
The Role of Nutritional Lifestyle and Physical Activity in Multiple Sclerosis Pathogenesis and Management: A Narrative Review

 , ,
, , 



Abstract
:1. Introduction
2. Nutritional Lifestyle as Risk Factor or Complementary Treatment for Patients with Multiple Sclerosis
2.1. Food-Derived Factors
2.2. Diets
2.2.1. Low-Carbohydrate Diets
2.2.2. Gluten-Free Diet (GFD)
2.2.3. Mediterranean Diet (MD)
2.2.4. Low-Fat Diet: Swank and McDougall Diets
2.2.5. Fasting-Mimicking Diets (FMD)
2.2.6. Western Diets (WD)
3. Clinical Impact of Exercise in Patients with Multiple Sclerosis
3.1. Biological and Radiological Modifications Induced by Physical Activity in EAE/MS
3.1.1. Pre-Clinical Studies
3.1.2. Clinical Studies
3.2. Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wootla, B.; Eriguchi, M.; Rodriguez, M. Is multiple sclerosis an autoimmune disease? Autoimmune Dis. 2012, 2012, 969657. [Google Scholar] [CrossRef]
- Nourbakhsh, B.; Mowry, E.M. Multiple sclerosis risk factors and pathogenesis. Continuum 2019, 25, 596–610. [Google Scholar] [CrossRef]
- Drehmer, E.; Platero, J.L.; Carrera-Juliá, S.; Moreno, M.L.; Tvarijonaviciute, A.; Navarro, M.Á.; Lopez-Rodriguez, M.M.; de la Rubia Orti, J.E. The relation between eating habits and abdominal fat, anthropometry, PON1 and IL-6 levels in patients with multiple sclerosis. Nutrients 2020, 12, 744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Y.; Van Der Mei, I.A.F.; Simpson, S.; Ponsonby, A.L.; Lucas, R.M.; Tettey, P.; Charlesworth, J.; Kostner, K.; Taylor, B.V. Lipid-related genetic polymorphisms significantly modulate the association between lipids and disability progression in multiple sclerosis. J. Neurol. Neurosurg. Psychiatry 2019, 90, 636–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo-Barreiro, L.; Eixarch, H.; Montalban, X.; Espejo, C. Combined therapies to treat complex diseases: The role of the gut microbiota in multiple sclerosis. Autoimmun. Rev. 2018, 17, 165–174. [Google Scholar] [CrossRef]
- Schepici, G.; Silvestro, S.; Bramanti, P.; Mazzon, E. The Gut Microbiota in Multiple Sclerosis: An Overview of Clinical Trials. Cell Transplant. 2019, 28, 1507–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wade, D.T.; Young, C.A.; Chaudhuri, K.R.; Davidson, D.L.W. A randomised placebo controlled exploratory study of vitamin B-12, lofepramine, and L-phenylalanine (the “Cari Loder regime”) in the treatment of multiple sclerosis. J. Neurol. Neurosurg. Psych. 2002, 73, 246–249. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, D.A.; Schandl, E.K. Carbon monoxide, vitamin B6, and multiple sclerosis: A theory of interrelationship. Am. J. Clin. Nutr. 1973, 26, 890–896. [Google Scholar] [CrossRef]
- Goldberg, P. Multiple sclerosis: Vitamin D and calcium as environmental determinants of prevalence. Int. J. Environ. Stud. 1974, 6, 19–27. [Google Scholar] [CrossRef]
- Goldberg, P.; Fleming, M.C.; Picard, E.H. Multiple sclerosis: Decreased relapse rate through dietary supplementation with calcium, magnesium and vitamin D. Med. Hypotheses 1983, 21, 193–200. [Google Scholar] [CrossRef]
- Salemi, G.; Gueli, M.C.; Vitale, F.; Battaglieri, F.; Guglielmini, E.; Ragonese, P.; Trentacosti, A.; Massenti, M.F.; Savettieri, G.; Bono, A. Blood lipids, homocysteine, stress factors, and vitamins in clinically stable multiple sclerosis patients. Lipids Health Dis. 2010, 18, 9. [Google Scholar] [CrossRef] [Green Version]
- Radzikowska, U.; Rinaldi, A.O.; Sözener, Z.Ç.; Karaguzel, D.; Wojcik, M.; Cypryk, K.; Akdis, M.; Sokolowska, M. The influence of dietary fatty acids on immune responses. Nutrients 2019, 11, 2990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unoda, K.; Doi, Y.; Nakajima, H.; Yamane, K.; Hosokawa, T.; Ishida, S.; Kimura, F.; Hanafusa, T. Eicosapentaenoic acid (EPA) induces peroxisome proliferator-activated receptors and ameliorates experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2013, 256, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Yen, J.H.; Ganea, D. Docosahexaenoic acid prevents dendritic cell maturation, inhibits antigen-specific Th1/Th17 differentiation and suppresses experimental autoimmune encephalomyelitis. Brain Behav. Immun. 2011, 25, 872–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinto, L.; Marracci, G.; Baldauf-Wagner, S.; Strehlow, A.; Yadav, V.; Stuber, L.; Bourdette, D. Omega-3 fatty acid supplementation decreases matrix metalloproteinase-9 production in relapsing-remitting multiple sclerosis. Prostaglandins Leukot. Essent. Fat Acids 2009, 80, 131–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouchaki, E.; Afarini, M.; Abolhassani, J.; Mirhosseini, N.; Bahmani, F.-; Masoud, S.A.; Asemi, Z. High-dose ω-3 fatty acid plus Vitamin D3 supplementation affects clinical symptoms and metabolic status of patients with multiple sclerosis: A randomized controlled clinical trial. J. Nutr. 2018, 148, 1380–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haider, L. Inflammation, Iron, Energy Failure, and Oxidative Stress in the Pathogenesis of Multiple Sclerosis. Oxid. Med. Cell. Longev. 2015, 2015, 725370. [Google Scholar] [CrossRef]
- Hendriks, J.J.A.; Alblas, J.; Van Der Pol, S.M.A.; Van Tol, E.A.F.; Dijkstra, C.D.; De Vries, H.E. Flavonoids influence monocytic GTPase activity and are protective in experimental allergic encephalitis. J. Exp. Med. 2004, 200, 1667–1672. [Google Scholar] [CrossRef] [Green Version]
- Sternberg, Z.; Chadha, K.; Lieberman, A.; Drake, A.; Hojnacki, D.; Weinstock-Guttman, B.; Munschauer, F. Immunomodulatory responses of peripheral blood mononuclear cells from multiple sclerosis patients upon in vitro incubation with the flavonoid luteolin: Additive effects of IFN-β. J. Neuroinflamm. 2009, 6, 28. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, X.; Ciric, B.; Ma, C.G.; Gran, B.; Rostami, A.; Zhang, G.X. Therapeutic effect of baicalin on experimental autoimmune encephalomyelitis is mediated by SOCS3 regulatory pathway. Sci. Rep. 2015, 5, 17407. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.Y.; Cha, H.J.; Choi, E.O.; Kim, C.H.; Kim, G.Y.; Yoo, Y.H.; Hwang, H.J.; Parkm, H.T.; Yoon, H.M.; Choi, Y.H. Activation of the Nrf2/HO-1 signaling pathway contributes to the protective effects of baicalein against oxidative stress-induced DNA damage and apoptosis in HEI193 schwann cells. Int. J. Med. Sci. 2019, 16, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Li, X.K.; Funeshima-Fuji, N.; Kimura, H.; Matsumoto, Y.; Isaka, Y.; Takahara, S. Amelioration of experimental autoimmune encephalomyelitis by curcumin treatment through inhibition of IL-17 production. Int. Immunopharmacol. 2009, 9, 575–581. [Google Scholar] [CrossRef]
