FTO and PLAG1 Genes Expression and FTO Methylation Predict Changes in Circulating Levels of Adipokines and Gastrointestinal Peptides in Children

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microarray Analysis
2.2. Methylation Analysis
2.2.1. DNA and Selection of Fragments
2.2.2. Restriction Enzyme Digestion
2.2.3. qPCR
2.2.4. Methylation Data Processing and Statistical Analysis
2.3. Study Protocol
2.4. Laboratory Measurements
2.5. Anthropometric Measurements
2.6. Statistical Analysis
3. Results
3.1. Spearman’s Correlation Results of Adipokines with FTO Gene Methylation, Expression and PLAG1 Gene Expression
3.2. Spearman’s Correlation Results of Gastrointestinal Tract Hormones with FTO Gene Methylation, Expression and PLAG1 Gene Expression
3.3. Multiple Linear Regression Model Explaining the Effect of FTO Gene Methylation, FTO Gene Expression and PLAG1 Gene Expression on the Plasma Concentrations of Adipokines and Gastrointestinal Tract Hormones
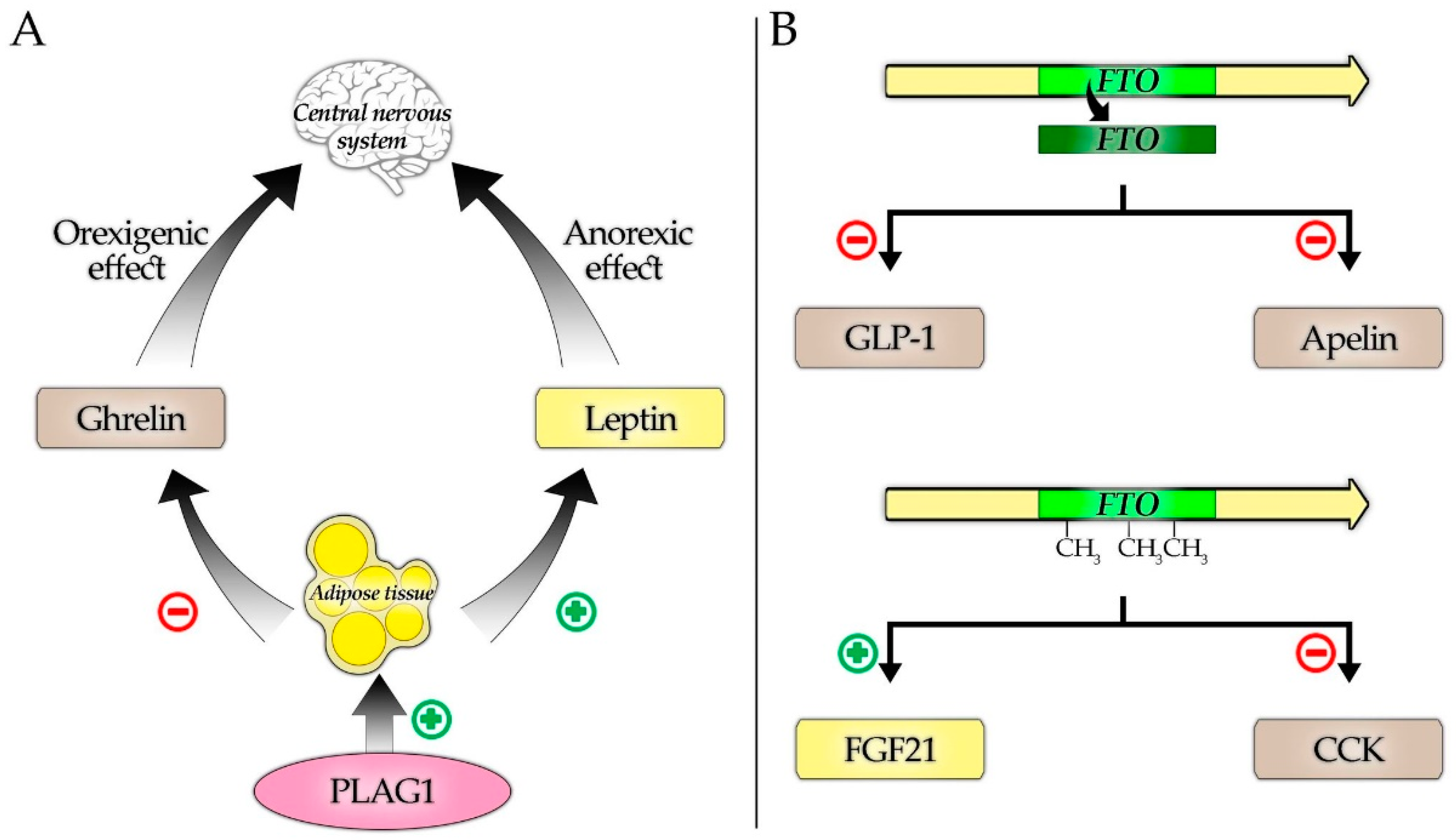
4. Discussion
4.1. FTO Gene
4.1.1. FTO and Adipokines
4.1.2. FTO and Gastrointestinal Tract Hormones
4.2. PLAG1 Gene
4.2.1. PLAG1 and Adipokines
4.2.2. PLAG1 and Gastrointestinal Tract Hormones
4.3. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Overweight/Obesity. Available online: https://www.who.int/data/gho/data/themes/topics/indicator-groups/indicator-group-details/GHO/overweight-obesity (accessed on 9 May 2021).
