
Insulin-Induced Cardiomyocytes Hypertrophy That Is Prevented by Taurine via β-alanine-Sensitive Na+-Taurine Symporter


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Cells
2.2. Quantitative 3D Confocal Microscopy
2.3. DNA, RNA and Protein Extraction
2.4. Reverse Transcription PCR Analysis
2.5. Western Blot
2.6. Densitometry
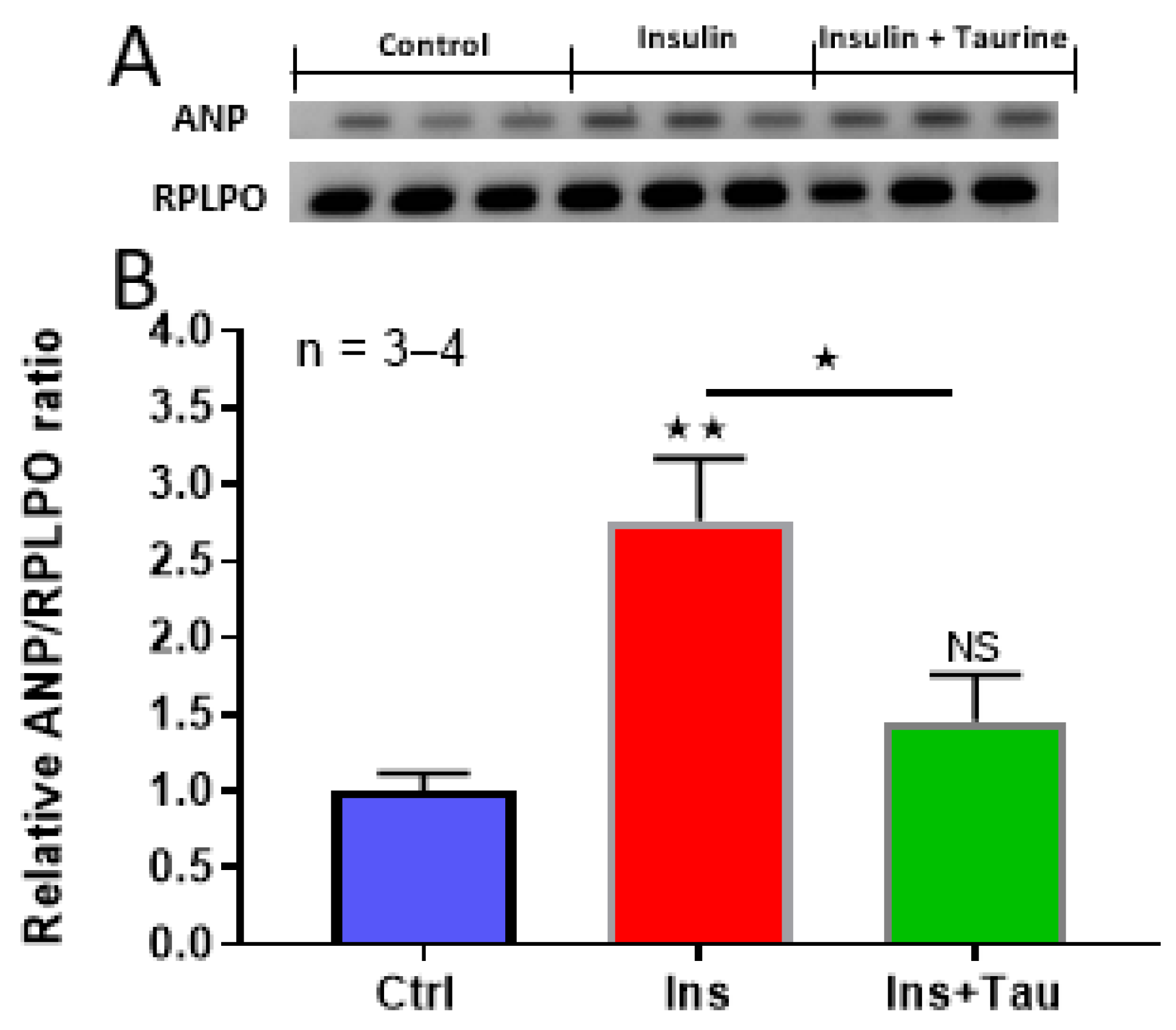
3. Results
3.1. Modulation of the Whole Cell and Nuclear Volumes by a 48-h Treatment with Insulin (80 μU/mL), Taurine (20 mM), and β-alanine (500 μM) of Ventricular Cardiomyocytes
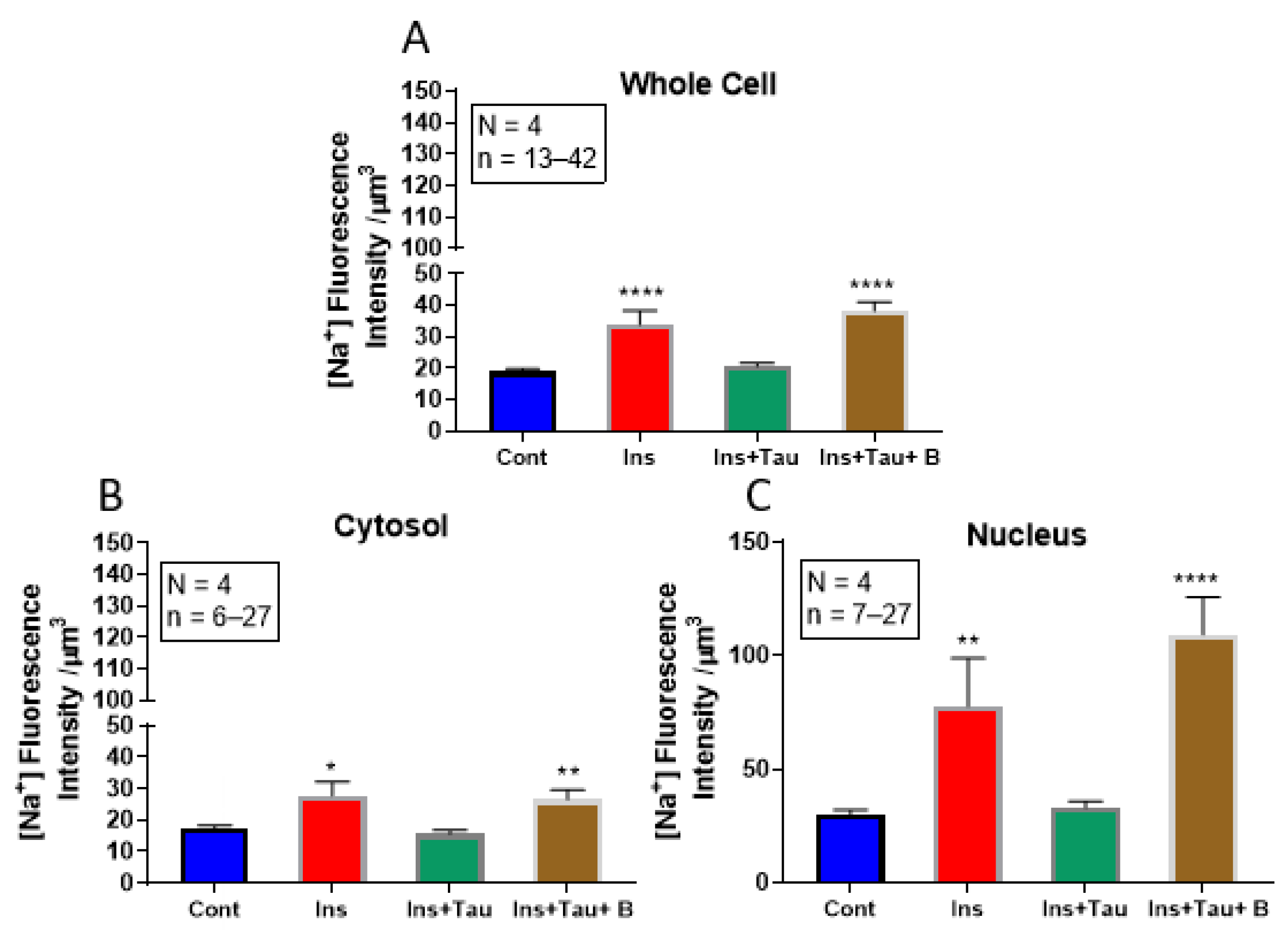
3.2. Modulation of Cytosolic and Nuclear Sodium Levels by Insulin, Insulin + Taurine, and Insulin + Taurine + β-alanine in Ventricular Cardiomyocytes
3.3. Modulation of Cytosolic and Nuclear Calcium Levels by Insulin and Taurine in Adult Rat Ventricular Cardiomyocytes
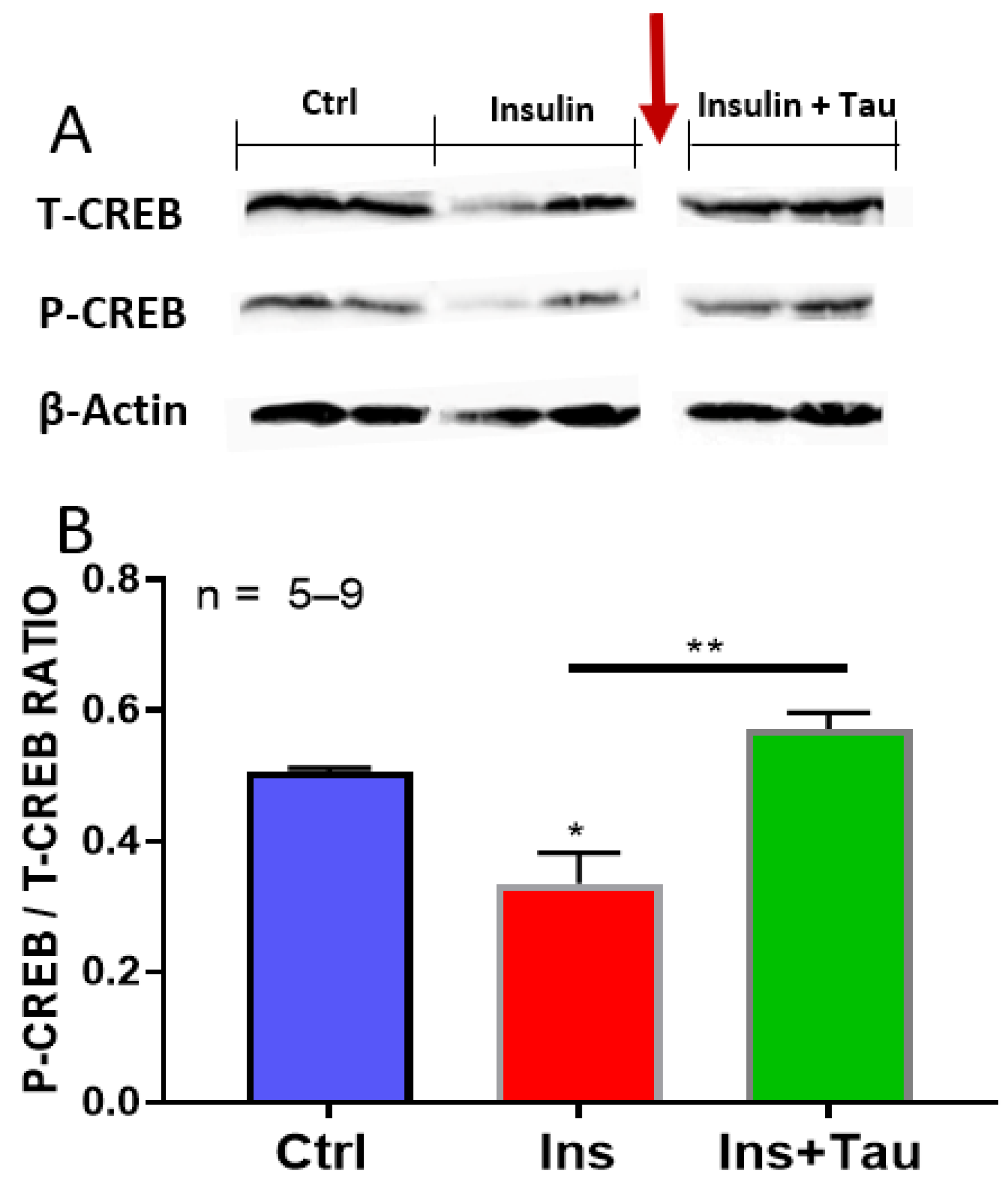
3.4. Effect of Long-Term Treatment with Insulin and Insulin + Taurine on the Ratio of pCREB/tCREB
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hunter, J.J.; Chien, K.R. Signaling Pathways for Cardiac Hypertrophy and Failure. N. Engl. J. Med. 1999, 341, 1276–1283. [Google Scholar] [CrossRef]
- Lemmens, K.; Doggen, K.; De Keulenaer, G.W. Role of Neuregulin-1/ErbB Signaling in Cardiovascular Physiology and Disease: Implications for Therapy of Heart Failure. Circulation 2007, 116, 954–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samak, M.; Fatullayev, J.; Sabashnikov, A.; Zeriouh, M.; Schmack, B.; Farag, M.; Popov, A.-F.; Dohmen, P.M.; Choi, Y.-H.; Wahlers, T.; et al. Cardiac Hypertrophy: An Introduction to Molecular and Cellular Basis. Med. Sci. Monit. Basic Res. 2016, 22, 75–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bkaily, G.; Econornos, D.; Potvin, L.; Ardilouze, J.-L.; Marriott, C.; Corcos, J.; Bonneau, D.; Fong, C.N. Blockade of Insulin Sensitive Steady-State R-Type Ca2+ Channel by PN 200-110 in Heart and Vascular Smooth Muscle. Mol. Cell. Biochem. 