- Natarajan, C.; Bright, J.J. Curcumin Inhibits Experimental Allergic Encephalomyelitis by Blocking IL-12 Signaling Through Janus Kinase-STAT Pathway in T Lymphocytes. J. Immunol. 2002, 168, 6506–6513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahey, A.J.; Adrian Robins, R.; Constantinescu, C.S. Curcumin modulation of IFN-β and IL-12 signalling and cytokine induction in human T cells. J. Cell. Mol. Med. 2007, 11, 1129–1137. [Google Scholar] [CrossRef]
- Dolati, S.; Ahmadi, M.; Aghebti-Maleki, L.; Nikmaram, A.; Marofi, F.; Rikhtegar, R.; Ayromlou, H.; Yousefi, M. Nanocurcumin is a potential novel therapy for multiple sclerosis by influencing inflammatory mediators. Pharmacol. Rep. 2018, 70, 1158–1167. [Google Scholar] [CrossRef]
- Seyedzadeh, M.H.; Safari, Z.; Zare, A.; Gholizadeh Navashenaq, J.; Razavi, S.A.; Kardar, G.A.; Khorramizadeh, M.R. Study of curcumin immunomodulatory effects on reactive astrocyte cell function. Int. Immunopharmacol. 2014, 22, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.P.; Hegde, V.L.; Hofseth, L.J.; Nagarkatti, M.; Nagarkatti, P.S. Resveratrol (trans-3,5,4′-trihydroxystilbene) ameliorates experimental allergic encephalomyelitis, primarily via induction of apoptosis in T cells involving activation of aryl hydrocarbon receptor and estrogen receptor. Mol. Pharmacol. 2007, 72, 1508–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imler, T.J.; Petro, T.M. Decreased severity of experimental autoimmune encephalomyelitis during resveratrol administration is associated with increased IL-17+IL-10+ T cells, CD4- IFN-γ+ cells, and decreased macrophage IL-6 expression. Int. Immunopharmacol. 2009, 9, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Shindler, K.S.; Ventura, E.; Dutt, M.; Elliott, P.; Fitzgerald, D.C.; Rostami, A. Oral resveratrol reduces neuronal damage in a model of multiple sclerosis. J. Neurophthalmol. 2010, 30, 328–339. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Li, S.P.; Fu, J.S.; Zhang, S.; Bai, L.; Guo, L. Resveratrol defends blood-brain barrier integrity in experimental autoimmune encephalomyelitis mice. J. Neurophysiol. 2016, 116, 2173–2179. [Google Scholar] [CrossRef] [Green Version]
- Gandy, K.A.O.; Zhang, J.; Nagarkatti, P.; Nagarkatti, M. Resveratrol (3, 5, 4′-Trihydroxy-trans-Stilbene) Attenuates a Mouse Model of Multiple Sclerosis by Altering the miR-124/Sphingosine Kinase 1 Axis in Encephalitogenic T Cells in the Brain. J. Neuroimmune Pharmacol. 2019, 14, 462–477. [Google Scholar] [CrossRef] [PubMed]
- Riccio, P.; Rossano, R.; Larocca, M.; Trotta, V.; Mennella, I.; Vitaglione, P.; Ettore, M.; Graverini, A.; De Santis, A.; Di Monte, E.; et al. Anti-inflammatory nutritional intervention in patients with relapsing-remitting and primary-progressive multiple sclerosis: A pilot study. Exp. Biol. Med. 2016, 241, 620–635. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Ren, Z.; Xu, Y.; Xiao, S.; Meydani, S.N.; Wu, D. Epigallocatechin-3-gallate ameliorates experimental autoimmune encephalomyelitis by altering balance among CD4 + T-cell subsets. Am. J. Pathol. 2012, 180, 221–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellmann-Strobl, J.; Paul, F.; Wuerfel, J.; Dörr, J.; Infante-Duarte, C.; Heidrich, E.; Kortgen, B.; Brandt, A.; Pfuller, C.; Radbruch, H.; et al. Epigallocatechin Gallate in Relapsing-Remitting Multiple Sclerosis: A Randomized, Placebo-Controlled Trial. Neurol. Neuroimmunol. Neuroinflamm. 2021, 8, e9813. [Google Scholar] [CrossRef]
- Johnston, G.A.R. Flavonoid nutraceuticals and ionotropic receptors for the inhibitory neurotransmitter GABA. Neurochem. Int. 2015, 89, 120–125. [Google Scholar] [CrossRef]
- Mähler, A.; Steiniger, J.; Bock, M.; Klug, L.; Parreidt, N.; Lorenz, M.; Zimmermann, B.F.; Krannich, A.; Paul, F.; Boschmann, M. Metabolic response to epigallocatechin-3-gallate in relapsing-remitting multiple sclerosis: A randomized clinical trial. Am. J. Clin. Nutr. 2015, 101, 487–495. [Google Scholar] [CrossRef]
- Platero, J.L.; Cuerda-Ballester, M.; Sancho-Cantus, D.; Benlloch, M.; Ceron, J.J.; Rubio, C.P.; Garcia-Pardo, M.P.; Lopez-Rodriguez, M.M.; de La Rubia Orti, J.E. The impact of epigallocatechin gallate and coconut oil treatment on cortisol activity and depression in multiple sclerosis patients. Life 2021, 11, 353. [Google Scholar] [CrossRef] [PubMed]
- Coe, S.; Cossington, J.; Collett, J.; Soundy, A.; Izadi, H.; Ovington, M.; Durkin, L.; Kirsten, M.; Clegg, M.; Cavey, A.; et al. A randomised double-blind placebo-controlled feasibility trial of flavonoid-rich cocoa for fatigue in people with relapsing and remitting multiple sclerosis. J. Neurol. Neurosurg. Psych. 2019, 90, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Herden, L.; Weissert, R. The effect of coffee and caffeine consumption on patients with multiple sclerosis-related fatigue. Nutrients 2020, 12, 2262. [Google Scholar] [CrossRef]
- Wang, T.; Xi, N.N.; Chen, Y.; Shang, X.F.; Hu, Q.; Chen, J.F.; Zheng, R.J. Chronic caffeine treatment protects against experimental autoimmune encephalomyelitis in mice: Therapeutic window and receptor subtype mechanism. Neuropharmacology 2014, 86, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Q.; Chen, Y.Y.; Wang, X.S.; Wu, S.Z.; Yang, H.M.; Xu, H.Q.; He, J.C.; Wang, X.T.; Chen, J.F.; Zheng, R.Y. Chronic caffeine treatment attenuates experimental autoimmune encephalomyelitis induced by guinea pig spinal cord homogenates in Wistar rats. Brain Res. 2010, 1309, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Caslin, B.; Maguire, C.; Karmakar, A.; Mohler, K.; Wylie, D.; Melamed, E. Alcohol shifts gut microbial networks and ameliorates a murine model of neuroinflammation in a sex-specific pattern. Proc. Natl. Acad. Sci. USA 2019, 116, 25808–25815. [Google Scholar] [CrossRef] [PubMed]
- Salari, S.; Khomand, P.; Arasteh, M.; Yousefzamani, B.; Hassanzadeh, K. Zinc sulphate: A reasonable choice for depression management in patients with multiple sclerosis: A randomized, double-blind, placebo-controlled clinical trial. Pharmacol. Rep. 2015, 67, 606–609. [Google Scholar] [CrossRef] [PubMed]
- Stoye, D.; Schubert, C.; Goihl, A.; Guttek, K.; Reinhold, A.; Brocke, S.; Grungreiff, K.; Reinhold, D. Zinc aspartate suppresses T cell activation in vitro and relapsing experimental autoimmune encephalomyelitis in SJL/J mice. BioMetals 2012, 25, 529–539. [Google Scholar] [CrossRef]
- Fitzgerald, K.C.; Munger, K.L.; Hartung, H.P.; Freedman, M.; Montalbán, X.; Edan, G.; Wicklein, E.M.; Radue, E.M.; Kappos, L.; Pohl, C. Sodium intake and multiple sclerosis activity and progression in BENEFIT. Ann. Neurol. 2017, 82, 20–29. [Google Scholar] [CrossRef]
- Hernandez, A.L.; Kitz, A.; Wu, C.; Lowther, D.E.; Rodriguez, D.M.; Vudattu, N.; Deng, S.; Herold, K.C.; Kuchroo, V.K.; Kleinewietfeld, M.; et al. Sodium chloride inhibits the suppressive function of FOXP3+ regulatory T cells. J. Clin. Investig. 2015, 125, 4212–4222. [Google Scholar] [CrossRef] [Green Version]
