Antidiabetic Effects of Flavan-3-ols and Their Microbial Metabolites



Abstract
1. Introduction
2. Search Criteria
3. Flavan-3-ols: Intake and Metabolism
4. Antidiabetic Effects of Flavan-3-ols: In Vitro and In Vivo Studies
4.1. Glucose Absorption in the Gut
4.2. Insulin Signaling Pathways and Glucose Peripheral Uptake
4.3. Beta Cell Viability and Function
4.4. Endogenous Glucose Production
4.5. Incretin Effect
4.6. Other Mechanisms
5. Antidiabetic Effects of Flavan-3-ols: Clinical Intervention Trials
6. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th ed. Diabetes Res. Clin. Prac. 2019, 157, 107843. [Google Scholar]
- Patterson, C.C.; Dahlquist, G.G.; Gyürüs, E.; Green, A.; Soltész, G.; EURODIAB Study Group. Incidence trends for childhood type 1 diabetes in Europe during 1989–2003 and predicted new cases 2005–2020: A multicentre prospective registration study. Lancet 2009, 373, 2027–2033. [Google Scholar] [PubMed]
- Cersosimo, E.; Triplitt, C.; Solis-Herrera, C.; Mandarino, L.J.; DeFronzo, R.A. Pathogenesis of Type 2 Diabetes Mellitus. In Endotext [Internet]; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., Dungan, K., Grossman, A., Hershman, J.M., Kaltsas, G., Koch, C., Kopp, P., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2018. Available online: https://www.ncbi.nlm.nih.gov/books/NBK279115/ (accessed on 27 February 2018).
- Stein, S.A.; Lamos, E.M.; Davis, S.N. A review of the efficacy and safety of oral antidiabetic drugs. Expert Opin. Drug Saf. 2013, 12, 153–175. [Google Scholar] [CrossRef]
- Vogiatzoglou, A.; Mulligan, A.A.; Lentjes, M.A.H.; Luben, R.N.; Spencer, J.P.E.; Schroeter, H.; Khaw, K.-T.; Kuhnle, G.G.C. Flavonoid Intake in European Adults (18 to 64 Years). PLoS ONE 2015, 10, e0128132. [Google Scholar] [CrossRef]
- Wang, X.; Ouyang, Y.Y.; Liu, J.; Zhao, G. Flavonoid intake and risk of CVD: A systematic review and meta-analysis of prospective cohort studies. Br. J. Nutr. 2014, 111, 1–11. [Google Scholar] [CrossRef]
- Lei, L.; Yang, Y.; He, H.; Chen, E.; Du, L.; Dong, J.; Yang, J. Flavan-3-ols consumption and cancer risk: A meta-analysis of epidemiologic studies. Oncotarget 2016, 7, 73573–73592. [Google Scholar] [PubMed]
- Iso, H.; Date, C.; Wakai, K.; Fukui, M.; Tamakoshi, A.; JACC Study Group. The Relationship between Green Tea and Total Caffeine Intake and Risk for Self-Reported Type 2 Diabetes among Japanese Adults. Ann. Intern. Med. 2006, 144, 554–562. [Google Scholar] [CrossRef] [PubMed]
- The InterAct Consortium. Tea consumption and incidence of type 2 diabetes in Europe: The EPIC-InterAct case-cohort study. PLoS ONE 2012, 7, e36910. [Google Scholar] [CrossRef]
- Nguyen, C.T.; Lee, A.H.; Pham, N.M.; Do, V.V.; Ngu, N.D.; Tran, B.Q.; Binns, C. Habitual tea drinking associated with a lower risk of type 2 diabetes in Vietnamese adults. Asia Pac. J. Clin. Nutr. 2018, 27, 701–706. [Google Scholar] [CrossRef]
- Monagas, M.; Urpi-Sarda, M.; Sánchez-Patán, F.; Llorach, R.; Garrido, I.; Gómez-Cordovés, C.; Andres-Lacueva, C.; Bartolomé, B. Insights into the metabolism and microbial biotransformation of dietary flavan-3-ols and the bioactivity of their metabolites. Food Funct. 2010, 1, 233–253. [Google Scholar] [CrossRef]
- Hellstrom, J.K.; Torronen, A.R.; Mattila, P.H. Proanthocyanidins in common food products of plant origin. J. Agric. Food Chem. 2009, 57, 7899–7906. [Google Scholar] [CrossRef] [PubMed]
- Fruit and Vegetable Phytochemicals: Chemistry Nutritional Value and Stability, 1st ed.; De la Rosa, L.A., Alvarez-Parrilla, E., Gonzalez-Aguilar, G.A., Eds.; Wiley-Blackwell: Ames, IA, USA, 2009. [Google Scholar]
- Knaze, V.; Zamora-Ros, R.; Lujan-Barroso, L.; Romieu, I.; Scalbert, A.; Slimani, N.; Riboli, E.; Van Rossum, C.; Bueno-De-Mesquita, H.B.; Trichopoulou, A.; et al. Intake estimation of total and individual flavan-3-ols, proanthocyanidins and theaflavins, their food sources and determinants in the European Prospective Investigation into Cancer and Nutrition (EPIC) study. Br. J. Nutr. 2011, 108, 1095–1108. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed]
- Crozier, A.; Del Rio, D.; Clifford, M.N. Bioavailability of dietary flavonoids and phenolic compounds. Mol. Asp. Med. 2010, 31, 446–467. [Google Scholar] [CrossRef]
- Selma, M.V.; Espín, J.C.; Tomás-Barberán, F.A. Interaction between Phenolics and Gut Microbiota: Role in Human Health. J. Agric. Food Chem. 2009, 57, 6485–6501. [Google Scholar] [CrossRef] [PubMed]
- Espín, J.C.; González-Sarrías, A.; Tomás-Barberán, F.A. The gut microbiota: A key factor in the therapeutic effects of (poly)phenols. Biochem. Pharmacol. 2017, 139, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xu, P.; Wang, Y.; Wang, Y.; Hochstetter, D. Combined Effects of Green Tea Extracts, Green Tea Polyphenols or Epigallocatechin Gallate with Acarbose on Inhibition against α-Amylase and α-Glucosidase in Vitro. Molecules 2013, 18, 11614–11623. [Google Scholar] [CrossRef] [PubMed]
- Yilmazer-Musa, M.; Griffith, A.M.; Michels, A.J.; Schneider, E.; Frei, B. Inhibition of α-Amylase and α-Glucosidase Activity by Tea and Grape Seed Extracts and their Constituent Catechins. J. Agric. Food Chem. 2012, 60, 8924–8929. [Google Scholar] [CrossRef]
- Wasai, M.; Fujimura, Y.; Nonaka, H.; Kitamura, R.; Murata, M.; Tachibana, H. Postprandial glycaemia-lowering effect of a green tea cultivar Sunrouge and cultivar-specific metabolic profiling for determining bioactivity-related ingredients. Sci. Rep. 2018, 8, 16041. [Google Scholar] [CrossRef]
- Mrabti, H.N.; Jaradat, N.; Fichtali, I.; Ouedrhiri, W.; Jodeh, S.; Ayesh, S.; Cherrah, Y.; Faouzi, M.E.A. Separation, Identification, and Antidiabetic Activity of Catechin Isolated from Arbutus unedo L. Plant Roots. Plants 2018, 7, 31. [Google Scholar] [CrossRef]
- Bräunlich, M.; Slimestad, R.; Wangensteen, H.; Brede, C.; Malterud, K.E.; Barsett, H. Extracts, Anthocyanins and Procyanidins from Aronia melanocarpa as Radical Scavengers and Enzyme Inhibitors. Nutrients 2013, 5, 663–678. [Google Scholar] [CrossRef] [PubMed]
- Tong, T.; Ren, N.; Soomi, P.; Wu, J.; Guo, N.; Kang, H.; Kim, E.; Wu, Y.; He, P.; Tu, Y.; et al. Theaflavins Improve Insulin Sensitivity through Regulating Mitochondrial Biosynthesis in Palmitic Acid-Induced HepG2 Cells. Molecules 2018, 23, 3382. [Google Scholar] [CrossRef] [PubMed]
- Ojelabi, O.A.; Lloyd, K.P.; De Zutter, J.K.; Carruthers, A. Red wine and green tea flavonoids are cis-allosteric activators and competitive inhibitors of glucose transporter 1 (GLUT1)-mediated sugar uptake. J. Biol. Chem. 2018, 293, 19823–19834. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zhao, Y.; Suo, S.; Liu, Y.; Zhao, B. Green tea catechins ameliorate adipose insulin resistance by improving oxidative stress. Free. Rad. Biol. Med. 2012, 52, 1648–1657. [Google Scholar] [CrossRef]
