Potential Cardiovascular and Metabolic Beneficial Effects of ω-3 PUFA in Male Obesity Secondary Hypogonadism Syndrome

 , , , and
, , , and
Abstract
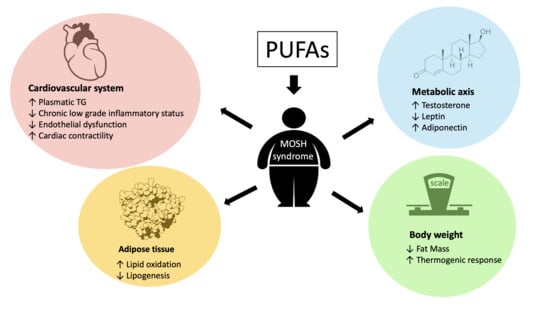
1. Introduction
2. Methods
3. Structure, Metabolic Pathways and Dietary Sources of PUFA
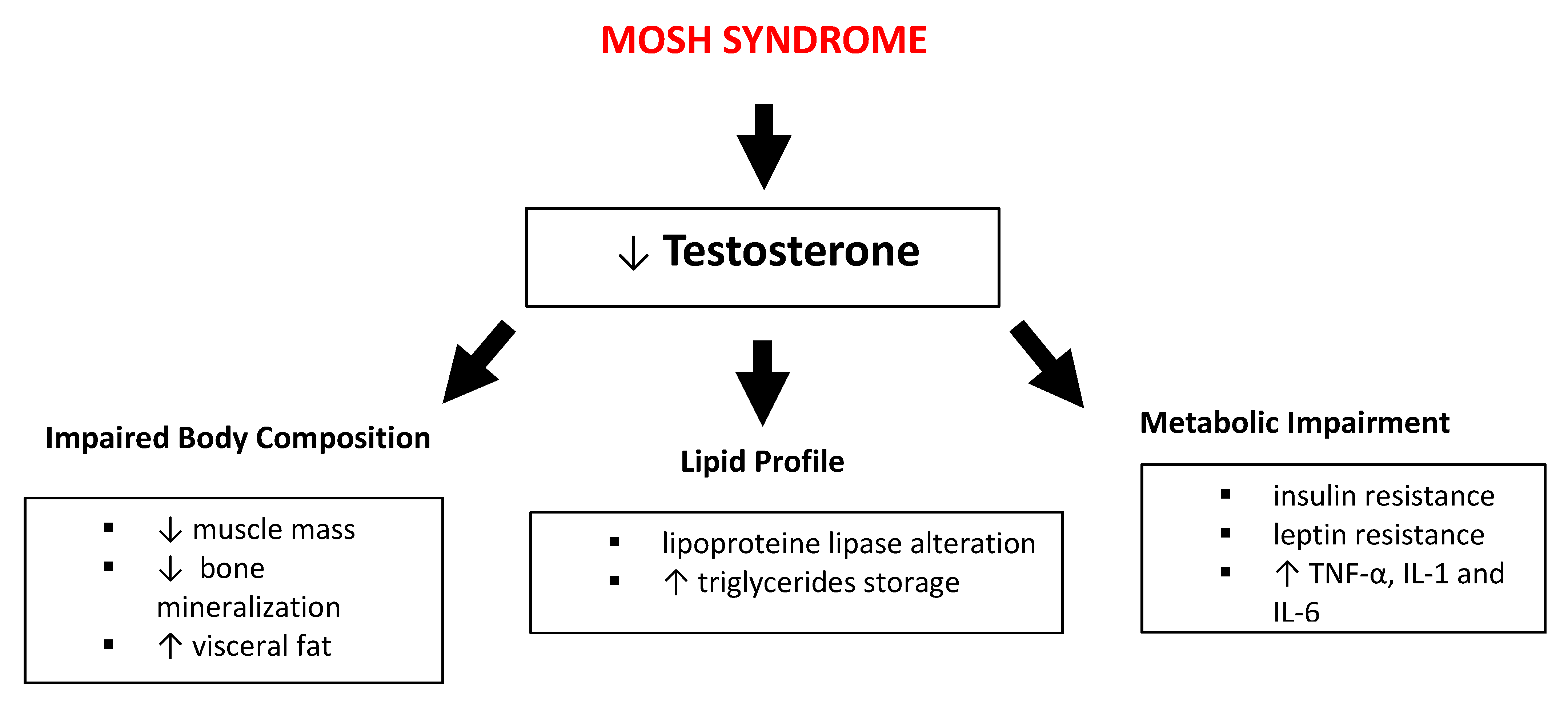
4. Male Obesity Secondary Hypogonadism (MOSH) Syndrome Definition
4.1. Role of PUFA in Cardiovascular Disease
4.2. Impact of PUFA Consumption on Body Weight
4.3. PUFA and Metabolic Axis
5. Summary and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviation List
| AA | Arachidonic acid |
| AHA | American Heart Association |
| Akt | Protein kinase B |
| ALA | α-linoleic acid |
| AMI | Myocardial infarction |
| BAT | Brown adipose tissue |
| BMI | Body mass index |
| CAD | Caspase-activated DNase |
| CAT | Catalase |
| CNCD | Chronic non-communicable degenerative disease |
| CRP | C-reactive protein |
| CV | Cardiovascular |
| CVD | Cardiovascular disease |
| DART | Diet And Reinfarction Trial |
| DGLA | Dihomo-γ-linoleic acid |
| DHA | Docosahexaenoic acid |
| DPA | Docosapentaenoic acid |
| eNOS | Endothelial nitric oxide synthase |
| EPA | Eicosapentaenoic acid |
| FAs | Fatty acids |
| FM | Fat mass |
| FSH | Follicle-stimulating hormone |
| Gal-3 | Galectin-3 |
| GISSI | Gruppo Italiano per lo Studio della Streptochinasi nell’Infarto |
| GLA | γ-linoleic acid |
| GnRH | Gonadotropin-Releasing Hormone |
| HC | Hip circumference |
| HDL | High-density lipoprotein |
| HOMA-IR | Homeostatic model assessment-insulin resistance |
| HT | High triglyceride |
| IL | Interleukin |
| LA | Linoleic acid |
| LDL | Low-density lipoprotein |
| LH | Luteinizing hormone |
| LTS | Leucotriens |
| LXS | Lipoxines |
| MIP | Macrophage inflammatory protein |
| MOSH | Male obesity secondary hypogonadism |
| mTOR | Mammalian target of rapamycin |
| MUFAs | Monounsaturated fatty acids |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| PG | Prostaglandins |
| PPARα | Peroxisome proliferator- activated receptor α |
| PUFA | Polyunsaturated fatty acid |
| S6K1 | Ribosomal protein S6 kinase beta-1 |
| SFAs | Saturated fatty acids |
| SNPs | Single-nucleotide polymorphism |
| SOD | Superoxide dismutase |
| STAT3 | Signal transducer and activator of transcription 3 |
| T | Testosterone |
| T2DM | Type 2 diabetes mellitus |
| TG | Triglycerides |
| TNF-α | Tumor necrosis factor- α |
| Tx | Thromboxanes |
| VCAM-1 | Vascular cell adhesion molecule-1 |
| VHT | Very high triglyceride |
| WAT | White adipose tissue |
| WC | Waist circumference |
| WHO | World Health Organization |
References
- Barnes, S. Nutritional genomics, polyphenols, diets, and their impact on dietetics. J. Am. Diet. Assoc. 2008, 108, 1888–1895. [Google Scholar] [CrossRef] [PubMed]
- Christensen, J.H. Omega-3 polyunsaturated Fatty acids and heart rate variability. Front. Physiol. 2011, 2, 84. [Google Scholar] [CrossRef] [PubMed]
- Raphael, W.; Sordillo, L.M. Dietary polyunsaturated fatty acids and inflammation: The role of phospholipid biosynthesis. Int. J. Mol. Sci. 2013, 14, 21167–21188. [Google Scholar] [CrossRef] [PubMed]
- Bender, N.; Portmann, M.; Heg, Z.; Hofmann, K.; Zwahlen, M.; Egger, M. Fish or n3-PUFA intake and body composition: A systematic review and meta-analysis. Obes. Rev. 2014, 15, 657–665. [Google Scholar] [CrossRef]
- Ofei, F. Obesity—A preventable disease. Ghana. Med. J. 2005, 39, 98–101. [Google Scholar]
- de Mello, A.H.; Uberti, M.F.; de Farias, B.X.; de Souza, N.A.R.; Rezin, G.T. n-3 PUFA and obesity: From peripheral tissues to the central nervous system. Br. J. Nutr. 2018, 119, 1312–1323. [Google Scholar] [CrossRef]
- Obesity, W. Obesity Classification. Available online: https://www.worldobesity.org/about/about-obesity/obesity-classification (accessed on 1 June 2020).
