Attenuating Effect of Peruvian Cocoa Populations on the Acute Asthmatic Response in Brown Norway Rats


 , , ,
, , ,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cocoa Population Characterization
2.1.1. Phenolic Compounds and Antioxidant Activity
2.1.2. Methylxanthine Quantification
2.2. Animals
2.3. In vitro Study
2.3.1. Peritoneal Macrophages and Lymphocytes Culture
2.3.2. Radical Oxygen Species (ROS) Production
2.3.3. M1 and M2 Characterization
2.3.4. IgG and Tumor Necrosis Factor (TNF)-α Quantification by ELISA
2.4. In vivo Study
2.4.1. Diets and Animal Groups
2.4.2. Sensitization and Induction of an Anaphylactic Response
2.4.3. Body Temperature Monitoring
2.4.4. Motor Activity Assessment
2.4.5. Sample Collection
2.4.6. Quantification of Plasma Rat Mast Cell Protease II
2.4.7. Antibody Quantification
2.4.8. Quantification of Cysteinyl Leukotriene (CysLT)
2.4.9. Urine Polyphenols
2.5. Statistical Analysis
3. Results
3.1. Cocoa Peruvian Populations Characterization
3.2. In Vitro Effects of Cocoa Peruvian Populations
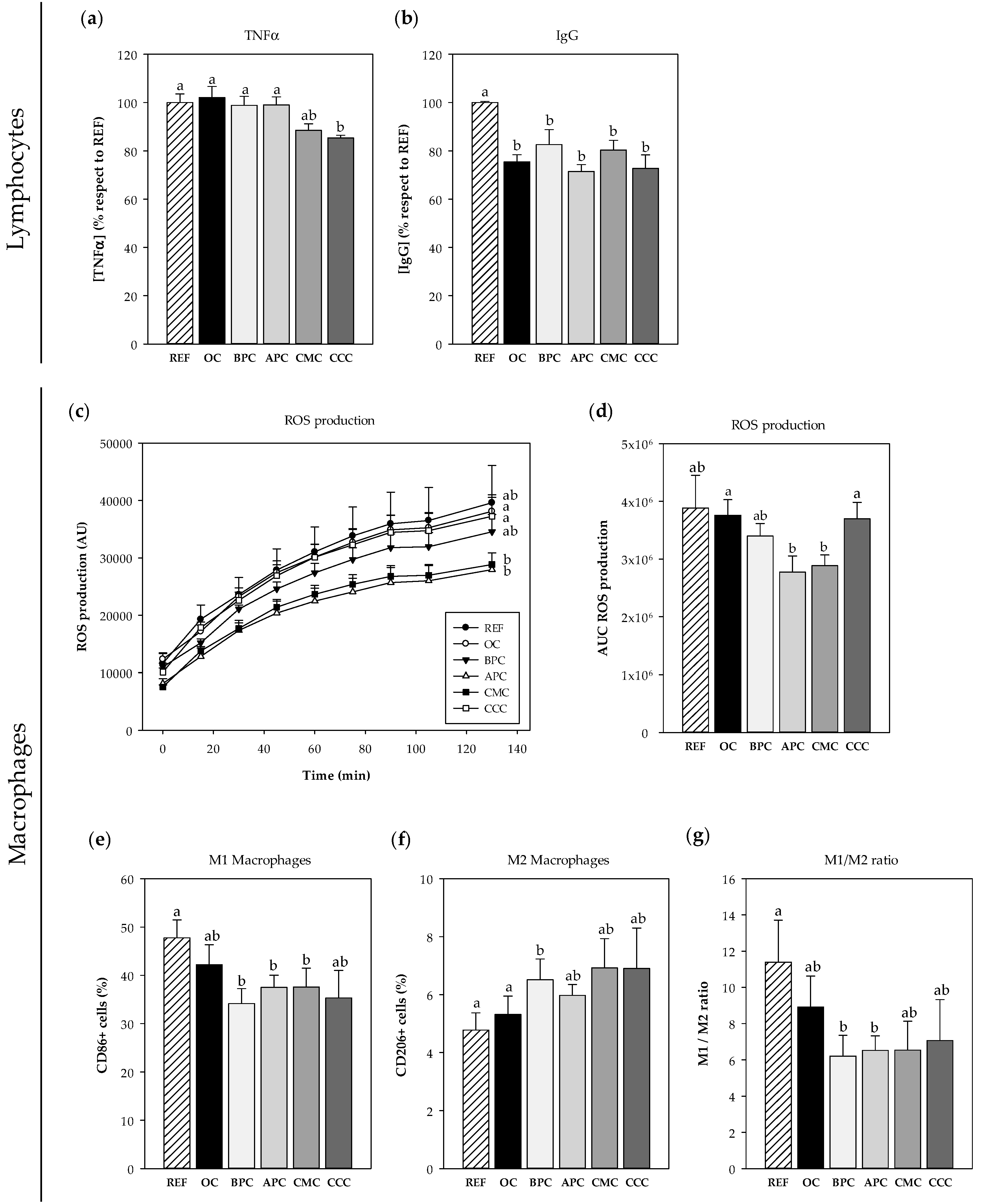
3.2.1. Effects on Spleen Cells
3.2.2. Effects on Macrophages
3.3. In Vivo Effects of Cocoa Peruvian Populations
3.3.1. Body Weight and Food and Water Intake
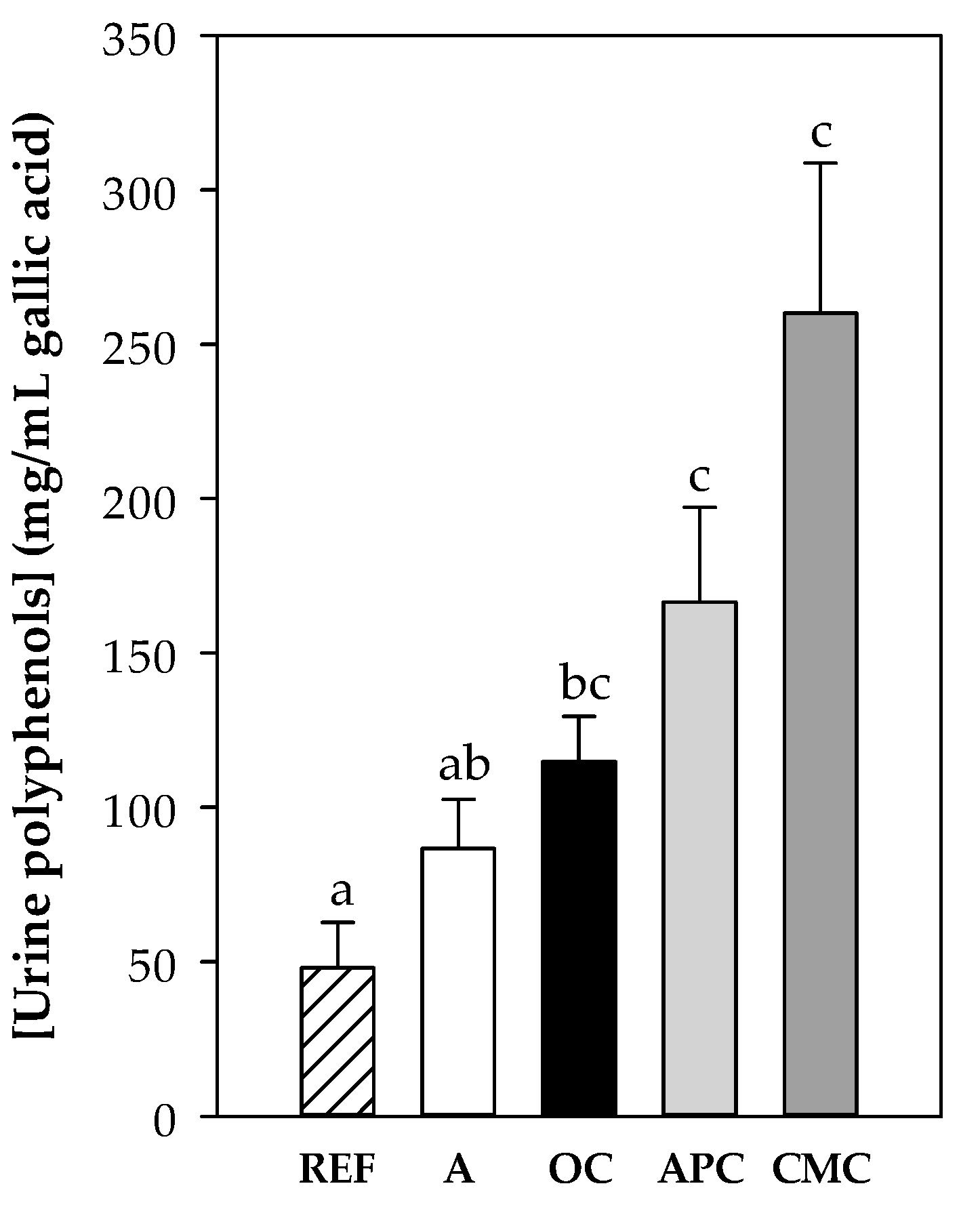
3.3.2. Content of Polyphenols in Urine
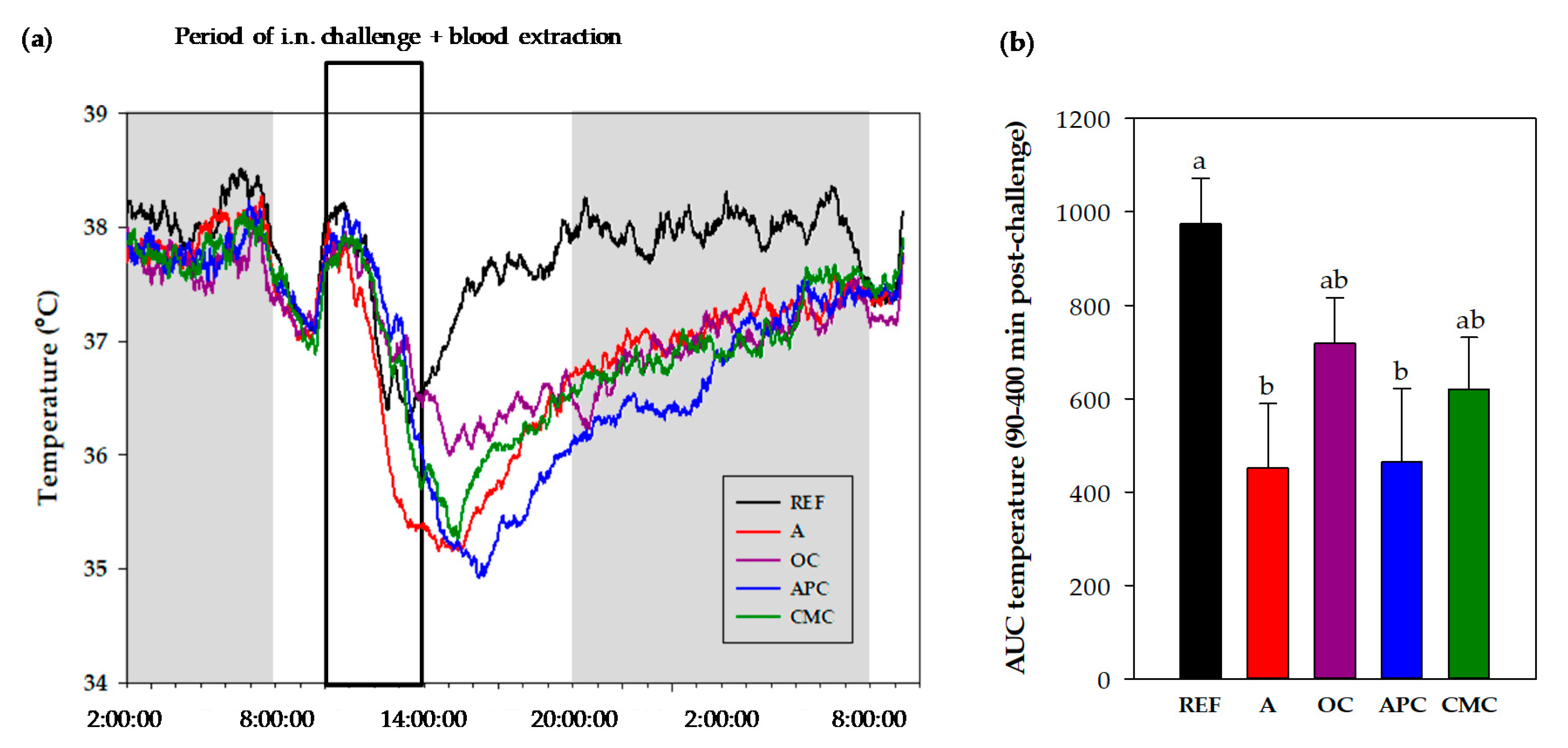
3.3.3. Changes in Body Temperature After i.n. Challenge
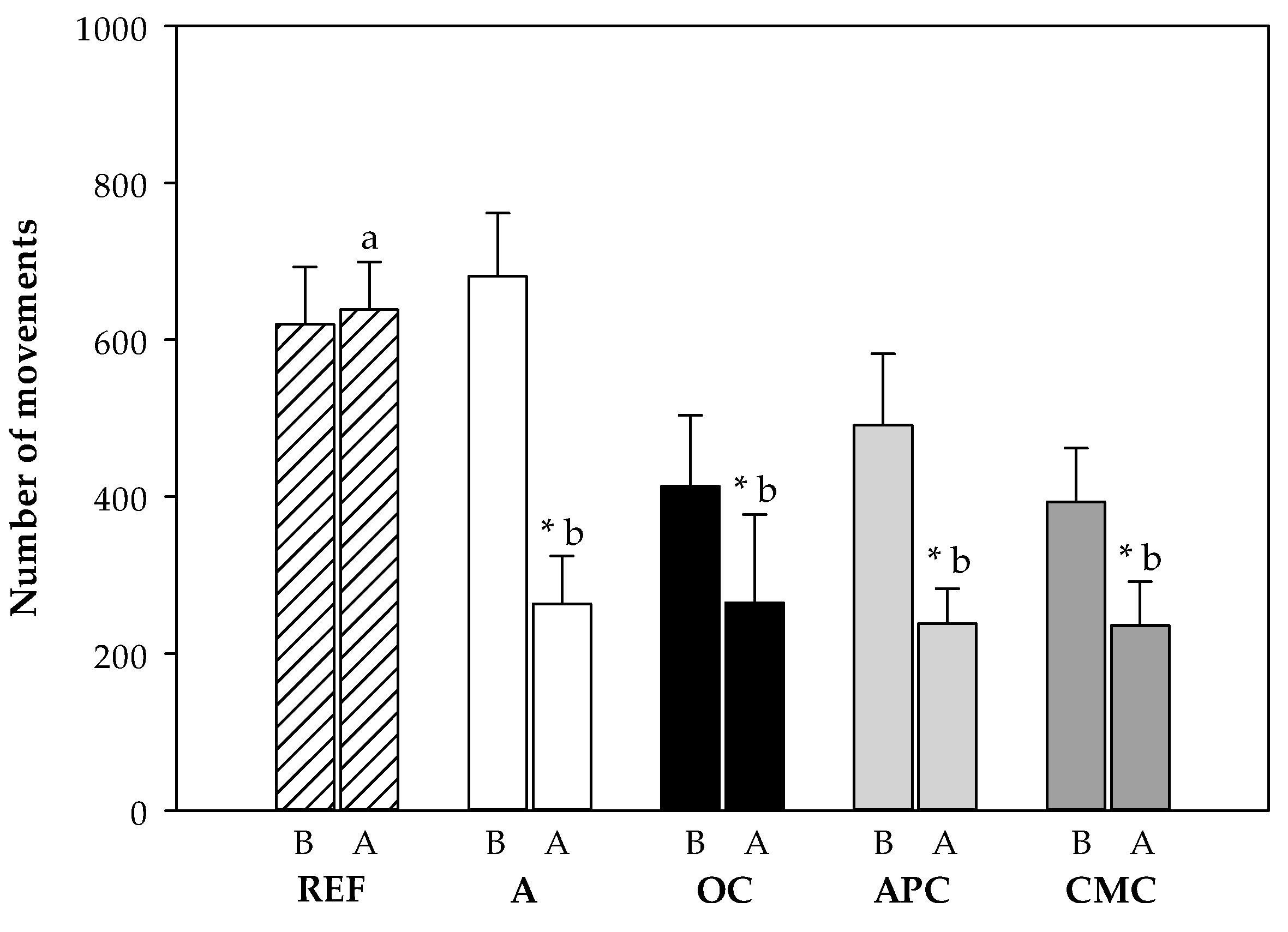
3.3.4. Changes in Motor Activity After i.n. Challenge
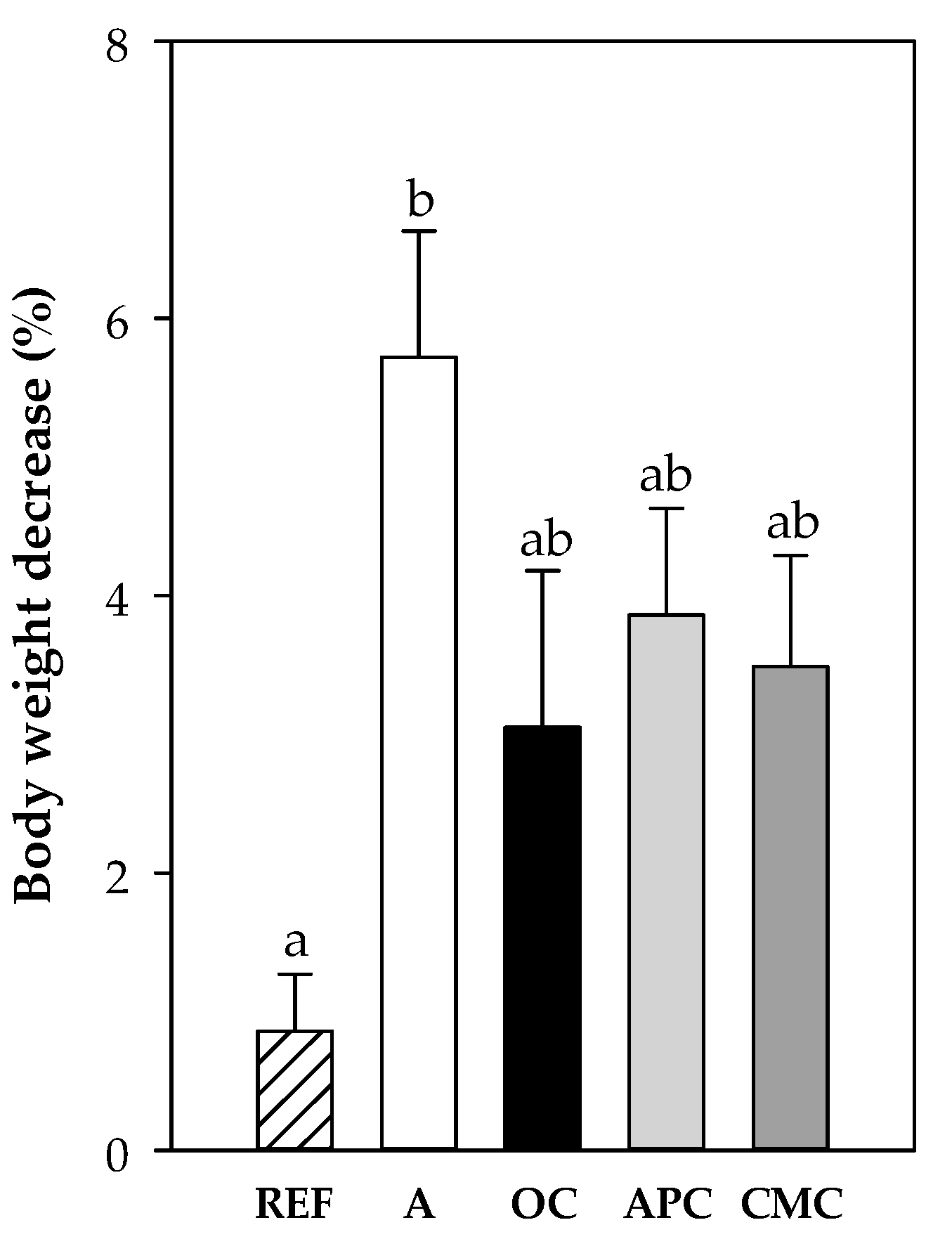
3.3.5. Changes in Body Weight After i.n. Challenge
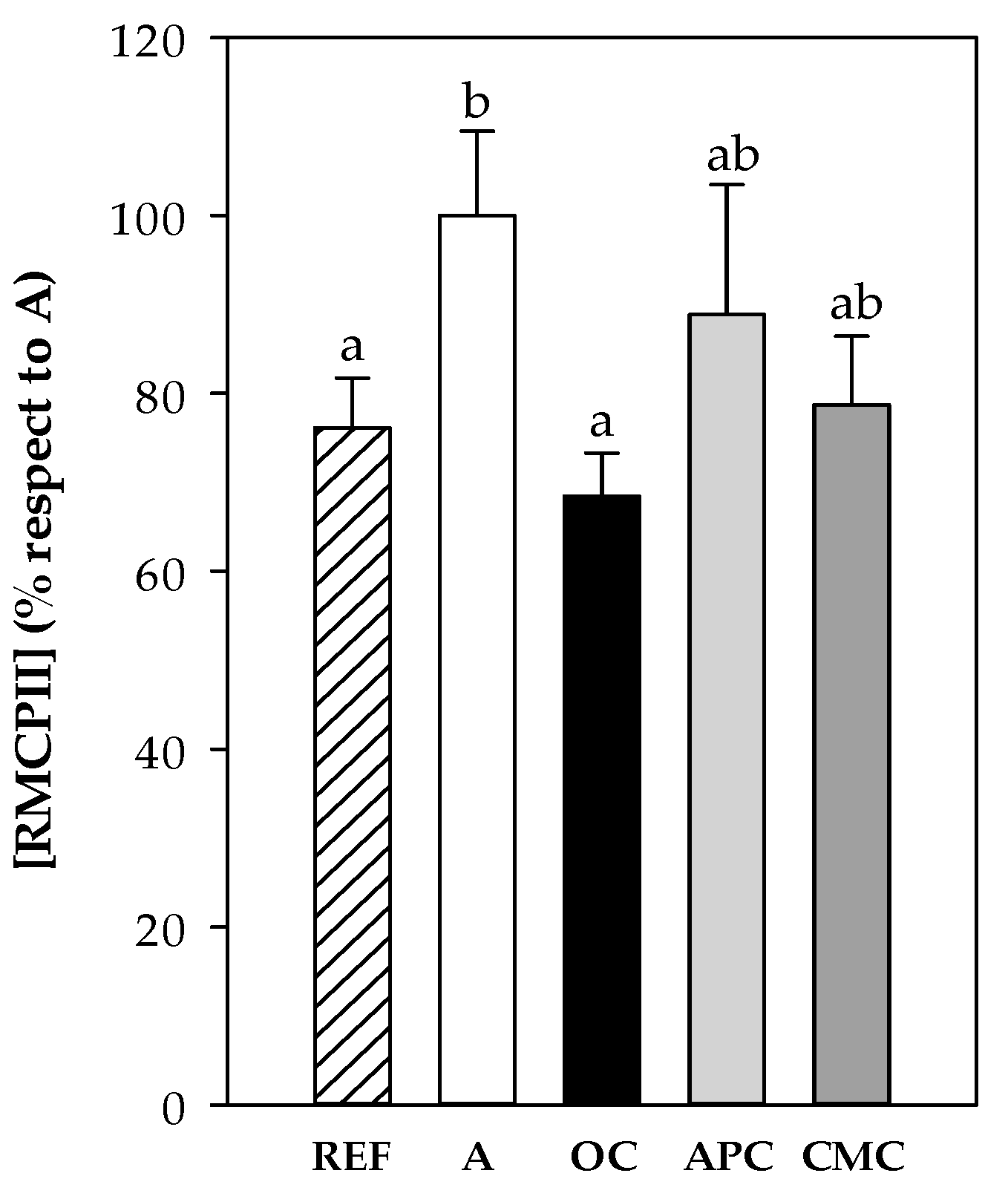
3.3.6. Rat Mast Cell Protease II (RMCPII)
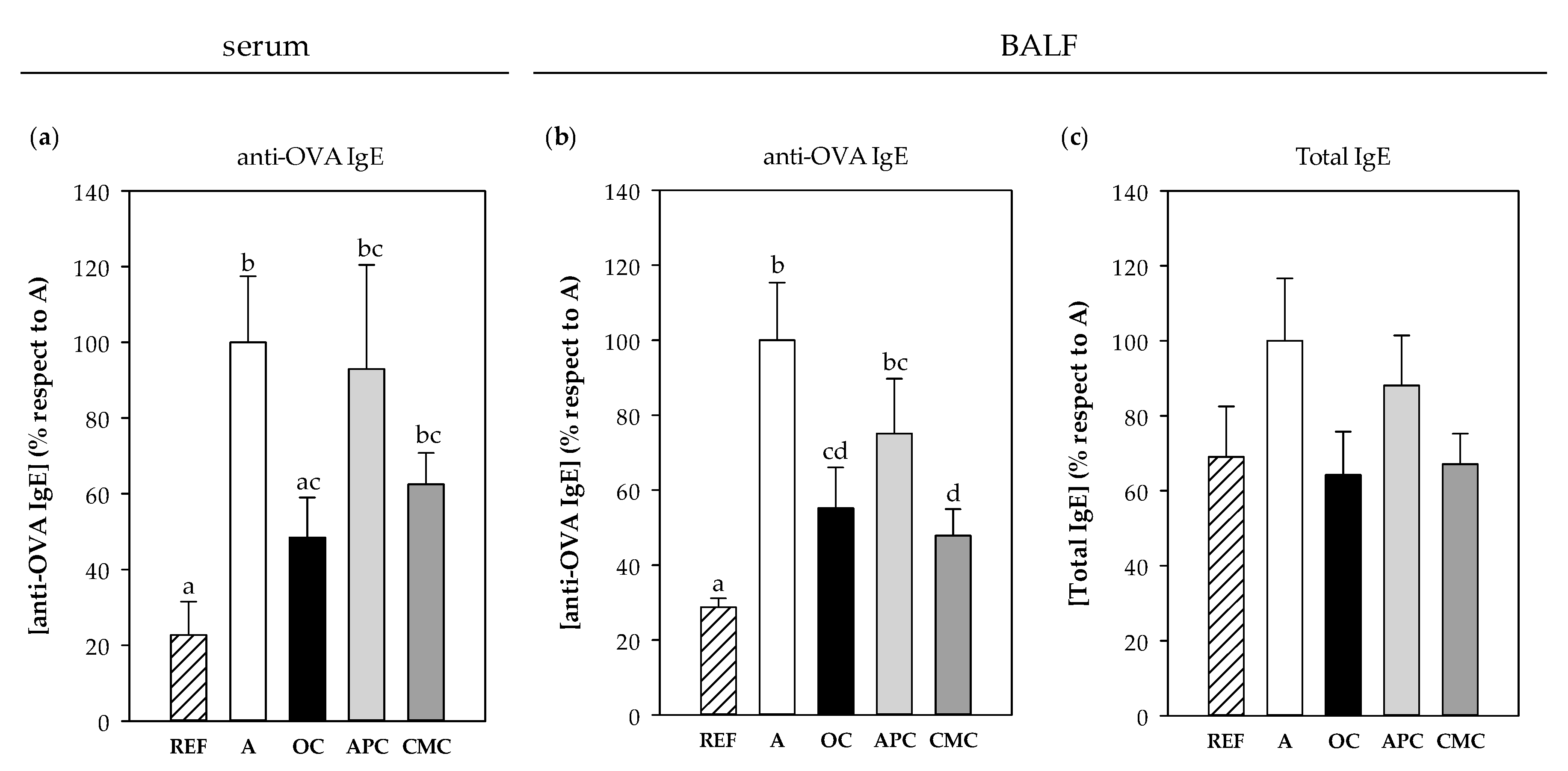
3.3.7. IgE Antibodies
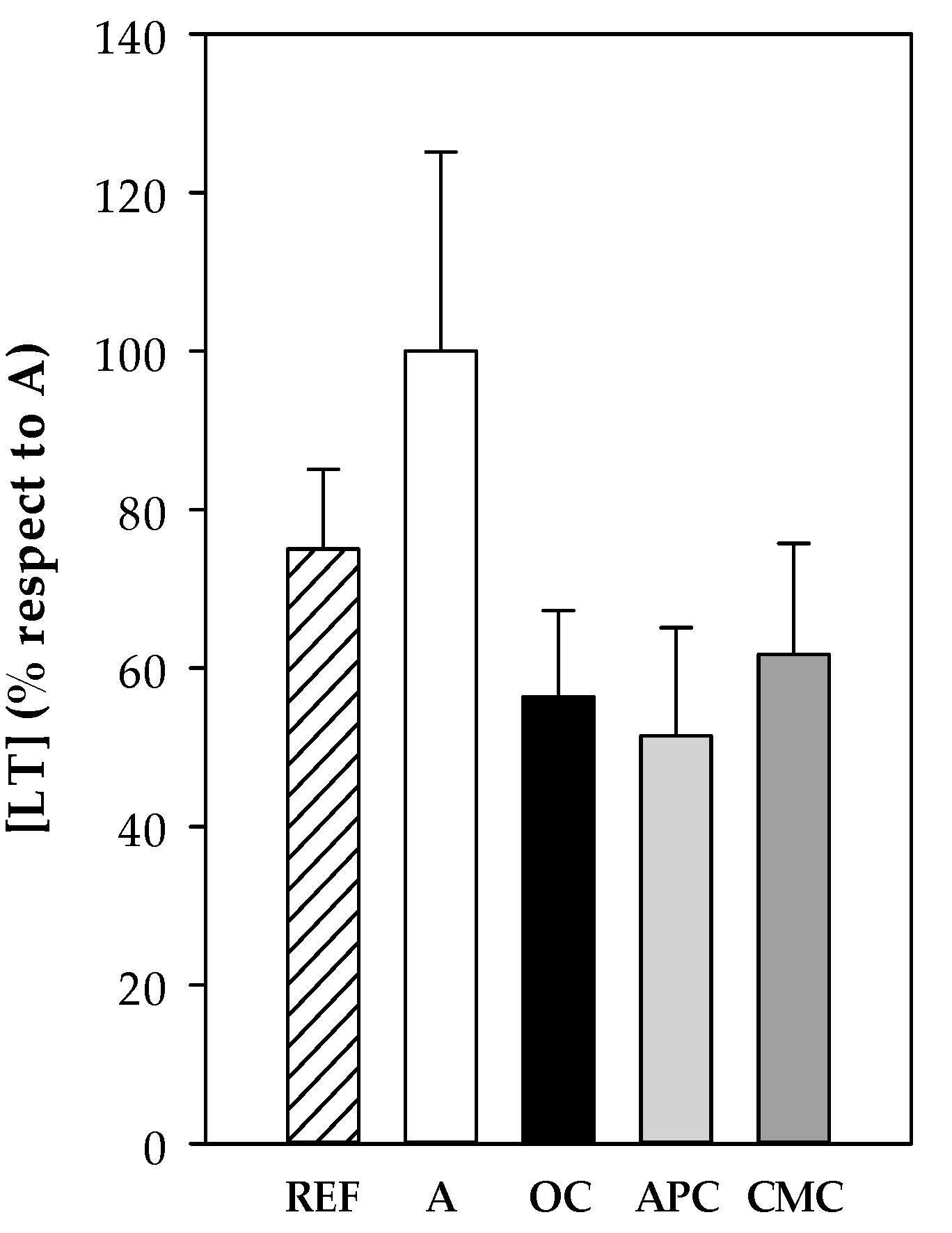
3.3.8. Leukotrienes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Prevention of Allergy and Allergic Asthma; World Health Organization: Geneva, Switzerland, 2003. [Google Scholar]
- Papi, A.; Brightling, C.; Pedersen, S.E.; Reddel, H.K. Asthma. Lancet 2018, 391, 783–800. [Google Scholar] [CrossRef]
- Mishra, V.; Banga, J.; Silveyra, P. Oxidative stress and cellular pathways of asthma and inflammation: Therapeutic strategies and pharmacological targets HHS Public Access. Pharmacol. Ther. 2018, 181, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Curtis, J.L. Cell-mediated Adaptive Immune Defense of the Lungs. Proc. Am. Thorac. Soc. 2005, 2, 412–416. [Google Scholar] [CrossRef] [PubMed]
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef]