- Takagaki, A.; Yoshioka, Y.; Yamashita, Y.; Nagano, T.; Ikeda, M.; Hara-Terawaki, A.; Seto, R.; Ashida, H. Effects of Microbial Metabolites of (-)-Epigallocatechin Gallate on Glucose Uptake in L6 Skeletal Muscle Cell and Glucose Tolerance in ICR Mice. Biol. Pharm. Bull. 2019, 42, 212–221. [Google Scholar] [CrossRef]
- Scazzocchio, B.; Varì, R.; Filesi, C.; D’Archivio, M.; Santangelo, C.; Giovannini, C.; Iacovelli, A.; Silecchia, G.; Li Volti, G.; Galvano, F.; et al. Cyanidin-3-O-β-glucoside and protocatechuic acid exert insulin-like effects by upregulating PPARγ activity in human omental adipocytes. Diabetes 2011, 60, 2234–2244. [Google Scholar] [CrossRef]
- Álvarez-Cilleros, D.; Martín, M.Á.; Ramos, S. (−)-Epicatechin and the Colonic 2,3-Dihydroxybenzoic Acid Metabolite Regulate Glucose Uptake, Glucose Production, and Improve Insulin Signaling in Renal NRK-52E Cells. Mol. Nutr. Food Res. 2018, 62, 1700470. [Google Scholar] [CrossRef]
- Álvarez-Cilleros, D.; Martín, M.Á.; Ramos, S. Protective effects of (−)-epicatechin and the colonic metabolite 3,4-dihydroxyphenylacetic acid against glucotoxicity-induced insulin signalling blockade and altered glucose uptake and production in renal tubular NRK-52E cells. Food Chem. Toxicol. 2018, 120, 119–128. [Google Scholar] [CrossRef]
- Kaneko, Y.K.; Takii, M.; Kojima, Y.; Yokosawa, H.; Ishikawa, T. Structure-Dependent Inhibitory Effects of Green Tea Catechins on Insulin Secretion from Pancreatic β-Cells. Biol. Pharm. Bull. 2015, 38, 476–481. [Google Scholar] [CrossRef][Green Version]
- Yang, K.; Chan, C. Epicatechin potentiation of glucose-stimulated insulin secretion in INS-1 cells is not dependent on its antioxidant activity. Acta Pharmacol. Sin. 2018, 39, 893–902. [Google Scholar] [CrossRef]
- Koonyosying, P.; Uthaipibull, C.; Fucharoen, S.; Koumoutsea, E.V.; Porter, J.B.; Srichairatanakool, S. Decrement in Cellular Iron and Reactive Oxygen Species, and Improvement of Insulin Secretion in a Pancreatic Cell Line Using Green Tea Extract. Pancreas 2019, 48, 636–643. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Wang, T.; Chen, L.; Yu, B.; Jia, Q.; Chen, K.; Fan, H.; Li, Y.; Wang, H. Trimer procyanidin oligomers contribute to the protective effects of cinnamon extracts on pancreatic β-cells in vitro. Acta Pharmacol. Sin. 2016, 37, 1083–1090. [Google Scholar] [CrossRef]
- Mo, Y.; Lei, J.; Sun, Y.; Zhang, Q.; Wei, G. Conformational Ensemble of hIAPP Dimer: Insight into the Molecular Mechanism by which a Green Tea Extract inhibits hIAPP Aggregation. Sci. Rep. 2016, 6, 33076. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Abedini, A.; Plesner, A.; Verchere, C.B.; Raleigh, D.P. The flavanol (-)-epigallocatechin 3-gallate inhibits amyloid formation by islet amyloid polypeptide, disaggregates amyloid fibrils, and protects cultured cells against IAPP-induced toxicity. Biochemistry 2010, 49, 8127–8133. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.X.; Zhang, Q.; Ma, G.L.; Chen, C.H.; He, Y.M.; Xu, L.H.; Zhang, Y.; Zhou, G.R.; Li, Z.H.; Yang, H.J.; et al. Influence of Aluminium and EGCG on Fibrillation and Aggregation of Human Islet Amyloid Polypeptide. J. Diabetes Res. 2016, 2016, 1–14. [Google Scholar] [CrossRef]
- Fernández-Millán, E.; Ramos, S.; Alvarez, C.; Bravo, L.; Goya, L.; Martín, M.Á. Microbial phenolic metabolites improve glucose-stimulated insulin secretion and protect pancreatic beta cells against tert-butyl hydroperoxide-induced toxicity via ERKs and PKC pathways. Food Chem. Toxicol. 2014, 66, 245–253. [Google Scholar] [CrossRef]
- Carrasco-Pozo, C.; Gotteland, M.; Castillo, R.L.; Chen, C. 3,4-Dihydroxyphenylacetic acid, a microbiota-derived metabolite of quercetin, protects against pancreatic β-cells dysfunction induced by high cholesterol. Exp. Cell Res. 2015, 334, 270–282. [Google Scholar] [CrossRef]
- Bitner, B.F.; Ray, J.D.; Kener, K.B.; Herring, J.A.; Tueller, J.A.; Johnson, D.K.; Tellez Freitas, C.M.; Fausnacht, D.W.; Allen, M.E.; Thomson, A.H.; et al. Common gut microbial metabolites of dietary flavonoids exert potent protective activities in β-cells and skeletal muscle cells. J. Nutr. Biochem. 2018, 62, 95–107. [Google Scholar] [CrossRef]
- Waltner-Law, M.E.; Wang, X.L.; Law, B.K.; Hall, R.K.; Nawano, M.; Granner, D.K. Epigallocatechin Gallate, a Constituent of Green Tea, Represses Hepatic Glucose Production. J. Biol. Chem. 2002, 277, 34933–34940. [Google Scholar] [CrossRef]
- Collins, Q.F.; Liu, H.Y.; Pi, J.; Liu, Z.; Quon, M.J.; Cao, W. Epigallocatechin-3-gallate (EGCG), a green tea polyphenol, suppresses hepatic gluconeogenesis through 5’-AMP-activated protein kinase. J. Boil. Chem. 2007, 282, 30143–30149. [Google Scholar] [CrossRef]
- Yadollah, S.; Kazemipour, N.; Bakhtiyari, S.; Nazifi, S. Palmitate-induced insulin resistance is attenuated by Pioglitazone and EGCG through reducing the gluconeogenic key enzymes expression in HepG2 cells. J. Med. Life 2017, 10, 244–249. [Google Scholar] [PubMed]
- González-Abuín, N.; Martínez-Micaelo, N.; Blay, M.; Green, B.D.; Pinent, M.; Ardévol, A. Grape-seed procyanidins modulate cellular membrane potential and nutrient-induced GLP-1 secretion in STC-1 cells. Am. J. Physiol. Cell Physiol. 2013, 306, C485–C492. [Google Scholar] [CrossRef]
- Kim, J.J.; Tan, Y.; Xiao, L.; Sun, Y.L.; Qu, X. Green tea polyphenol epigallocatechin-3-gallate enhance glycogen synthesis and inhibit lipogenesis in hepatocytes. BioMed. Res. Int. 2013, 2013, 1–8. [Google Scholar] [CrossRef]
- Vazquez-Prieto, M.A.; Bettaieb, A.; Haj, F.G.; Fraga, C.G.; Oteiza, P.I. (−)-Epicatechin prevents TNFα-induced activation of signaling cascades involved in inflammation and insulin sensitivity in 3T3-L1 adipocytes. Arch. Bioch. Biophys. 2012, 527, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Najmanová, I.; Pourová, J.; Vopršalová, M.; Pilarˇová, V.; Semecký, V.; Nováková, L.; Mladeˇnka, P. Flavonoid metabolite 3-(3-hydroxyphenyl) propionic acid formed by human microflora decreases arterial blood pressure in rats. Mol. Nutr. Food Res. 2016, 60, 981–991. [Google Scholar] [CrossRef]
- Qian, Y.; Babu, P.V.A.; Symons, J.D.; Jalili, T. Metabolites of flavonoid compounds preserve indices of endothelial cell nitric oxide bioavailability under glucotoxic conditions. Nutr. Diabetes 2017, 7, e286. [Google Scholar] [CrossRef]