- Cohen, D.A. Obesity and the built environment: Changes in environmental cues cause energy imbalances. Int. J. Obes. 2008, 32 (Suppl. 7), S137–S142. [Google Scholar] [CrossRef]
- Obesity, W. Prevalence of Obesity. Available online: https://www.worldobesity.org/about/about-obesity/prevalence-of-obesity (accessed on 3 April 2020).
- Chooi, Y.C.; Ding, C.; Magkos, F. The epidemiology of obesity. Metabolism 2019, 92, 6–10. [Google Scholar] [CrossRef]
- Knight, J.A. Diseases and disorders associated with excess body weight. Ann. Clin. Lab. Sci. 2011, 41, 107–121. [Google Scholar]
- Kyrou, I.; Randeva, H.S.; Tsigos, C.; Kaltsas, G.; Weickert, M.O. Clinical Problems Caused by Obesity. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., Dungan, K., Grossman, A., Hershman, J.M., Kaltsas, G., Koch, C., Kopp, P., et al., Eds.; Endotext.com: South Dartmouth, MA, USA, 2000. [Google Scholar]
- World Health Organization. Cardiovascular Diseases. Available online: https://www.who.int/health-topics/cardiovascular-diseases#tab=tab_1 (accessed on 1 June 2020).
- Di Daniele, N.; Noce, A.; Vidiri, M.F.; Moriconi, E.; Marrone, G.; Annicchiarico-Petruzzelli, M.; D’Urso, G.; Tesauro, M.; Rovella, V.; De Lorenzo, A. Impact of Mediterranean diet on metabolic syndrome, cancer and longevity. Oncotarget 2017, 8, 8947–8979. [Google Scholar] [CrossRef]
- Canale, M.P.; Manca di Villahermosa, S.; Martino, G.; Rovella, V.; Noce, A.; De Lorenzo, A.; Di Daniele, N. Obesity-related metabolic syndrome: Mechanisms of sympathetic overactivity. Int. J. Endocrinol. 2013, 2013, 865965. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzo, A.; Noce, A.; Moriconi, E.; Rampello, T.; Marrone, G.; Di Daniele, N.; Rovella, V. MOSH Syndrome (Male Obesity Secondary Hypogonadism): Clinical Assessment and Possible Therapeutic Approaches. Nutrients 2018, 10, 474. [Google Scholar] [CrossRef] [PubMed]
- Calderon, B.; Gomez-Martin, J.M.; Vega-Pinero, B.; Martin-Hidalgo, A.; Galindo, J.; Luque-Ramirez, M.; Escobar-Morreale, H.F.; Botella-Carretero, J.I. Prevalence of male secondary hypogonadism in moderate to severe obesity and its relationship with insulin resistance and excess body weight. Andrology 2016, 4, 62–67. [Google Scholar] [CrossRef]
- Anderson, J.L.; May, H.T.; Lappe, D.L.; Bair, T.; Le, V.; Carlquist, J.F.; Muhlestein, J.B. Impact of Testosterone Replacement Therapy on Myocardial Infarction, Stroke, and Death in Men With Low Testosterone Concentrations in an Integrated Health Care System. Am. J. Cardiol. 2016, 117, 794–799. [Google Scholar] [CrossRef] [PubMed]
- Wiktorowska-Owczarek, A.; Berezinska, M.; Nowak, J.Z. PUFAs: Structures, Metabolism and Functions. Adv. Clin. Exp. Med. 2015, 24, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Keum, Y.S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef]
- Das, U.N. Essential Fatty acids—A review. Curr. Pharm. Biotechnol. 2006, 7, 467–482. [Google Scholar] [CrossRef]
- Tvrzicka, E.; Kremmyda, L.S.; Stankova, B.; Zak, A. Fatty acids as biocompounds: Their role in human metabolism, health and disease--a review. Part 1: Classification, dietary sources and biological functions. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc. Czech. Repub. 2011, 155, 117–130. [Google Scholar] [CrossRef]
- Adkins, Y.; Kelley, D.S. Mechanisms underlying the cardioprotective effects of omega-3 polyunsaturated fatty acids. J. Nutr. Biochem. 2010, 21, 781–792. [Google Scholar] [CrossRef]
- Dessi, M.; Noce, A.; Bertucci, P.; Noce, G.; Rizza, S.; De Stefano, A.; Manca di Villahermosa, S.; Bernardini, S.; De Lorenzo, A.; Di Daniele, N. Plasma and erythrocyte membrane phospholipids and fatty acids in Italian general population and hemodialysis patients. Lipids Health Dis. 2014, 13, 54. [Google Scholar] [CrossRef]
- Yang, L.G.; Song, Z.X.; Yin, H.; Wang, Y.Y.; Shu, G.F.; Lu, H.X.; Wang, S.K.; Sun, G.J. Low n-6/n-3 PUFA Ratio Improves Lipid Metabolism, Inflammation, Oxidative Stress and Endothelial Function in Rats Using Plant Oils as n-3 Fatty Acid Source. Lipids 2016, 51, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Li, F.; Li, L.; Fan, J.; Sun, X.; Yin, Y. n-6:n-3 PUFA ratio is involved in regulating lipid metabolism and inflammation in pigs. Br. J. Nutr. 2014, 111, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. Evolutionary aspects of diet, the omega-6/omega-3 ratio and genetic variation: Nutritional implications for chronic diseases. Biomed. Pharmacother. 2006, 60, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Gammone, M.A.; Riccioni, G.; Parrinello, G.; D’Orazio, N. Omega-3 Polyunsaturated Fatty Acids: Benefits and Endpoints in Sport. Nutrients 2018, 11, 46. [Google Scholar] [CrossRef] [PubMed]
- Merida-Ortega, A.; Rothenberg, S.J.; Torres-Sanchez, L.; Schnaas, L.; Hernandez-Alcaraz, C.; Cebrian, M.E.; Garcia-Hernandez, R.M.; Ogaz-Gonzalez, R.; Lopez-Carrillo, L. Polyunsaturated fatty acids and child neurodevelopment among a population exposed to DDT: A cohort study. Environ. Health 2019, 18, 17. [Google Scholar] [CrossRef]
- Liu, W.; Xie, X.; Liu, M.; Zhang, J.; Liang, W.; Chen, X. Serum omega-3 Polyunsaturated Fatty Acids and Potential Influence Factors in Elderly Patients with Multiple Cardiovascular Risk Factors. Sci. Rep. 2018, 8, 1102. [Google Scholar] [CrossRef]
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. EDC-2: The Endocrine Society’s Second Scientific Statement on Endocrine-Disrupting Chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef]
- Fernandez, C.J.; Chacko, E.C.; Pappachan, J.M. Male Obesity-related Secondary Hypogonadism—Pathophysiology, Clinical Implications and Management. Eur. Endocrinol. 2019, 15, 83–90. [Google Scholar] [CrossRef]
- Kelly, D.M.; Jones, T.H. Testosterone: A metabolic hormone in health and disease. J. Endocrinol. 2013, 217, R25–R45. [Google Scholar] [CrossRef]
- Clarke, H.; Dhillo, W.S.; Jayasena, C.N. Comprehensive Review on Kisspeptin and Its Role in Reproductive Disorders. Endocrinol. Metab. 2015, 30, 124–141. [Google Scholar] [CrossRef]