- Wilck, N.; Matus, M.G.; Kearney, S.M.; Olesen, S.W.; Forslund, K.; Bartolomaeus, H.; Haase, S.; Mahler, A.; Balogh, A.; Marko, L.; et al. Salt-responsive gut commensal modulates TH17 axis and disease. Nature 2017, 551, 585–589. [Google Scholar] [CrossRef]
- Nourbakhsh, B.; Graves, J.; Casper, T.C.; Lulu, S.; Waldman, A.; Belman, A.; Greenberg, B.; Weinstock-Guttman, B.; Aaen, G.; Tillema, J.M.; et al. Dietary salt intake and time to relapse in paediatric multiple sclerosis. J. Neurol. Neurosurg. Psych. 2016, 87, 1350–1353. [Google Scholar] [CrossRef] [Green Version]
- McDonald, J.; Graves, J.; Waldman, A.; Lotze, T.; Schreiner, T.; Belman, A.; Greenberg, B.; Weinstock-Guttman, B.; Aaen, G.; Tillema, J.M. A case-control study of dietary salt intake in pediatric-onset multiple sclerosis. Mult. Scler. Relat. Disord. 2016, 6, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Farez, M.F.; Fiol, M.P.; Gaitán, M.I.; Quintana, F.J.; Correale, J. Sodium intake is associated with increased disease activity in multiple sclerosis. J. Neurol. Neurosurg. Psychiatry 2015, 86, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Na, S.Y.; Janakiraman, M.; Leliavski, A.; Krishnamoorthy, G. High-salt diet suppresses autoimmune demyelination by regulating the blood–brain barrier permeability. Proc. Natl. Acad. Sci. USA 2021, 118, e2025944118. [Google Scholar] [CrossRef]
- Sharif, K.; Watad, A.; Bragazzi, N.L.; Adawi, M.; Amital, H.; Shoenfeld, Y. Coffee and autoimmunity: More than a mere hot beverage! Autoimmun. Rev. 2017, 16, 712–721. [Google Scholar] [CrossRef]
- Simpson, S.; Blizzard, L.; Otahal, P.; Van Der Mei, I.; Taylor, B. Latitude is significantly associated with the prevalence of multiple sclerosis: A meta-analysis. J. Neurol. Neurosurg. Psychiatry 2011, 82, 1132–1141. [Google Scholar] [CrossRef]
- Ascherio, A.; Munger, K.L.; White, R.; Köchert, K.; Simon, K.C.; Polman, C.H.; Freedman, M.S.; Hartung, H.P.; Miller, D.H.; Montalban, X.; et al. Vitamin D as an early predictor of multiple sclerosis activity and progression. JAMA Neurol. 2014, 71, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.; Taylor, B.; Blizzard, L.; Ponsonby, A.L.; Pittas, F.; Tremlett, H.; Dwyer, T.; Gies, P.; van der Mei, I. Higher 25-hydroxyvitamin D is associated with lower relapse risk in multiple sclerosis. Ann. Neurol. 2010, 68, 193–203. [Google Scholar] [CrossRef]
- Jagannath, V.A.; Filippini, G.; Di Pietrantonj, C.; Asokan, G.V.; Robak, E.W.; Whamond, L.; Robinson, S.A. Vitamin D for the management of multiple sclerosis. Cochrane Database Syst. Rev. 2018, 9, CD008422. [Google Scholar] [CrossRef] [PubMed]
- Feige, J.; Moser, T.; Bieler, L.; Schwenker, K.; Hauer, L.; Sellner, J. Vitamin D supplementation in multiple sclerosis: A critical analysis of potentials and threats. Nutrients 2020, 12, 783. [Google Scholar] [CrossRef] [Green Version]
- Mallone, F.; Lucchino, L.; Franzone, F.; Marenco, M.; Carlesimo, S.C.; Moramarco, A. High-dose vitamin B supplementation for persistent visual deficit in multiple sclerosis: A pilot study. Drug. Discov. Ther. 2020, 14, 122–128. [Google Scholar] [CrossRef]
- Nanduri, R.; Mahajan, S.; Bhagyaraj, E.; Sethi, K.; Kalra, R.; Chandra, V.; Gupta, P. The active form of vitamin D transcriptionally represses Smad7 signaling and activates extracellular signal-regulated kinase (ERK) to inhibit the differentiation of a inflammatory T helper cell subset and suppress experimental autoimmune encephalomyelitis. J. Biol. Chem. 2015, 290, 12222–12236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jafarzadeh, A.; Azizi, S.V.; Arabi, Z.; Ahangar-Parvin, R.; Mohammadi-Kordkhayli, M.; Larussa, T.; Khatami, L.; Nemati, M. Vitamin D down-regulates the expression of some Th17 cell-related cytokines, key inflammatory chemokines, and chemokine receptors in experimental autoimmune encephalomyelitis. Nutr. Neurosci. 2019, 22, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Sloka, S.; Silva, C.; Wang, J.; Yong, V.W. Predominance of Th2 polarization by Vitamin D through a STAT6-dependent mechanism. J. Neuroinflamm. 2011, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Chen, J.; Zheng, C.; Wu, J.; Cheng, Y.; Zhu, S.; Lin, C.; Cao, Q.; Zhu, J.; Jin, T. 1,25-dihydroxyvitamin D3-induced dendritic cells suppress experimental autoimmune encephalomyelitis by increasing proportions of the regulatory lymphocytes and reducing T helper type 1 and type 17 cells. Immunology 2017, 152, 414–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farias, A.S.; Spagnol, G.S.; Bordeaux-Rego, P.; Oliveira, C.O.F.; Fontana, A.G.M.; de Paula, R.F.O.; Santos, M.P.A.; Pradella, F.; Moraes, A.S.; Oliveira, E.C.; et al. Vitamin D3 induces IDO+ tolerogenic DCs and enhances treg, reducing the severity of EAE. CNS Neurosci. Ther. 2013, 19, 269–277. [Google Scholar] [CrossRef]
- Feng, X.; Wang, Z.; Howlett-Prieto, Q.; Einhorn, N.; Causevic, S.; Reder, A.T. Vitamin D enhances responses to interferon-β in MS. Neurol. Neuroimmunol. Neuroinflamm. 2019, 6, e622. [Google Scholar] [CrossRef] [Green Version]
- Rolf, L.; Muris, A.H.; Bol, Y.; Damoiseaux, J.; Smolders, J.; Hupperts, R. Vitamin D3 supplementation in multiple sclerosis: Symptoms and biomarkers of depression. J. Neurol. Sci. 2017, 378, 30–35. [Google Scholar] [CrossRef]
- Shirazi, H.A.; Rasouli, J.; Ciric, B.; Wei, D.; Rostami, A.; Zhang, G.X. 1,25-Dihydroxyvitamin D3 suppressed experimental autoimmune encephalomyelitis through both immunomodulation and oligodendrocyte maturation. Exp. Mol. Pathol. 2017, 102, 515–521. [Google Scholar] [CrossRef]
- Chiuso-Minicucci, F.; Ishikawa, L.L.W.; Mimura, L.A.N.; De Campos Fraga-Silva, T.F.; França, T.G.D.; Zorzella-Pezavento, S.F.G.; Marques, C.; Ikoma, M.R.V.; Sartori, A. Treatment with vitamin D/MOG association suppresses experimental autoimmune encephalomyelitis. PLoS ONE 2015, 10, e0125836. [Google Scholar] [CrossRef] [PubMed]
- Hoepner, R.; Bagnoud, M.; Pistor, M.; Salmen, A.; Briner, M.; Synn, H.; Schrewe, L.; Guse, K.; Ahmadi, F.; Demir, S.; et al. Vitamin D increases glucocorticoid efficacy via inhibition of mTORC1 in experimental models of multiple sclerosis. Acta Neuropathol. 2019, 138, 443–456. [Google Scholar] [CrossRef] [Green Version]
- Ji, Z.; Fan, Z.; Zhang, Y.; Yu, R.; Yang, H.; Zhou, C.; Luo, J.; Ke, Z.J. Thiamine Deficiency Promotes T Cell Infiltration in Experimental Autoimmune Encephalomyelitis: The Involvement of CCL2. J. Immunol. 2014, 193, 2157–2167. [Google Scholar] [CrossRef]
- de la Rubia Ortí, J.E.; Cuerda-Ballester, M.; Drehmer, E.; Carrera-Juliá, S.; Motos-Muñoz, M.; Cunha-Pérez, C.; Benlloch, M.; Rodriguez, M.M.L. Vitamin B1 intake in multiple sclerosis patients and its impact on depression presence: A pilot study. Nutrients 2020, 12, 2655. [Google Scholar] [CrossRef]