- Álvarez-Cilleros, D.; Ramos, S.; Goya, L.; Martín, M.Á. Colonic metabolites from flavanols stimulate nitric oxide production in human endothelial cells and protect against oxidative stress-induced toxicity and endothelial dysfunction. Food Chem. Toxicol. 2018, 115, 88–97. [Google Scholar] [CrossRef]
- Verzelloni, E.; Pellacani, C.; Tagliazucchi, D.; Tagliaferri, S.; Calani, L.; Costa, L.G.; Brighenti, F.; Borges, G.; Crozier, A.; Conte, A.; et al. Antiglycative and neuroprotective activity of colon-derived polyphenol catabolites. Mol. Nutr. Food Res. 2011, 55, S35–S43. [Google Scholar] [CrossRef]
- Giménez-Bastida, J.A.; Zielinski, H.; Piskula, M.; Zielinska, D.; Szawara-Nowak, D. Buckwheat bioactive compounds, their derived phenolic metabolites and their health benefits. Mol. Nutr. Food Res. 2017, 61, 1600475. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Xue, Y.; Lu, X.; Shao, Q.; Cao, Y.; Wu, Z.; Chen, G. The Effects of Different Degrees of Procyanidin Polymerization on the Nutrient Absorption and Digestive Enzyme Activity in Mice. Molecules 2018, 23, 2916. [Google Scholar] [CrossRef]
- Yamashita, Y.; Okabe, M.; Natsume, M.; Ashida, H. Cacao liquor procyanidin extract improves glucose tolerance by enhancing GLUT4 translocation and glucose uptake in skeletal muscle. J. Nutr. Sci. 2012, 1, E2. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.; Okabe, M.; Natsume, M.; Ashida, H. Cacao liquor procyanidins prevent postprandial hyperglycaemia by increasing glucagon-like peptide-1 activity and AMP-activated protein kinase in mice. J. Nutr. Sci. 2019, 8, E2. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.; Wang, L.; Nanba, F.; Ito, C.; Toda, T.; Ashida, H. Procyanidin Promotes Translocation of Glucose Transporter 4 in Muscle of Mice through Activation of Insulin and AMPK Signaling Pathways. PLoS ONE 2016, 11, e0161704. [Google Scholar] [CrossRef]
- Ueda-Wakagi, M.; Nagayasu, H.; Yamashita, Y.; Ashida, H. Green Tea Ameliorates Hyperglycemia by Promoting the Translocation of Glucose Transporter 4 in the Skeletal Muscle of Diabetic Rodents. Int. J. Mol. Sci. 2019, 20, 2436. [Google Scholar] [CrossRef] [PubMed]
- Cremonini, E.; Bettaieb, A.; Haj, F.G.; Fraga, C.G.; Oteiza, P. (−)-Epicatechin improves insulin sensitivity in high fat diet-fed mice. Arch. Biochem. Biophys. 2016, 599, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Bettaieb, A.; Vazquez Prieto, M.A.; Rodriguez Lanzi, C.; Miatello, R.M.; Haj, F.G.; Fraga, C.G.; Oteiza, P.I. (−)-Epicatechin mitigates high-fructose-associated insulin resistance by modulating redox signaling and endoplasmic reticulum stress. Free Radic. Biol. Med. 2014, 72, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Othman, A.I.; El-Sawi, M.R.; El-Missiry, M.A.; Abukhalil, M.H. Epigallocatechin-3-gallate protects against diabetic cardiomyopathy through modulating the cardiometabolic risk factors, oxidative stress, inflammation, cell death and fibrosis in streptozotocin-nicotinamide-induced diabetic rats. Biomed. Pharmacother. 2017, 94, 362–373. [Google Scholar] [CrossRef]
- Cedó, L.; Castell-Auví, A.; Pallarès, V.; Blay, M.; Ardévol, A.; Pinent, M. Grape Seed Procyanidin Extract Improves Insulin Production but Enhances Bax Protein Expression in Cafeteria-Treated Male Rats. Int. J. Food Sci. 2013, 2013, 875314. [Google Scholar] [CrossRef][Green Version]
- Gan, L.; Meng, Z.; Xiong, R.; Guo, J.; Lu, X.; Zheng, Z.; Deng, Y.; Luo, B.; Zou, F.; Li, H. Green tea polyphenol epigallocatechin-3-gallate ameliorates insulin resistance in non-alcoholic fatty liver disease mice. Acta Pharmacol. Sin. 2015, 36, 597–605. [Google Scholar] [CrossRef]
- Ortsäter, H.; Grankvist, N.; Wolfram, S.; Kuehn, N.; Sjöholm, A. Diet supplementation with green tea extract epigallocatechin gallate prevents progression to glucose intolerance in db/db mice. Nutr. Metab. 2012, 9, 11. [Google Scholar] [CrossRef]
- Franko, A.; Rodriguez Camargo, D.C.; Böddrich, A.; Garg, D.; Rodríguez Camargo, A.; Rathkolb, B.; Janik, D.; Aichler, M.; Feuchtinger, A.; Neff, F.; et al. Epigallocatechin gallate (EGCG) reduces the intensity of pancreatic amyloid fibrils in human islet amyloid polypeptide (hIAPP) transgenic mice. Sci. Rep. 2018, 8, 1116. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Zhen, W.; Yuskavage, J.; Liu, D. Epigallocatechin gallate delays the onset of type 1 diabetes in spontaneous non-obese diabetic mice. Br. J. Nutr. 2011, 105, 1218–1225. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Yuskavage, J.; Liu, D. Dietary flavonol epicatechin prevents the onset of type 1 diabetes in nonobese diabetic mice. J. Agric. Food Chem. 2013, 61, 4303–4309. [Google Scholar] [CrossRef]
- Yamashita, Y.; Okabe, M.; Nasume, M.; Ashida, H. Cinnamtannin A2, a Tetrameric Procyanidin, Increases GLP-1 and Insulin Secretion in Mice. Biosc. Biotech. Biochem. 2013, 77, 888–891. [Google Scholar] [CrossRef] [PubMed]
- Samarghandian, S.; Azimi-Nezhad, M.; Farkhondeh, T. Catechin Treatment Ameliorates Diabetes and Its Complications in Streptozotocin-Induced Diabetic Rats. Dose-Response 2017, 15, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Haidari, F.; Omidian, K.; Rafiei, H.; Zarei, M.; Mohamad Shahi, M. Green Tea (Camellia sinensis) Supplementation to Diabetic Rats Improves Serum and Hepatic Oxidative Stress Markers. IJPR 2013, 12, 109–114. [Google Scholar] [PubMed]
- De Sandoval-Muñiz, R.J.; Vargas-Guerrero, B.; Flores-Alvarado, L.J.; Gurrola-Díaz, C.M. Glucotransporters: Clinical, molecular and genetic aspects. Gac. Med. Mex. 2016, 152, 547–557. [Google Scholar]
- Hajiaghaalipour, F.; Khalilpourfarshbafi, M.; Arya, A. Modulation of Glucose Transporter Protein by Dietary Flavonoids in Type 2 Diabetes Mellitus. Int. J. Biol. Sci. 2015, 11, 508–524. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Cohrs, C.M.; Stertmann, J.; Bozsak, R.; Speier, S. Human beta cell mass and function in diabetes: Recent advances in knowledge and technologies to understand disease pathogenesis. Mol. Metab. 2017, 6, 943–957. [Google Scholar] [CrossRef]
- Haataja, L.; Gurlo, T.; Huang, C.J.; Butler, P.C. Islet amyloid in type 2 diabetes, and the toxic oligomer hypothesis. Endocr. Rev. 2008, 29, 303–316. [Google Scholar]
- Del Prato, S. Role of glucotoxicity and lipotoxicity in the pathophysiology of Type 2 diabetes mellitus and emerging treatment strategies. Diab. Med. 2009, 26, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Cersosimo, E.; Solis-Herrera, C.; Trautmann, M.E.; Malloy, J.; Triplitt, C.L. Assessment of pancreatic β-cell function: Review of methods and clinical applications. Curr. Diabetes Rev. 2014, 10, 2–42. [Google Scholar] [CrossRef] [PubMed]