- Roseweir, A.K.; Millar, R.P. The role of kisspeptin in the control of gonadotrophin secretion. Hum. Reprod. Update 2009, 15, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Herbst, K.L.; Bhasin, S. Testosterone action on skeletal muscle. Curr. Opin. Clin. Nutr. Metab. Care 2004, 7, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Giagulli, V.A.; Castellana, M.; Murro, I.; Pelusi, C.; Guastamacchia, E.; Triggiani, V.; De Pergola, G. The Role of Diet and Weight Loss in Improving Secondary Hypogonadism in Men with Obesity with or without Type 2 Diabetes Mellitus. Nutrients 2019, 11, 2975. [Google Scholar] [CrossRef] [PubMed]
- Di Renzo, L.; Gualtieri, P.; Romano, L.; Marrone, G.; Noce, A.; Pujia, A.; Perrone, M.A.; Aiello, V.; Colica, C.; De Lorenzo, A. Role of Personalized Nutrition in Chronic-Degenerative Diseases. Nutrients 2019, 11, 1707. [Google Scholar] [CrossRef] [PubMed]
- Denke, M.A. Diet, lifestyle, and nonstatin trials: Review of time to benefit. Am. J. Cardiol. 2005, 96, 3F–10F. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamid, A.S.; Martin, N.; Bridges, C.; Brainard, J.S.; Wang, X.; Brown, T.J.; Hanson, S.; Jimoh, O.F.; Ajabnoor, S.M.; Deane, K.H.; et al. Polyunsaturated fatty acids for the primary and secondary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2018, 7, CD012345. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, R.; Warnakula, S.; Kunutsor, S.; Crowe, F.; Ward, H.A.; Johnson, L.; Franco, O.H.; Butterworth, A.S.; Forouhi, N.G.; Thompson, S.G.; et al. Association of dietary, circulating, and supplement fatty acids with coronary risk: A systematic review and meta-analysis. Ann. Intern. Med. 2014, 160, 398–406. [Google Scholar] [CrossRef]
- Rhee, J.J.; Kim, E.; Buring, J.E.; Kurth, T. Fish Consumption, Omega-3 Fatty Acids, and Risk of Cardiovascular Disease. Am. J. Prev. Med. 2017, 52, 10–19. [Google Scholar] [CrossRef]
- Siscovick, D.S.; Barringer, T.A.; Fretts, A.M.; Wu, J.H.; Lichtenstein, A.H.; Costello, R.B.; Kris-Etherton, P.M.; Jacobson, T.A.; Engler, M.B.; Alger, H.M.; et al. Omega-3 Polyunsaturated Fatty Acid (Fish Oil) Supplementation and the Prevention of Clinical Cardiovascular Disease: A Science Advisory From the American Heart Association. Circulation 2017, 135, e867–e884. [Google Scholar] [CrossRef]
- Wilber, C.G.; Levine, V.E. Fat metabolism in Alaskan eskimos. Exp. Med. Surg. 1950, 8, 422–425. [Google Scholar]
- Kagawa, Y.; Nishizawa, M.; Suzuki, M.; Miyatake, T.; Hamamoto, T.; Goto, K.; Motonaga, E.; Izumikawa, H.; Hirata, H.; Ebihara, A. Eicosapolyenoic acids of serum lipids of Japanese islanders with low incidence of cardiovascular diseases. J. Nutr. Sci. Vitaminol. 1982, 28, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Bang, H.O.; Dyerberg, J.; Sinclair, H.M. The composition of the Eskimo food in north western Greenland. Am. J. Clin. Nutr. 1980, 33, 2657–2661. [Google Scholar] [CrossRef] [PubMed]
- Peter, S.; Chopra, S.; Jacob, J.J. A fish a day, keeps the cardiologist away!—A review of the effect of omega-3 fatty acids in the cardiovascular system. Indian. J. Endocrinol. Metab. 2013, 17, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Burr, M.L.; Fehily, A.M.; Gilbert, J.F.; Rogers, S.; Holliday, R.M.; Sweetnam, P.M.; Elwood, P.C.; Deadman, N.M. Effects of changes in fat, fish, and fibre intakes on death and myocardial reinfarction: Diet and reinfarction trial (DART). Lancet 1989, 2, 757–761. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J.; American Heart Association; Nutrition, C. Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease. Circulation 2002, 106, 2747–2757. [Google Scholar] [CrossRef]
- Marchioli, R.; Schweiger, C.; Tavazzi, L.; Valagussa, F. Efficacy of n-3 polyunsaturated fatty acids after myocardial infarction: Results of GISSI-Prevenzione trial. Gruppo Italiano per lo Studio della Sopravvivenza nell’Infarto Miocardico. Lipids 2001, 36 (Suppl. 1), S119–S126. [Google Scholar] [CrossRef]
- GISSI-Prevenzione Investigators. Dietary supplementation with n-3 polyunsaturated fatty acids and vitamin E after myocardial infarction: Results of the GISSI-Prevenzione trial. Gruppo Italiano per lo Studio della Sopravvivenza nell’Infarto miocardico. Lancet 1999, 354, 447–455. [Google Scholar] [CrossRef]
- Tavazzi, L.; Maggioni, A.P.; Marchioli, R.; Barlera, S.; Franzosi, M.G.; Latini, R.; Lucci, D.; Nicolosi, G.L.; Porcu, M.; Tognoni, G.; et al. Effect of n-3 polyunsaturated fatty acids in patients with chronic heart failure (the GISSI-HF trial): A randomised, double-blind, placebo-controlled trial. Lancet 2008, 372, 1223–1230. [Google Scholar] [CrossRef]
- Ness, A.R.; Hughes, J.; Elwood, P.C.; Whitley, E.; Smith, G.D.; Burr, M.L. The long-term effect of dietary advice in men with coronary disease: Follow-up of the Diet and Reinfarction trial (DART). Eur. J. Clin. Nutr. 2002, 56, 512–518. [Google Scholar] [CrossRef]
- Zehr, K.R.; Walker, M.K. Omega-3 polyunsaturated fatty acids improve endothelial function in humans at risk for atherosclerosis: A review. Prostaglandins Lipid. Mediat. 2018, 134, 131–140. [Google Scholar] [CrossRef]
- Marion-Letellier, R.; Savoye, G.; Ghosh, S. Polyunsaturated fatty acids and inflammation. IUBMB Life 2015, 67, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Montagnani, M.; Funahashi, T.; Shimomura, I.; Quon, M.J. Adiponectin stimulates production of nitric oxide in vascular endothelial cells. J. Biol. Chem. 2003, 278, 45021–45026. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.; Stone, N.J.; Ballantyne, C.; Bittner, V.; Criqui, M.H.; Ginsberg, H.N.; Goldberg, A.C.; Howard, W.J.; Jacobson, M.S.; Kris-Etherton, P.M.; et al. Triglycerides and cardiovascular disease: A scientific statement from the American Heart Association. Circulation 2011, 123, 2292–2333. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Bulchandani, D. Why do omega-3 fatty acids lower serum triglycerides? Curr. Opin. Lipidol. 2006, 17, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Bays, H.E.; Ballantyne, C.M.; Kastelein, J.J.; Isaacsohn, J.L.; Braeckman, R.A.; Soni, P.N. Eicosapentaenoic acid ethyl ester (AMR101) therapy in patients with very high triglyceride levels (from the Multi-center, plAcebo-controlled, Randomized, double-blINd, 12-week study with an open-label Extension [MARINE] trial). Am. J. Cardiol. 2011, 108, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Kastelein, J.J.; Maki, K.C.; Susekov, A.; Ezhov, M.; Nordestgaard, B.G.; Machielse, B.N.; Kling, D.; Davidson, M.H. Omega-3 free fatty acids for the treatment of severe hypertriglyceridemia: The EpanoVa fOr Lowering Very high triglyceridEs (EVOLVE) trial. J. Clin. Lipidol. 2014, 8, 94–106. [Google Scholar] [CrossRef]