- Bentham, J.; Di Cesare, M.; Bilano, V.; Bixby, H.; Zhou, B.; Stevens, G.A.; Riley, L.M.; Taddei, C.; Hajifathalian, K.; Lu, Y.; et al. Worldwide Trends in Body-Mass Index, Underweight, Overweight, and Obesity from 1975 to 2016: A Pooled Analysis of 2416 Population-Based Measurement Studies in 128·9 Million Children, Adolescents, and Adults. Lancet 2017, 390, 2627–2642. [Google Scholar] [CrossRef] [Green Version]
- Kadakia, R.; Zheng, Y.; Zhang, Z.; Zhang, W.; Josefson, J.L.; Hou, L. Association of Cord Blood Methylation with Neonatal Leptin: An Epigenome Wide Association Study. PLoS ONE 2019, 14, e0226555. [Google Scholar] [CrossRef] [Green Version]
- Aslibekyan, S.; Do, A.N.; Xu, H.; Li, S.; Irvin, M.R.; Zhi, D.; Tiwari, H.K.; Absher, D.M.; Shuldiner, A.R.; Zhang, T.; et al. CPT1A Methylation Is Associated with Plasma Adiponectin. Nutr. Metab. Cardiovasc. Dis. 2017, 27, 225–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakatochi, M.; Ichihara, S.; Yamamoto, K.; Ohnaka, K.; Kato, Y.; Yokota, S.; Hirashiki, A.; Naruse, K.; Asano, H.; Izawa, H.; et al. Epigenome-Wide Association Study Suggests That SNPs in the Promoter Region of RETN Influence Plasma Resistin Level via Effects on DNA Methylation at Neighbouring Sites. Diabetologia 2015, 58, 2781–2790. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Kohli, S.; Dua, S.; Thinlas, T.; Mohammad, G.; Pasha, M.A.Q. Genetic Differences and Aberrant Methylation in the Apelin System Predict the Risk of High-Altitude Pulmonary Edema. Proc. Natl. Acad. Sci. USA 2015, 112, 6134–6139. [Google Scholar] [CrossRef] [Green Version]
- Wiemerslage, L.; Islam, R.; Van Der Kamp, C.; Cao, H.; Olivo, G.; Ence-Eriksson, F.; Castillo, S.; Larsen, A.L.; Bandstein, M.; Dahlberg, L.S.; et al. A DNA Methylation Site within the KLF13 Gene Is Associated with Orexigenic Processes Based on Neural Responses and Ghrelin Levels. Int. J. Obes. 2017, 41, 990–994. [Google Scholar] [CrossRef]
- Li, P.; Gao, X.; Sun, X.; Li, W.; Yi, B.; Zhu, L. A Novel Epigenetic Mechanism of FXR Inhibiting GLP-1 Secretion via MiR-33 and Its Downstream Targets. Biochem. Biophys. Res. Commun. 2019, 517, 629–635. [Google Scholar] [CrossRef]
- Płatek, T.; Polus, A.; Góralska, J.; Raźny, U.; Dziewońska, A.; Micek, A.; Dembińska-Kieć, A.; Solnica, B.; Malczewska-Malec, M. Epigenetic Regulation of Processes Related to High Level of Fibroblast Growth Factor 21 in Obese Subjects. Genes 2021, 12, 307. [Google Scholar] [CrossRef]
- Chandra, R.; Wang, Y.; Shahid, R.A.; Vigna, S.R.; Freedman, N.J.; Liddle, R.A. Immunoglobulin-like Domain Containing Receptor 1 Mediates Fat-Stimulated Cholecystokinin Secretion. J. Clin. Investig. 2013, 123, 3343–3352. [Google Scholar] [CrossRef] [Green Version]
- Osinski, C.; Le Gléau, L.; Poitou, C.; de Toro-Martin, J.; Genser, L.; Fradet, M.; Soula, H.A.; Leturque, A.; Blugeon, C.; Jourdren, L.; et al. Type 2 Diabetes Is Associated with Impaired Jejunal Enteroendocrine GLP-1 Cell Lineage in Human Obesity. Int. J. Obes. 2021, 45, 170–183. [Google Scholar] [CrossRef]
- Reinehr, T.; Kratzsch, J.; Kiess, W.; Andler, W. Circulating Soluble Leptin Receptor, Leptin, and Insulin Resistance before and after Weight Loss in Obese Children. Int. J. Obes. 2005, 29, 1230–1235. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; López, M.; Rahmouni, K. The Cellular and Molecular Bases of Leptin and Ghrelin Resistance in Obesity. Nat. Rev. Endocrinol. 2017, 13, 338–351. [Google Scholar] [CrossRef]
- Kamińska, A.; Kopczyńska, E.; Bronisz, A.; Żmudzińska, M.; Bieliński, M.; Borkowska, A.; Tyrakowski, T.; Junik, R. An Evaluation of Visfatin Levels in Obese Subjects. Endokrynol. Pol. 2010, 61, 169–173. [Google Scholar]
- Taşkesen, D.; Kirel, B.; Us, T. Serum Visfatin Levels, Adiposity and Glucose Metabolism in Obese Adolescents. J. Clin. Res. Pediatr. Endocrinol. 2012, 4, 76–81. [Google Scholar] [CrossRef]
- Castan-Laurell, I.; Dray, C.; Attané, C.; Duparc, T.; Knauf, C.; Valet, P. Apelin, Diabetes, and Obesity. Endocrine 2011, 40, 1–9. [Google Scholar] [CrossRef]