- Solé, D.; Aranda, C.S.; Wandalsen, G.F. Asthma: Epidemiology of disease control in Latin America—Short review. Asthma Res. Pract. 2017, 3, 4–9. [Google Scholar] [CrossRef]
- Mallol, J.; Solé, D.; Baeza-Bacab, M.; Aguirre-Camposano, V.; Soto-Quiros, M.; Baena-Cagnani, C. Regional variation in asthma symptom prevalence in Latin American children. J. Asthma 2010, 47, 644–650. [Google Scholar] [CrossRef]
- Castell, M.; Pérez-Cano, F.J.; Abril-Gil, M.; Franch, À. Flavonoids on Allergy. Curr. Pharm. Des. 2014, 20, 973–987. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects. J. Funct. Foods 2015, 1–78. [Google Scholar] [CrossRef]
- Joskova, M.; Sadlonova, V.; Nosalova, G.; Novakova, E.; Franova, S. Polyphenols and their components in experimental allergic asthma. In Respiratory Regulation—The Molecular Approach; Pokorski, M., Ed.; Springer: Dordrecht, The Netherlands, 2013; pp. 91–98. [Google Scholar]
- Ayala-Mata, F.; Barrera-Mendoza, C.C.; Cortés-Rojo, C.; Montoya-Pérez, R.d.C.; García-Pérez, M.E.; Rodríguez-Orozco, A.R. Antioxidants in asthma: Polyphenols. Med. Interna Mex. 2019, 35, 223–234. [Google Scholar]
- Ramiro, E.; Franch, À.; Castellote, C.; Andrés-Lacueva, C.; Izquierdo-Pulido, M.; Castell, M. Effect of Theobroma cacao flavonoids on immune activation of a lymphoid cell line. Br. J. Nutr. 2005, 93, 859–866. [Google Scholar] [CrossRef]
- Ramiro, E.; Franch, A.; Castellote, C.; Pérez-Cano, F.; Permanyer, J.; Izquierdo-Pulido, M.; Castell, M. Flavonoids from Theobroma cacao down-regulate inflammatory mediators. J. Agric. Food Chem. 2005, 53, 8506–8511. [Google Scholar] [CrossRef]
- Ramiro-Puig, E.; Castell, M. Cocoa: Antioxidant and immunomodulator. Br. J. Nutr. 2009, 101, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Castell, M.; Franch, A.; Castellote, C. Effect of a diet rich in cocoa flavonoids on experimental acute inflammation. In Flavonoids: Biosynthesis, Biological Effects and Dietary Sources; Keller, R.B., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2009; Volume 6, pp. 213–229. ISBN 9781607416227. [Google Scholar]
- Massot-Cladera, M.; Franch, À.; Pérez-Cano, F.J.; Castell, M. Cocoa and cocoa fibre differentially modulate IgA and IgM production at mucosal sites. Br. J. Nutr. 2016, 115, 1539–1546. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Camps-Bossacoma, M.; Massot-Cladera, M.; Abril-Gil, M.; Franch, A.; Pérez-Cano, F.J.; Castell, M. Cocoa Diet and Antibody Immune Response in Preclinical Studies. Front. Nutr. 2017, 4, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Abril-Gil, M.; Massot-Cladera, M.; Pérez-Cano, F.J.; Castellote, C.; Franch, A.; Castell, M. A diet enriched with cocoa prevents IgE synthesis in a rat allergy model. Pharmacol. Res. 2012, 65, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Abril-Gil, M.; Pérez-Cano, F.J.; Franch, À.; Castell, M. Effect of a cocoa-enriched diet on immune response and anaphylaxis in a food allergy model in Brown Norway rats. J. Nutr. Biochem. 2016, 27, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Camps-Bossacoma, M.; Abril-Gil, M.; Saldaña-Ruiz, S.; Franch, À.; Pérez-Cano, F.J.; Castell, M. Cocoa diet prevents antibody synthesis and modifies lymph node composition and functionality in a rat oral sensitization model. Nutrients 2016, 8, 242. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Lagunas, M.J.; Vicente, F.; Pereira, P.; Castell, M.; Pérez-Cano, F.J. Relationship between cocoa intake and healthy status: A pilot study in university students. Molecules 2019, 24, 812. [Google Scholar] [CrossRef]
- Febrianto, N.A.; Zhu, F. Diversity in Composition of Bioactive Compounds among 26 Cocoa Genotypes. J. Agric. Food Chem. 2019, 67, 9501–9509. [Google Scholar] [CrossRef]
- Oracz, J.; Zyzelewicz, D.; Nebesny, E. The Content of Polyphenolic Compounds in Cocoa Beans (Theobroma cacao L.), Depending on Variety, Growing Region, and Processing Operations: A Review. Crit. Rev. Food Sci. Nutr. 2015, 55, 1176–1192. [Google Scholar] [CrossRef]
- Bertoldi, D.; Barbero, A.; Camin, F.; Caligiani, A.; Larcher, R. Multielemental fingerprinting and geographic traceability of Theobroma cacao beans and cocoa products. Food Control 2016, 65, 46–53. [Google Scholar] [CrossRef]
- Caligiani, A.; Marseglia, A.; Prandi, B.; Palla, G.; Sforza, S. Influence of fermentation level and geographical origin on cocoa bean oligopeptide pattern. Food Chem. 2016, 211, 431–439. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, R.N.; Grimbs, S.; Behrends, B.; Bernaert, H.; Ullrich, M.S.; Kuhnert, N. Origin-based polyphenolic fingerprinting of Theobroma cacao in unfermented and fermented beans. Food Res. Int. 2017, 99, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.; van Zonneveld, M.; Loo, J.; Hodgkin, T.; Galluzzi, G.; van Etten, J. Present Spatial Diversity Patterns of Theobroma cacao L. in the Neotropics Reflect Genetic Differentiation in Pleistocene Refugia Followed by Human-Influenced Dispersal. PLoS ONE 2012, 7, e47676. [Google Scholar] [CrossRef] [PubMed]
- Zarrillo, S.; Gaikwad, N.; Lanaud, C.; Powis, T.; Viot, C.; Lesur, I.; Fouet, O.; Argout, X.; Guichoux, E.; Salin, F.; et al. The use and domestication of Theobroma cacao during the mid-Holocene in the upper Amazon. Nat. Ecol. Evol. 2018, 2, 1879–1888. [Google Scholar] [CrossRef]