- Maki, K.C.; Orloff, D.G.; Nicholls, S.J.; Dunbar, R.L.; Roth, E.M.; Curcio, D.; Johnson, J.; Kling, D.; Davidson, M.H. A highly bioavailable omega-3 free fatty acid formulation improves the cardiovascular risk profile in high-risk, statin-treated patients with residual hypertriglyceridemia (the ESPRIT trial). Clin. Ther. 2013, 35, 1400–1411.e3. [Google Scholar] [CrossRef]
- Balakumar, P.; Taneja, G. Fish oil and vascular endothelial protection: Bench to bedside. Free Radic. Biol. Med. 2012, 53, 271–279. [Google Scholar] [CrossRef]
- Wang, T.M.; Chen, C.J.; Lee, T.S.; Chao, H.Y.; Wu, W.H.; Hsieh, S.C.; Sheu, H.H.; Chiang, A.N. Docosahexaenoic acid attenuates VCAM-1 expression and NF-kappaB activation in TNF-alpha-treated human aortic endothelial cells. J. Nutr. Biochem. 2011, 22, 187–194. [Google Scholar] [CrossRef]
- Dyerberg, J.; Bang, H.O.; Stoffersen, E.; Moncada, S.; Vane, J.R. Eicosapentaenoic acid and prevention of thrombosis and atherosclerosis? Lancet 1978, 2, 117–119. [Google Scholar] [CrossRef]
- Haglund, O.; Mehta, J.L.; Saldeen, T. Effects of fish oil on some parameters of fibrinolysis and lipoprotein(a) in healthy subjects. Am. J. Cardiol. 1994, 74, 189–192. [Google Scholar] [CrossRef]
- DeFilippis, A.P.; Rai, S.N.; Cambon, A.; Miles, R.J.; Jaffe, A.S.; Moser, A.B.; Jones, R.O.; Bolli, R.; Schulman, S.P. Fatty acids and TxA(2) generation, in the absence of platelet-COX-1 activity. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 428–433. [Google Scholar] [CrossRef]
- Matsumoto, M.; Sata, M.; Fukuda, D.; Tanaka, K.; Soma, M.; Hirata, Y.; Nagai, R. Orally administered eicosapentaenoic acid reduces and stabilizes atherosclerotic lesions in ApoE-deficient mice. Atherosclerosis 2008, 197, 524–533. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 polyunsaturated fatty acids and inflammatory processes: Nutrition or pharmacology? Br. J. Clin. Pharmacol. 2013, 75, 645–662. [Google Scholar] [CrossRef]
- Calder, P.C. n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. Am. J. Clin. Nutr. 2006, 83, 1505S–1519S. [Google Scholar] [CrossRef]
- Egert, S.; Stehle, P. Impact of n-3 fatty acids on endothelial function: Results from human interventions studies. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Casanova, M.A.; Medeiros, F.; Trindade, M.; Cohen, C.; Oigman, W.; Neves, M.F. Omega-3 fatty acids supplementation improves endothelial function and arterial stiffness in hypertensive patients with hypertriglyceridemia and high cardiovascular risk. J. Am. Soc. Hypertens. 2017, 11, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Del-Rio-Navarro, B.E.; Leija-Martinez, J.; Torres-Alcantara, S.; Ruiz-Bedolla, E.; Hernandez-Cadena, L.; Barraza-Villarreal, A.; Romero-Nava, R.; Sanchez-Munoz, F.; Villafana, S.; et al. Effect of omega-3 fatty acids supplementation combined with lifestyle intervention on adipokines and biomarkers of endothelial dysfunction in obese adolescents with hypertriglyceridemia. J. Nutr. Biochem. 2019, 64, 162–169. [Google Scholar] [CrossRef]
- Rantanen, J.M.; Riahi, S.; Johansen, M.B.; Schmidt, E.B.; Christensen, J.H. Effects of Marine n-3 Polyunsaturated Fatty Acids on Heart Rate Variability and Heart Rate in Patients on Chronic Dialysis: A Randomized Controlled Trial. Nutrients 2018, 10, 1313. [Google Scholar] [CrossRef]
- Hidayat, K.; Yang, J.; Zhang, Z.; Chen, G.C.; Qin, L.Q.; Eggersdorfer, M.; Zhang, W. Effect of omega-3 long-chain polyunsaturated fatty acid supplementation on heart rate: A meta-analysis of randomized controlled trials. Eur. J. Clin. Nutr. 2018, 72, 805–817. [Google Scholar] [CrossRef]
- Sagara, M.; Njelekela, M.; Teramoto, T.; Taguchi, T.; Mori, M.; Armitage, L.; Birt, N.; Birt, C.; Yamori, Y. Effects of docosahexaenoic Acid supplementation on blood pressure, heart rate, and serum lipids in Scottish men with hypertension and hypercholesterolemia. Int. J. Hypertens. 2011, 2011, 809198. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.B.; Notay, K.; Klingel, S.L.; Chabowski, A.; Mutch, D.M.; Millar, P.J. Docosahexaenoic acid reduces resting blood pressure but increases muscle sympathetic outflow compared with eicosapentaenoic acid in healthy men and women. Am. J. Physiol. Heart Circ. Physiol. 2019, 316, H873–H881. [Google Scholar] [CrossRef] [PubMed]
- Ghio, S.; Scelsi, L.; Latini, R.; Masson, S.; Eleuteri, E.; Palvarini, M.; Vriz, O.; Pasotti, M.; Gorini, M.; Marchioli, R.; et al. Effects of n-3 polyunsaturated fatty acids and of rosuvastatin on left ventricular function in chronic heart failure: A substudy of GISSI-HF trial. Eur. J. Heart Fail 2010, 12, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Heydari, B.; Abdullah, S.; Pottala, J.V.; Shah, R.; Abbasi, S.; Mandry, D.; Francis, S.A.; Lumish, H.; Ghoshhajra, B.B.; Hoffmann, U.; et al. Effect of Omega-3 Acid Ethyl Esters on Left Ventricular Remodeling After Acute Myocardial Infarction: The OMEGA-REMODEL Randomized Clinical Trial. Circulation 2016, 134, 378–391. [Google Scholar] [CrossRef]
- Laake, K.; Seljeflot, I.; Schmidt, E.B.; Myhre, P.; Tveit, A.; Norseth, J.; Arnesen, H.; Solheim, S. Galectin-3, a marker of cardiac remodeling, is inversely related to serum levels of marine omega-3 fatty acids. A cross-sectional study. JRSM Cardiovasc. Dis. 2017, 6, 2048004017729984. [Google Scholar] [CrossRef]
- Takahashi, Y.; Ide, T. Dietary n-3 fatty acids affect mRNA level of brown adipose tissue uncoupling protein 1, and white adipose tissue leptin and glucose transporter 4 in the rat. Br. J. Nutr. 2000, 84, 175–184. [Google Scholar] [CrossRef]
- Clarke, S.D. Polyunsaturated fatty acid regulation of gene transcription: A mechanism to improve energy balance and insulin resistance. Br. J. Nutr. 2000, 83 (Suppl. 1), S59–S66. [Google Scholar] [CrossRef]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef]
- Di Nunzio, M.; Valli, V.; Bordoni, A. Pro- and anti-oxidant effects of polyunsaturated fatty acid supplementation in HepG2 cells. Prostaglandins Leukot. Essent. Fatty Acids 2011, 85, 121–127. [Google Scholar] [CrossRef]
- CREA. Linee Guida per Una Sana Alimentazione. Available online: https://www.crea.gov.it/web/alimenti-e-nutrizione/-/linee-guida-per-una-sana-alimentazione-2018 (accessed on 15 June 2020).