- Costantini, A.; Nappo, A.; Pala, M.I.; Zappone, A. High dose thiamine improves fatigue in multiple sclerosis. BMJ Case Rep. 2013, 2013, bcr2013009144. [Google Scholar] [CrossRef] [Green Version]
- Rawji, K.S.; Young, A.M.H.; Ghosh, T.; Michaels, N.J.; Mirzaei, R.; Kappen, J.; Kolehmainen, K.L.; Alaeiilkhchi, N.; Lozinski, B.; Mishra, M.K.; et al. Niacin-mediated rejuvenation of macrophage/microglia enhances remyelination of the aging central nervous system. Acta Neuropathol. 2020, 139, 893–909. [Google Scholar] [CrossRef] [Green Version]
- Branger, P.; Parienti, J.J.; Derache, N.; Kassis, N.; Assouad, R.; Maillart, E.; Defer, G. Relapses During High-Dose Biotin Treatment in Progressive Multiple Sclerosis: A Case-Crossover and Propensity Score-Adjusted Prospective Cohort. Neurotherapeutics 2020, 17, 989–993. [Google Scholar] [CrossRef]
- Tourbah, A.; Lebrun-Frenay, C.; Edan, G.; Clanet, M.; Papeix, C.; Vukusic, S.; De Sèze, J.; Debouverie, M.; Gout, O.; Clavelou, P.; et al. MD1003 (high-dose biotin) for the treatment of progressive multiple sclerosis: A randomised, double-blind, placebo-controlled study. Mult. Scler. 2016, 22, 1719–1731. [Google Scholar] [CrossRef] [Green Version]
- Sedel, F.; Bernard, D.; Mock, D.M.; Tourbah, A. Targeting demyelination and virtual hypoxia with high-dose biotin as a treatment for progressive multiple sclerosis. Neuropharmacology 2016, 110, 644–653. [Google Scholar] [CrossRef] [Green Version]
- Bitarafan, S.; Saboor-Yaraghi, A.; Sahraian, M.-A.; Soltani, D.; Nafissi, S.; Togha, M.; Moghadam, N.B.; Roostaei, T.; Honarvar, N.M.; Harirchian, M.H. Effect of Vitamin A Supplementation on Fatigue and Depression in Multiple Sclerosis Patients: A Double-blind Placebo-controlled Clinical Trial. Iran. J. Allergy Asthma Immunol. 2016, 15, 13–19. [Google Scholar]
- Goudarzvand, M.; Javan, M.; Mirnajafi-Zadeh, J.; Mozafari, S.; Tiraihi, T. Vitamins E and D3 attenuate demyelination and potentiate remyelination processes of hippocampal formation of rats following local injection of ethidium bromide. Cell. Mol. Neurobiol. 2010, 30, 289–299. [Google Scholar] [CrossRef]
- Xue, H.; Ren, H.; Zhang, L.; Sun, X.; Wang, W.; Zhang, S.; Zhao, J.; Ming, L. Alpha-tocopherol ameliorates experimental autoimmune encephalomyelitis through the regulation of Th1 cells. Iran. J. Basic Med. Sci. 2016, 19, 861–866. [Google Scholar]
- McDonald, T.J.W.; Cervenka, M.C. Ketogenic Diets for Adult Neurological Disorders. Neurotherapeutics 2018, 15, 1018–1031. [Google Scholar] [CrossRef] [Green Version]
- Milder, J.B.; Liang, L.P.; Patel, M. Acute oxidative stress and systemic Nrf2 activation by the ketogenic diet. Neurobiol. Dis. 2010, 40, 238–244. [Google Scholar] [CrossRef] [Green Version]
- Youm, Y.H.; Nguyen, K.Y.; Grant, R.W.; Goldberg, E.L.; Bodogai, M.; Kim, D.; D’Agostino, D.; Pkanavsky, N.; Lupfer, C.; Kanneganti, T.D.; et al. The ketone metabolite β-hydroxybutyrate blocks NLRP3 inflammasome-mediated inflammatory disease. Nat. Med. 2015, 21, 263–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swidsinski, A.; Dörffel, Y.; Loening-Baucke, V.; Gille, C.; Göktas, O.; Reißhauer, A.; Neuhaus, J.; Weylandt, K.-H.; Guschin, A.; Bock, M. Reduced mass and diversity of the colonic microbiome in patients with multiple sclerosis and their improvement with ketogenic diet. Front. Microbiol. 2017, 8, 1141. [Google Scholar] [CrossRef]
- Choi, I.Y.; Piccio, L.; Childress, P.; Bollman, B.; Ghosh, A.; Brandhorst, S.; Suarez, J.; Michalsen, A.; Cross, A.H.; Morgan, T.E.; et al. A Diet Mimicking Fasting Promotes Regeneration and Reduces Autoimmunity and Multiple Sclerosis Symptoms. Cell. Rep. 2016, 15, 2136–2146. [Google Scholar] [CrossRef] [Green Version]
- Brenton, J.N.; Banwell, B.; Christina Bergqvist, A.G.; Lehner-Gulotta, D.; Gampper, L.; Leytham, E.; Coleman, R.; Goldman, M.D. Pilot study of a ketogenic diet in relapsing-remitting MS. Neurol. Neuroimmunol. Neuroinflamm. 2019, 6, e565. [Google Scholar] [CrossRef] [Green Version]
- Wahls, T.L.; Chenard, C.A.; Snetselaar, L.G. Review of two popular eating plans within the multiple sclerosis community: Low saturated fat and modified paleolithic. Nutrients 2019, 11, 352. [Google Scholar] [CrossRef] [Green Version]
- Langley, M.R.; Triplet, E.M.; Scarisbrick, I.A. Dietary influence on central nervous system myelin production, injury, and regeneration. Biochim. Biophys. Acta Mol. Bas. Dis. 2020, 1866, 165779. [Google Scholar] [CrossRef]
- Irish, A.; Erickson, C.; Wahls, T.; Snetselaar, L.; Darling, W. Randomized control trial evaluation of a modified Paleolithic dietary intervention in the treatment of relapsing-remitting multiple sclerosis: A pilot study. Degener. Neurol. Neuromuscul. Dis. 2017, 7, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masullo, L.; Papas, M.A.; Cotugna, N.; Baker, S.; Mahoney, L.; Trabulsi, J. Complementary and Alternative Medicine Use and Nutrient Intake Among Individuals with Multiple Sclerosis in the United States. J. Community Health 2015, 40, 153–160. [Google Scholar] [CrossRef]
- Thomsen, H.L.; Jessen, E.B.; Passali, M.; Frederiksen, J.L. The role of gluten in multiple sclerosis: A systematic review. Mult. Scler. Rel. Dis. 2019, 27, 156–163. [Google Scholar] [CrossRef]
- Passali, M.; Josefsen, K.; Frederiksen, J.L.; Antvorskov, J.C. Current evidence on the efficacy of gluten-free diets in multiple sclerosis, psoriasis, type 1 diabetes and autoimmune thyroid diseases. Nutrients 2020, 12, 2316. [Google Scholar] [CrossRef]
- Rodrigo, L.; Hernandez-Lahoz, C.; Fuentes, D.; Mauri, G.; Alvarez, N.; Vega, J.; Gonzalez, S. Randomised Clinical Trial Comparing the Efficacy of a Gluten-Free Diet Versus A Regular Diet in A Series of Relapsing-Remitting Multiple Sclerosis Patients. Int. J. Neurol. Neurother. 2014, 1, 012. [Google Scholar] [CrossRef]
- Lee, J.E.; Bisht, B.; Hall, M.J.; Rubenstein, L.M.; Louison, R.; Klein, D.T.; Wahks, T.L. A Multimodal, Nonpharmacologic Intervention Improves Mood and Cognitive Function in People with Multiple Sclerosis. J. Am. Coll. Nutr. 2017, 36, 150–168. [Google Scholar] [CrossRef] [Green Version]
- Bach-Faig, A.; Berry, E.M.; Lairon, D.; Reguant, J.; Trichopoulou, A.; Dernini, S.; Medina, F.X.; Battino, M.; Belahsen, R.; Miranda, G.; et al. Mediterranean diet pyramid today. Science and cultural updates. Public Health Nutr. 2011, 12A, 2274–2284. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Lacoba, R.; Pardo-Garcia, I.; Amo-Saus, E.; Escribano-Sotos, F. Mediterranean diet and health outcomes: A systematic meta-review. Eur. J Public Health 2018, 28, 955–961. [Google Scholar] [CrossRef] [Green Version]
- Esposito, S.; Bonavita, S.; Sparaco, M.; Gallo, A.; Tedeschi, G. The role of diet in multiple sclerosis: A review. Nutritional neuroscience. Nutr. Neurosci. 2018, 21, 377–390. [Google Scholar] [CrossRef]