- United Nations Conference on Trade and Development. International Cocoa Agreement; United Nations: Geneva, Switzerland, 2010; ISBN 9780857091253. [Google Scholar]
- Eskes, A.B.; Rodriguez, C.A.C.; Cruz Condori, D.; Seguine, E.; Garcia Carrion, L.F.; Lachenaud, P. Large genetic diversity for fine-flavor traits unveiled in cacao (Theobroma cacao L.) with special attention to the native Chunco variety in Cusco, Peru. Agrotrópica (Itabuna) 2018, 30, 157–174. [Google Scholar] [CrossRef]
- García Carrión, L.F. Catálogo de Cultivares de cacao del Perú; Ministerio de Agricultura y Riego, Dirección General de Competitividad Agraria: Lima, Peru, 2010.
- Secretaria del Convenio sobre la Diversidad Biológica. Protocolo de Nagoya Sobre Acceso a los Recursos Genéticos y Participación Justa y Equitativa en los Beneficios que se Deriven de su Utilización al Convenio Sobre la Diversidad Biológica; Secretaría del Convenio sobre la Diversidad Biológica: Montreal, QC, Canada, 2011; ISBN 92-9225-310-7. [Google Scholar]
- Laird, S.; Wynberg, R.; Rourke, M.; Humphries, F.; Muller, M.R.; Lawson, C. Rethink the expansion of access and benefit sharing. Science 2020, 367, 1200–1202. [Google Scholar] [CrossRef]
- Pedan, V.; Fischer, N.; Rohn, S. An online NP-HPLC-DPPH method for the determination of the antioxidant activity of condensed polyphenols in cocoa. Food Res. Int. 2016, 89, 890–900. [Google Scholar] [CrossRef]
- Shetty, K.; Curtis, O.F.; Levin, R.E.; Witkowsky, R.; Ang, W. Prevention of Vitrification Aßociated with in vitro Shoot Culture of Oregano. (Origanum vulgare) by Pseudomonas spp. J. Plant Physiol. 1995, 147, 447–451. [Google Scholar] [CrossRef]
- Xu, G.; Ye, X.; Chen, J.; Liu, D. Effect of heat treatment on the phenolic compounds and antioxidant capacity of citrus peel extract. J. Agric. Food Chem. 2007, 55, 330–335. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Marinova, D.; Ribarova, F.; Atanassova, M. Total Phenolics and Total Flavonoids in Bulgarian Fruits and Vegetables. J. Univ. Chem. Technol. Metall. 2005, 40, 255–260. [Google Scholar]
- Srdjenovic, B.; Djordjevic-Milic, V.; Grujic, N.; Injac, R.; Lepojevic, Z. Simultaneous HPLC determination of caffeine, theobromine, and theophylline in food, drinks, and herbal products. J. Chromatogr. Sci. 2008, 46, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Romero, S.; Pérez-Cano, F.J.; Ramiro-Puig, E.; Franch, A.; Castell, M. Cocoa intake attenuates oxidative stress associated with rat adjuvant arthritis. Pharmacol. Res. 2012, 66, 207–212. [Google Scholar] [CrossRef]