- Summers, L.K.; Fielding, B.A.; Bradshaw, H.A.; Ilic, V.; Beysen, C.; Clark, M.L.; Moore, N.R.; Frayn, K.N. Substituting dietary saturated fat with polyunsaturated fat changes abdominal fat distribution and improves insulin sensitivity. Diabetologia 2002, 45, 369–377. [Google Scholar] [CrossRef]
- Couet, C.; Delarue, J.; Ritz, P.; Antoine, J.M.; Lamisse, F. Effect of dietary fish oil on body fat mass and basal fat oxidation in healthy adults. Int. J. Obes. Relat. Metab. Disord 1997, 21, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Wang, T.; Heianza, Y.; Zheng, Y.; Sun, D.; Kang, J.H.; Pasquale, L.R.; Rimm, E.B.; Manson, J.E.; Hu, F.B.; et al. Habitual consumption of long-chain n-3 PUFAs and fish attenuates genetically associated long-term weight gain. Am. J. Clin. Nutr. 2019, 109, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, L.K.; Wiener, H.W.; Aslibekyan, S.; Allison, D.B.; Havel, P.J.; Stanhope, K.L.; O’Brien, D.M.; Hopkins, S.E.; Lemas, D.J.; Boyer, B.B.; et al. Linkage and association analysis of obesity traits reveals novel loci and interactions with dietary n-3 fatty acids in an Alaska Native (Yup’ik) population. Metabolism 2015, 64, 689–697. [Google Scholar] [CrossRef]
- Coelho, M.; Oliveira, T.; Fernandes, R. Biochemistry of adipose tissue: An endocrine organ. Arch. Med. Sci. 2013, 9, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Glazier, B.J.; Hinkel, B.C.; Cao, J.; Liu, L.; Liang, C.; Shi, H. Neuroendocrine Regulation of Energy Metabolism Involving Different Types of Adipose Tissues. Int. J. Mol. Sci. 2019, 20, 2707. [Google Scholar] [CrossRef] [PubMed]
- Casas-Agustench, P.; Lopez-Uriarte, P.; Bullo, M.; Ros, E.; Gomez-Flores, A.; Salas-Salvado, J. Acute effects of three high-fat meals with different fat saturations on energy expenditure, substrate oxidation and satiety. Clin. Nutr. 2009, 28, 39–45. [Google Scholar] [CrossRef]
- Itoh, M.; Suganami, T.; Hachiya, R.; Ogawa, Y. Adipose tissue remodeling as homeostatic inflammation. Int. J. Inflam. 2011, 2011, 720926. [Google Scholar] [CrossRef]
- Fuentes, E.; Fuentes, F.; Vilahur, G.; Badimon, L.; Palomo, I. Mechanisms of chronic state of inflammation as mediators that link obese adipose tissue and metabolic syndrome. Mediators. Inflamm. 2013, 2013, 136584. [Google Scholar] [CrossRef]
- Lund, A.S.; Hasselbalch, A.L.; Gamborg, M.; Skogstrand, K.; Hougaard, D.M.; Heitmann, B.L.; Kyvik, K.O.; Sorensen, T.I.; Jess, T. N-3 polyunsaturated fatty acids, body fat and inflammation. Obes. Facts 2013, 6, 369–379. [Google Scholar] [CrossRef]
- Howe, P.; Buckley, J. Metabolic health benefits of long-chain omega-3 polyunsaturated fatty acids. Mil. Med. 2014, 179, 138–143. [Google Scholar] [CrossRef]
- Gray, B.; Steyn, F.; Davies, P.S.; Vitetta, L. Omega-3 fatty acids: A review of the effects on adiponectin and leptin and potential implications for obesity management. Eur. J. Clin. Nutr. 2013, 67, 1234–1242. [Google Scholar] [CrossRef] [PubMed]
- Parra, D.; Ramel, A.; Bandarra, N.; Kiely, M.; Martinez, J.A.; Thorsdottir, I. A diet rich in long chain omega-3 fatty acids modulates satiety in overweight and obese volunteers during weight loss. Appetite 2008, 51, 676–680. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.M.; Halaas, J.L. Leptin and the regulation of body weight in mammals. Nature 1998, 395, 763–770. [Google Scholar] [CrossRef]
- Halaas, J.L.; Gajiwala, K.S.; Maffei, M.; Cohen, S.L.; Chait, B.T.; Rabinowitz, D.; Lallone, R.L.; Burley, S.K.; Friedman, J.M. Weight-reducing effects of the plasma protein encoded by the obese gene. Science 1995, 269, 543–546. [Google Scholar] [CrossRef]
- Auwerx, J.; Staels, B. Leptin. Lancet 1998, 351, 737–742. [Google Scholar] [CrossRef]
- Houseknecht, K.L.; Baile, C.A.; Matteri, R.L.; Spurlock, M.E. The biology of leptin: A review. J. Anim. Sci. 1998, 76, 1405–1420. [Google Scholar] [CrossRef]
- Cameron, A.J.; Welborn, T.A.; Zimmet, P.Z.; Dunstan, D.W.; Owen, N.; Salmon, J.; Dalton, M.; Jolley, D.; Shaw, J.E. Overweight and obesity in Australia: The 1999-2000 Australian Diabetes, Obesity and Lifestyle Study (AusDiab). Med. J. Aust. 2003, 178, 427–432. [Google Scholar] [CrossRef]
- Madsen, E.L.; Bruun, J.M.; Skogstrand, K.; Hougaard, D.M.; Christiansen, T.; Richelsen, B. Long-term weight loss decreases the nontraditional cardiovascular risk factors interleukin-18 and matrix metalloproteinase-9 in obese subjects. Metabolism 2009, 58, 946–953. [Google Scholar] [CrossRef]
- Legradi, G.; Emerson, C.H.; Ahima, R.S.; Flier, J.S.; Lechan, R.M. Leptin prevents fasting-induced suppression of prothyrotropin-releasing hormone messenger ribonucleic acid in neurons of the hypothalamic paraventricular nucleus. Endocrinology 1997, 138, 2569–2576. [Google Scholar] [CrossRef]
- Enriori, P.J.; Sinnayah, P.; Simonds, S.E.; Garcia Rudaz, C.; Cowley, M.A. Leptin action in the dorsomedial hypothalamus increases sympathetic tone to brown adipose tissue in spite of systemic leptin resistance. J. Neurosci. 2011, 31, 12189–12197. [Google Scholar] [CrossRef]
- Finck, B.N.; Johnson, R.W. Tumor necrosis factor-alpha regulates secretion of the adipocyte-derived cytokine, leptin. Microsc. Res. Tech. 2000, 50, 209–215. [Google Scholar] [CrossRef]
- Shamsuzzaman, A.S.; Winnicki, M.; Wolk, R.; Svatikova, A.; Phillips, B.G.; Davison, D.E.; Berger, P.B.; Somers, V.K. Independent association between plasma leptin and C-reactive protein in healthy humans. Circulation 2004, 109, 2181–2185. [Google Scholar] [CrossRef] [PubMed]
- Corsonello, A.; Perticone, F.; Malara, A.; De Domenico, D.; Loddo, S.; Buemi, M.; Ientile, R.; Corica, F. Leptin-dependent platelet aggregation in healthy, overweight and obese subjects. Int. J. Obes. Relat. Metab. Disord. 2003, 27, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Perez-Matute, P.; Perez-Echarri, N.; Martinez, J.A.; Marti, A.; Moreno-Aliaga, M.J. Eicosapentaenoic acid actions on adiposity and insulin resistance in control and high-fat-fed rats: Role of apoptosis, adiponectin and tumour necrosis factor-alpha. Br. J. Nutr. 2007, 97, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.; Wang, Y.; Keshaw, H.; Xu, L.Y.; Lam, K.S.; Cooper, G.J. The fat-derived hormone adiponectin alleviates alcoholic and nonalcoholic fatty liver diseases in mice. J. Clin. Invest. 2003, 112, 91–100. [Google Scholar] [CrossRef]