- Zhang, X.; Yeung, D.C.Y.; Karpisek, M.; Stejskal, D.; Zhou, Z.G.; Liu, F.; Wong, R.L.C.; Chow, W.S.; Tso, A.W.K.; Lam, K.S.L.; et al. Serum FGF21 Levels Are Increased in Obesity and Are Independently Associated with the Metabolic Syndrome in Humans. Diabetes 2008, 57, 1246–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baek, J.; Nam, H.-K.; Rhie, Y.-J.; Lee, K.-H. Serum FGF21 Levels in Obese Korean Children and Adolescents. J. Obes. Metab. Syndr. 2017, 26, 204–209. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Yin, J.; Fu, J.; Li, L.; Grant, S.F.A.; Li, C.; Li, M.; Mi, J.; Li, M.; Gao, S. FGF21 Deficiency Is Associated with Childhood Obesity, Insulin Resistance and Hypoadiponectinaemia: The BCAMS Study. Diabetes Metab. 2017, 43, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Reinehr, T.; Woelfle, J.; Roth, C.L. Lack of Association between Apelin, Insulin Resistance, Cardiovascular Risk Factors, and Obesity in Children: A Longitudinal Analysis. Metabolism 2011, 60, 1349–1354. [Google Scholar] [CrossRef]
- Tapan, S.; Tascilar, E.; Abaci, A.; Sonmez, A.; Kilic, S.; Erbil, M.K.; Ozcan, O. Decreased Plasma Apelin Levels in Pubertal Obese Children. J. Pediatr. Endocrinol. Metab. 2010, 23, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Vehapoglu, A.; Ustabas, F.; Ozgen, T.I.; Terzioglu, S.; Cermik, B.B.; Ozen, O.F. Role of Circulating Adipocytokines Vaspin, Apelin, and Visfatin in the Loss of Appetite in Underweight Children: A Pilot Trial. J. Pediatr. Endocrinol. Metab. 2015, 28, 1065–1071. [Google Scholar] [CrossRef]
- Weiss, R.; Dufour, S.; Groszmann, A.; Petersen, K.; Dziura, J.; Taksali, S.E.; Shulman, G.; Caprio, S. Low Adiponectin Levels in Adolescent Obesity: A Marker of Increased Intramyocellular Lipid Accumulation. J. Clin. Endocrinol. Metab. 2003, 88, 2014–2018. [Google Scholar] [CrossRef] [Green Version]
- Araki, S.; Dobashi, K.; Kubo, K.; Asayama, K.; Shirahata, A. High Molecular Weight, Rather than Total, Adiponectin Levels Better Reflect Metabolic Abnormalities Associated with Childhood Obesity. J. Clin. Endocrinol. Metab. 2006, 91, 5113–5116. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.S.; Lee, W.J.; Funahashi, T.; Tanaka, S.; Matsuzawa, Y.; Chao, C.L.; Chen, C.L.; Tai, T.Y.; Chuang, L.M. Plasma Adiponectin Levels in Overweight and Obese Asians. Obes. Res. 2002, 10, 1104–1110. [Google Scholar] [CrossRef] [PubMed]
- Tschöp, M.; Weyer, C.; Tataranni, P.A.; Devanarayan, V.; Ravussin, E.; Heiman, M.L. Circulating Ghrelin Levels Are Decreased in Human Obesity. Diabetes 2001, 50, 707–709. [Google Scholar] [CrossRef] [Green Version]
- Hira, T.; Pinyo, J.; Hara, H. What Is GLP-1 Really Doing in Obesity? Trends Endocrinol. Metab. 2020, 31, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Gerber, M.; Boettner, A.; Seidel, B.; Lammert, A.; Bär, J.; Schuster, E.; Thiery, J.; Kiess, W.; Kratzsch, J. Serum Resistin Levels of Obese and Lean Children and Adolescents: Biochemical Analysis and Clinical Relevance. J. Clin. Endocrinol. Metab. 2005, 90, 4503–4509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Chan, J.L.; Yiannakouris, N.; Kontogianni, M.; Estrada, E.; Seip, R.; Orlova, C.; Mantzoros, C.S. Circulating Resistin Levels Are Not Associated with Obesity or Insulin Resistance in Humans and Are Not Regulated by Fasting or Leptin Administration: Cross-Sectional and Interventional Studies in Normal, Insulin-Resistant, and Diabetic Subjects. J. Clin. Endocrinol. Metab. 2003, 88, 4848–4856. [Google Scholar] [CrossRef] [Green Version]
- Zou, C.C.; Liang, L.; Hong, F.; Fu, J.F.; Zhao, Z.Y. Serum Adiponectin, Resistin Levels and Non-Alcoholic Fatty Liver Disease in Obese Children. Endocr. J. 2005, 52, 519–524. [Google Scholar] [CrossRef] [Green Version]
- Rehfeld, J.F. Measurement of Cholecystokinin in Plasma with Reference to Nutrition Related Obesity Studies. Nutr. Res. 2020, 76, 1–8. [Google Scholar] [CrossRef]
- FTO Gene Symbol Report | HUGO Gene Nomenclature Committee. Available online: https://www.genenames.org/data/gene-symbol-report/#!/hgnc_id/HGNC:24678 (accessed on 14 May 2021).