- Sedaghat, F.; Jessri, M.; Behrooz, M.; Mirghotbi, M.; Rashidkhani, B. Mediterranean diet adherence and risk of multiple sclerosis: A case-control study. Asia Pac. J. Clin. Nutr. 2016, 25, 377–384. [Google Scholar] [CrossRef]
- Esposito, S.; Sparaco, M.; Maniscalco, G.T.; Signoriello, E.; Lanzillo, R.; Russo, C.; Carmisciano, L.; Cepparulo, S.; Lavorgna, L.; Gallo, A.; et al. Lifestyle and Mediterranean diet adherence in a cohort of Southern Italian patients with Multiple Sclerosis. Mult. Scler. Rel. Dis. 2021, 47, 102636. [Google Scholar] [CrossRef]
- Giacometti, J.; Grubić-Kezele, T. Olive Leaf Polyphenols Attenuate the Clinical Course of Experimental Autoimmune Encephalomyelitis and Provide Neuroprotection by Reducing Oxidative Stress, Regulating Microglia and SIRT1, and Preserving Myelin Integrity. Oxid. Med. Cell Longev. 2020, 2020, 6125638. [Google Scholar] [CrossRef]
- Swank, R.L. Multiple sclerosis; a correlation of its incidence with dietary fat. Am. J. Med. Sci. 1950, 220, 421–430. [Google Scholar] [CrossRef]
- Swank, R.L. Treatment of multiple sclerosis with low-fat diet. AMA Arch. Neurol. Psychiatry 1953, 69, 91–103. [Google Scholar] [CrossRef]
- Wahls, T.; Scott, M.O.; Alshare, Z.; Rubenstein, L.; Darling, W.; Carr, L.; Smith, K.; Chenard, C.A.; LaRocca, N.; Snetselaar, L. Dietary approaches to treat MS-related fatigue: Comparing the modified Paleolithic (Wahls Elimination) and low saturated fat (Swank) diets on perceived fatigue in persons with relapsing-remitting multiple sclerosis: Study protocol for a randomized controlled trial. Trials 2018, 19, 309. [Google Scholar] [CrossRef]
- Swank, R.L.; Goodwin, J. Review of MS patient survival on a Swank low saturated fat diet. Nutrition 2003, 19, 161–162. [Google Scholar] [CrossRef]
- McDougall, J.; Thomas, L.E.; McDougall, C.; Moloney, G.; Saul, B.; Finnell, J.S.; Rischardson, K.; Petersen, K.M. Effects of 7 days on an ad libitum low-fat vegan diet: The McDougall Program cohort. Nutr. J. 2014, 13, 99. [Google Scholar] [CrossRef] [Green Version]
- Yadav, V.; Marracci, G.; Kim, E.; Spain, R.; Cameron, M.; Overs, S.; Riddehough, A.; Li, D.K.B.; McDougall, J.; Lovera, J.; et al. Low-fat, plant-based diet in multiple sclerosis: A randomized controlled trial. Mult. Scler. Rel. Dis. 2016, 9, 80–90. [Google Scholar] [CrossRef] [Green Version]
- Schüpbach, R.; Wegmüller, R.; Berguerand, C.; Bui, M.; Herter-Aeberli, I. Micronutrient status and intake in omnivores, vegetarians and vegans in Switzerland. Eur. J. Nutr. 2017, 56, 283–293. [Google Scholar] [CrossRef]
- Roman, S.N.; Fitzgerald, K.C.; Beier, M.; Mowry, E.M. Safety and feasibility of various fasting-mimicking diets among people with multiple sclerosis. Mult. Scler. Rel. Dis. 2020, 42, 102149. [Google Scholar] [CrossRef]
- Mattson, M.P.; Longo, V.D.; Harvie, M. Impact of intermittent fasting on health and disease processes. Ageing Res. Rev. 2017, 39, 46–58. [Google Scholar] [CrossRef]
- Spaulding, C.C.; Walford, R.L.; Effros, R.B. Calorie restriction inhibits the age-related dysregulation of the cytokines TNF-α and IL-6 in C3B10RF1 mice. Mech. Ageing Dev. 1997, 1–3, 87–94. [Google Scholar] [CrossRef]
- Mojaverrostami, S.; Pasbakhsh, P.; Madadi, S.; Nekoonam, S.; Zarini, D.; Noori, L.; Shiri, E.; Salama, M.; Zibara, K.; Kashani, I.R. Calorie restriction promotes remyelination in a Cuprizone-Induced demyelination mouse model of multiple sclerosis. Metab. Brain Dis. 2020, 35, 1211–1224. [Google Scholar] [CrossRef]
- Esquifino, A.I.; Cano, P.; Jimenez-Ortega, V.; Fernández-Mateos, M.P.; Cardinali, D.P. Immune response after experimental allergic encephalomyelitis in rats subjected to calorie restriction. J. Neuroinflamm. 2007, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Bai, M.; Wang, Y.; Han, R.; Xu, L.; Huang, M.; Zhao, J.; Lin, Y.; Song, S.; Chen, Y. Intermittent caloric restriction with a modified fasting-mimicking diet ameliorates autoimmunity and promotes recovery in a mouse model of multiple sclerosis. J. Nutr. Biochem. 2021, 87, 108493. [Google Scholar] [CrossRef]
- Cignarella, F.; Cantoni, C.; Ghezzi, L.; Salter, A.; Dorsett, Y.; Chen, L.; Philips, D.; Weinstock, G.M.; Fontana, L.; Cross, A.H.; et al. Intermittent Fasting Confers Protection in CNS Autoimmunity by Altering the Gut Microbiota. Cell. Metab. 2018, 27, 1222–1235. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, K.C.; Vizthum, D.; Henry-Barron, B.; Schweitzer, A.; Cassard, S.D.; Kossoff, E.; Hartman, A.L.; Kapogiannis, D.; Sullivan, P.; Baer, D.J.; et al. Effect of intermittent vs. daily calorie restriction on changes in weight and patient-reported outcomes in people with multiple sclerosis. Mult. Scler. Rel. Dis. 2018, 23, 33–39. [Google Scholar] [CrossRef]
- Bock, M.; Karber, M.; Kuhn, H. Ketogenic diets attenuate cyclooxygenase and lipoxygenase gene expression in multiple sclerosis. EBioMedicine 2018, 36, 293–303. [Google Scholar] [CrossRef] [Green Version]
- Most, J.; Tosti, V.; Redman, L.M.; Fontana, L. Calorie restriction in humans: An update. Ageing Res. Rev. 2017, 39, 36–45. [Google Scholar] [CrossRef]
- Marrie, R.A.; Cohen, J.; Stuve, O.; Trojano, M.; Sørensen, P.S.; Reingold, S.; Cutter, G.; Reider, N. A systematic review of the incidence and prevalence of comorbidity in multiple sclerosis: Overview. Mult. Scler. 2015, 21, 263–281. [Google Scholar] [CrossRef]
- Haghikia, A.; Jörg, S.; Duscha, A.; Berg, J.; Manzel, A.; Waschbisch, A.; Hammer, A.; Kee, D.H.; May, C.; Wilck, N.; et al. Dietary Fatty Acids Directly Impact Central Nervous System Autoimmunity via the Small Intestine. Immunity 2015, 43, 817–829. [Google Scholar] [CrossRef] [Green Version]
- Timmermans, S.; Bogie, J.F.J.; Vanmierlo, T.; Lütjohann, D.; Stinissen, P.; Hellings, N.; Hendriks, J.J.A. High fat diet exacerbates neuroinflammation in an animal model of multiple sclerosis by activation of the renin angiotensin system. J. Neuroimmune Pharmacol. 2014, 9, 209–217. [Google Scholar] [CrossRef]
- Azary, S.; Schreiner, T.; Graves, J.; Waldman, A.; Belman, A.; Weinstock-Guttman, B.; Aaen, G.; Tillema, J.M.; Mar, S.; Hart, J.; et al. Contribution of dietary intake to relapse rate in early paediatric multiple sclerosis. J. Neurol. Neurosurg. Psych. 2018, 89, 28–33. [Google Scholar] [CrossRef]
- Black, L.J.; Rowley, C.; Sherriff, J.; Pereira, G.; Ponsonby, A.L.; Lucas, R.M. A healthy dietary pattern associates with a lower risk of a first clinical diagnosis of central nervous system demyelination. Mult. Scler. 2019, 25, 1514–1525. [Google Scholar] [CrossRef]
- Albrechtsen, M.T.; Langeskov-Christensen, M.; Jørgensen, M.L.K.; Dalgas, U.; Hansen, M. Is diet associated with physical capacity and fatigue in persons with multiple sclerosis? –Results from a pilot study. Mult. Scler. Rel. Dis. 2020, 40, 101921. [Google Scholar] [CrossRef]