- Berg, A.H.; Combs, T.P.; Du, X.; Brownlee, M.; Scherer, P.E. The adipocyte-secreted protein Acrp30 enhances hepatic insulin action. Nat. Med. 2001, 7, 947–953. [Google Scholar] [CrossRef]
- Dadson, K.; Liu, Y.; Sweeney, G. Adiponectin action: A combination of endocrine and autocrine/paracrine effects. Front. Endocrinol. 2011, 2, 62. [Google Scholar] [CrossRef]
- Dimiter, D. Effect of omega-3 fatty acids on plasma adiponectin levels in Metabolic syndrome subjects. Inter. J. Obes. 2007, 31. [Google Scholar]
- Whitehouse, A.S.; Tisdale, M.J. Downregulation of ubiquitin-dependent proteolysis by eicosapentaenoic acid in acute starvation. Biochem. Biophys. Res. Commun. 2001, 285, 598–602. [Google Scholar] [CrossRef]
- Noce, A.; Marrone, G.; Rovella, V.; Cusijmano, A.; Di Daniele, N. Beneficial effects of physical activity on uremic sarcopenia. Med. Dello Sport 2018. [Google Scholar] [CrossRef]
- Andrade-Vieira, R.; Han, J.H.; Marignani, P.A. Omega-3 polyunsaturated fatty acid promotes the inhibition of glycolytic enzymes and mTOR signaling by regulating the tumor suppressor LKB1. Cancer Biol. Ther. 2013, 14, 1050–1058. [Google Scholar] [CrossRef] [PubMed]
- Howe, P.; Coates, A.; Murphy, K.; Pettman, T.; Milte, C.; Buckley, J. Higher erythrocyte LCn-3 PUFA content is associated with a healthier body composition. Australas. Med. J. 2014, 1, 60. [Google Scholar]
- Wing, S.S.; Goldberg, A.L. Glucocorticoids activate the ATP-ubiquitin-dependent proteolytic system in skeletal muscle during fasting. Am. J. Physiol. 1993, 264, E668–E676. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Ω-3 Series | Foods |
|---|---|
| ALA |  Green leafy vegetables, nuts, soybeans, flaxseed Green leafy vegetables, nuts, soybeans, flaxseed |
| EPA | Fish oil(herring, salmon, sardine, cod liver), Fish(caviar black and red, herring, salmon) |
| DHA |  Fish oil, herring, caviar black and red, salmon Fish oil, herring, caviar black and red, salmon |
| Ω-6 Series | Foods |
|---|---|
| LA |  Olive oil, seed oil Olive oil, seed oil |
| GLA |  Borage oil, black currant oil Borage oil, black currant oil |
| DGLA |  Human milk Human milk |
| AA |  Meat, dairy, shelfish, human milk Meat, dairy, shelfish, human milk |
| Type of the Study | Reference | Year | Type of Intervention | Primary Outcome | Conclusions |
|---|---|---|---|---|---|
| Animal | Wang, T.M. et al. [63] | 2011 | 40 ApoE (-/-) knockout mice randomized into 5 groups: 1 control group fed normal chow diet, 4 groups fed chow diet supplemented with 200 mg/kg/day of (i) DHA, (ii) EPA, (iii) LA or (iv) AA, for 10 weeks. | DHA supplementation reduced the expression of VCAM-1 in a dose-dependent manner in TNF-α -activated aortic endothelial cells. | DHA supplementation acts at endothelial level by exercising a vasoprotective role. |
| Matsumoto, M. et al. [67] | 2008 | 12 ApoE-deficient mice were fed with Western diet and randomized into two groups: (i) 5% EPA supplementation (ii) without EPA supplementation, for 13 weeks. | The EPA supplementation group showed a reduction in the development of atherosclerotic lesions. Lesions presented a great amount collagen and smooth muscle cells and lower amount of macrophages. | EPA has anti-inflammatory and stabilizing effects on the atherosclerotic plaque. | |
| Human | Rhee J.J. et al. [42] | 2017 | Prospective cohort study on a total of 39,876 women aged ≥45 years without CV diseases subjected to questionnaires on food frequency. | During the follow-up (1993–2014) period there were no associations between the consumption of fish and ω-3 PUFA and CV disease. | The consumption of ω-3 PUFA does not reduce CV risk. |
| GISSI-Prevenzione Investigators [51] | 1999 | Controlled study conducted on 11,324 patients who survived myocardial infarction. They were randomized into 4 groups taking: (i) ω-3 PUFA supplements (1 g per day), (ii) vitamin E (300 mg per day), (iii) both or (iv) neither, for 3–5 years. | The groups with ω-3 PUFA supplementation and with ω-3 PUFA + vitamin E supplementation, presented a reduced risk of death due to CV causes, equally. | Supplementation with ω-3 PUFA reduces the risk of CV mortality. | |
| Bays, H.E. et al. [59] | 2011 | Double blind controlled study conducted on 229 subjects with highly elevated blood levels of triglycerides (≥500 mg/dL), which were randomized into 3 groups: (i) 4 g/day supplementation of EPA ethyl ester (ii) 2 g/day supplementation of EPA ethyl ester or (iii) placebo, for 12 weeks. | The supplementation of 4 g/day EPA ethyl ester reduced triglyceride levels by 33.1% whereas 2 g/day supplementation led to 19.2% reduction, both compared to placebo. | ω-3 PUFA supplementation can be useful to counteract hypertriglyceridemia. | |
| Kastelein, J.J. et al. [60] | 2014 | Double-blind controlled study conducted on subjects with highly elevated blood levels of triglycerides (≥500 mg/dL) which were randomized into 4 groups: (i) control group (4 g/day of olive oil), (ii) 2g/day of ω-3 PUFA, (iii) 3 g/day of ω-3 PUFA, (iv) 4 g/day of ω-3 PUFA, for 12 weeks in combination with a nutrition education program. | 25.9%, 25.5% and 30.9% reduction in blood triglycerides with supplementation of 2, 3 and 4 g/day of ω-3 PUFA respectively, compared to placebo group. | ω-3 PUFA supplementation can be used in lowering hypertriglyceridemia. | |
| Maki, K.C. et al. [61] | 2013 | 647 patients with triglyceride values ≥200 mg/dL and <500 mg/dL were randomized to 6 weeks of treatment with control capsules (4 g/d of olive oil), 2 g/day ω-3 PUFA + 2 g/day olive oil or 4 g/d of ω-3 PUFA. | 14.6% and 20.6% reduction in triglyceride levels and 3.9% and 6.9% reduction in non-HDL cholesterol levels with supplementation of 2 g/day and 4 g/d respectively ω-3 PUFA. | ω-3 PUFA supplementation can be useful in the control of dyslipidemias. | |
| Casanova, M.A. et al. [71] | 2017 | 29 adults with hypertriglyceridemia were divided into: high risk CV patients and low risk CV patients, randomized to take ω -3 PUFA (1800 mg/day) or ciprofibrate (100 mg/day) for 12 weeks. | In high-risk patients, pulse wave velocity was reduced and flow-mediated dilation was increased by ω-3 PUFA. | ω -3 PUFA improved arterial stiffness and endothelial function. | |