- Abril-Gil, M.; Garcia-Just, A.; Pérez-Cano, F.J.; Franch, À.; Castell, M. Development and characterization of an effective food allergy model in Brown Norway rats. PLoS ONE 2015, 10, e0125314. [Google Scholar] [CrossRef]
- Estruel-Amades, S.; Massot-Cladera, M.; Garcia-Cerdà, P.; Pérez-Cano, F.J.; Franch, À.; Castell, M.; Camps-Bossacoma, M. Protective effect of hesperidin on the oxidative stress induced by an exhausting exercise in intensively trained rats. Nutrients 2019, 11, 783. [Google Scholar] [CrossRef]
- Wu, L.; Magaz, A.; Maughan, E.; Oliver, N.; Darbyshire, A.; Loizidou, M.; Emberton, M.; Birchall, M.; Song, W. Cellular responses to thermoresponsive stiffness memory elastomer nanohybrid scaffolds by 3D-TIPS. Acta Biomater. 2019, 85, 157–171. [Google Scholar] [CrossRef]
- Yu, T.; Gao, M.; Yang, P.; Liu, D.; Wang, D.; Song, F.; Zhang, X.; Liu, Y. Insulin promotes macrophage phenotype transition through PI3K/Akt and PPAR-γ signaling during diabetic wound healing. J. Cell. Physiol. 2019, 234, 4217–4231. [Google Scholar] [CrossRef]
- Rodríguez-Palmero, M.; Franch, À.; Castell, M.; Pelegrí, C.; Pérez-Cano, F.J.; Kleinschnitz, C.; Stoll, G.; Hünig, T.; Castellote, C. Effective treatment of adjuvant arthritis with a stimulatory CD28-specific monoclonal antibody. J. Rheumatol. 2006, 33, 110–118. [Google Scholar]
- Ramiro-Puig, E.; Pérez-Cano, F.J.; Ramírez-Santana, C.; Castellote, C.; Izquierdo-Pulido, M.; Permanyer, J.; Franch, A.; Castell, M. Spleen lymphocyte function modulated by a cocoa-enriched diet. Clin. Exp. Immunol. 2007, 149, 535–542. [Google Scholar] [CrossRef]
- Périz, M.; Pérez-cano, F.J.; Rodríguez-lagunas, M.J.; Cambras, T. Development and characterization of an allergic asthma rat model for interventional studies. Int. J. Mol. Sci. 2020, 21, 3841. [Google Scholar] [CrossRef]
- Abril-Gil, M.; Garcia-Just, A.; Cambras, T.; Pérez-Cano, F.J.; Castellote, C.; Franch, À.; Castell, M. Motor activity as an unbiased variable to assess anaphylaxis in allergic rats. Exp. Biol. Med. 2015, 240, 1373–1377. [Google Scholar] [CrossRef] [PubMed]
- Jafari, M.; Ansari-Pour, N. Why, when and how to adjust your P values? Cell J. 2019, 20, 604–607. [Google Scholar]
- Ellam, S.; Williamson, G. Cocoa and Human Health. Annu. Rev. Nutr. 2013, 33, 105–128. [Google Scholar] [CrossRef] [PubMed]
- Ferri, C.; Desideri, G.; Ferri, L.; Proietti, I.; Di Agostino, S.; Martella, L.; Mai, F.; Di Giosia, P.; Grassi, D. Cocoa, Blood Pressure, and Cardiovascular Health. J. Agric. Food Chem. 2015, 63, 9901–9909. [Google Scholar] [CrossRef]