- Hatting, M.; Tavares, C.D.; Sharabi, K.; Rines, A.K.; Puigserver, P. Insulin regulation of gluconeogenesis. Ann. N. Y. Acad. Sci. 2018, 1411, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M.A.; Meier, J.J. Incretin hormones: Their role in health and disease. Diabetes Obes. Metab. 2018, 1, 5–21. [Google Scholar] [CrossRef]
- Strat, K.M.; Rowley, T.J.; Smithson, A.T.; Tessem, J.S.; Hulver, M.W.; Liu, D.; Davy, B.M.; Davy, K.P.; Neilson, A.P. Mechanisms by which cocoa flavanols improve metabolic syndrome and related disorders. J. Nutr. Biochem. 2016, 35, 1–21. [Google Scholar] [CrossRef]
- Kohlgruber, A.; Lynch, L. Adipose tissue inflammation in the pathogenesis of type 2 diabetes. Curr. Diabetes Rep. 2015, 15, 92. [Google Scholar] [CrossRef]
- Petrie, J.R.; Guzik, T.J.; Touyz, R.M. Diabetes, Hypertension, and Cardiovascular Disease: Clinical Insights and Vascular Mechanisms. Can. J. Cardiol. 2018, 34, 575–584. [Google Scholar] [CrossRef]
- Vlassara, H.; Uribarri, J. Advanced glycation end products (AGE) and diabetes: Cause, effect, or both? Curr. Diabetes Rep. 2014, 14, 453. [Google Scholar] [CrossRef]
- Zhang, H.; Su, S.; Yu, X.; Li, Y. Dietary epigallocatechin 3-gallate supplement improves maternal and neonatal treatment outcome of gestational diabetes mellitus: A double-blind randomised controlled trial. J. Hum. Nutr. Diet. 2017, 30, 753–758. [Google Scholar] [CrossRef]
- Grassi, D.; Desideri, G.; Necozione, S.; Lippi, C.; Casale, R.; Properzi, G.; Blumberg, J.B.; Ferri, C. Blood pressure is reduced and insulin sensitivity increased in glucose-intolerant, hypertensive subjects after 15 days of consuming high-polyphenol dark chocolate. J. Nutr. 2008, 138, 1671–1676. [Google Scholar] [CrossRef]
- Curtis, P.J.; Sampson, M.; Potter, J.; Dhatariya, K.; Kroon, P.A.; Cassidy, A. Chronic ingestion of flavan-3-ols and isoflavones improves insulin sensitivity and lipoprotein status and attenuates estimated 10-year CVD risk in medicated postmenopausal women with type 2 diabetes: A 1-year, double-blind, randomized, controlled trial. Diabetes Care 2012, 35, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-H.; Liao, Y.-L.; Lin, S.-C.; Tsai, T.-H.; Huang, C.-J.; Chou, P. Does supplementation with green tea extract improve insulin resistance in obese type 2 diabetics? A randomized, double-blind, and placebo-controlled clinical trial. Altern. Med. Rev. 2011, 16, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Nagao, T.; Meguro, S.; Hase, T.; Otsuka, K.; Komikado, M.; Tokimitsu, I.; Yamamoto, T.; Yamamoto, K. A catechin-rich beverage improves obesity and blood glucose control in patients with type 2 diabetes. Obesity 2008, 17, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Fukino, Y.; Ikeda, A.; Maruyama, K.; Aoki, N.; Okubo, T.; Iso, H. Randomized controlled trial for an effect of green tea-extract powder supplementation on glucose abnormalities. Eur. J. Clin. Nutr. 2008, 62, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Fukino, Y.; Shimbo, M.; Aoki, N.; Okubo, T.; Iso, H. Randomized controlled trial for an effect of green tea consumption on insulin resistance and inflammation markers. J. Nutr. Sci. Vitaminol. 2005, 51, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Ryu, O.H.; Lee, J.; Lee, K.W.; Kim, H.Y.; Seo, J.A.; Kim, S.G.; Kim, N.H.; Baik, S.-H.; Choi, D.S.; Choi, K.M. Effects of green tea consumption on inflammation, insulin resistance and pulse wave velocity in type 2 diabetes patients. Diabetes Res. Clin. Pract. 2006, 71, 356–358. [Google Scholar] [CrossRef]
- Kar, P.; Laight, D.; Rooprai, H.K.; Shaw, K.M.; Cummings, M. Effects of grape seed extract in Type 2 diabetic subjects at high cardiovascular risk: A double blind randomized placebo controlled trial examining metabolic markers, vascular tone, inflammation, oxidative stress and insulin sensitivity. Diabet. Med. 2009, 26, 526–531. [Google Scholar] [CrossRef]
- Dicks, L.; Kirch, N.; Gronwald, D.; Wernken, K.; Zimmermann, B.F.; Helfrich, H.P.; Ellinger, S. Regular Intake of a Usual Serving Size of Flavanol-Rich Cocoa Powder Does Not Affect Cardiometabolic Parameters in Stably Treated Patients with Type 2 Diabetes and Hypertension-A Double-Blinded, Randomized, Placebo-Controlled Trial. Nutrients 2018, 10, 1435. [Google Scholar] [CrossRef]
- Stote, K.S.; Clevidence, B.A.; Novotny, J.A.; Henderson, T.; Radecki, S.V.; Baer, D.J. Effect of cocoa and green tea on biomarkers of glucose regulation, oxidative stress, inflammation and hemostasis in obese adults at risk for insulin resistance. Eur. J. Clin. Nutr. 2012, 66, 1153–1159. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Betts, N.M.; Leyva, M.J.; Fu, D.; Aston, C.E.; Lyons, T.J. Acute Cocoa Supplementation Increases Postprandial HDL Cholesterol and Insulin in Obese Adults with Type 2 Diabetes after Consumption of a High-Fat Breakfast. J. Nutr. 2015, 145, 2325–2332. [Google Scholar] [CrossRef]
- Rynarzewski, J.; Dicks, L.; Zimmermann, B.F.; Stoffel-Wagner, B.; Ludwig, N.; Helfrich, H.P.; Ellinger, S. Impact of a Usual Serving Size of Flavanol-Rich Cocoa Powder Ingested with a Diabetic-Suitable Meal on Postprandial Cardiometabolic Parameters in Type 2 Diabetics-A Randomized, Placebo-Controlled, Double-Blind Crossover Study. Nutrients 2019, 11, 417. [Google Scholar] [CrossRef]
- Del Rio, D.; Calani, L.; Cordero, C.; Salvatore, S.; Pellegrini, N.; Brighenti, F. Bioavailability and catabolism of green tea flavan-3-ols in humans. Nutrition 2010, 26, 1110–1116. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, J.I.; Borges, G.; Momma, T.Y.; Spencer, J.P.E.; Keen, C.L.; Crozier, A.; Schroeter, H. The metabolome of [2-(14) C] (-)-epicatechin in humans: Implications for the assessment of efficacy, safety, and mechanisms of action of polyphenolic bioactives. Sci. Rep. 2016, 6, 29034. [Google Scholar] [CrossRef] [PubMed]
- Urpi-Sarda, M.; Monagas, M.; Khan, N.; Llorach, R.; Lamuela-Raventós, R.M.; Jáuregui, O.; Estruch, R.; Izquierdo-Pulido, M.; Andrés-Lacueva, C. Targeted metabolic profiling of phenolics in urine and plasma after regular consumption of cocoa by liquid chromatography–tandem mass spectrometry. J. Chromatogr. A 2009, 1216, 7258–7267. [Google Scholar] [CrossRef] [PubMed]
- Wiese, S.; Esatbeyoglu, T.; Winterhalter, P.; Kruse, H.-P.; Winkler, S.; Bub, A.; Kulling, S.E. Comparative biokinetics and metabolism of pure monomeric, dimeric, and polymeric flavan-3-ols: A randomized cross-over study in humans. Mol. Nutr. Food Res. 2015, 59, 610–621. [Google Scholar] [PubMed]