1992, 117, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Bkaily, G.; Naik, R.; Jaalouk, D.; Jacques, D.; Economos, D.; D’Orléans-Juste, P.; Pothier, P. Endothelin-1 and Insulin Activate the Steady-State Voltage Dependent R-Type Ca2+ Channel in Aortic Smooth Muscle Cells via a Pertussis Toxin and Cholera Toxin Sensitive G-Protein. Mol. Cell. Biochem. 1998, 183, 39–47. [Google Scholar] [CrossRef]
- Azuma, M.; Takahashi, K.; Fukuda, T.; Ohyabu, Y.; Yamamoto, I.; Kim, S.; Iwao, H.; Schaffer, S.W.; Azuma, J. Taurine Attenuates Hypertrophy Induced by Angiotensin II in Cultured Neonatal Rat Cardiac Myocytes. Eur. J. Pharmacol. 2000, 403, 181–188. [Google Scholar] [CrossRef]
- Singh, P.; Marcu, K.B.; Goldring, M.B.; Otero, M. Phenotypic Instability of Chondrocytes in Osteoarthritis: On a Path to Hypertrophy. Ann. N. Y. Acad. Sci. 2019, 1442, 17–34. [Google Scholar] [CrossRef]
- Bkaily, G.; Haddad, G.; Jaalouk, D.; Gros-Louis, N.; Benchekroun, M.T.; Naik, R.; Pothier, P.; D’Orléans-Juste, P.; Bui, M.; Wang, S.; et al. Modulation of Ca2+ and Na+ Transport by Taurine in Heart and Vascular Smooth Muscle. Adv. Exp. Med. Biol. 1996, 403, 263–273. [Google Scholar] [CrossRef]
- Bkaily, G.; Jaalouk, D.; Haddad, G.; Gros-Louis, N.; Simaan, M.; Naik, R.; Pothier, P. Modulation of Cytosolic and Nuclear Ca2+ and Na+ Transport by Taurine in Heart Cells. Mol. Cell. Biochem. 1997, 170, 1–8. [Google Scholar] [CrossRef]
- Bkaily, G.; Jaalouk, D.; Sader, S.; Shbaklo, H.; Pothier, P.; Jacques, D.; D’Orléans-Juste, P.; Cragoe, E.J.; Bose, R. Taurine Indirectly Increases [Ca]i by Inducing Ca2+ Influx through the Na(+)-Ca2+ Exchanger. Mol. Cell. Biochem. 1998, 188, 187–197. [Google Scholar] [CrossRef]
- Bkaily, G.; Simon, Y.; Jazzar, A.; Najibeddine, H.; Normand, A.; Jacques, D. High Na+ Salt Diet and Remodeling of Vascular Smooth Muscle and Endothelial Cells. Biomedicines 2021, 9, 883. [Google Scholar] [CrossRef]