- Desch, S.; Schmidt, J.; Kobler, D.; Sonnabend, M.; Eitel, I.; Sareban, M.; Rahimi, K.; Schuler, G.; Thiele, H. Effect of Cocoa Products on blood pressure: Systematic review and meta-analysis. Am. J. Hypertens. 2010, 23, 97–103. [Google Scholar] [CrossRef]
- Seem, S.A.; Yuan, Y.V.; Tou, J.C. Chocolate and chocolate constituents influence bone health and osteoporosis risk. Nutrition 2019, 65, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Andújar, I.; Recio, M.C.; Giner, R.M.; Ríos, J.L. Cocoa polyphenols and their potential benefits for human health. Oxid. Med. Cell. Longev. 2012, 2012, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Pallares Pallares, A.; Estupiñán A, M.R.; Perea Villamil, J.A.; López Giraldo, L.J. Impacto de la fermentación y secado sobre el contenido de polifenoles y capacidad antioxidante del clon de cacao CCN-51. Rev. ION Bucaramanga 2016, 29, 7–21. [Google Scholar]
- Dugo, L.; Belluomo, M.G.; Fanali, C.; Russo, M.; Cacciola, F.; Maccarrone, M.; Sardanelli, A.M. Effect of Cocoa Polyphenolic Extract on Macrophage Polarization from Proinflammatory M1 to Anti-Inflammatory M2 State. Oxidative Med. Cell. Longev. 2017, 2017, 1–11. [Google Scholar] [CrossRef]
- Ramiro-Puig, E.; Pérez-Cano, F.J.; Ramos-Romero, S.; Pérez-Berezo, T.; Castellote, C.; Permanyer, J.; Franch, À.; Izquierdo-Pulido, M.; Castell, M. Intestinal immune system of young rats influenced by cocoa-enriched diet. J. Nutr. Biochem. 2008, 19, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Massot-Cladera, M.; Abril-Gil, M.; Torres, S.; Franch, A.; Castell, M.; Pérez-Cano, F.J. Impact of cocoa polyphenol extracts on the immune system and microbiota in two strains of young rats. Br. J. Nutr. 2014, 112, 1944–1954. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Berezo, T.; Ramiro-Puig, E.; Pérez-Cano, F.J.; Castellote, C.; Permanyer, J.; Franch, A.; Castell, M. Influence of a cocoa-enriched diet on specific immune response in ovalbumin-sensitized rats. Mol. Nutr. Food Res. 2009, 53, 389–397. [Google Scholar] [CrossRef]
- Gibson, S.; Mackeller, A.; Newlands, G.F.J.; Miller, H.R.P. Phenotypic expression of mast cell granule proteinases. Distribution of mast cell proteinases I and II in the rat digestive system. Immunology 1987, 62, 621–627. [Google Scholar] [PubMed]
- Das, M.; Ram, A.; Ghosh, B. Luteolin alleviates bronchoconstriction and airway hyperreactivity in ovalbumin sensitized mice. Inflamm. Res. 2003, 52, 101–106. [Google Scholar]
