Antitumor, Anti-inflammatory and Antiallergic Effects of Agaricus blazei Mushroom Extract and the Related Medicinal Basidiomycetes Mushrooms, Hericium erinaceus and Grifola frondosa: A Review of Preclinical and Clinical Studies

 , ,
, ,
Abstract
1. Introduction
2. Material and Methods
2.1. Antitumor Effects of ABM-Preclinical Studies
2.2. Antitumor Effects of AbM—Clinical Studies
2.3. Antitumor Effects of GF
2.4. Antitumor Effects of HE
3. Anti-inflammatory Effects of AbM and HE
3.1. AbM
3.2. HE
3.3. GF
4. Antiallergic Effects of AbM and GF Extracts
5. Safety of AbM, GF and HE
6. Discussion
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Firenzuoli, F.; Gori, L.; Lombardo, G. The Medicinal Mushroom Agaricus blazei Murrill: Review of Literature and Pharmaco-Toxicological Problems. Evid. Based Complement Altern. Med. 2008, 5, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Fu, Z.; Han, C. The Medicinal Values of Culinary-Medicinal Royal Sun Mushroom (Agaricus blazei Murrill). Evid. Based Complement Altern. Med. 2013, 2013, 842619. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Biedron, R.; Tangen, J.M.; Maresz, K.; Hetland, G. Agaricus blazei Murill immunomodulating properties and health benefits. Funct. Food Health Dis. 2012, 2, 428–447. [Google Scholar] [CrossRef]
- Rop, O.; Mlcek, J.; Jurikova, T. Beta-glucans in higher fungi and their health effects. Nutr. Rev. 2009, 67, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Du, B.; Bian, Z.; Xu, B. Beta-glucans from edible and medicinal mushrooms: Characteristics, physiochemical and biological activities. J. Food Compos. Anal. 2015, 41, 165–173. [Google Scholar] [CrossRef]
- Itoh, H.; Ito, H.; Hibasami, H. Blazein of a New Steroid Isolated from Agaricus blazei Murrill (Himematsutake) Induces Cell Death and Morphological Change Indicative of Apoptotic Chromatin Condensation in Human Lung Cancer LU99 and Stomach Cancer KATO III Cells. Oncol. Rep. 2008, 20, 1359–1361. [Google Scholar]
- Shimizu, T.; Kawai, J.; Ouchi, K.; Kikuchi, H.; Osima, Y.; Hidemi, R. Agarol, an Ergosterol Derivative from Agaricus blazei, Induces Caspase-Independent Apoptosis in Human Cancer Cells. Int. J. Oncol. 2016, 48, 1670–1678. [Google Scholar] [CrossRef]
- Endo, M.; Beppu, H.; Akiyama, H.; Wakamatsu, K.; Ito, S.; Kawamoto, Y.; Shimpo, K.; Sumiya, T.; Koike, T.; Matsui, T. Agaritine Purified from Agaricus blazei Murrill Exerts Anti-Tumor Activity against Leukemic Cells. Biochim. Biophys. Acta 2010, 1800, 669–673. [Google Scholar] [CrossRef]
- Kim, S.P.; Kang, M.Y.; Kim, J.H.; Nam, S.H.; Friedman, M. Composition and mechanism of antitumor effects of Hericium erinaceus mushroom extracts in tumor-bearing mice. J. Agric. Food Chem. 2011, 28, 599861–599869. [Google Scholar] [CrossRef]
- Masuda, Y.; Inoue, H.; Ohta, H.; Miyake, A.; Konishi, M.; Nanba, H. Oral administration of soluble β-glucans extracted from Grifola frondosa induces systemic antitumor immune response and decreases immunosuppression in tumor-bearing mice. Int. J. Cancer 2013, 133, 108–119. [Google Scholar] [CrossRef]