- Hamali, B.; Pichler, S.; Wischnitzki, E.; Schicker, K.; Burger, M.; Holy, M.; Jaentsch, K.; Molin, M.; Sehr, E.M.; Kudlacek, O.; et al. Identification and Characterization of the Fasciola Hepatica Sodium- and Chloride-Dependent Taurine Transporter. PLoS Negl. Trop. Dis. 2018, 12, e0006428. [Google Scholar] [CrossRef] [Green Version]
- Pasantes-Morales, H.; Quesada, O.; Cárabez, A.; Huxtable, R.J. Effects of the Taurine Transport Antagonist, Guanidinoethane Sulfonate, and β-Alanine on the Morphology of Rat Retina. J. Neurosci. Res. 1983, 9, 135–143. [Google Scholar] [CrossRef]
- Dantzler, W.H.; Silbernagl, S. Renal Tubular Reabsorption of Taurine, Gamma-Aminobutyric Acid (GABA) and Beta-Alanine Studied by Continuous Microperfusion. Pflügers Archiv 1976, 367, 123–128. [Google Scholar] [CrossRef]
- Steven, A.; Friedrich, M.; Jank, P.; Heimer, N.; Budczies, J.; Denkert, C.; Seliger, B. What Turns CREB on? And off? And Why Does It Matter? Cell. Mol. Life Sci. 2020, 77, 4049–4067. [Google Scholar] [CrossRef]
- Truong, V.; Anand-Srivastava, M.B.; Srivastava, A.K. Role of Cyclic AMP Response Element Binding Protein (CREB) in Angiotensin II-Induced Responses in Vascular Smooth Muscle Cells. Can. J. Physiol. Pharmacol. 2021, 99, 30–35. [Google Scholar] [CrossRef]
- Watson, P.A.; Reusch, J.E.B.; McCune, S.A.; Leinwand, L.A.; Luckey, S.W.; Konhilas, J.P.; Brown, D.A.; Chicco, A.J.; Sparagna, G.C.; Long, C.S.; et al. Restoration of CREB Function Is Linked to Completion and Stabilization of Adaptive Cardiac Hypertrophy in Response to Exercise. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H246–H259. [Google Scholar] [CrossRef] [PubMed]