| Huang, F. et al. [72] | 2019 | 69 normal weight adolescents and 70 obese adolescents with hypertriglyceridemia were treated with a lifestyle intervention and randomized for ω -3 PUFA or placebo supplementation for 12 weeks. | After 12 weeks of omega-3 supplementation, adolescents showed a significant reduction in triglycerides, leptin, RBP4, ADMA and E-selectin compared to the placebo group and compared to lifestyle intervention alone. | ω-3 PUFA supplementation associated with a healthy lifestyle improves dyslipidemia, insulin resistance and endothelial dysfunction. | |
| Rantanen, J.M. et al. [73] | 2018 | Randomized, double-blind, controlled trial of 112 chronic dialysis patients from Denmark randomized for daily supplementation of 2 g marine ω-3 PUFA or control group, for three months. | In the group with daily supplementation with ω-3 PUFA there was a reduction in heart rate of 2.5 beats per minute, evaluated by 48-h Holter monitoring. | ω-3 PUFA could contribute to vagal modulation that could be protective against malignant ventricular arrhythmias. | |
| Sagara, M. et al. [75] | 2011 | 38 men with arterial hypertension and/or hypercholesterolemia were randomized for a five-week dietary supplement with 2 g of DHA vs active placebo (1 g of olive oil). | Significant reduction in systolic and diastolic blood pressure, heart rate and HDL-C increase in the group with supplementation of DHA. | DHA supplementation can reduce coronary heart disease risk factors. | |
| Lee, J.B. et al. [76] | 2019 | Randomized double-blind study of 86 healthy young men and women to evaluate the effects of oral supplementation of 3 g/day of (i) EPA, (ii) DHA or (iii) olive oil for 12 weeks. | Reduction of systolic and diastolic BP at rest after DHA and olive oil supplementation compared to EPA; DHA supplementation enhances peripheral vasoconstrictor outflow. | DHA supplementation may represent a valid support for patients with high chronic BP. | |
| Ghio, S. et al. [77] | 2010 | 608 patients with chronic symptomatic heart failure were randomized to take (i) ω-3 PUFA or (ii) placebo supplementation. Echocardiography was performed at baseline and after 1, 2 and 3 years. | Left ventricular ejection fraction increase after ω-3 PUFA supplementation of 8.1% at 1 year, 11.1% at 2 years and 11.5% at 3 years. | ω-3 PUFA supplementation can significantly improve left ventricular ejection fraction in patients with symptomatic heart failure. | |
| Tavazzi, L. et al. [52] | 2008 | Controlled, double-blind study in patients with chronic heart failure randomized for the supplementation of 1 g/day of (i) ω-3 PUFA or (ii) placebo, followed for a median of 3.9 years. | 57% of patients in the ω-3 PUFA supplement group, compared to 59% in the placebo group, died or were hospitalized for CV reasons. | ω-3 PUFA supplementation may provide a small benefit in terms of mortality and hospitalization for CV reasons in heart failure patients. | |
| Heydari, B. et al. [78] | 2016 | Controlled, double-blind study on 358 patients after acute MI, randomized for (i) ω-3 PUFA supplementation (4 g/die) or (ii) placebo and underwent baseline assessment by CMR 4-28 days after MI, with 6 months follow-up. | After 6 months of PUFA treatment, the follow-up CMR revealed a significant reduction in left ventricular end-systolic volume indexed and myocardial extra-cellular volume fraction and ST2, fibrosis marker, compared to placebo. | PUFA have an important effect on phenotypes of myocardial tissue after MI. | |
| Laake, L. et al. [79] | 2017 | Evaluation of the relationship between serum level of marine PUFA ω-3 and ω-6 and biomarkers of fibrosis and cardiac remodeling (ST2 and Galectin-3) in 299 elderly patients 2–8 weeks after acute MI. | Galectin-3 levels were inversely related to EPA and DHA and positively related to the ω -6/ω -3 ratio. | ω-3 PUFA display positive effect on cardiac remodeling in acute MI elderly patients. |
| Type of the Study | Reference | Year | Type of Intervention | Primary Outcome | Conclusions |
|---|---|---|---|---|---|
| In vitro | Di Nunzio, M. et al. [83] | 2011 | Supplementation of HepG2 cells with different PUFAs produced various effects on cytotoxicity, oxidation and on antioxidant defenses. | Supplementation with ARA highlighted the induction of oxidative damage, on the contrary, DHA supplementation induced an enhancement in antioxidant defenses. | Each PUFA seems to exert certain actions, on the basis of chemical structure. |
| Human | Summers, L.K. et al. [85] | 2002 | 17 subjects (6 with T2DM, 6 non-obese and 5 obese without T2DM) were randomized in a crossover study to follow two 5-week periods (one period with a diet rich in saturated fatty acids and one period with a diet rich in polyunsaturated fatty acids). | Insulin sensitivity and plasma LDL cholesterol concentrations ameliorated in subjects that followed a diet rich in PUFA compared with the subjects that followed a diet rich in saturated fatty acids. The authors observed a reduction in abdominal subcutaneous fat. | These dietary patterns suggest an improvement in insulin sensitivity, reducing the risk of developing T2DM. |
| Couet, C. et al. [86] | 1997 | Six volunteers were fed with a control diet (52% carbohydrates, 16% protein, 32% fat; no FO) ad libitum for 3 weeks and, 10–12 weeks later, 6 g of fats, of the same diet, were replaced with 6 g/d of FO for a further 3 weeks. | After the dietetic treatment with FO, there was observed a decrease in body fat mass and basal respiratory quotient and an increase of basal lipid oxidation. | Dietary FO causes a reduction of body fat mass and induces lipid oxidation in healthy adults. | |
| Huang, T. et al. [87] | 2019 | The authors tested interactions of ω-3 PUFA habitual consumption and overall genetic susceptibility on long-term weight change. | Food-sourced ω-3 PUFA assumption showed substantial interactions with GRS on long-term changes in body weight. | High intake of ω-3 PUFAs is related to an attenuation of genetic association with long-term weight gain. | |
| Vaughan, L.K. et al. [88] | 2015 | The authors evaluated body composition, plasma adipokines and ghrelin in 982 subjects. They investigated gene–diet interactions. | The authors observed a linkage for all obesity-related traits. They identified new regions of interest for adiponectin and body circumferences. They reported that ω-3 PUFAs are able to modify the link with obesity-related traits. | These authors speculated on the interaction between gene-obesity tract and the pathophysiology obesity. | |