- Rodriguez-Mateos, A.; Feliciano, R.P.; Boeres, A.; Weber, T.; Dos Santos, C.N.; Ventura, M.R.; Heiss, C. Cranberry (poly)phenol metabolites correlate with improvements in vascular function: A double-blind, randomized, controlled, dose-response, crossover study. Mol. Nutr. Food Res. 2016, 60, 2130–2140. [Google Scholar] [CrossRef]
- Castello, F.; Costabile, G.; Bresciani, L.; Tassotti, M.; Naviglio, D.; Luongo, D.; Ciciola, P.; Vitale, M.; Vetrani, C.; Galaverna, G.; et al. Bioavailability and pharmacokinetic profile of grape pomace phenolic compounds in humans. Arch. Biochem. Biophys. 2018, 646, 1–9. [Google Scholar]
- Feliciano, R.P.; Boeres, A.; Massacessi, L.; Istas, G.; Ventura, M.R.; Nunes dos Santos, C.; Heiss, C.; Rodriguez-Mateos, A. Identification and quantification of novel cranberry-derived plasma and urinary (poly) phenols. Arch. Biochem. Biophys. 2016, 599, 31–41. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Martini, S.; Conte, A. Protocatechuic and 3,4-Dihydroxyphenylacetic Acids Inhibit Protein Glycation by Binding Lysine through a Metal-Catalyzed Oxidative Mechanism. J. Agric. Food Chem. 2019, 67, 7821–7831. [Google Scholar] [CrossRef]
- Pourová, J.; Najmanová, I.; Vopršalová, M.; Migkos, T.; Pilařová, V.; Applová, L.; Nováková, L.; Mladěnka, P. Two flavonoid metabolites, 3,4-dihydroxyphenylacetic acid and 4-methylcatechol, relax arteries ex vivo and decrease blood pressure in vivo. Vascul. Pharmacol. 2018, 111, 36–43. [Google Scholar] [CrossRef]


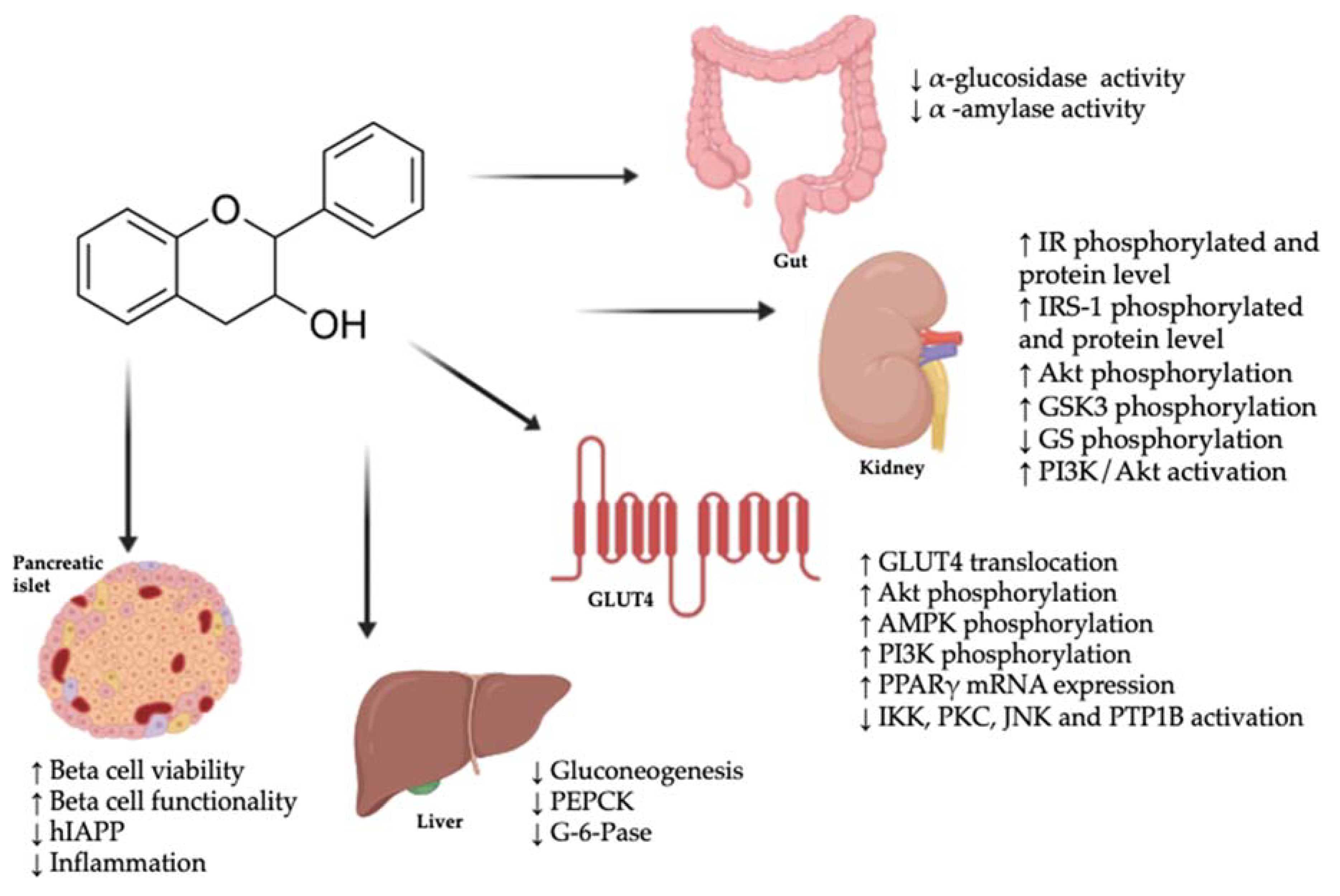{kind=link}
| In Vitro Test | Flavan-3-ol | Concentration/Dose | Results | Ref. |
|---|---|---|---|---|
| Glucose absorption in the gut | ||||
| Inhibition of α-glucosidase and α-amylase activity | GTE, GTP, EGCG | α-amylase: IC50 = 1370.812 ± 59.081–1849.612 ± 73.475 µg/mL α-glucosidase: IC50 = 4.421 ± 0.018–10.019 ± 0.017 µg/mL | Inhibition of α-glucosidase by GTE was stronger than by acarbose (IC50 = 4822.783 ± 26.042 µg/mL) and the other compounds but had no effect on α-amylase. | [19] |
| Combination of GTE, GTP, EGCG, and acarbose at low concentrations had synergistic suppressive effects on α-glucosidase. | ||||
| α-amylase was inhibited at high concentrations of GTP and EGCG, but lower than that of acarbose (IC50 = 2715.654 ± 24.709 µg/mL). | ||||
| Inhibition of α-amylase and α-glucosidase activity | GSE, tea extracts, C, EC, EGC, EGCG, GCG, ECG | α-amylase: IC50 = 8.7 ± 0.8–378 ± 134 µg/L α-glucosidase: IC50 = 0.3 ± 0.1–31 µg/L | α-amylase was only inhibited by GTE extract similarly to acarbose. | [20] |
| α-glucosidase was significantly inhibited by all compounds except C, EC following this order: Teavigo® > EGCG > GTE> GSE > GCG > WTE > ECG | ||||
| Inhibition of α-glucosidase activity | EGCG, ECG, EGCG3”Me, ECG3”Me | IC50 = 8.1−61.1 µM | Inhibition of α-glucosidase EGCG3”Me > EGCG > ECG3”Me > ECG | [21] |
| α-glucosidase inhibition assay | C | IC50 = 87.55 µg/mL | The α-glucosidase inhibitory potency was greater than acarbose (IC50 = 199.53 µg/mL). | [22] |
| Inhibition of α-glucosidase activity | Procyanidins B2, B5 and C1 | IC50 = 4.7 ± 0.2, 5.5 ± 0.1 and 3.8 ± 0.2 µg/mL | Trimeric procyanidin (C1) exerted the strongest inhibitory activity. Inhibitory effect was stronger than for acarbose (130.0 ± 20.0 µg/mL). | [23] |
| Insulin signaling pathways and glucose peripheral uptake | ||||
| Glucose uptake assay and insulin signaling pathway in HepG2 cells treated with PA | Theaflavin mixture (TF, TF-3-G, TF-3′-G, and TFDG) | 2.5–10 μg/mL | Increased 2-NBDG uptake. Increased membrane bound GLUT4 protein level and Akt phosphorylation. Decreased IRS-1 phosphorylation at Ser307. Increase of mtDNA copy number. Downregulation of PGC-1β mRNA level and increase of PRC mRNA expression. | [24] |
| GLUT1-mediated uptake of 3-O-methylglucose in human red blood cells | EGCG and ECG | - | Uptake of 0.1 mM 3MG was dose-dependently inhibited. | [25] |
| Glucose uptake, GLUT4 translocation, and JNK phosphorylation in insulin resistant 3T3-L1 adipocytes | EGCG | 0.1–5 µM | At 5 µM, increased glucose uptake. Dose-dependent reversion of Dex- and TNFα-induced JNK phosphorylation. At 1 µM, increased GLUT4 translocation. | [26] |
| GLUT4 translocation in L6 skeletal muscle cells | 5-(3,5-dihydroxyphenyl)-γ-VL | 1 and 3 µM | 3 µM promoted the strongest effect on GLUT4 translocation. AMPK phosphorylation increased. | [27] |
| Glucose transport in human and murine 3T3-L1 adipocytes stimulated or not with insulin | PCA | 100 µmol/L | Reversion of oxLDL-induced decrease in glucose uptake and GLUT4 translocation. Reversion of oxLDL-induced decrease of adiponectin mRNA expression and secretion, and of PPARγ mRNA expression and activity. | [28] |
| Insulin signaling, glucose uptake, and glucose production in rat renal NRK-52E cells | EC, 2,3-DHB, 3,4-DHPA, 3-HPP and VA | 5–20 µM | Glucose uptake, glucose production, and PEPCK reduced after treatment with EC (5–20 µM) and 2,3-DBH (20 µM). | [29] |
| IR and IRS-1 phosphorylated and total protein levels increased at 10 µM EC and 20 µM 2,3-DHB. Increased phosphorylation of Akt and GSK3. The inhibition of the PI3K/Akt pathway was restrained. | ||||
| Insulin signaling and glucose uptake and production in rat renal NRK-52E cells treated with high glucose | EC, 3,4-DHPA, 2,3-DHB and 3-HPP | 5–20 µM | The altered glucose uptake and production caused by high glucose was prevented by EC (5–20 µM) and 3,4-DHPA (10–20 µM). At 10 µM, tyrosine phosphorylated, and total levels of IR increased. The PI3K/Akt pathway and AMPK were activated and the PEPCK expression was reduced. | [30] |
| Beta cell viability and function | ||||
| GSIS in INS-1 cell. [Ca2+] oscillations induced by glucose in INS-1 cells | EGCG, GCG, EC, C, EGC, GC, ECG, CG | 10–100 μM | GSIS was decreased by 10 and 30 μM EGCG. GSIS was terminated by 100 μM EGCG and 100 μM GCG. EGC nearly abolished GSIS at 100 μM, GC and ECG partly inhibited it. EC, C, and CG did not show any effect. 100 μM EGCG decreased the oscillation of intracellular calcium. | [31] |
| GSIS in SFA-treated INS-1 cell; ROS production in high-glucose and H2O2-treated INS-1 cell | EC | 0.3 μmol/L 30 µmol/L | Increase of GSIS. Reversion of SFA-induced inhibition of CaMKII phosphorylation. Reduced ROS production. | [32] |
| Insulin production in iron-loaded RINm5F pancreatic cells. Iron and ROS levels in RINm5F pancreatic cells | GTE | 1–20 µM EGCG 1–10 µM EGCG | Dose-dependent increase of insulin secretion. | [33] |
| Dose-dependent decrease of iron and ROS levels. | ||||