- Friedman, M. Chemistry, Nutrition, and Health-Promoting Properties of Hericium erinaceus (Lion’s Mane) Mushroom Fruiting Bodies and Mycelia and Their Bioactive Compounds. J. Agric. Food Chem. 2015, 63, 7108–7123. [Google Scholar] [CrossRef] [PubMed]
- Klaus, A.; Kozarski, M.; Vunduk, J.; Todorovic, N.; JakovljeviĆ, D.; Zizak, Z.; Pavlović, V.; Levic, S.; Nikšić, M.; Van Griensven, L.J.L.D. Biological potential of extracts of the wild edible Basidiomycete mushroom Grifola frondosa. Food Res. Int. 2015, 67, 272–283. [Google Scholar] [CrossRef]
- Bernardshaw, S.; Johnson, E.; Hetland, G. An extract of the mushroom Agaricus blazei Murill administered orally protects against systemic Streptococcus pneumoniae infection in mice. Scand. J. Immunol. 2005, 62, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Bernardshaw, S.; Hetland, G.; Grinde, B.; Johnson, E. An extract of the mushroom Agaricus blazei Murill protects against lethal septicemia in a mouse model of fecal peritonitis. Shock 2006, 25, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Ellertsen, L.K.; Hetland, G. An extract of the medicinal mushroom Agaricus blazei Murill can protect against allergy. Clin. Mol. Allergy 2009, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Hetland, G.; Eide, D.M.; Tangen, J.M.; Haugen, M.H.; Mirlashari, M.R.; Paulsen, J.E. The Agaricus blazei-Based Mushroom Extract, Andosan™, Protects against Intestinal Tumorigenesis in the A/J Min/+ Mouse. PLoS ONE 2016, 11, e0167754. [Google Scholar] [CrossRef] [PubMed]
- Baecker, A.; Liu, X.; La Vecchia, C.; Zhang, Z.F. Worldwide incidence of hepatocellular carcinoma cases attributable to major risk factors. Eur. J. Cancer Prev. 2018, 27, 205–212. [Google Scholar] [CrossRef]
- McGuigan, A.; Kelly, P.; Turkington, R.C.; Jones, C.; Coleman, H.G.; McCain, R.S. Pancreatic cancer: A review of clinical diagnosis, epidemiology, treatment and outcomes. World J. Gastroenterol. 2018, 24, 4846–4861. [Google Scholar] [CrossRef]
- Watari, J.; Chen, N.; Amenta, P.S.; Fukui, H.; Oshima, T.; Tomita, T.; Miwa, H.; Lim, K.J.; Das, K.M. Helicobacter pylori associated chronic gastritis, clinical syndromes, precancerous lesions, and pathogenesis of gastric cancer development. World J. Gastroenterol. 2014, 20, 5461–5573. [Google Scholar] [CrossRef]
- Morgner, A.; Bayerdörffer, E.; Neubauer, A.; Stolte, M. Gastric MALT lymphoma and its relationship to Helicobacter pylori infection: Management and pathogenesis of the disease. Microsc. Res. Tech. 2000, 48, 349–356. [Google Scholar] [CrossRef]
- Saginala, K.; Barsouk, A.; Aluru, J.S.; Rawla, P.; Padala, S.A.; Barsouk, A. Epidemiology of Bladder Cancer. Med. Sci. 2020, 8, 15. [Google Scholar] [CrossRef]
- Perletti, G.; Monti, E.; Magri, V.; Cai, T.; Cleves, A.; Trinchieri, A.; Montanari, E. The association between prostatitis and prostate cancer. Systematic review and meta-analysis. Arch. Ital. Urol. Androl. 2017, 89, 259–265. [Google Scholar] [CrossRef]