- Kudryavtseva, O.; Aalkjaer, C.; Matchkov, V.V. Vascular Smooth Muscle Cell Phenotype Is Defined by Ca2+-Dependent Transcription Factors. FEBS J. 2013, 280, 5488–5499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schauer, I.E.; Knaub, L.A.; Lloyd, M.; Watson, P.A.; Gliwa, C.; Lewis, K.E.; Chait, A.; Klemm, D.J.; Gunter, J.M.; Bouchard, R.; et al. CREB Downregulation in Vascular Disease: A Common Response to Cardiovascular Risk. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, C.; MacDonald, C.; Willett, B.; Houslay, M.D. Analysis of the Adenylate Cyclase Signalling System, and Alterations Induced by Culture with Insulin, in a Novel SV40-DNA-Immortalized Hepatocyte Cell Line (P9 Cells). Biochem. J. 1994, 300 Pt 3, 835–842. [Google Scholar] [CrossRef] [Green Version]
- Moxham, C.M.; Malbon, C.C. Insulin Action Impaired by Deficiency of the G-Protein Subunit Giα2. Nature 1996, 379, 840–844. [Google Scholar] [CrossRef]
- Bkaily, G.; Sperelakis, N.; Doane, J. A New Method for Preparation of Isolated Single Adult Myocytes. Am. J. Physiol. 1984, 247, H1018–H1026. [Google Scholar] [CrossRef]
- Bkaily, G.; Jazzar, A.; Normand, A.; Simon, Y.; Al-Khoury, J.; Jacques, D. Taurine and Cardiac Disease: State of the Art and Perspectives. Can. J. Physiol. Pharmacol. 2020, 98, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Kahn, S.E. The Relative Contributions of Insulin Resistance and Beta-Cell Dysfunction to the Pathophysiology of Type 2 Diabetes. Diabetologia 2003, 46, 3–19. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.-J.; Arneja, A.S.; Tappia, P.S.; Dhalla, N.S. The Potential Health Benefits of Taurine in Cardiovascular Disease. Exp. Clin. Cardiol. 2008, 13, 57–65. [Google Scholar] [PubMed]
- Bkaily, G.; Pothier, P.; D’Orléans-Juste, P.; Simaan, M.; Jacques, D.; Jaalouk, D.; Belzile, F.; Hassan, G.; Boutin, C.; Haddad, G.; et al. The Use of Confocal Microscopy in the Investigation of Cell Structure and Function in the Heart, Vascular Endothelium and Smooth Muscle Cells. Mol. Cell. Biochem. 1997, 172, 171–194. [Google Scholar] [CrossRef] [PubMed]