- Langdon, D.W.; Amato, M.P.; Boringa, J.; Brochet, B.; Foley, F.; Fredrikson, S.; Hamalainen, P.; Hartung, H.P.; Krupp, L.; Penner, J.K.; et al. Recommendations for a brief international cognitive assessment for multiple sclerosis (BICAMS). Mult. Scler. 2012, 18, 891–898. [Google Scholar] [CrossRef] [Green Version]
- Chiaravalloti, N.D.; DeLuca, J. Cognitive impairment in multiple sclerosis. Lancet Neurol. 2008, 7, 1139–1151. [Google Scholar] [CrossRef]
- Miller, E.; Morel, A.; Redlicka, J.; Miller, I.; Saluk, J. Pharmacological and non-pharmacological therapies of cognitive impairment in multiple sclerosis. Curr. Neuropharmacol. 2018, 16, 475–483. [Google Scholar] [CrossRef]
- Rademacher, A.; Joisten, N.; Proschinger, S.; Bloch, W.; Gonzenbach, R.; Kool, J.; Langdon, D.; Bansi, J.; Zimmer, P. Cognitive Impairment Impacts. Exercise Effects on Cognition in Multiple Sclerosis. Front. Neurol. 2021, 11, 619500. [Google Scholar] [CrossRef]
- Benedict, R.H.B.; DeLuca, J.; Phillips, G.; LaRocca, N.; Hudson, L.D.; Rudick, R. Validity of the Symbol Digit Modalities Test as a cognitive performance outcome measure for multiple sclerosis. Mult. Scler. 2017, 23, 721–733. [Google Scholar] [CrossRef]
- Mayo, C.D.; Harrison, L.; Attwell-Pope, K.; Stuart-Hill, L.; Gawryluk, J.R. A pilot study of the impact of an exercise intervention on brain structure, cognition, and psychosocial symptoms in individuals with relapsing-remitting multiple sclerosis. Pilot Feasibility Stud. 2021, 7, 65. [Google Scholar] [CrossRef]
- Langeskov-Christensen, M.; Hvid, L.G.; Jensen, H.B.; Nielsen, H.H.; Petersen, T.; Stenager, E.; Hämäläinen, P.; Dalgas, U. Efficacy of high-intensity aerobic exercise on cognitive performance in people with multiple sclerosis: A randomized controlled trial. Mult. Scler. 2020, 27, 1585–1596. [Google Scholar] [CrossRef]
- Oken, B.S.; Kishiyama, S.; Zajdel, D.; Bourdette, D.; Carlsen, J.; Haas, M.; Hugos, C.; Kraemer, D.F.; Lawrence, J.; Mass, M. Randomized controlled trial of yoga and exercise in multiple sclerosis. Neurology 2004, 62, 2058–2064. [Google Scholar] [CrossRef]
- Baquet, L.; Hasselmann, H.; Patra, S.; Stellmann, J.P.; Vettorazzi, E.; Engel, A.K.; Rosenkranz, S.C.; Poettgen, J.; Gold, S.M.; Schulz, K.H.; et al. Short-term interval aerobic exercise training does not improve memory functioning in relapsing-remitting multiple sclerosis-a randomized controlled trial. Peer J. 2018, 6, e6037. [Google Scholar] [CrossRef] [Green Version]
- Kucuk, F.; Kara, B.; Poyraz, E.C.; Idiman, E. Improvements in cognition, quality of life, and physical performance with clinical Pilates in multiple sclerosis: A randomized controlled trial. J. Phys. Ther. Sci. 2016, 28, 761–768. [Google Scholar] [CrossRef] [Green Version]
- Gharakhanlou, R.; Wesselmann, L.; Rademacher, A.; Lampit, A.; Negaresh, R.; Kaviani, M.; Oberste, M.; Motl, R.W.; Sandroff, B.M.; Bansi, J.; et al. Exercise training and cognitive performance in persons with multiple sclerosis: A systematic review and multilevel meta-analysis of clinical trials. Mult. Scler. 2020, 11, 1352458520917935. [Google Scholar] [CrossRef]
- Dalgas, U.; Stenager, E.; Ingemann-Hansen, T. Multiple sclerosis and physical exercise: Recommendations for the application of resistance-, endurance-and combined training. Mult. Scler. 2008, 14, 35–53. [Google Scholar] [CrossRef]
- Ozkul, C.; Guclu-Gunduz, A.; Irkec, C.; Fidan, I.; Aydin, Y.; Ozkan, T.; Yazici, G. Effect of combined exercise training on serum brain-derived neurotrophic factor, suppressors of cytokine signaling 1 and 3 in patients with multiple sclerosis. J. Neuroimmunol. 2018, 316, 121–129. [Google Scholar] [CrossRef]
- Ozkul, C.; Guclu-Gunduz, A.; Eldemir, K.; Apaydin, Y.; Yazici, G.; Irkec, C. Combined exercise training improves cognitive functions in multiple sclerosis patients with cognitive impairment: A single-blinded randomized controlled trial. Mult. Scler. Rel. Dis. 2020, 45, 102419. [Google Scholar] [CrossRef]
- Pons, V.; Rivest, S. Beneficial Roles of Microglia and Growth Factors in MS, a Brief Review. Front. Cell. Neurosci. 2020, 14, 284. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, D.; Oliveira-Lima, O.C.; Vitarelli da Silva, T.; Faraco, C.C.F.; Ribeiro Lete, H.; Juliano, M.A.; Moreira dos Santos, D.; Bethea, J.R.; Brambilla, R.; Orian, J.M.; et al. Differential brain and spinal cord cytokine and BDNF levels in experimental autoimmune encephalomyelitis are modulated by prior and regular exercise. J. Neuroimmunol. 2013, 264, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Li, Z.; Wang, Y.; Xue, X.; Ma, W.; Zhang, Y.; Wang, Y. Effects of moderate- versus high- intensity swimming training on inflammatory and CD4+ T cell subset profiles in experimental autoimmune encephalomyelitis mice. J. Neuroimmunol. 2019, 328, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Pedard, M.; Cefis, M.; Ennequin, G.; Quirié, A.; Garnier, P.; Prigent-Tessier, A.; Pernet, N.; Marie, C. Brain-derived Neurotrophic Factor Pathway after Downhill and Uphill Training in Rats. Medicine and science in sports and exercise. Med. Sci. Sports Exerc. 2019, 51, 27–34. [Google Scholar] [CrossRef]
- Baranowski, B.J.; MacPherson, R. Acute exercise induced BDNF-TrkB signalling is intact in the prefrontal cortex of obese, glucose-intolerant male mice. Applied physiology, nutrition, and metabolism. Appl. Physiol. Nutr. Metab. 2018, 43, 1083–1089. [Google Scholar] [CrossRef] [PubMed]
- Souza, P.S.; Goncalves, E.D.; Pedroso, G.S.; Farias, H.R.; Junqueira, S.C.; Marcon, R.; Tuon, T.; Cola, M.; Silveira, P.C.L.; Santos, A.R.; et al. Physical exercise attenuates experimental autoimmune encephalomyelitis by inhibiting peripheral immune response and blood-brain barrier disruption. Mol. Neurobiol. 2017, 54, 4723–4737. [Google Scholar] [CrossRef] [PubMed]
- Mandolesi, G.; Buttitta, S.; Fresegna, D.; De Vito, F.; Rizzo, F.R.; Musella, A.; Guadalupi, L.; Vanni, V.; Stampanoni Bassi, M.; Buttari, F.; et al. Voluntary running wheel attenuates motor deterioration and brain damage in cuprizone-induced demyelination. Neurobiol. Dis. 2019, 129, 102–117. [Google Scholar] [CrossRef]
- Alvarenga-Filho, H.; Sacramento, P.M.; Ferreira, T.B.; Hygino, J.; Abreu, J.; Carvalho, S.R.; Wing, A.C.; Alvarenga, R.; Bento, C. Combined exercise training reduces fatigue and modulates the cytokine profile of T-cells from multiple sclerosis patients in response to neuromediators. J. Neuroimmunol. 2016, 293, 91–99. [Google Scholar] [CrossRef]
- Lozinski, B.M.; Yong, V.W. Exercise and the brain in multiple sclerosis. Mult. Scler. 2020, 30, 1352458520969099. [Google Scholar] [CrossRef] [PubMed]