- Tweedie, S.; Braschi, B.; Gray, K.; Jones, T.E.M.; Seal, R.L.; Yates, B.; Bruford, E.A. Genenames.Org: The HGNC and VGNC Resources in 2021. Nucleic Acids Res. 2021, 49, D939–D946. [Google Scholar] [CrossRef]
- Frayling, T.M.; Timpson, N.J.; Weedon, M.N.; Zeggini, E.; Freathy, R.M.; Lindgren, C.M.; Perry, J.R.B.; Elliott, K.S.; Lango, H.; Rayner, N.W.; et al. A Common Variant in the FTO Gene Is Associated with Body Mass Index and Predisposes to Childhood and Adult Obesity. Science 2007, 316, 889–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Justice, A.E.; Chittoor, G.; Blanco, E.; Graff, M.; Wang, Y.; Albala, C.; Santos, J.L.; Angel, B.; Lozoff, B.; Voruganti, V.S.; et al. Genetic Determinants of BMI from Early Childhood to Adolescence: The Santiago Longitudinal Study. Pediatr. Obes. 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- da Silva, T.E.R.; Andrade, N.L.; de Oliveira Cunha, D.; Leão-Cordeiro, J.A.B.; Vilanova-Costa, C.A.S.T.; Silva, A.M.T.C. The FTO Rs9939609 Polymorphism and Obesity Risk in Teens: Evidence-Based Meta-Analysis. Obes. Res. Clin. Pract. 2018, 12, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Monteiro, C.; Matos, A.; You, J.; Fraga, A.; Pereira, C.; Catalán, V.; Rodríguez, A.; Gómez-Ambrosi, J.; Frühbeck, G.; et al. Epigenome-Wide DNA Methylation Profiling of Periprostatic Adipose Tissue in Prostate Cancer Patients with Excess Adiposity-a Pilot Study. Clin. Epigenetics 2018, 10, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirchner, H.; Sinha, I.; Gao, H.; Ruby, M.A.; Schönke, M.; Lindvall, J.M.; Barrès, R.; Krook, A.; Näslund, E.; Dahlman-Wright, K.; et al. Altered DNA Methylation of Glycolytic and Lipogenic Genes in Liver from Obese and Type 2 Diabetic Patients. Mol. Metab. 2016, 5, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zou, D.; Li, Z.; Gao, R.; Sang, J.; Zhang, Y.; Li, R.; Xia, L.; Zhang, T.; Niu, G.; et al. EWAS Atlas: A Curated Knowledgebase of Epigenome-Wide Association Studies. Nucleic Acids Res. 2019, 47, D983–D988. [Google Scholar] [CrossRef] [PubMed]
- Mehrdad, M.; Doaei, S.; Gholamalizadeh, M.; Fardaei, M.; Fararouei, M.; Eftekhari, M.H. Association of FTO Rs9939609 Polymorphism with Serum Leptin, Insulin, Adiponectin, and Lipid Profile in Overweight Adults. Adipocyte 2020, 9, 51–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genis-Mendoza, A.D.; Martínez-Magaña, J.J.; Ruiz-Ramos, D.; Gonzalez-Covarrubias, V.; Tovilla-Zarate, C.A.; Narvaez, M.L.L.; Castro, T.B.G.; Juárez-Rojop, I.E.; Nicolini, H. Interaction of FTO Rs9939609 and the Native American-Origin ABCA1 p.Arg230Cys with Circulating Leptin Levels in Mexican Adolescents Diagnosed with Eating Disorders: Preliminary Results. Psychiatry Res. 2020, 291, 113270. [Google Scholar] [CrossRef]
- Duicu, C.; MǍrginean, C.O.; VoidǍzan, S.; Tripon, F.; BǍnescu, C. FTO Rs 9939609 SNP Is Associated with Adiponectin and Leptin Levels and the Risk of Obesity in a Cohort of Romanian Children Population. Medicine (Baltimore) 2016, 95, e3709. [Google Scholar] [CrossRef]
- Ghafarian-Alipour, F.; Ziaee, S.; Ashoori, M.R.; Zakeri, M.S.; Boroumand, M.A.; Aghamohammadzadeh, N.; Abbasi-Majdi, M.; Shool, F.; Asbaghi, N.S.; Mohammadi, A.; et al. Association between FTO Gene Polymorphisms and Type 2 Diabetes Mellitus, Serum Levels of Apelin and Androgen Hormones among Iranian Obese Women. Gene 2018, 641, 361–366. [Google Scholar] [CrossRef]
- Majdi, M.A.; Mohammadzadeh, N.A.; Lotfi, H.; Mahmoudi, R.; Alipour, F.G.; Shool, F.; Moghanloo, M.N.; Porfaraj, S.; Zarghami, N. Correlation of Resistin Serum Level with Fat Mass and Obesity-Associated Gene (FTO) Rs9939609 Polymorphism in Obese Women with Type 2 Diabetes. Diabetes Metab. Syndr. Clin. Res. Rev. 2017, 11, S715–S720. [Google Scholar] [CrossRef]
- López-Bermejo, A.; Petry, C.J.; Díaz, M.; Sebastiani, G.; De Zegher, F.; Dunger, D.B.; Ibáñez, L. The Association between the FTO Gene and Fat Mass in Humans Develops by the Postnatal Age of Two Weeks. J. Clin. Endocrinol. Metab. 2008, 93, 1501–1505. [Google Scholar] [CrossRef] [Green Version]