- Bkaily, G.; Al-Khoury, J.; Simon, Y.; Jacques, D. Intracellular Free Calcium Measurement Using Confocal Imaging. Methods Mol. Biol. 2017, 1527, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Bkaily, G.; Jacques, D.; Pothier, P. Use of Confocal Microscopy to Investigate Cell Structure and Function. Methods Enzymol. 1999, 307, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Jacques, D.; Sader, S.; Perreault, C.; Fournier, A.; Pelletier, G.; Beck-Sickinger, A.G.; Descorbeth, M. Presence of Neuropeptide Y and the Y1 Receptor in the Plasma Membrane and Nuclear Envelope of Human Endocardial Endothelial Cells: Modulation of Intracellular Calcium. Can. J. Physiol. Pharmacol. 2003, 81, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Bkaily, G.; Sleiman, S.; Stephan, J.; Asselin, C.; Choufani, S.; Kamal, M.; Jacques, D.; Gobeil, F.; D’Orléans-Juste, P. Angiotensin II AT1 Receptor Internalization, Translocation and de Novo Synthesis Modulate Cytosolic and Nuclear Calcium in Human Vascular Smooth Muscle Cells. Can. J. Physiol. Pharmacol. 2003, 81, 274–287. [Google Scholar] [CrossRef]
- Bkaily, G.; Chahine, M.; Al-Khoury, J.; Avedanian, L.; Beier, N.; Scholz, W.; Jacques, D. Na(+)-H(+) Exchanger Inhibitor Prevents Early Death in Hereditary Cardiomyopathy. Can. J. Physiol. Pharmacol. 2015, 93, 923–934. [Google Scholar] [CrossRef] [Green Version]
- Chahine, M.; Bkaily, G.; Nader, M.; Al-Khoury, J.; Jacques, D.; Beier, N.; Scholz, W. NHE-1-Dependent Intracellular Sodium Overload in Hypertrophic Hereditary Cardiomyopathy: Prevention by NHE-1 Inhibitor. J. Mol. Cell. Cardiol. 2005, 38, 571–582. [Google Scholar] [CrossRef]
- Bkaily, G.; Abou Abdallah, N.; Simon, Y.; Jazzar, A.; Jacques, D. Vascular Smooth Muscle Remodeling in Health and Disease. Can. J. Physiol. Pharmacol. 2020, 99, 171–178. [Google Scholar] [CrossRef]
- Gardner, D.G. Natriuretic Peptides: Markers or Modulators of Cardiac Hypertrophy? Trends Endocrinol. Metab. 2003, 14, 411–416. [Google Scholar] [CrossRef]