- Bonfiglio, T.; Olivero, G.; Vergassola, M.; Di Cesare Mannelli, L.; Pacini, A.; Iannuzzi, F.; Summa, M.; Bertorelli, L.; Feligioni, M.; Ghelardini, C.; et al. Environmental training is beneficial to clinical symptoms and cortical presynaptic defects in mice suffering from experimental autoimmune encephalomyelitis. Neuropharmacology 2019, 145, 75–86. [Google Scholar] [CrossRef]
- Shahidi, S.H.; Kordi, M.R.; Rajabi, H.; Malm, C.; Shah, F.; Quchan, A.S.K. Exercise modulates the levels of growth inhibitor genes before and after multiple sclerosis. J. Neuroimmunol. 2020, 341, 577172. [Google Scholar] [CrossRef]
- Magalon, K.; Cantarella, C.; Monti, G.; Cayre, M.; Durbec, P. Enriched environment promotes adult neural progenitor cell mobilization in mouse demyelination models. Eur. J. Neurosci. 2007, 25, 761–771. [Google Scholar] [CrossRef]
- Jensen, S.K.; Michaels, N.J.; Ilyntskyy, S.; Keough, G.H.; Kovalchuk, O.; Yong, V.W. Multimodal enhancement of remyelination by exercise with a pivotal role for oligodendroglial PGC1α. Cell Rep. 2018, 24, 3167–3179. [Google Scholar] [CrossRef] [Green Version]
- Dalgas, U.; Langeskov-Christensen, M.; Stenager, E.; Riemenschneider, M.; Hvid, L.G. Exercise as Medicine in Multiple Sclerosis—Time for a Paradigm Shift: Preventive, Symptomatic, and Disease-Modifying Aspects and Perspectives. Curr. Neurol. Neurosci. Rep. 2019, 19, 88. [Google Scholar] [CrossRef]
- Pryor, W.P.; Freeman, K.G.; Larson, R.D.; Edwards, G.L.; White, L.J. Chronic Exercise Confers Neuroprotection in Experimental Autoimmune Encephalomyelitis. J. Neurosci. Res. 2015, 93, 697–706. [Google Scholar] [CrossRef]
- Rossi, S.; Furlan, R.; De Chiara, V.; Musella, A.; Lo Giudice, T.; Mataluni, G.; Cavasinni, F.; Cantarella, C.; Bernardi, G.; Muzio, L.; et al. Exercise attenuates the clinical, synaptic, and dendritic abnormalities of experimental autoimmune encephalomyelitis. Neurobiol. Dis. 2009, 36, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.I.; White, L.J. Effect of 10-day forced treadmill training on neurotrophic factors in experimental autoimmune encephalomyelitis. Appl. Physiol. Nutr. Metab. 2013, 38, 194–199. [Google Scholar] [CrossRef]
- Joisten, N.; Rademacher, A.; Warnke, C.; Proschinger, S.; Schenk, A.; Walzik, D.; Knoop, A.; Thevis, M.; Steffen, S.; Gonzenbach, R.; et al. Exercise Diminishes Plasma Neurofilament Light Chain and Reroutes the Kynurenine Pathway in Multiple Sclerosis. Neurol. Neuroimmunol. Neuroinflamm. 2021, 8, e982. [Google Scholar] [CrossRef] [PubMed]
- Faramarzi, M.; Banitalebi, E.; Raisi, Z.; Samieyan, M.; Saberi, Z.; Ghahfarrokhi, M.M.; Negaresh, R.; Motl, R.W. Effect of combined exercise training on pentraxins and pro- inflammatory cytokines in people with multiple sclerosis as a function of disability status. Cytokine 2020, 134, 155196. [Google Scholar] [CrossRef]
- Banitalebi, E.; Ghahfarrokhi, M.M.; Negaresh, R.; Kazemi, A.; Faramarzi, M.; Motl, R.W.; Zimmer, P. Exercise improves neurotrophins in multiple sclerosis independent of disability status. Mult. Scler. Rel. Dis. 2020, 43, 102143. [Google Scholar] [CrossRef] [PubMed]
- Gravesteijn, A.S.; Beckerman, H.; de Jong, B.A.; Hulst, H.E.; de Groot, V. Neuroprotective effects of exercise in people with progressive multiple sclerosis (Exercise PRO-MS): Study protocol of a phase II trial. BMC Neurol. 2020, 20, 177. [Google Scholar] [CrossRef]
- Proschinger, S.; Joisten, N.K.; Rademacker, A.; Schlagheck, M.L.; Walzik, D.; Metcalfe, A.J.; Oberste, M.; Warnke, C.; Bloch, W.; Schenk, A.; et al. Influence of combined functional resistance and endurance exercise over 12 weeks on matrix metalloproteinase-2 serum concentration in persons with relapsing-remitting multiple sclerosis—A community-based randomized controlled trial. BMC Neurol. 2019, 19, 314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves, A.R.; Devasahayam, A.J.; Kelly, l.P.; Pretty, R.W.; Ployhman, M. Exercise-Induced Brain Excitability Changes in Progressive Multiple Sclerosis. J. Neurol. Phys. Ther. 2020, 44, 132–144. [Google Scholar] [CrossRef]
- Stellmann, J.P.; Maarouf, A.; Schulz, K.H.; Baquet, L.; Pottgen, J.; Patra, S.; Penner, J.K.; Gelliβen, S.; Ketels, G.; Besson, P.; et al. Aerobic Exercise Induces Functional and Structural Reorganization of CNS Networks in Multiple Sclerosis: A Randomized Controlled Trial. Front. Hum. Neurosci. 2020, 14, 255. [Google Scholar] [CrossRef]
- Savsek, L.; Stergar, T.; Strojnik, V.; Ihan, I.; Koren, A.; Spliclin, Z.; Jazbec, S.S. Impact of aerobic exercise on clinical and magnetic resonance imaging biomarkers in persons with Multiple Sclerosis: An exploratory randomized controlled trial. J. Rehabil. Med. 2021, 53, jrm00178. [Google Scholar] [CrossRef] [PubMed]
- Kjølhede, T.; Siemonsen, S.; Wenzel, D.; Stellmann, J.P.; Ringgaard, S.; Pedersen, B.G.; Stenager, E.; Petersen, T.; Vissing, K.; Heesen, C.; et al. Can resistance training impact MRI outcomes in relapsing-remitting multiple sclerosis? Mult. Scler. 2018, 24, 1356–1365. [Google Scholar] [CrossRef] [PubMed]
- Langeskov-Christensen, M.; Hvid, L.G.; Nygaard, M.K.E.; Ringgaard, H.B.; Jensen, H.B.; Nielsen, H.H.; Petersen, T.; Stenager, E.; Eskildsen, S.F.; Dalgas, U. Efficacy of High-Intensity Aerobic Exercise on Brain MRI Measures in Multiple Sclerosis. Neurology 2021, 96, e203–e213. [Google Scholar] [CrossRef]
- Riemenschneider, M.; Hvid, L.G.; Ringgaard, S.; Nygaard, M.K.E.; Eskildsen, S.F.; Petersen, T.; Stenager, E.; Dalgas, U. Study protocol: Randomised controlled trial evaluating exercise therapy as a supplemental treatment strategy in early multiple sclerosis: The Early Multiple Sclerosis Exercise Study (EMSES). BMJ Open 2021, 11, e043699. [Google Scholar] [CrossRef] [PubMed]
- Kalb, R.; Brown, T.R.; Coote, S.; Costello, K.; Dalgas, U.; Garmon, E.; Giesser, B.; Halper, J.; Karpatkin, H.; Keller, J.; et al. Exercise and lifestyle physical activity recommendations for people with multiple sclerosis throughout the disease course. Mult. Scler. 2020, 26, 1459–1469. [Google Scholar] [CrossRef] [PubMed]
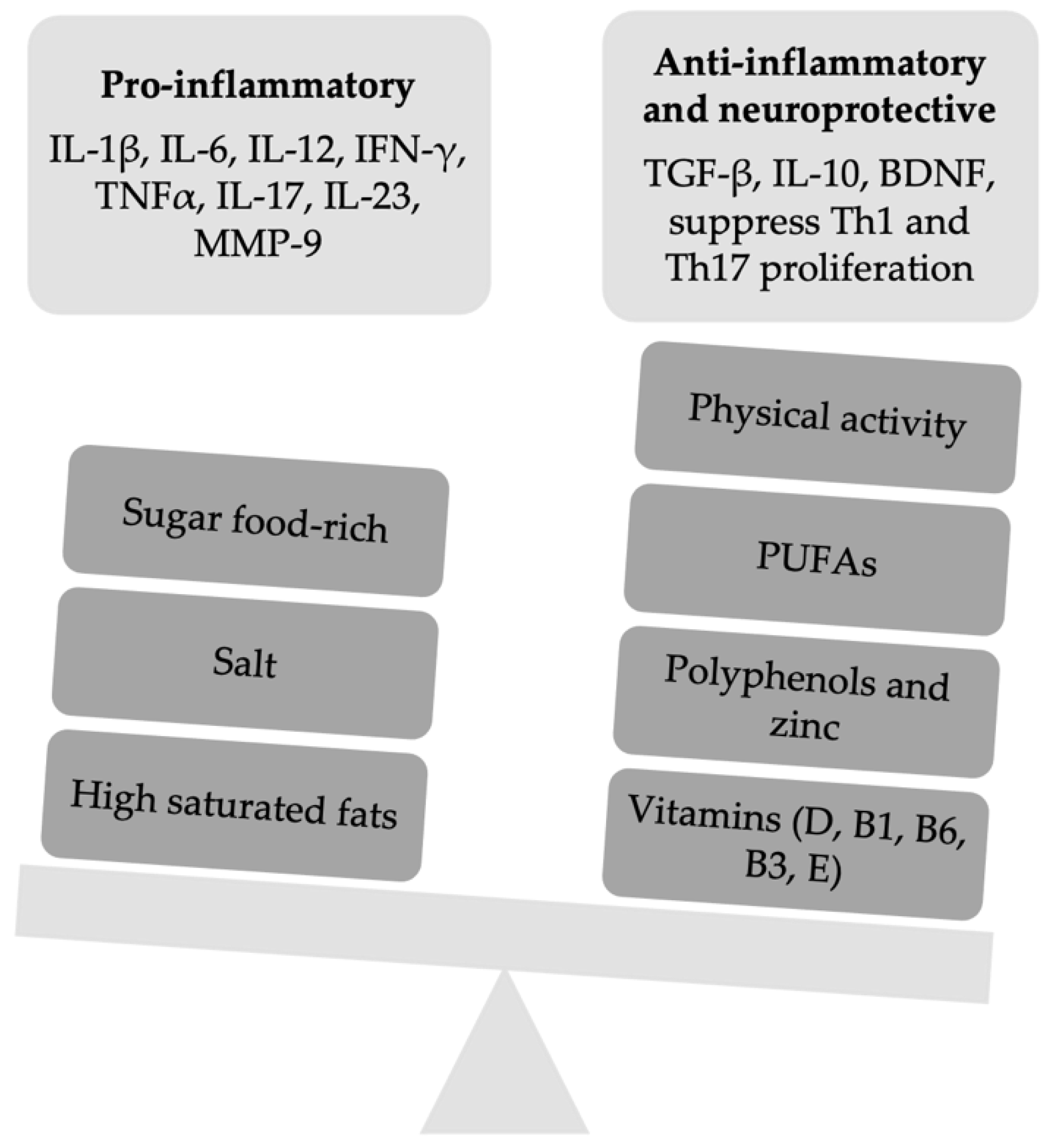

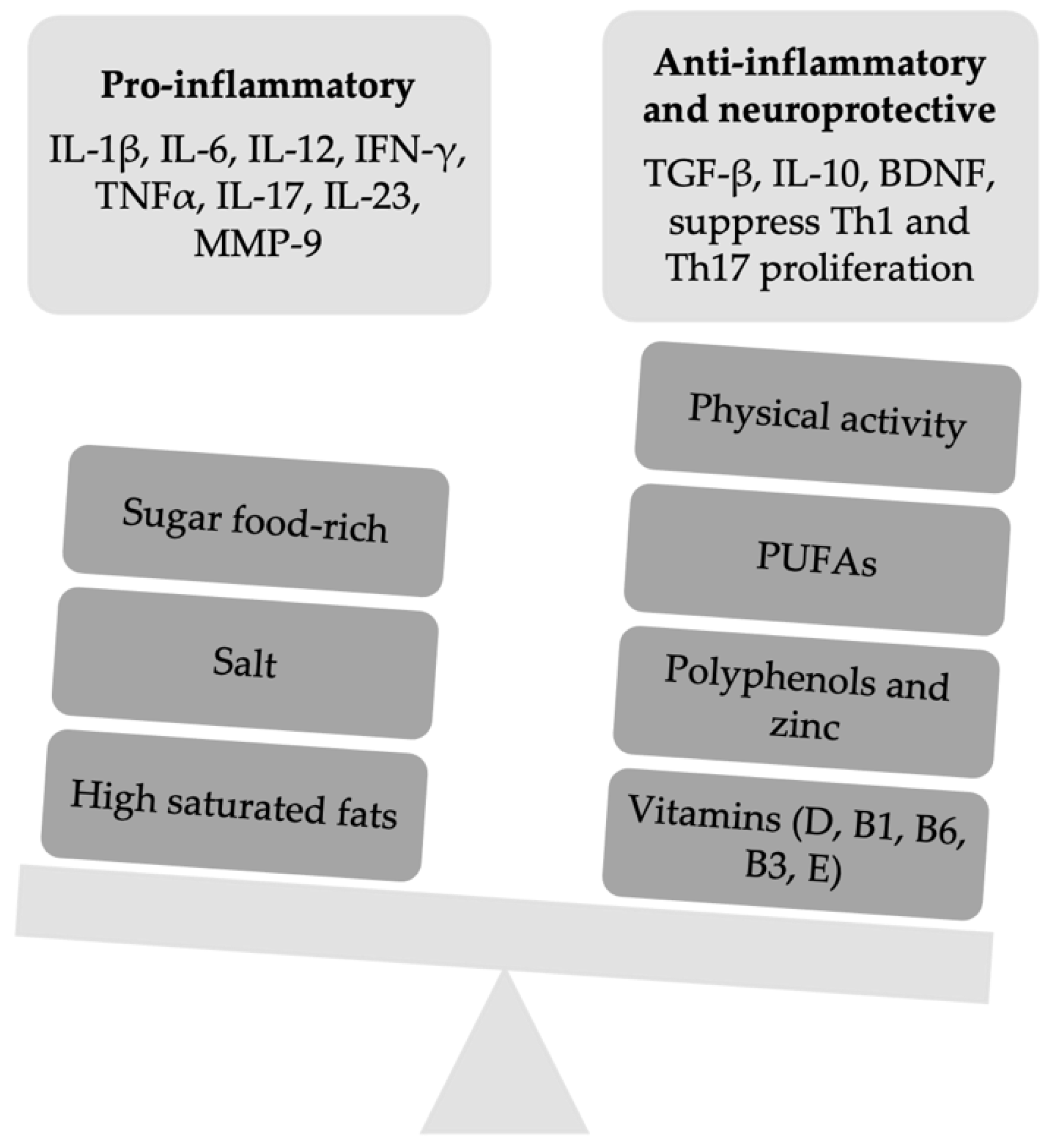{kind=link}
| Food-Derived Factor | Molecular Mechanism Proposed | Benefits on PwMS | References |
|---|---|---|---|
| PUFAs | EPA improves EAE by reducing IFN-γ and IL-17 production, specifically in CNS lesions infiltrated by CD4 T cells, and it enhances PPARs. DHA reduces the expression of transcription factors in DCs for Th1 and Th17 differentiation as well as the mortality in mice with EAE | Omega-3 improve the metabolic profile, EDSS score, and reduces inflammatory markers such as MMP-9, TNF α, IL-1β, and IL-6 | [14,15,16,17] |
| Food-Derived Factor | Molecular Mechanism Proposed | Benefits on PwMS | References |
|---|---|---|---|
