR-1339 and high fat diet induced hyperlipidemic rats. Food Chem. Toxicol. 2013, 60, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Meenu, M.; Xu, B. A critical review on anti-diabetic and anti-obesity effects of dietary resistant starch. Crit. Rev. Food Sci. Nutr. 2019, 59, 3019–3031. [Google Scholar] [CrossRef]
- Pirillo, A.; Casula, M.; Olmastroni, E.; Norata, G.D.; Catapano, A.L. Global epidemiology of dyslipidaemias. Nat. Rev. Cardiol. 2021, 18, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Tian, Z.; Li, M.; Li, B.; Cui, W. Effects of Probiotic Supplementation Regulating Blood Lipid Levels in Overweight or Obese Subjects: A Meta-analysis. Food Funct. 2019, 86, 104711. [Google Scholar]
- Yoo, S.-R.; Kim, Y.-J.; Park, D.-Y.; Jung, U.-J.; Jeon, S.-M.; Ahn, Y.-T.; Huh, C.-S.; McGregor, R.; Choi, M.S. Probiotics L. plantarum and L. curvatus in Combination Alter Hepatic Lipid Metabolism and Suppress Diet-Induced Obesity. Obesity 2013, 21, 2571–2578. [Google Scholar] [CrossRef]
- Barrea, L.; Muscogiuri, G.; Laudisio, D.; Di Somma, C.; Salzano, C.; Pugliese, G.; de Alteriis, G.; Colao, A.; Savastano, S. Phase Angle: A Possible Biomarker to Quantify Inflammation in Subjects with Obesity and 25(OH)D Deficiency. Nutrients 2019, 11, 1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liou, C.J.; Lee, Y.K.; Ting, N.C.; Chen, Y.L.; Shen, S.C.; Wu, S.J.; Huang, W.C. Protective Effects of Licochalcone A Ameliorates Obesity and Non-Alcoholic Fatty Liver Disease Via Promotion of the Sirt-1/AMPK Pathway in Mice Fed a High-Fat Diet. Cells 2019, 8, 447. [Google Scholar] [CrossRef] [Green Version]
- Jianming, L.; Jiamei, W.; Wang, Z.; Liu, G.; Zhang, Q. Antiobesity Effect of Flaxseed Polysaccharide via Inducing Satiety due to Leptin Resistance Removal and Promoting Lipid Metabolism through the AMP-Activated Protein Kinase (AMPK) Signaling Pathway. J. Agric. Food Chem. 2019, 67, 7040–7049. [Google Scholar]
- Sag, D.; Carling, D.; Stout, R.D.; Suttles, J. Adenosine 5′-Monophosphate-Activated Protein Kinase Promotes Macrophage Polarization to an Anti-Inflammatory Functional Phenotype. J. Immunol. 2008, 181, 8633–8641. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, C.; Zhang, L.; Zhao, Y.; Duan, C.; Zhang, X.; Gao, L.; Li, S. Lactobacillus plantarum NA136 improves the non-alcoholic fatty liver disease by modulating the AMPK/Nrf2 pathway. Appl. Microbiol. Biotechnol. 2019, 103, 5843–5850. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Li, Y.; Du, Y.; Guo, L.; Chen, M.; Huang, X.; Yang, F.; Hong, J.; Kong, X. Konjaku flour reduces obesity in mice by modulating the composition of the gut microbiota. Int. J. Obes. 2019, 43, 1631–1643. [Google Scholar] [CrossRef]
- Zheng, S.; Huang, H.; Li, Y.; Wang, Y.; Zheng, Y.; Liang, J.; Zhang, S.; Liu, M.; Fang, Z. Yin-xing-tong-mai decoction attenuates atherosclerosis via activating PPAR gamma-LXR alpha-ABCA1/ABCG1 pathway. Pharmacol. Res. 2021, 169, 105639. [Google Scholar] [CrossRef]
- Liver X receptors in lipid metabolism: Opportunities for drug discovery. Nat. Rev. Drug Discov. 2014, 13, 433–444. [CrossRef]
- Hafiane, A.; Gasbarrino, K.; Daskalopoulou, S.S. The role of adiponectin in cholesterol efflux and HDL biogenesis and metabolism. Metab.-Clin. Exp. 2019, 100, 153953. [Google Scholar] [CrossRef]
- Truax, A.D.; Chen, L.; Tam, J.W.; Cheng, N.; Guo, H.; Koblansky, A.A.; Chou, W.C.; Wilson, J.E.; Brickey, W.J.; Petrucelli, A.; et al. The Inhibitory Innate Immune Sensor NLRP12 Maintains a Threshold against Obesity by Regulating Gut Microbiota Homeostasis. Cell Host Microbe 2018, 24, 364–378. [Google Scholar] [CrossRef] [Green Version]