- Magno, F.C.C.M.; Guaraná, H.C.; Fonseca, A.C.P.; Cabello, G.M.K.; Carneiro, J.R.I.; Pedrosa, A.P.; Ximenes, A.C.; Rosado, E.L. Influence of FTO Rs9939609 Polymorphism on Appetite, Ghrelin, Leptin, IL6, TNFα Levels, and Food Intake of Women with Morbid Obesity. Diabetes, Metab. Syndr. Obes. Targets Ther. 2018, 11, 199–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karra, E.; O’Daly, O.G.; Choudhury, A.I.; Yousseif, A.; Millership, S.; Neary, M.T.; Scott, W.R.; Chandarana, K.; Manning, S.; Hess, M.E.; et al. A Link between FTO, Ghrelin, and Impaired Brain Food-Cue Responsivity. J. Clin. Investig. 2013, 123, 3539–3551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorling, J.L.; Clayton, D.J.; Jones, J.; Carter, W.G.; Thackray, A.E.; King, J.A.; Pucci, A.; Batterham, R.L.; Stensel, D.J. A Randomized Crossover Trial Assessing the Effects of Acute Exercise on Appetite, Circulating Ghrelin Concentrations, and Butyrylcholinesterase Activity in Normal-Weight Males with Variants of the Obesity-Linked FTO Rs9939609 Polymorphism. Am. J. Clin. Nutr. 2019, 110, 1055–1066. [Google Scholar] [CrossRef] [PubMed]
- PLAG1 Gene Symbol Report | HUGO Gene Nomenclature Committee. Available online: https://www.genenames.org/data/gene-symbol-report/#!/hgnc_id/HGNC:9045 (accessed on 14 May 2021).
- PLAG1—Zinc Finger Protein PLAG1—Homo Sapiens (Human)—PLAG1 Gene & Protein. Available online: https://www.uniprot.org/uniprot/Q6DJT9 (accessed on 23 April 2021).
- Bateman, A.; Martin, M.-J.; Orchard, S.; Magrane, M.; Agivetova, R.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; Bursteinas, B.; et al. UniProt: The Universal Protein Knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Warrington, N.M.; Beaumont, R.N.; Horikoshi, M.; Day, F.R.; Helgeland, Ø.; Laurin, C.; Bacelis, J.; Peng, S.; Hao, K.; Feenstra, B.; et al. Maternal and Fetal Genetic Effects on Birth Weight and Their Relevance to Cardio-Metabolic Risk Factors. Nat. Genet. 2019, 51, 804–814. [Google Scholar] [CrossRef]
- Hübel, C.; Gaspar, H.A.; Coleman, J.R.I.; Finucane, H.; Purves, K.L.; Hanscombe, K.B.; Prokopenko, I.; Graff, M.; Ngwa, J.S.; Workalemahu, T.; et al. Genomics of Body Fat Percentage May Contribute to Sex Bias in Anorexia Nervosa. Am. J. Med. Genet. Part. B Neuropsychiatr. Genet. 2019, 180, 428–438. [Google Scholar] [CrossRef] [Green Version]
- Shungin, D.; Winkler, T.; Croteau-Chonka, D.C.; Ferreira, T.; Locke, A.E.; Mägi, R.; Strawbridge, R.J.; Pers, T.H.; Fischer, K.; Justice, A.E.; et al. New Genetic Loci Link Adipose and Insulin Biology to Body Fat Distribution. Nature 2015, 518, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Czogała, W.; Czogała, M.; Strojny, W.; Wątor, G.; Wołkow, P.; Wójcik, M.; Multanowski, M.B.; Tomasik, P.; Wędrychowicz, A.; Kowalczyk, W.; et al. Methylation and Expression of FTO and PLAG1 Genes in Childhood Obesity: Insight into Anthropometric Parameters and Glucose-Lipid Metabolism. Nutrients 2021, 13, 1683. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Kokubun, S.; Itoi, E.; Roach, H.I. Improved Quantification of DNA Methylation Using Methylation-Sensitive Restriction Enzymes and Real-Time PCR. Epigenetics 2007, 2, 86–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redshaw, N.; Huggett, J.F.; Taylor, M.S.; Foy, C.A.; Devonshire, A.S. Quantification of Epigenetic Biomarkers: An Evaluation of Established and Emerging Methods for DNA Methylation Analysis. BMC Genom. 2014, 15, 1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howe, K.L.; Achuthan, P.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Ridwan Amode, M.; Armean, I.M.; Azov, A.G.; Bennett, R.; Bhai, J.; et al. Ensembl 2021. Nucleic Acids Res. 2021, 49, D884–D891. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New Capabilities and Interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- R: The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 13 May 2021).
- Smyth, G.K. Linear Models and Empirical Bayes Methods for Assessing Differential Expression in Microarray Experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3, 3. [Google Scholar] [CrossRef]
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: Open Software Development for Computational Biology and Bioinformatics. Genome Biol. 2004, 5, R80. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- WHO Anthro Survey Analyser and Other Tools. Available online: https://www.who.int/toolkits/child-growth-standards/software (accessed on 4 March 2021).
- FTO Gene—GeneCards | FTO Protein | FTO. Antibody. Available online: https://www.genecards.org/cgi-bin/carddisp.pl?gene=FTO&keywords=FTO (accessed on 16 April 2021).