- Canavan, C.; Abrams, K.R.; Mayberry, J. Meta-analysis: Colorectal and small bowel cancer risk in patients with Crohn’s disease. Aliment. Pharmacol. Ther. 2006, 23, 1097–1104. [Google Scholar] [CrossRef]
- Zhou, Q.; Shen, Z.F.; Wu, B.S.; Xu, C.B.; He, Z.Q.; Chen, T.; Shang, H.T.; Xie, C.F.; Huang, S.Y.; Chen, Y.G.; et al. Risk of Colorectal Cancer in Ulcerative Colitis Patients: A Systematic Review and Meta-Analysis. Gastroenterol. Res. Pract. 2019, 2019, 5363261. [Google Scholar] [CrossRef]
- Kodama, N.; Komuta, K.; Sakai, N.; Nanba, H. Effects of D-Fraction, a polysaccharide from Grifola frondosa on tumor growth involve activation of NK cells. Biol. Pharm. Bull. 2002, 25, 1647–1650. [Google Scholar] [CrossRef]
- Lu, X. Impact of IL-12 in Cancer. Curr. Cancer Drug Targets 2017, 17, 682–697. [Google Scholar] [CrossRef]
- Romagnani, S. Induction of TH1 and TH2 responses: A key role for the ‘natural’ immune response? Immunol. Today 1992, 13, 379–381. [Google Scholar] [CrossRef]
- Chou, W.Y.; Lai, P.Y.; Hu, J.M.; Hsu, C.H.; Chen, Y.C.; Tian, Y.F.; You, S.L.; Hsiao, C.W.; Chou, Y.C.; Sun, C.A. Association between atopic dermatitis and colorectal cancer risk: A nationwide cohort study. Medicine 2020, 99, e18530. [Google Scholar] [CrossRef]
- Jiang, X.; Dimou, N.L.; Zhu, Z.; Bonilla, C.; Lewis, S.J.; Lindström, S.; Kraft, P.; Tsilidis, K.K.; Martin, R.M. Allergy, asthma, and the risk of breast and prostate cancer: A Mendelian randomization study. Cancer Causes Control. 2020, 31, 273–282. [Google Scholar] [CrossRef]
- Muir, A.B.; Whelan, K.A.; Dougherty, M.K.; Aaron, B.; Navarre, B.; Aceves, S.S.; Dellon, E.S.; Jensen, E.T. The potential for malignancy from atopic disorders and allergic inflammation: A systematic review and meta-analysis. Clin. Exp. Allergy 2020, 50, 147–159. [Google Scholar] [CrossRef]
- Strachan, D.P. Hay fever, hygiene, and household size. BMJ 1989, 299, 1259–1260. [Google Scholar] [CrossRef] [PubMed]
- Ellertsen, L.K.; Wiker, H.G.; Egeberg, N.T.; Hetland, G. Allergic sensitisation in tuberculosis and leprosy patients. Int. Arch. Allergy Immunol. 2005, 138, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Ellertsen, L.K.; Storla, D.G.; Diep, L.M.; Brokstad, K.A.; Wiker, H.G.; Hetland, G. Allergic sensitisation in tuberculosis patients at the time of diagnosis and following chemotherapy. BMC Infect. Dis. 2009, 9, 100. [Google Scholar] [CrossRef] [PubMed]
- Itoh, H.; Ito, H.; Amano, H.; Noda, H. Inhibitory action of a (1-->6)-beta-D-glucan-protein complex (F III-2-b) isolated from Agaricus blazei Murill (“himematsutake”) on Meth A fibrosarcoma-bearing mice and its antitumor mechanism. Jpn. J. Pharmacol. 1994, 66, 265–271. [Google Scholar] [CrossRef]
- Ito, H.; Shimura, K.; Itoh, H.; Kawade, M. Antitumor effects of a new polysaccharide-protein complex (ATOM) prepared from Agaricus blazei (Iwade strain 101) “Himematsutake” and its mechanisms in tumor-bearing mice. Anticancer Res. 1997, 17, 277–284. [Google Scholar]