| Cell viability and GSIS in PA- and H2O2-treated INS-1 pancreatic beta cells. H2O2-stimulated ROS production | Cinnam-tannin B1, procyanidin C1, cinnam-tannin D1 | 12.5–100 μmol/L | Dose-dependent increase of cell viability. | [34] |
| GSIS increase at 25 µmol/L. | ||||
| Decreased ROS production. | ||||
| Inhibition of hIAPP aggregation and molecular mechanism | EGCG | - | Blockage of inter-peptide hydrophobic/aromatic interactions and intra-peptide interactions. | [35] |
| Abolishment of β-hairpin-containing three-stranded β-sheet conformation. | ||||
| Shift of hIAPP dimer toward loosely packed coil-rich conformations. | ||||
| Amyloid formation by IAPP and disaggregation of amyloid fibrils with thioflavin-T binding assay and TEM. Cell viability in mixture IAPP:EGCG on rat INS-1 | EGCG | 3.2–32 µM | At 32 µM, inhibition of amyloid formation by IAPP. IAPP:EGCG (3.2 µM) complex did not seed amyloid formation by IAPP. Disaggregation of IAPP. Increased cell viability of INS-1 cells to 77%. | [36] |
| hIAPP fibrillation and aggregation | EGCG | 2–32 µM | Inhibition of hIAPP fibrillation. | [37] |
| Formation of amorphous aggregates instead of ordered fibrils. | ||||
| Beta cell function of rat INS-1E pancreatic beta cells and rat pancreatic islets | 3,4-DHPA, 2,3-DHB and 3-HPP | 1–5 µM | 3,4-DHPA and 3-HPP enhanced GSIS (5 and 1 µM, respectively). Under oxidative stress, 3,4-DHPA and 3-HPP reduced ROS and carbonyl group production, and GSIS returned to control levels. PKC and ERKs phosphorylation improved. | [38] |
| Beta cell function of Min6 pancreatic beta cells incubated with cholesterol | 3,4-DHPA | 10–250 µM | 3,4-DHPA reversed the diminished insulin secretion induced by cholesterol. It protected beta cells against apoptosis, oxidative stress, and mitochondrial dysfunction. | [39] |
| Beta cell function and glucose utilization in rat INS-1 beta cells and human skeletal muscle | EC, HA, HVA and 5-PVA | 5–100 µM | EC (10 and 25 µM), HA, and 5-PVA (25 µM) provoked glucose oxidation in skeletal muscle. After oxidative insult, skeletal mitochondrial function was conserved. In beta cells, EC (100 µM) and metabolites (5–100 µM) stimulated GSIS. | [40] |
| Endogenous glucose production | ||||
| Glucose production and PEPCK/G-6-Pase gene expression in H4IIE rat hepatoma cells incubated with pyruvate and lactate | EGCG | 12.5–100 µM | At 25 µM, glucose production was repressed comparable to that of insulin. | [41] |
| Dose-dependent reduction of PEPCK mRNA as well as G-6-Pase. PI3K inhibitor LY 294,002 reversed the repression of EGCG on PEPCK and G-6-Pase gene expression. NAC and SOD reversed the increased protein-tyrosine phosphorylation and reversed PEPCK and G-6-Pase gene repression. | ||||
| Gluconeogenesis and PEPCK/G-6-Pase gene expression in mouse cAMP-Dex-stimulated hepatocytes | EGCG | 0.25–1 µM | Dose-dependent attenuation of gluconeogenesis. Expression of PEPCK and G-6-Pase genes was blocked. | [42] |
| Activation of AMPK mediated by CaMKK and ROS-dependent. | ||||
| Gluconeogenesis pathway in palmitate-induced insulin resistant HepG2 cells | EGCG | 40 μM | Expression of PEPCK and G-6-Pase was reduced by 53% and 67%, respectively. Glucose production was reduced by 50%. | [43] |
| Incretin effect | ||||
| Plasma membrane potential and GLP-1 secretion in STC-1 cells under basal and nutrient-stimulated conditions | GSPE | 0.05–50 mg/L | At 0.05 and 0.5 mg/L, membrane depolarization. At 50 mg/L, hyperpolarization and suppression of GLP-1 secretion. | [44] |
| Under nutrient-stimulation, 50 mg/L limited membrane depolarization and reduced GLP-1 secretion. | ||||
| Insulin-stimulated glycogen synthesis and lipogenesis in high-glucose treated human hepatoma HepG2 cells | GTP (60% EGCG) | 0.1–10 µM | Enhanced glycogen synthesis, increased phosphorylation of Ser9 GSK3ß and Ser641 GS. | [45] |
| Inhibition of lipogenesis through enhanced expression of phosphorylated AMPKα and acetyl CoA carboxylase. | ||||
| Inflammation | ||||
| TNFα-induced activation of NF-κB, MAPKs, AP-1, and PPARγ in differentiated white 3T3-L1 adipocytes | EC | 0.5–10 µM | Dose-dependent decrease of JNK, ERK1/2, and p-38 phosphorylation, and nuclear AP-1-DNA binding. Inhibition of NF-κB signaling cascade activation, preventing p65 nuclear transport and nuclear NF-κB-DNA binding. Altered transcription of genes (MCP-1, IL-6, TNFα, resistin, PTP1B). Attenuation of decreased PPARγ expression. | [46] |
| Vasodilation | ||||
| Vasodilation of pre-contracted isolated rat aortic rings | 3-PP, 4-HPP, 3,4-DHPP, 4-HPA, 3,4-DHPA, HVA, 3-HB, PhG, 4-MC, m-CoA, 3-HPP and 3-HPA | 100 nM | 3-HPP had the strongest vasodilatory activity, which was NO and endothelium-dependent. | [47] |
| NO production by human aortic endothelial cells under glucotoxic conditions | 3-HPP | 1 µM | Insulin-stimulated increase in NO production was preserved, as well as phosphorylation of Akt and eNOS. The increase in ROS and RNS was prevented. | [48] |
| Endothelial function in human EA.hy926 endothelial cells | 3,4-DHPA, 2,3-DHB and 3-HPP | 10–12 µM | 3,4-DHPA and a mixture of the metabolites increased the NO generation and phosphorylation of eNOS, Akt, and AMPK. Under oxidative stress, cell viability was improved by the metabolites and reduced eNOS phosphorylation was reversed. ROS generation and phosphorylation of ERK and JNK were reversed. | [49] |
| Antiglycative activity | ||||
| AGEs formation in BSA/glucose system and glyoxal trapping ability | PG, 3,4-DHPP, DHFA, 3-HPA, 3,4-DHPA and HVA | 2–50 µmol/L | DHFA at 10 μmol/L significantly inhibited albumin glycation. At 2 µmol/L, a mix of 3-HPA, 3,4-DHPA, and HVA inhibited glycation. PG, 3,4-DHPP, and 3,4-DHPA had a glyoxal trapping ability of 60%, 90%, and 65%, respectively. | [50] |
| AGEs formation in BSA/glucose and BSA/MGO systems | 3,4-DHPA, 3-HPA and HVA | 1 mM | The order of AGEs’ inhibition ability was: rutin > quercetin > 3,4-DHPA > aminoguanidine > 3-HPA > HVA | [51] |
| In Vivo Model | Treatment | Dose/Route/Period | Results | Ref. |
|---|---|---|---|---|
| Glucose absorption in the gut | ||||
| Inhibition of α-amylase activity in mice. | High vs. low DP proanthocyanidins | 150 mg/kg/day. Oral. 56 days. | High DP proanthocyanidins had a stronger inhibition rate of digestive enzyme activity than the low DP group (0.20 ± 0.03 vs. 0.27 ± 0.06 U mg/prot in small intestine, 0.26 ± 0.04 vs. 0.35 ± 0.04 U mg/prot in pancreas) | [52] |
| Insulin signaling pathways and glucose peripheral uptake | ||||