- Canfora, E.; Jocken, J.W.; Blaak, E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; Meex, R.C.R.; Venema, K.; Blaak, E.E. Gut microbial metabolites in obesity, NAFLD and T2DM. Nat. Rev. Endocrinol. 2019, 15, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Ji, Y.; Jung, H.-Y.; Park, H.; Kang, J.; Choi, S.-H.; Shin, H.; Hyun, C.K.; Kim, K.T.; Holzapfel, W.H. Lactobacillus plantarum HAC01 regulates gut microbiota and adipose tissue accumulation in a diet-induced obesity murine model. Appl. Microbiol. Biotechnol. 2017, 101, 1605–1614. [Google Scholar] [CrossRef] [PubMed]
- Den Besten, G.; Bleeker, A.; Gerding, A.; van Eunen, K.; Havinga, R.; van Dijk, T.H.; Oosterveer, M.H.; Jonker, J.W.; Groen, A.K.; Reijngoud, D.-J.; et al. Short-Chain Fatty Acids Protect Against High-Fat Diet-Induced Obesity via a PPAR-Dependent Switch From Lipogenesis to Fat Oxidation. Diabetes 2015, 64, 2398–2408. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Diet | Treatment |
|---|---|---|
| NC (Normal control) | Normal diet | 0.2 mL PBS per day |
| HFD (High-fat diet) | High-fat diet | 0.2 mL PBS per day |
| JL1 (L. rhamnosus JL1) | High-fat diet | 0.2 mL PBS + 109 CFU/mL L. rhamnosus JL1 per day |
| Genes | Primer Sequences | |
|---|---|---|
| Forward (5′–3′) | Reverse (5′–3′) | |
| β-actin | CTACCTCATGAAGATCCTGACC | CACAGCTTCTCTTTGATGTCAC |
| AMPK | CAACTATCGATCTTGCCAAAGG | AACAGGAGAAGAGTCAAGTGAG |
| PPAR-γ | TCCATTCACAAGAGCTGACCC | GGTGGAGATGCAGGTTCTACT |
| LXR-α | TGACTTTGCCAAACAGCTCC | TGTACCTCCGTGACGTCTC |
| SREBP-1C | CTGTGTGACCTGCTTCTTGT | CTCATGTAGGAACACCCTCC |
| Group | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| NC (g) | 18.33 ± 1.05 a | 19.05 ± 0.94 a | 20.42 ± 0.70 b | 21.59 ± 0.84 b | 21.77 ± 1.75 c | 21.59 ± 1.2 b | 21.63 ± 0.57 c | 21.32 ± 0.71 c | 22.05 ± 0.77 b | 22.22 ± 0.64 c |
| HFD (g) | 18.29 ± 0.65 a | 18.77 ± 0.77 ab | 21.53 ± 0.52 a | 23.12 ± 0.60 a | 24.51 ± 0.39 a | 23.71 ± 0.81 a | 23.91 ± 1.05 ab | 24.04 ± 0.94 a | 25.15 ± 0.69 a | 26.59 ± 1.17 a |
| JL1 (g) | 18.25 ± 0.84 a | 18.44 ± 1.33 b | 21.34 ± 1.25 a | 21.28 ± 1.30 b | 23.07 ± 1.12 b | 23.27 ± 1.42 a | 23.50 ± 1.56 ab | 23.38 ± 0.86 b | 22.83 ± 0.96 b | 23.78 ± 0.84 b |
| Group | TC (mmol/L) | TG (mmol/L) | LDL-C (mmol/L) | HDL-C (mmol/L) |
|---|---|---|---|---|
| NC | 3.85 ± 0.38 c | 1.34 ± 0.28 c | 0.51 ± 0.07 c | 4.74 ± 1.33 a |
| HFD | 6.09 ± 0.73 a | 2.02 ± 0.38 a | 0.98 ± 0.14 a | 3.20 ± 1.04 b |
| JL1 | 4.98 ± 0.68 b | 1.69 ± 0.23 b | 0.69 ± 0.08 b | 4.30 ± 0.90 ab |
| IL-1β (ng/L) | IL-6 (ng/L) | IFN-γ (ng/L) | TNF-α (ng/L) | |
|---|---|---|---|---|
| NC | 16.98 ± 2.00 c | 45.76 ± 9.21 b | 185.86 ± 19.16 c | 187.20 ± 19.63 c |
| HFD | 30.23 ± 6.57 a | 69.55 ± 10.56 a | 284.30 ± 16.04 a | 297.54 ± 29.44 a |
| JL1 | 24.17 ± 5.08 b | 49.48 ± 11.65 b | 227.80 ± 11.28 b | 257.60 ± 29.79 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.; Zheng, J.; Zong, X.; Yang, X.; Zhang, Y.; Man, C.; Jiang, Y. Preventive Effect and Molecular Mechanism of Lactobacillus rhamnosus JL1 on Food-Borne Obesity in Mice. Nutrients 2021, 13, 3989. https://doi.org/10.3390/nu13113989
Yang M, Zheng J, Zong X, Yang X, Zhang Y, Man C, Jiang Y. Preventive Effect and Molecular Mechanism of Lactobacillus rhamnosus JL1 on Food-Borne Obesity in Mice. Nutrients. 2021; 13(11):3989. https://doi.org/10.3390/nu13113989
Chicago/Turabian StyleYang, Mo, Jiapeng Zheng, Xinran Zong, Xinyan Yang, Yu Zhang, Chaoxin Man, and Yujun Jiang. 2021. "Preventive Effect and Molecular Mechanism of Lactobacillus rhamnosus JL1 on Food-Borne Obesity in Mice" Nutrients 13, no. 11: 3989. https://doi.org/10.3390/nu13113989
APA StyleYang, M., Zheng, J., Zong, X., Yang, X., Zhang, Y., Man, C., & Jiang, Y. (2021). Preventive Effect and Molecular Mechanism of Lactobacillus rhamnosus JL1 on Food-Borne Obesity in Mice. Nutrients, 13(11), 3989. https://doi.org/10.3390/nu13113989