- Choi, J.R.; Lee, C.M.; Jung, I.D.; Lee, J.S.; Jeong, Y.I.; Chang, J.H.; Park, H.J.; Choi, I.W.; Kim, J.S.; Shin, Y.K.; et al. Apigenin protects ovalbumin-induced asthma through the regulation of GATA-3 gene. Int. Immunopharmacol. 2009, 9, 918–924. [Google Scholar] [CrossRef]
- Tanaka, T.; Takahashi, R. Flavonoids and asthma. Nutrients 2013, 5, 2128–2143. [Google Scholar] [CrossRef]
- Enomoto, T.; Nagasako-Akazome, Y.; Kanda, T.; Ikeda, M.; Dake, Y. Clinical effects of apple polyphenols on persistent allergic rhinitis: A randomized double-blind placebo-controlled parallel arm study. J. Investig. Allergol. Clin. Immunol. 2006, 16, 283–289. [Google Scholar]
- Cooling, D.S. Theophylline toxicity. J. Emerg. Med. 1993, 11, 415–425. [Google Scholar] [CrossRef]
- Kraft, M.; Torvik, J.A.; Trudeau, J.B.; Wenzel, S.E.; Martin, R.J. Theophylline: Potential antiinflammatory effects in nocturnal asthma. J. Allergy Clin. Immunol. 1996, 97, 1242–1246. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OC | BPC | APC | CMC | CCC | |
|---|---|---|---|---|---|
| Total phenolics (mg gallic acid equivalents/g) | 24.11 ± 1.15 ab | 21.07 ± 0.98 a | 28.69 ± 0.80 c | 30.44 ± 0.56 d | 25.30 ± 0.38 b |
| Total flavonoids (mg catechin equivalents/g) | 34.82 ± 1.01 a | 36.59 ± 0.98 a | 50.35 ± 0.95 b | 56.62 ± 1.12 c | 45.60 ± 1.26 d |
| Theobromine (mg/100 g) | 560.75 ± 0.45 a | 491.39 ± 0.93 b | 564.71 ± 0.37 a | 604.19 ± 0.28 d | 573.36 ± 0.30 e |
| Theophylline (mg/100 g) | 1.41 ± 0.00 a | 1.37 ± 0.00 b | 1.40 ± 0.00 c | 1.39 ± 0.00 d | 1.54 ± 0.00 e |
| Caffeine (mg/100 g) | 236.35 ± 0.06 a | 280.74 ± 0.56 b | 275.10 ± 0.11c | 360.53 ± 0.27 d | 324.55 ± 0.15 e |
| FRAP activity (µmol Fe2+/g) | 275.29 ± 12.28 ab | 261.39 ± 14.15 a | 344.31 ± 11.80 c | 309.55 ± 6.7 b | 308.32 ± 4.79 b |
| DPPH activity (µg TEAC/g) | 25.36 ± 0.24 ab | 19.62 ± 0.38 c | 29.77 ± 0.70 a | 25.38 ± 0.94 bd | 24.39 ± 0.07 d |
| Time (days) | REF | A | OC | APC | CMC |
|---|---|---|---|---|---|
| −7 | 57.01 ± 5.04 | 57.52 ± 3.32 | 56.43 ± 3.59 | 56.46 ± 3.54 | 56.97 ± 2.63 |
| −3 | 67.38 ± 5.37 | 68.60 ± 3.57 | 65.37 ± 3.86 | 64.84 ± 3.80 | 64.03 ± 2.90 |
| 0 a | 75.96 ± 5.37 | 76.97 ± 3.55 | 72.44 ± 3.79 | 72.66 ± 4.03 | 71.91 ± 2.79 |
| 4 | 85.07 ± 4.80 | 84.76 ± 3.10 | 79.31 ± 3.65 | 79.21 ± 3.80 | 78.47 ± 2.59 |
| 7 b | 91.57 ± 4.66 | 91.33 ± 3.57 | 82.73 ± 3.50 | 85.10 ± 3.94 | 84.48 ± 2.72 |
| 11 | 98.29 ± 5.52 | 93.70 ± 3.76 | 88.29 ± 3.34 | 89.31 ± 3.99 | 89.43 ± 2.67 |
| 14 | 104.03 ± 4.66 | 99.94 ± 3.74 | 94.59 ± 4.16 | 94.96 ± 4.28 | 94.07 ± 2.82 |