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinform. 2016, 54, 1.30.1–1.30.33. [Google Scholar] [CrossRef]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference Sequence (RefSeq) Database at NCBI: Current Status, Taxonomic Expansion, and Functional Annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef] [Green Version]
- Jia, G.; Yang, C.G.; Yang, S.; Jian, X.; Yi, C.; Zhou, Z.; He, C. Oxidative Demethylation of 3-Methylthymine and 3-Methyluracil in Single-Stranded DNA and RNA by Mouse and Human FTO. FEBS Lett. 2008, 582, 3313–3319. [Google Scholar] [CrossRef] [Green Version]
- Han, Z.; Niu, T.; Chang, J.; Lei, X.; Zhao, M.; Wang, Q.; Cheng, W.; Wang, J.; Feng, Y.; Chai, J. Crystal Structure of the FTO Protein Reveals Basis for Its Substrate Specificity. Nature 2010, 464, 1205–1209. [Google Scholar] [CrossRef]
- Speakman, J.R. The ‘Fat Mass and Obesity Related’ (FTO) Gene: Mechanisms of Impact on Obesity and Energy Balance. Curr. Obes. Rep. 2015, 4, 73–91. [Google Scholar] [CrossRef]
- Di Renzo, L.; Cioccoloni, G.; Falco, S.; Abenavoli, L.; Moia, A.; Sinibaldi Salimei, P.; De Lorenzo, A. Influence of FTO Rs9939609 and Mediterranean Diet on Body Composition and Weight Loss: A Randomized Clinical Trial NCT01890070 NCT. J. Transl. Med. 2018, 16, 308. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.Y.; Park, J.H.; Park, S.E.; Shon, J.; Park, Y.J. The Fat Mass- and Obesity-Associated (FTO) Gene to Obesity: Lessons from Mouse Models. Obesity 2018, 26, 1674–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labayen, I.; Ruiz, J.R.; Ortega, F.B.; Dalongeville, J.; Jiménez-Pavón, D.; Castillo, M.J.; De Henauw, S.; González-Gross, M.; Bueno, G.; Molnar, D.; et al. Association between the FTO Rs9939609 Polymorphism and Leptin in European Adolescents: A Possible Link with Energy Balance Control. The HELENA Study. Int. J. Obes. 2011, 35, 66–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Luo, X.; Liu, M.; Chen, L. Function and Regulation of Apelin/APJ System in Digestive Physiology and Pathology. J. Cell Physiol. 2019, 234, 7796–7810. [Google Scholar] [CrossRef] [PubMed]
- Kadoglou, N.P.E.; Vrabas, I.S.; Kapelouzou, A.; Lampropoulos, S.; Sailer, N.; Kostakis, A.; Liapis, C.D.; Angelopoulou, N. The Impact of Aerobic Exercise Training on Novel Adipokines, Apelin and Ghrelin, in Patients with Type 2 Diabetes. Med. Sci. Monit. 2012, 18, CR290-5. [Google Scholar] [CrossRef] [Green Version]
- Son, J.S.; Chae, S.A.; Park, B.I.; Du, M.; Song, W. Plasma Apelin Levels in Overweight/Obese Adults Following a Single Bout of Exhaustive Exercise: A Preliminary Cross-Sectional Study. Endocrinol. Diabetes Nutr. 2019, 66, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Kadoglou, N.P.E.; Vrabas, I.S.; Kapelouzou, A.; Angelopoulou, N. The Association of Physical Activity with Novel Adipokines in Patients with Type 2 Diabetes. Eur. J. Intern. Med. 2012, 23, 137–142. [Google Scholar] [CrossRef]
- Chandra, R.; Liddle, R.A. Cholecystokinin. Curr. Opin. Endocrinol. Diabetes Obes. 2007, 14, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Crovesy, L.; Rosado, E.L. Interaction between Genes Involved in Energy Intake Regulation and Diet in Obesity. Nutrition 2019, 67–68, 110547. [Google Scholar] [CrossRef] [PubMed]
- Holst, J.J. The Physiology of Glucagon-like Peptide 1. Physiol. Rev. 2007, 87, 1409–1439. [Google Scholar] [CrossRef]
- Danaher, J.; Stathis, C.G.; Cooke, M.B. Similarities in Metabolic Flexibility and Hunger Hormone Ghrelin Exist between FTO Gene Variants in Response to an Acute Dietary Challenge. Nutrients 2019, 11, 2518. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Ngwa, J.S.; Van Rooij, F.J.A.; Zillikens, M.C.; Wojczynski, M.K.; Frazier-Wood, A.C.; Houston, D.K.; Kanoni, S.; Lemaitre, R.N.; Luan, J.N.A.; et al. Genome-Wide Meta-Analysis of Observational Studies Shows Common Genetic Variants Associated with Macronutrient Intake. Am. J. Clin. Nutr. 2013, 97, 1395–1402. [Google Scholar] [CrossRef] [Green Version]
- PLAG1 Gene—GeneCards | PLAG1 Protein | PLAG1 Antibody. Available online: https://www.genecards.org/cgi-bin/carddisp.pl?gene=PLAG1&keywords=PLAG1 (accessed on 16 April 2021).