- Galderisi, M.; Anderson, K.M.; Wilson, P.W.; Levy, D. Echocardiographic Evidence for the Existence of a Distinct Diabetic Cardiomyopathy (the Framingham Heart Study). Am. J. Cardiol. 1991, 68, 85–89. [Google Scholar] [CrossRef]
- Grossman, E.; Shemesh, J.; Shamiss, A.; Thaler, M.; Carroll, J.; Rosenthal, T. Left Ventricular Mass in Diabetes-Hypertension. Arch. Intern. Med. 1992, 152, 1001–1004. [Google Scholar] [CrossRef]
- Lin, X.; Yang, P.; Reece, E.A.; Yang, P. Pregestational Type 2 Diabetes Mellitus Induces Cardiac Hypertrophy in the Murine Embryo through Cardiac Remodeling and Fibrosis. Am. J. Obstet. Gynecol. 2017, 217, 216.e1–216.e13. [Google Scholar] [CrossRef]
- Sadoshima, J.; Izumo, S. The Cellular and Molecular Response of Cardiac Myocytes to Mechanical Stress. Annu. Rev. Physiol. 1997, 59, 551–571. [Google Scholar] [CrossRef]
- Gerdes, A.M.; Liu, Z.; Zimmer, H.G. Changes in Nuclear Size of Cardiac Myocytes during the Development and Progression of Hypertrophy in Rats. Cardioscience 1994, 5, 203–208. [Google Scholar]
- Jacques, D.; Bkaily, G. Endocardial Endothelial Cell Hypertrophy Takes Place during the Development of Hereditary Cardiomyopathy. Mol. Cell. Biochem. 2019, 453, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Komi, P. Strength and Power in Sport: Olympic Encyclopedia of Sports Medicine; John Wiley & Sons, Inc.: Chichester, UK, 2002; ISBN 978-1-4051-4059-1. [Google Scholar]
- Ulrich-Lai, Y.M.; Figueiredo, H.F.; Ostrander, M.M.; Choi, D.C.; Engeland, W.C.; Herman, J.P. Chronic Stress Induces Adrenal Hyperplasia and Hypertrophy in a Subregion-Specific Manner. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E965–E973. [Google Scholar] [CrossRef] [PubMed]