| GLUT4 translocation in ICR mice. GLUT4 translocation in C57BL/6 mice. | CLP (EC, C, procyanidin) | 250 mg/kg. Oral. Single dose. Diet with 0.5% (w/w). Oral. 7 days. | Enhanced GLUT4 translocation in skeletal muscle of ICR mice after a single dose following glucose load. | [53] |
| Enhanced GLUT4 translocation in skeletal muscle of C57BL/6 mice after consecutive administration of CLP. | ||||
| GLUT-4 expression and JNK phosphorylation in KK-Ay mice. | GTP | 150–300 mg/kg/day. Oral. 4 weeks. | Decreased glucose levels and enhanced glucose tolerance. JNK phosphorylation in adipose tissues was reduced and GLUT4 expression was increased. ROS content was reduced. | [26] |
| OGTT and GLUT4 translocation in skeletal muscle of ICR mice. AMPK activation in ICR mice. | CLP and high vs. low DP pro-cyanidins | 10 mg/kg. Oral. Single dose. | Reduction of plasma glucose levels after OGTT. Promotion of GLUT4 translocation by high and low DP procyanidins. Activation of AMPK-signaling pathway. Increased phosphorylation of IRβ, IRS-1, and P13K in muscle. Low-DP increased phosphorylation of Akt. | [54] |
| Increased insulin secretion in plasma. | ||||
| GLUT4 translocation in skeletal muscle of ICR mice. Activation of insulin and AMPK signaling pathways in ICR mice soleus muscle. | EC, Procyanidin B2, Procya-nidin C1, PA4-1 and cinnamtannin A2 | 10 μg/kg. Oral. Single dose. | Reduction of hyperglycemia after an OGTT. Increase of GLUT4 translocation. | [55] |
| Promotion of AMPK, PI3K, and Akt phosphorylation. | ||||
| Glucose uptake in STZ-induced T1D Wistar/ST rats. Glucose uptake in KK-Ay mice. | GTE (C, EC, GC, EGC, CG, ECG, GCG, EGCG and caffeine) | 2 g/L. Oral. 12 d. 2 g/L. Oral. 63 days vs. 42 days. | Significantly lower blood glucose level after OGTT. Increased GLUT4 translocation. Reduction in STZ-induced increase in plasma fructosamine and HbA1c. | [56] |
| Significantly lower blood glucose level after OGTT. Increased GLUT4 translocation. Reduced fructosamine and HbA1c concentration. | ||||
| Glucose intolerance of HFD-induced obese and diabetic C57BL/6 mice. | EGCG | 75 mg/kg. Oral. Single dose. | Blood glucose increased until 15 min (30 min in control), and rapidly decreased thereafter. It was significantly lower than in control group. | [56] |
| Insulin sensitivity in HFD-induced obese and diabetic C57BL/6 mice. | EC | 20 mg/kg. Oral. 15 weeks. | Increase of insulin was prevented. Phosphorylation of IRS-1 and Akt was increased, while that of PKC, JNK, IKK, and PTP1B was downregulated. | [57] |
| IRes and effect on insulin signaling cascade in HFr-fed rats. | EC | 20 mg/kg. Oral. 8 weeks. | Reversion of impaired activation of IR, IRS-1, Akt, and ERK1/2 induced by HFr diet. Reversion of upregulation of PKC, IKK, JNK, and PTP1B induced by HFr. Inhibition of HFr-induced increase of expression and activation of NADPH oxidase, expression of cytokines and chemokines, and activation of redox-sensitive signals. | [58] |
| Plasma glucose level in ICR mice during OGTT and GLUT4 translocation of soleus muscle. | 5-(3,5-dihydroxy-phenyl)-γ-VL | 32 mg/kg. Oral. Single dose. | Suppression of postprandial hyperglycemia at 15 and 30 min after OGTT. Increased GLUT4 translocation. Increased phosphorylation of AMPK. | [27] |
| Beta cell viability and function | ||||
| Glycemia, insulin, and HbA1c glycation on nicotinamide and STZ-induced diabetic rats. | EGCG | 2 mg/kg. Oral. 15 days. | Glucose, HbA1c, and HOMA-IR decreased. Insulin increased. | [59] |
| Insulin synthesis and apoptosis in male Wistar cafeteria-induced obese rats. | GSPE | 25 mg/kg. Oral. 21 days. | Decreased HOMA-IR and HOMA-ß. | [60] |
| Decreased expression of Cpe. | ||||
| Increase in Bax protein levels. | ||||
| IRes, insulin clearance, and serum lipids in NAFLD C57BL/6 mice. | EGCG | 10, 20, 40 mg/kg/day. i.p. 4 weeks. | Reduction of fasting blood glucose and serum insulin. Decrease of GSIS was dose-dependently reversed. Metabolic clearance rate of insulin and IDE increased. Dose-dependent decrease of serum TC, TG, and LDL. Dose-dependent increase of serum HDL. | [61] |
| Antidiabetic effects in a db/db diabetic mouse model. | EGCG | 10 g/kg diet, 1% (w/w). Oral. 10 weeks. | After an OGTT, fasting blood glucose levels decreased similarly to rosiglitazone. No changes in HOMA-IR or QUICKI. Increase in number and size of pancreatic islets comparable to rosiglitazone. | [62] |
| hIAPP amyloidogenesis in hIAPP transgenic mice | EGCG | 0.4 mg/mL. Oral. 3 weeks. | Reduction of amyloid fibril intensity of hIAPP in the pancreas of non-diabetic transgenic mice. | [63] |
| Development of T1D and protective effect on pancreatic islets in NOD mice. | EGCG | 0.05% in drinking water. Oral. 32 weeks. | Delayed onset of T1D. Increased plasma insulin levels compared to control. Decreased HbA1c. Increased concentration of cytokine IL-10 level. Increased islet viability when exposed to pro-inflammatory cytokines. | [64] |
| Immunity modulation and prevention of T1D in NOD mice. | EC | 0.5% in drinking water. Oral. Single dose. | Prevention of T1D onset. Blood glucose levels decreased within the first 60 min of OGTT. HbA1c concentration reduced compared to control group. | [65] |
| Plasma insulin levels were higher than in untreated group. Pancreatic islet mass improved. High proportion of immune cell-free islets. Enhanced plasma IL-10 and IL-12 levels. | ||||
| Incretin effect | ||||
| Plasma GLP-1 in ICR mice. | High vs. low DP pro-cyanidins | 10 mg/kg. Oral. Single dose. | Increased GLP-1 secretion in plasma. | [54] |
| GLP-1 and plasma insulin levels in male ICR mice. | Cinnam-tannin A2 | 10 µg/kg. Oral. Single dose. | Increase of plasma insulin level. Increase of GLP-1 secretion levels in plasma 60 min after administration. | [66] |
| Increased phosphorylation of IRß and IRS-1 in vitro in skeletal muscle. | ||||
| Oxidative stress | ||||
| Oxidative damage and serum lipid profile in STZ-induced diabetic rats. | C | 20–80 mg/kg/day. i.p. 4 weeks. | Dose-dependent decrease of blood glucose levels. Dose-dependent increase of SOD, GST, and CAT activity. Dose-dependent decrease of TC, TG, LDL, and apoB. Dose-dependent increase of HDL and apo A-I. | [67] |
| Serum glucose levels and serum and hepatic biomarkers of oxidative stress in STZ-induced diabetic rats. | GTE | 100 and 200 mg/kg. Oral. 4 weeks. | Decreased serum glucose levels, as well as serum and hepatic MDA concentration with 200 mg/kg for 4 weeks. TAC increased. | [68] |
| Intervention | Study Design | Population | Duration | Parameter | Results | Ref. |
|---|---|---|---|---|---|---|
| Single polyphenols | ||||||