- Van Dyck, F.; Declercq, J.; Braem, C.; Van de Ven, W. PLAG1, the Prototype of the PLAG Gene Family: Versatility in Tumour Development (Review). Int. J. Oncol. 2007, 30, 765–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zatkova, A.; Rouillard, J.M.; Hartmann, W.; Lamb, B.J.; Kuick, R.; Eckart, M.; Von Schweinitz, D.; Koch, A.; Fonatsch, C.; Pietsch, T.; et al. Amplification and Overexpression of the IGF2 Regulator PLAG1 in Hepatoblastoma. Genes Chromosom. Cancer 2004, 39, 126–137. [Google Scholar] [CrossRef]
- GWAS Catalog. Available online: https://www.ebi.ac.uk/gwas/home (accessed on 6 March 2021).
- Buniello, A.; Macarthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E.; et al. The NHGRI-EBI GWAS Catalog of Published Genome-Wide Association Studies, Targeted Arrays and Summary Statistics 2019. Nucleic Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Street, M.E.; Smerieri, A.; Montanini, L.; Predieri, B.; Iughetti, L.; Valenzise, M.; De Luca, F.; Vigone, M.; Weber, G.; Maghnie, M.; et al. Interactions among Pro-Inflammatory Cytokines, IGF System and Thyroid Function in Pre-Pubertal Obese Subjects. J. Biol. Regul. Homeost. Agents 2013, 27, 259–266. [Google Scholar]
- Seck, T.; Englaro, P.; Blum, W.F.; Scheidt-Nave, C.; Rascher, W.; Ziegler, R.; Pfeilschifter, J. Leptin Concentrations in Serum from a Randomly Recruited Sample of 50- to 80-Year-Old Men and Women: Positive Association with Plasma Insulin-like Growth Factors (IGFs) and IGF-Binding Protein-3 in Lean, but Not in Obese, Individuals. Eur. J. Endocrinol. 1998, 138, 70–75. [Google Scholar] [CrossRef] [Green Version]
- Kleiman, A.; Keats, E.C.; Chan, N.G.; Khan, Z.A. Elevated IGF2 Prevents Leptin Induction and Terminal Adipocyte Differentiation in Hemangioma Stem Cells. Exp. Mol. Pathol. 2013, 94, 126–136. [Google Scholar] [CrossRef]
- Uchida, A.; Zechner, J.F.; Mani, B.K.; Park, W.M.; Aguirre, V.; Zigman, J.M. Altered Ghrelin Secretion in Mice in Response to Diet-Induced Obesity and Roux-En-Y Gastric Bypass. Mol. Metab. 2014, 3, 717–730. [Google Scholar] [CrossRef]
- de Mello, V.D.F.; Kolehmanien, M.; Schwab, U.; Pulkkinen, L.; Uusitupa, M. Gene Expression of Peripheral Blood Mononuclear Cells as a Tool in Dietary Intervention Studies: What Do We Know so Far? Mol. Nutr. Food Res. 2012, 56, 1160–1172. [Google Scholar] [CrossRef] [PubMed]
- Reynés, B.; Priego, T.; Cifre, M.; Oliver, P.; Palou, A. Peripheral Blood Cells, a Transcriptomic Tool in Nutrigenomic and Obesity Studies: Current State of the Art. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1006–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohane, I.S.; Valtchinov, V.I. Quantifying the white blood cell transcriptome as an accessible window to the multiorgan transcriptome. Bioinformatics 2012, 28, 538–545. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Gene | Localization | Fragment Size | No. of CCGG Sites | Forward | Reverse |
|---|---|---|---|---|---|
| FTO | upstream | 216bp | 2 | CAACTCCAGGGCCTTCTC | GGAGCCTGCCATGTTTCT |
| PLAG1 | exon1 | 202bp | 2 | ACAATGGCTGCTGGAAAGA | CCCTGATATTTCTCCCGCTAAA |
| Baseline Characteristics | Study Group N = 26 |
|---|---|
| boys/girls n (%) | 16/10 (61.54%/38.46%) |
| Age (years) | 13.98 ± 2.6 |
| Height (cm) | 164.85 ± 13.9 |
| Weight (kg) | 76.62 ± 24.98 |
| BMI (kg/m2) | 27.87 ± 7.48 |
| BMI (percentile) | 82.47 ± 30.02 |
| BMI SD | 2.05 ± 1.64 |
| Baseline Characteristics | Study Group N = 26 |
|---|---|
| FTO gene methylation (%) | 4.35 ± 3.9 |
| FTO expression | 266.72 ± 87.46 |
| PLAG1 gene methylation (%) | 0.08 ± 0.08 |
| PLAG1 expression | 82 ± 21.73 |
| AUC Adiponectin (µg/mL/h) | 7.01 ± 4.48 |
| AUC apelin (ng/mL/h) | 4.96 ± 3.47 |
| AUC leptin (ng/mL/h) | 55.08 ± 49.81 |
| AUC leptin receptor (ng/mL/h) | 21.44 ± 13.7 |
| AUC resistin (ng/mL/h) | 7.47 ± 1.94 |
| AUC visfatin | 21.86 ± 10.83 |
| AUC FGF21 (ng/mL/h) | 264.75 ± 157.18 |
| AUC Cholecystokinin (ng/mL/h) | 4.61 ± 2.72 |