| 18 | 109.49 ± 4.88 | 105.90 ± 3.62 | 97.99 ± 4.68 | 97.98 ± 4.15 | 99.27 ± 2.92 |
| 21 | 115.80 ± 4.86 | 112.00 ± 3.72 | 104.26 ± 4.58 | 104.24 ± 4.49 | 105.77 ± 2.89 |
| 25 | 118.29 ± 5.58 | 115.68 ± 3.62 | 107.66 ± 4.93 | 108.10 ± 4.39 | 107.65 ± 2.52 |
| 28 | 123.73 ± 5.16 | 118.09 ± 2.46 | 109.46 ± 4.52 | 111.95 ± 4.03 | 112.97 ± 2.71 |
| Time (h) | REF | A | OC | APC | CMC |
|---|---|---|---|---|---|
| −2 to 0 | 37.7 ± 0.11 | 37.6 ± 0.10 | 37.8 ± 0.18 | 37.9 ± 0.18 | 37.7 ± 0.13 |
| 0 to 2 | 37.1 ± 0.61 | 36.7 ± 0.29 * | 37.1 ± 0.19 | 36.1 ± 0.44 * | 36.2 ± 0.33 * |
| 2 to 4 | 36.7 ± 0.51 *,a | 35.1 ± 0.58 *,b | 36.2 ± 0.24 *,a | 35.3 ± 0.68 *,a | 35.8 ± 0.32 *,a |
| 4 to 6 | 37.5 ± 0.14 *,a | 35.5 ± 0.41 *,b | 36.2 ± 0.37 *,b | 35.6 ± 0.44 *,b | 36.1 ± 0.43 *,b |
| 6 to 8 | 37.7 ± 0.11 a | 36.2 ± 0.23 *,b | 36.5 ± 0.44 *,b | 36.1 ± 0.28 *,b | 36.3 ± 0.39 *,b |
| 8 to 10 | 37.9 ± 0.11 a | 36.6 ± 0.17 *,b | 36.4 ± 0.58 b | 36.4 ± 0.36 *,b | 36.7 ± 0.37 *,b |
| 10 to 12 | 38.0 ± 0.09 a | 36.9 ± 0.18 *,b | 36.9 ± 0.50 a | 36.5 ± 0.40 *,b | 36.8 ± 0.39 b |
| 12 to 14 | 37.9 ± 0.12 a | 37.1 ± 0.18 *,b | 37.0 ± 0.52 a | 36.8 ± 0.30 *,b | 36.9 ± 0.38 b |
| 14 to 16 | 38.1 ± 0.09 a | 37.2 ± 0.12 *,b | 37.1 ± 0.60 a | 37.2 ± 0.25 *,b | 37.0 ± 0.40 b |
| 16 to 18 | 38.0 ± 0.14 a | 37.3 ± 0.18 *,b | 37.2 ± 0.64 a | 37.4 ± 0.24 a | 37.4 ± 0.27 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Périz, M.; Pérez-Cano, F.J.; Cambras, T.; Franch, À.; Best, I.; Pastor-Soplin, S.; Castell, M.; Massot-Cladera, M. Attenuating Effect of Peruvian Cocoa Populations on the Acute Asthmatic Response in Brown Norway Rats. Nutrients 2020, 12, 2301. https://doi.org/10.3390/nu12082301
Périz M, Pérez-Cano FJ, Cambras T, Franch À, Best I, Pastor-Soplin S, Castell M, Massot-Cladera M. Attenuating Effect of Peruvian Cocoa Populations on the Acute Asthmatic Response in Brown Norway Rats. Nutrients. 2020; 12(8):2301. https://doi.org/10.3390/nu12082301
Chicago/Turabian StylePériz, Marta, Francisco J. Pérez-Cano, Trinitat Cambras, Àngels Franch, Ivan Best, Santiago Pastor-Soplin, Margarida Castell, and Malén Massot-Cladera. 2020. "Attenuating Effect of Peruvian Cocoa Populations on the Acute Asthmatic Response in Brown Norway Rats" Nutrients 12, no. 8: 2301. https://doi.org/10.3390/nu12082301
APA StylePériz, M., Pérez-Cano, F. J., Cambras, T., Franch, À., Best, I., Pastor-Soplin, S., Castell, M., & Massot-Cladera, M. (2020). Attenuating Effect of Peruvian Cocoa Populations on the Acute Asthmatic Response in Brown Norway Rats. Nutrients, 12(8), 2301. https://doi.org/10.3390/nu12082301