| 500 mg EGCG/day (one capsule/day) Control: 500 mg starch powder/day (one capsule/day) | CT, pc, d-b | n = 326 (women, GDM, 3rd trimester of pregnancy, Chinese, 25–34 years, ≈ 26kg/m2) | Until child’s birth | FBG, INS, HOMA-IR, QUICKI, HOMA-β, BW, BMI, neonatal complications at birth (LBW, hypoglycemia, RD, macrosomia, 1 and 5 min Apgar scores) | ↓ FBG *#, ↓ INS *#, ↓ HOMA-IR *#, ↑ QUICKI *#, ↓ HOMA-β *#, ↓ Neonatal complications at birth | [81] |
| Chocolate | ||||||
| FRC dark (100 g/day in 2 half-bar doses, 1008 mg TP, 36.12 g C). Control: FFWC | RCT, co | n = 19 (women 8, men 11, IGT + hypertension, 44.8 ± 8.0 years, 26.5 ± 1.9 kg/m2) | 15 days (+ 7-day run-in and 7-day washout phase) | FBG, INS, 3-h-PBG, 3-h-PINS, HOMA-IR, β-cell function (CIR120), QUCIKI, ISI, lipids (TC, LDL, HDL, TG), SPB und DBP (clinical + 24-h ABMP), FMD, hsCRP, plasma homo-cysteine, electrolytes, uric acid, fibrinogen | ↓ HOMA-IR *#, ↑ QUICKI *#, ↑ ISI *#, ↑ ISI0 *#, ↑ β-cell function (CIR120) *# → affected 3-h-PBG and 3-h-PINS ↓ SBP *# + DBP *#, ↓ 24-h ABMP *#, ↑ FMD *#, ↓ TC *#, ↓ LDL *# | [82] |
| 27 g/day FRC (2 × 13.5 g, 850 mg flavan-3-ols (90 mg EC) and 100 mg IsoF | RCT, pd, pc | n = 93 (women, postmeno-pausal, T2D, standard therapy; receiving TC-lowering therapy, UK, ≈ 62 years, ≈ 32 kg/m2) | 1 year | FBG, INS, HOMA-IR, HbA1c, QUICKI, lipids (TC, LDL, HDL, TG), 2-h ABPM, BW, 10-y total CHD risk | ↓ INS *#, ↓ HOMA-IR *#, ↑ QUICKI *#, ↓ TC:HDL ratio *#, ↓ LDL *#, ↑ HDL:LDL ratio *#, ↑ CHD risk * but ↓ CHD risk # | [83] |
| GTE | ||||||
| 1500 mg decaffeinated GTE (3 × 500 mg/day, 856 mg EGCG) | RCT, pc, d-b | n = 68 (women 44, men 24, obese T2D, Taiwanese, 51.3 ± 9.2 years, 29.7 ± 4.0 kg/m2) | 16 weeks | FBG, INS, HOMA-IR, HbA1c, leptin, ghrelin, adiponectin, lipids (TG, TC, LDL, HDL), SPB, DBP, creatinine, ALT, uric acid, BW, BMI, WC | ↓ HbA1c *, ↓ HOMA-IR *, ↓ INS *, ↑ ghrelin * (placebo too), ↓ WC * | [84] |
| 340 mL green tea + GTE/day (582.8 mg catechins/day) Control: 340 mL green tea + GTE/day (96.3 mg catechins/day) | CT, pd, d-b | n = 43 (women 25, men 18, T2D, no INS therapy, hypoglycemic drugs (n = 35)) | 12 weeks (+ 4-week run-in, 4-week follow-up period) | FBG, INS, HbA1c, lipids (TG, TC, FFAs), total keton bodies, remnant-like lipoprotein C, adiponectin, enzymes, total protein, albumin, urea nitrogen, uric acid, creatinine, electrolytes, hematology analysis, SBP, DBP, BW, BMI, WC, HC, WHR, FM (%) | At wk 12: ↑ INS *#, ↑ adiponectin *, ↓ TC #, ↓ FFAs *, ↓ total ketone Bodies *, ↓ WC *#, ↓ WHR *# At wk 8: ↓ SBP * | [85] |
| GTE powder (one packet/day, 544 mg PP, 456 mg C) | RCT, co | n = 60 (women 11, men 49; glucose abnormalities, T2D medication (n = 16), 71% IRes, Japanese, ≈ 54 years, ≈ 26 kg/m2) | 2 months/inter-vention | FBG, INS, HOMA-IR, HbA1c, lipids (TC, LDL, HDL, TG), hsCRP, SPB, DBP, BW, BMI, FM | ↓ HbA1c * | [86] |
| GTE powder (one packet/day, 544 mg PP, 456 mg C) | RCT, pd | n = 66 (women 13, men 53; (borderline) T2D, Japanese, ≈ 54 years, ≈ 26 kg/m2) | 2 months | FBG, INS, HOMA-IR, HbA1c, hsCRP, SPB, DBP, BW, BMI | ↓ FBG, ↓ INS, ↓ HOMA-IR, ↓ HbA1c *, ↓ SPB * and DBP *, ↓ BW * and BMI * | [87] |
| 900 mL green tea (9 g) Control: water | CT, co | n = 55 (women 24, men 32, T2D, 53.9 ± 7.7 years, 25.0 ± 2.2 kg/m2) | 4 weeks | FBG, INS, HOMA-IR, adiponectin, lipids (TC, LDL, HDL, TG), hsCRP, IL-6, arterial stiffness | No effects | [88] |
| GSE | ||||||
| 600 mg GSE/day (2 × 300 mg/day) | RCT, co, pc, d-b | n = 32 (women 16, men 16, T2D at high CV risks, oral glucose-lowering therapy (n = 19), UK, 61.8 ± 6.36 years, 30.2 ± 5.92 kg⁄m2) | 4 weeks/intervention (+2-week washout) | FBG, INS, HOMA-IR, HbA1c (only at baseline), fructosamine, lipids (TC, LDL, HDL, TG), liver function, hsCRP, endothelial function, oxidative stress (TAOS, GSH, GSSG), ACR | ↓ TC *, ↓ hsCRP *, ↓ fructosamie *, ↑ GSH * | [89] |
| Cacao | ||||||
| Cacao capsules (2,5 g/day ACTICOATM cacao powder, 207.5 mg Fla) | RCT, pc, pd, d-b | n = 35 (women 17, men 18, T2D + hypertension, dietetic and/or pharmacological treatment, 64.2 ± 1.5 years, ≈ 29 kg/m2 | 12 weeks | FBG, INS, HOMA-IR, HbA1c, lipids (TC, LDL, HDL, TG), SBP, DBD, creatinine, BW, BMI, WC, WHR, FM | ↓ WC * ↓ WHR * | [90] |
| Cacao beverages (2 × 28 g Cacao powder/day, 180, 400 or 900 mg Fla/day) Control: cacao beverages (30 mg Fla/day) | Explora-tory rando-mized study, co | n = 19 (women 10, men 9, obese at risk for IRes, IGT (n = 6), 46 ± 2.3 years, 36.8 ± 1.0 kg/m2) | 5 days (10 days washout) | FBG, INS, TG, hsCRP, ICAM, IL-6, total 8-isoprostane, SBP, DBP, BW, BMI, WC, FM After OGTT: AUC-BG, AUC-INS, 2h-PBG, 2h-PINS, 2h-TG, 2h-hsCRP, 2h-ICAM, 2h-IL-6, total 8-isopros-tane (1h and 1,5h), fibrinogen (1h and 1,5h), HOMA-IR, QUICKI, ISI | ↓ 8-isoprostane *, ↓ hsCRP *, ↓ IL-6 * (as the dose of Fla increased) | [91] |
| Cacao beverage (960 mg PP, 480 mg Fla) with high-fat breakfast (766 kcal, 50 g fat) | RCT, pc, co, d-b (1 week washout phase) | n = 18 (women 14, men 4, T2D, no insulin therapy, 56 ± 3.2 years, 35.3 ± 2.0 kg/m2) | Single dose (6-h study: 0, 1, 2, 3, 4, 5, 6 h) | Fasting + post-prandial: BG, INS, HOMA-IR, lipids (TC, LDL, HDL, TG), hsCRP, SBP, DBP, SAE, LAE; fasting: BW, BMI, WC | ↑ HDL# (1 h and 4 h, 6 h-AUC, overall ∆:1.5 ± 0.8 mg/dL), ↑ Ins# (4 h, overall ∆: 5.2 ± 3.2 mU/L), ↑ HOMA-IR# (4 h, 4 h-AUC, no overall), ↓ LAE# (2 h, overall ∆: −1.6 ± 0.7 mL/mm Hg) | [92] |
| 2.5 g cacao (5 capsules: 0.5 g ACTICOATM cacao powder, 40.4 mg EC) with diabetic-suitable breakfast. Control: cellulose | RCT, pc, co, d-b | n = 12 (women 3, men 9; T2D + overweight/obesity + hypertension, no insulin therapy; 68.0 ± 9.0 years) | Single dose (4-h study: 0, 2, 4h) (≥2 week wash-out) | Fasting & post-prandial: BG, INS, HOMA-IR, lipids (TC, LDL, HDL, TG), SBP, DBP, fasting: BW, WC, HC, WHR, FM | No effects | [93] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Márquez Campos, E.; Jakobs, L.; Simon, M.-C. Antidiabetic Effects of Flavan-3-ols and Their Microbial Metabolites. Nutrients 2020, 12, 1592. https://doi.org/10.3390/nu12061592
Márquez Campos E, Jakobs L, Simon M-C. Antidiabetic Effects of Flavan-3-ols and Their Microbial Metabolites. Nutrients. 2020; 12(6):1592. https://doi.org/10.3390/nu12061592
Chicago/Turabian StyleMárquez Campos, Estefanía, Linda Jakobs, and Marie-Christine Simon. 2020. "Antidiabetic Effects of Flavan-3-ols and Their Microbial Metabolites" Nutrients 12, no. 6: 1592. https://doi.org/10.3390/nu12061592
APA StyleMárquez Campos, E., Jakobs, L., & Simon, M.-C. (2020). Antidiabetic Effects of Flavan-3-ols and Their Microbial Metabolites. Nutrients, 12(6), 1592. https://doi.org/10.3390/nu12061592