| Casas-Agustench, P. et al. [91] | 2009 | 29 healthy men were randomized in a crossover trial. The authors compared the thermogenic effects of 3 isocaloric sources: (i) high in polyunsaturated fatty acids from walnuts, (ii) high in monounsaturated fatty acids from olive oil and (iii) high in saturated fatty acids from fat-rich dairy products. | Five hours postprandial thermogenesis was greater after the high-polyunsaturated meal (i), and after the high-monounsaturated meal (ii) compared to the high-in-saturated meal (iii). | The thermogenic response was influenced by the fat quality, although the action on substrate oxidation or satiety was unknown. | |
| Lund, A.S. et al. [94] | 2013 | 1212 healthy individuals were enrolled and the authors collected information on nutritional habits associated with different measures of body fat, and inflammatory biomarkers. | Absolute ω-3 PUFA intake presents inverse correlation with anthropometric measures of body fat and among ω-3 PUFA derivatives. In particular, only ALA was inversely associated with body fat measures. | Intake of ω-3 PUFA, in particular ALA, is positively associated with body fat. |
| Type of the Study | Reference | Year | Type of Intervention | Primary Outcome | Conclusions |
|---|---|---|---|---|---|
| In vitro | Andrade-Vieira, R. et al. [116] | 2013 | In vitro tests on MCF7 and HeLaS cell lines. | ω-3 PUFA increases the activation of the Akt-mTOR-S6K1 anabolic pathway. | ω-3 PUFA supplementation can improve cellular metabolism by the promotion of anticatabolite and anabolites production. |
| Perez-Matute, P. et al. [109] | 2007 | Male Wistar rats were fed a high-fat diet for 5 weeks: (i) with oral administration of EPA (1 g/kg) or (ii) without EPA administration. | The increase in body weight and FM was lower in the group treated with EPA. Moreover, EPA administration induced a decrease in food intake and an increase in leptin production and was able to prevent the increase in TNFα. | EPA supplementation can increase the feeling of satiety and reduce the inflammatory state induced by a high-fat diet. | |
| Takahashi, Y. et al. [80] | 2000 | 4 groups of rats were fed for 21 days as follows: (i) low-fat diet with 20 g of safflower oil/kg; (ii) high-fat diet (200 g/kg) rich in ω-6 with safflower oil; (iii) high-fat diet (200 g/kg) rich in ω-3 with perilla; (iv) high-fat diet (200 g/kg) rich in ω-3 with fish oil. | The high-fat diets rich in ω-3, compared to a low-fat diet, did not increase the WAT mass, but increased BAT. Moreover, the diets rich in ω-3, reducing the expression of GLUT-4 mRNA in WAT. | In rats, the gene expression of GLUT-4 mRNA in adipose tissue by ω-3, prevents body fat accumulation and regulates glucose metabolism. | |
| Whitehouse, A.S. et al. [114] | 2001 | Evaluation of the effect of EPA administration on soleus muscle proteolysis during acute fasting in rats compared to control group (olive oil). | Significant reduction of soleus muscle proteolysis in an EPA-treated group and attenuation of the proteasome “chymotryptic-like” enzyme activity. | EPA is able to inhibit protein proteolysis during acute starvation. | |
| Human | Madsen, E.L. et al. [103] | 2009 | Treatment of 68 subjects in abdominal obesity with a low-calorie diet (600–800 kcal/die) for 8 weeks followed by 36 months of randomized treatment with (i) orlistat or (ii) placebo, in association with the lifestyle intervention. | The decrease in body weight is associated with a significant reduction of IL-18, MMP-9 and leptin levels. | Long-term weight loss reduces non-traditional CV risk factors. |
| Parra, D. et al. [97] | 2008 | Appetite monitoring in 233 volunteers during the last 2 weeks of an 8-week low-calorie diet, associated with: (i) low ω-3 intake (<260 mg/die) or (ii) high ω-3 intake (>1300 mg/die). | The evaluation of VAS reveals lower hunger in the high-ω-3 group after dinner after 120 min. | ω-3 intake modulates postprandial satiety in obese and overweight subjects during the weight loss. | |
| Shamsuzzaman, A.S. et al. [107] | 2004 | Association study between plasma leptin and CRP in 100 healthy subjects. | Strong positive and significant association between leptin and CRP, even after adjustment for age, BMI, waist-hip ratio, smoking and alcohol consumption. | The study confirms a strong correlation between metabolic and inflammatory mechanisms. | |
| Dimiter, V. [113] | 2007 | 35 overweight and obese adults with metabolic syndrome were randomized into 2 groups: (i) treated with ω-3 and (ii) treated with placebo, for 3 months on a normal diet, without lifestyle changes. | Increased plasma concentrations of HDL-C and adiponectin and decreased plasma concentrations of triglycerides, HOMA-IR and CRP after ω-3 treatment. | ω-3 PUFA supplementation can improve the inflammatory status and lipid profile through adiponectin-dependent mechanisms in patients with metabolic syndrome. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noce, A.; Marrone, G.; Di Daniele, F.; Di Lauro, M.; Pietroboni Zaitseva, A.; Wilson Jones, G.; De Lorenzo, A.; Di Daniele, N. Potential Cardiovascular and Metabolic Beneficial Effects of ω-3 PUFA in Male Obesity Secondary Hypogonadism Syndrome. Nutrients 2020, 12, 2519. https://doi.org/10.3390/nu12092519
Noce A, Marrone G, Di Daniele F, Di Lauro M, Pietroboni Zaitseva A, Wilson Jones G, De Lorenzo A, Di Daniele N. Potential Cardiovascular and Metabolic Beneficial Effects of ω-3 PUFA in Male Obesity Secondary Hypogonadism Syndrome. Nutrients. 2020; 12(9):2519. https://doi.org/10.3390/nu12092519
Chicago/Turabian StyleNoce, Annalisa, Giulia Marrone, Francesca Di Daniele, Manuela Di Lauro, Anna Pietroboni Zaitseva, Georgia Wilson Jones, Antonino De Lorenzo, and Nicola Di Daniele. 2020. "Potential Cardiovascular and Metabolic Beneficial Effects of ω-3 PUFA in Male Obesity Secondary Hypogonadism Syndrome" Nutrients 12, no. 9: 2519. https://doi.org/10.3390/nu12092519
APA StyleNoce, A., Marrone, G., Di Daniele, F., Di Lauro, M., Pietroboni Zaitseva, A., Wilson Jones, G., De Lorenzo, A., & Di Daniele, N. (2020). Potential Cardiovascular and Metabolic Beneficial Effects of ω-3 PUFA in Male Obesity Secondary Hypogonadism Syndrome. Nutrients, 12(9), 2519. https://doi.org/10.3390/nu12092519