- Watson, P.A.; Nesterova, A.; Burant, C.F.; Klemm, D.J.; Reusch, J.E. Diabetes-Related Changes in CAMP Response Element-Binding Protein Content Enhance Smooth Muscle Cell Proliferation and Migration. J. Biol. Chem. 2001, 276, 46142–46150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eyster, C.A.; Matsuzaki, S.; Newhardt, M.F.; Giorgione, J.R.; Humphries, K.M. Diabetes Induced Decreases in PKA Signaling in Cardiomyocytes: The Role of Insulin. PLoS ONE 2020, 15, e0231806. [Google Scholar] [CrossRef]
- Bkaily, G.; Avedanian, L.; Al-Khoury, J.; Chamoun, M.; Semaan, R.; Jubinville-Leblanc, C.; D’Orléans-Juste, P.; Jacques, D. Nuclear Membrane R-Type Calcium Channels Mediate Cytosolic ET-1-Induced Increase of Nuclear Calcium in Human Vascular Smooth Muscle Cells. Can. J. Physiol. Pharmacol. 2015, 93, 291–297. [Google Scholar] [CrossRef]
- Goguen, J.M.; Halperin, M.L. Can Insulin Administration Cause an Acute Metabolic Acidosis in Vivo? An Experimental Study in Dogs. Diabetologia 1993, 36, 813–816. [Google Scholar] [CrossRef] [Green Version]
- Huxtable, R.J. Physiological Actions of Taurine. Physiol. Rev. 1992, 72, 101–163. [Google Scholar] [CrossRef] [Green Version]
- Khac, L.D.; Naze, S.; Harbon, S. Endothelin Receptor Type A Signals Both the Accumulation of Inositol Phosphates and the Inhibition of Cyclic AMP Generation in Rat Myometrium: Stimulation and Desensitization. Mol. Pharmacol. 1994, 46, 485–494. [Google Scholar] [PubMed]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative Stress and Antioxidant Defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karmazyn, M.; Kilić, A.; Javadov, S. The Role of NHE-1 in Myocardial Hypertrophy and Remodelling. J. Mol. Cell. Cardiol. 2008, 44, 647–653. [Google Scholar] [CrossRef]
- Bkaily, G.; Sperelakis, N. Injection of protein kinase inhibitor into cultured heart cells blocks calcium slow channels. Am. J. Physiol. 1984, 246 Pt 2, H630–H634. [Google Scholar] [CrossRef]