| Luteolin | Suppress the migration of PMBCs in animal model and prevent disease relapses by influencing the monocytic GTPase activity. | Reduction of EAE severity with protective effect on chronic EAE. Dose-dependent anti-inflammatory effect by reducing IL-1β, TNF-α, and MMP-9. The therapy with IFN-β has a summation effect with that of luteolin. | [19,20] |
| Baicalin | Reduces oxidative stress in myelin-producing cells through the Nrf2/HO-1 signaling pathway Reduces CNS inflammation by suppressing IL-17, IFN-γ, GM-CSF, IL1-β, IL-6, IL-1, and IL-23 | Improvement in the clinical score. Stopping baicalin results in recurrence of symptoms 7–8 days after the treatment. | [21,22] |
| Curcumin | Improves EAE through inhibition of the STAT3-phosphorylation and reduction of IL-12 production from microglial cells In PMBCs, suppressed IFN-γ and increased IFN-β Reduced production of MMP-9 in human astrocytes. | A dose of 80 mg daily for six months in pwRRMS reduced pro-inflammatory pathways compared to control without improving EDSS score. It might potentially reduce BBB permeability ameliorating MS clinical manifestation. | [23,24,25,26,27] |
| Resveratrol | In EAE, decreases the production of pro-inflammatory cytokines (e.g., TNF-α, IFN-γ, IL-2, IL-9, IL-12, and IL-17), induces T regulatory cells, and decreases in a dose-depended manner the BBB disruption by reducing the loss of TJ components (e.g., claudin-5, occludin). Contrasts EAE development through suppression of the miRNA-124/SK1 pathway. Neuroprotection during optic neuritis in an EAE model by reducing axonal loss. | Although 150 mg daily resveratrol supplementation in association with vitamin D in pwMS showed a reduction in serum levels of MMP-9, it did not demonstrate an improvement in signs and symptoms | [28,29,30,31,32,33] |
| Epigallocatechin gallate | Modulates GABAergic pathway. Improves EAE severity by reducing Th1 and Th17 cells. | It reduces IL-6, improving anxiety and depression. More in men than women, it improves the energy metabolism during exercise without any clinical and radiological effects | [34,35,36,37,38] |
| Cocoa | Increases cerebral blood flow. Antioxidant properties alleviate lipid peroxidation and axon damage | Mild reduction in fatigue and fatigability | [39] |
| Caffeine | In EAE after immunization with 10–30 mg/kg daily, caffeine reduces inflammatory cells in the spinal cord and neurological as well as IFN-γ production and disease severity. | An estimated amount of 250 to 300 mg caffeine intake (2–3 cups) improves fatigue and mental capacity, especially in patients with an EDSS higher than 0 but lower than 4 | [40,41,42] |
| Alcohol | In EAE, male-specific disease remission induced potentially via gut microbiota modulation. | Not available | [43] |
| Food-Derived Factor | Molecular Mechanism Proposed | Benefits on PwMS | References |
|---|---|---|---|
| Zinc | Medium dose (1.5 mg/kg) improves clinical score of EAE, suppressing T cell activation and pro-inflammatory cytokines, whereas high doses (6 mg/kg) lead to clinical worsening. | Zinc supplementation (220 mg zinc sulfate daily) is able to improve depression in MS patients without providing any benefit for movement disorders. | [44,45] |
| Salt | High-sodium intake might inhibit the functions of Treg cells, promoting the shift to a Th1-like phenotype. High-salt intake induces EAE exacerbation through changes in microbiota and enhanced Th17 cells differentiation. High-salt intake might enhance corticosterone serum levels, allowing the expression of TJ molecules suppressing CNS autoimmunity. | Some authors reported that salt intake did not have a relationship with the course or brain MRI activity of MS, whereas other ones reported that high-sodium intake is associated with increased MS relapses and brain MRI activity. However, in pediatric onset MS, no associations between salt intake and pediatric onset MS risk or time to relapses were found. | [46,47,48,49,50,51,52] |
| Food-Derived Factor | Molecular Mechanism Proposed | Benefits on PwMS | References |
|---|---|---|---|
| Vitamin D | In the EAE model, vitamin D3 suppresses Th17 and Th1 differentiation, enhancing the percentage of Treg cells. Reduces demyelination, incidence, and clinical score of EAE Increases the effectiveness of steroid therapy through mTORC1 inhibition.In EAE, high doses of vitamin D leading to hypercalcemia might promote T cells proliferation with clinical exacerbation | Vitamin D3 supplementation does not improve the depressive symptoms in MS. It enhances the response to some treatments for MS such as interferon-β | [60,61,62,63,64,65,66,67,68,69] |
| Vitamin B1 | In EAE, vitamin B1 deficiency causes Th1 and Th17 spinal cord infiltration | PwMS with reduced vitamin B1 levels and depression might benefit from supplementation (potential role in fatigue severity improvement) | [70,71,72] |
| Vitamin B3 | Potentiates beneficial effects on monocytes and macrophages in promoting remyelination in CNS aging | No data available | [73] |
| Vitamin B7 | It promotes myelin synthesis, and it reduces axon hypoxia in MS | Some authors reported benefits in worsening EDSS score in patients with progressive MS High doses might increase the risk of relapses | [74,75,76] |
| Vitamin A | Antioxidant | It might improve fatigue and depression during interferon therapy | [77] |
| Vitamin E | Hippocampal remyelination, and it suppresses INF-γ production and delays EAE progression | Not available | [78,79] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fanara, S.; Aprile, M.; Iacono, S.; Schirò, G.; Bianchi, A.; Brighina, F.; Dominguez, L.J.; Ragonese, P.; Salemi, G. The Role of Nutritional Lifestyle and Physical Activity in Multiple Sclerosis Pathogenesis and Management: A Narrative Review. Nutrients 2021, 13, 3774. https://doi.org/10.3390/nu13113774
Fanara S, Aprile M, Iacono S, Schirò G, Bianchi A, Brighina F, Dominguez LJ, Ragonese P, Salemi G. The Role of Nutritional Lifestyle and Physical Activity in Multiple Sclerosis Pathogenesis and Management: A Narrative Review. Nutrients. 2021; 13(11):3774. https://doi.org/10.3390/nu13113774
Chicago/Turabian StyleFanara, Salvatore, Maria Aprile, Salvatore Iacono, Giuseppe Schirò, Alessia Bianchi, Filippo Brighina, Ligia Juliana Dominguez, Paolo Ragonese, and Giuseppe Salemi. 2021. "The Role of Nutritional Lifestyle and Physical Activity in Multiple Sclerosis Pathogenesis and Management: A Narrative Review" Nutrients 13, no. 11: 3774. https://doi.org/10.3390/nu13113774
APA StyleFanara, S., Aprile, M., Iacono, S., Schirò, G., Bianchi, A., Brighina, F., Dominguez, L. J., Ragonese, P., & Salemi, G. (2021). The Role of Nutritional Lifestyle and Physical Activity in Multiple Sclerosis Pathogenesis and Management: A Narrative Review. Nutrients, 13(11), 3774. https://doi.org/10.3390/nu13113774