| AUC ghrelin (ng/mL/h) | 955.42 ± 302.77 |
| AUC GLP-1 (ng/mL/h) | 1.89 ± 0.82 |
| Adipokines | FTO Gene Methylation | FTO Expression | PLAG1 Expression | |||
|---|---|---|---|---|---|---|
| Spearman’s Correlation Coefficient r | p/pBH Value | Spearman’s Correlation Coefficient r | p/pBH Value | Spearman’s Correlation Coefficient r | p/pBH Value | |
| AUC adiponectin (µg/mL/h) | −0.29 | 0.203/0.271 | −0.221 | 0.336/0.384 | 0.007 | 0.975/0.975 |
| AUC apelin (ng/mL/h) | −0.388 | 0.067/0.322 | −0.758 | <0.001/<0.001 | −0.579 | 0.004/0.014 |
| AUC leptin (ng/mL/h) | 0.294 | 0.164/0.262 | 0.713 | <0.001/<0.001 | 0.676 | <0.001/0.002 |
| AUC leptin receptor (ng/mL/h) | −0.176 | 0.422/0.440 | −0.622 | 0.002/0.005 | −0.493 | 0.017/0.045 |
| AUC resistin (ng/mL/h) | −0.356 | 0.088/0.302 | −0.434 | 0.034/0.054 | −0.19 | 0.373/0.471 |
| AUC visfatin (ng/mL/h) | −0.364 | 0.105/0.252 | −0.442 | 0.045/0.064 | −0.263 | 0.249/0.374 |
| Gastrointestinal Tract Hormones | FTO Gene Methylation | FTO Expression | PLAG1 Expression | |||
|---|---|---|---|---|---|---|
| Spearman’s Correlation Coefficient r | p/pBH Value | Spearman’s Correlation Coefficient r | p/pBH Value | Spearman’s Correlation Coefficient r | p/pBH Value | |
| AUC CCK (ng/mL/h) | −0.508 | 0.011/0.044 | −0.613 | 0.001/0.008 | −0.489 | 0.015/0.12 |
| AUC FGF21 (ng/mL/h) | 0.556 | 0.005/0.04 | 0.525 | 0.008/0.016 | 0.236 | 0.266/0.304 |
| AUC Ghrelin (ng/mL/h) | −0.377 | 0.076/0.122 | −0.265 | 0.222/0.254 | −0.446 | 0.033/0.106 |
| AUC GLP-1 (ng/mL/h) | −0.371 | 0.081/0.118 | −0.638 | 0.001/0.003 | −0.457 | 0.029/0.155 |
| Hormone | Gene (Non-Standardized Regression Coefficient (B) ± SEM, p-Value) | R2adj | p/pBH-Value for R2 |
|---|---|---|---|
| AUC apelin (ng/mL/h) | FTO expression (−1.09 ± 0.51, 0.045) | 0.402 | 0.009/0.018 |
| AUC CCK (ng/mL/h) | FTO methylation (−0.25 ± 0.12, 0.05) | 0.435 | 0.002/0.008 |
| AUC FGF21 (ng/mL/h) | FTO methylation (20.51 ± 8.31, 0.023) | 0.297 | 0.029/0.046 |
| AUC ghrelin (ng/mL/h) | FTO methylation (−39.47 ± 16.94, 0.032) PLAG1 expression (−99.6 ± 33.3, 0.008) | 0.260 | 0.049/0.056 |
| AUC GLP-1 (ng/mL/h) | FTO expression (−0.25 ± 0.1, 0.020) | 0.364 | 0.004/0.011 |
| AUC leptin (ng/mL/h) | PLAG1 expression (10.44 ± 4.12, 0.020) | 0.537 | <0.001/0.006 |
| AUC leptin receptor (ng/mL/h) | FTO expression (−4.00 ± 1.83, 0.041) | 0.239 | 0.042/0.056 |
| AUC adiponectin (µg/mL/h) | FTO expression (−1.94 ± 0.80, 0.028) | 0.142 | 0.098/0.098 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Czogała, W.; Strojny, W.; Schab, M.; Grabowska, A.; Miklusiak, K.; Kowalczyk, W.; Łazarczyk, A.; Tomasik, P.; Skoczeń, S. FTO and PLAG1 Genes Expression and FTO Methylation Predict Changes in Circulating Levels of Adipokines and Gastrointestinal Peptides in Children. Nutrients 2021, 13, 3585. https://doi.org/10.3390/nu13103585
Czogała W, Strojny W, Schab M, Grabowska A, Miklusiak K, Kowalczyk W, Łazarczyk A, Tomasik P, Skoczeń S. FTO and PLAG1 Genes Expression and FTO Methylation Predict Changes in Circulating Levels of Adipokines and Gastrointestinal Peptides in Children. Nutrients. 2021; 13(10):3585. https://doi.org/10.3390/nu13103585
Chicago/Turabian StyleCzogała, Wojciech, Wojciech Strojny, Magdalena Schab, Agnieszka Grabowska, Karol Miklusiak, Wojciech Kowalczyk, Agnieszka Łazarczyk, Przemysław Tomasik, and Szymon Skoczeń. 2021. "FTO and PLAG1 Genes Expression and FTO Methylation Predict Changes in Circulating Levels of Adipokines and Gastrointestinal Peptides in Children" Nutrients 13, no. 10: 3585. https://doi.org/10.3390/nu13103585
APA StyleCzogała, W., Strojny, W., Schab, M., Grabowska, A., Miklusiak, K., Kowalczyk, W., Łazarczyk, A., Tomasik, P., & Skoczeń, S. (2021). FTO and PLAG1 Genes Expression and FTO Methylation Predict Changes in Circulating Levels of Adipokines and Gastrointestinal Peptides in Children. Nutrients, 13(10), 3585. https://doi.org/10.3390/nu13103585