- Smith, R.D.; Goldin, A.L. Protein kinase A phosphorylation enhances sodium channel currents in Xenopus oocytes. Am. J. Physiol. 1992, 263 Pt 1, C660–C666. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.D.; Scheuer, T.; Catterall, W.D. Voltage-dependent potentiation of L-type Ca2+ channels in skeletal muscle cells requires anchored cAMP-dependent protein kinase. Proc. Natl. Acad. Sci. USA 1994, 22, 11492–11496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, R.; Schlichter, L.C. PKA reduces the rat and human KCa3.1 current, CaM binding, and Ca2+ signaling, which requires Ser332/334 in the CaM-binding C terminus. J. Neurosci. 2014, 1, 33–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Chen, J.; Xia, P.; Stratakis, C.A.; Chen, Z. Loss of PKA regulatory subunit 1α aggravates cardiomyocyte necrosis and myocardial ischemia/reperfusion injury. J. Biol. Chem. 2021, 1, 8–10. [Google Scholar] [CrossRef]
- Carriba, P.; Pardo, L.; Parradamas, A.; Lichtenstein, M.P.; Saura, C.A.; Pujol, A.; Masgrau, R.; Galea, E. ATP and noradrenaline activate CREB in astrocytes via noncanonical Ca(2+) and cyclic AMP independent pathways. GLIA 2012, 9, 1330–1344. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jazzar, A.; Jacques, D.; Bkaily, G. Insulin-Induced Cardiomyocytes Hypertrophy That Is Prevented by Taurine via β-alanine-Sensitive Na+-Taurine Symporter. Nutrients 2021, 13, 3686. https://doi.org/10.3390/nu13113686
Jazzar A, Jacques D, Bkaily G. Insulin-Induced Cardiomyocytes Hypertrophy That Is Prevented by Taurine via β-alanine-Sensitive Na+-Taurine Symporter. Nutrients. 2021; 13(11):3686. https://doi.org/10.3390/nu13113686
Chicago/Turabian StyleJazzar, Ashley, Danielle Jacques, and Ghassan Bkaily. 2021. "Insulin-Induced Cardiomyocytes Hypertrophy That Is Prevented by Taurine via β-alanine-Sensitive Na+-Taurine Symporter" Nutrients 13, no. 11: 3686. https://doi.org/10.3390/nu13113686
APA StyleJazzar, A., Jacques, D., & Bkaily, G. (2021). Insulin-Induced Cardiomyocytes Hypertrophy That Is Prevented by Taurine via β-alanine-Sensitive Na+-Taurine Symporter. Nutrients, 13(11), 3686. https://doi.org/10.3390/nu13113686