- Takaku, T.; Kimura, Y.; Okuda, H. Isolation of an antitumor compound from Agaricus blazei Murill and its mechanism of action. J. Nutr. 2001, 131, 1409–1413. [Google Scholar] [CrossRef]
- Delmanto, R.D.; de Lima, P.L.; Sugui, M.M.; da Eira, A.F.; Salvadori, D.M.; Speit, G.; Ribeiro, L.R. Antimutagenic effect of Agaricus blazei Murrill mushroom on the genotoxicity induced by cyclophosphamide. Mutat. Res. 2001, 496, 15–21. [Google Scholar] [CrossRef]
- Ebina, T.; Fujimiya, Y. Antitumor effect of a peptide-glucan preparation extracted from Agaricus blazei in a double-grafted tumor system in mice. Biotherapy 1998, 11, 259–265. [Google Scholar] [CrossRef]
- Bertéli, M.B.; Lopes, A.D.; Colla, I.M.; Linde, G.A.; Colauto, N.B. Agaricus subrufescens: Substratum nitrogen concentration and mycelial extraction method on antitumor activity. An. Acad. Bras. Ciências 2016, 88, 2239–2246. [Google Scholar] [CrossRef]
- Rubel, R.; Santa, H.S.D.; Dos Santos, L.F.; Fernandes, L.C.; Figueiredo, B.C.; Soccol, C.R. Immunomodulatory and Antitumoral Properties of Ganoderma lucidum and Agaricus brasiliensis (Agaricomycetes) Medicinal Mushrooms. Int. J. Med. Mushrooms 2018, 20, 393–403. [Google Scholar] [CrossRef]
- Yeh, M.Y.; Shang, H.S.; Lu, H.F.; Chou, J.; Yeh, C.; Chang, J.B.; Hung, H.F.; Kuo, W.L.; Wu, L.Y.; Chung, J.G. Chitosan oligosaccharides in combination with Agaricus blazei Murill extract reduces hepatoma formation in mice with severe combined immunodeficiency. Mol. Med. Rep. 2015, 12, 133–140. [Google Scholar] [CrossRef]
- Lin, J.G.; Fan, M.J.; Tang, N.Y.; Yang, J.S.; Hsia, T.C.; Lin, J.J.; Lai, K.C.; Wu, R.S.; Ma, C.Y.; Wood, W.G.; et al. An extract of Agaricus blazei Murill administered orally promotes immune responses in murine leukemia BALB/c mice in vivo. Integr. Cancer Ther. 2012, 11, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.F.; Chen, Y.L.; Lee, M.H.; Shih, Y.L.; Hsu, Y.M.; Tang, M.C.; Lu, H.F.; Tang, N.Y.; Yang, S.T.; Chueh, F.S.; et al. Effect of Agaricus blazei Murrill extract on HT-29 human colon cancer cells in SCID mice in vivo. In Vivo 2011, 25, 673–677. [Google Scholar] [PubMed]
- Wu, M.F.; Lu, H.F.; Hsu, Y.M.; Tang, M.C.; Chen, H.C.; Lee, C.S.; Yang, Y.Y.; Yeh, M.Y.; Chung, H.K.; Huang, Y.P.; et al. Possible reduction of hepatoma formation by Smmu 7721 cells in SCID mice and metastasis formation by B16F10 melanoma cells in C57BL/6 mice by Agaricus blazei murill extract. In Vivo 2011, 25, 399–404. [Google Scholar]
- Pinto, A.V.; Martins, P.R.; Romagnoli, G.G.; Campanelli, A.P.; Terezan, A.P.; Filho, E.R.; Ferreira da Eira, A.; Kaneno, R. Polysaccharide fraction of Agaricus brasiliensis avoids tumor-induced IL-10 production and changes the microenvironment of subcutaneous Ehrlich adenocarcinoma. Cell Immunol. 2009, 256, 27–38. [Google Scholar] [CrossRef]
- Ziliotto, L.; Pinheiro, F.; Barbisan, L.F.; Rodrigues, M.A. Screening for in vitro and in vivo antitumor activities of the mushroom Agaricus blazei. Nutr. Cancer 2009, 61, 245–250. [Google Scholar] [CrossRef]
- Niu, Y.C.; Liu, J.C.; Zhao, X.M.; Wu, X.X. A low molecular weight polysaccharide isolated from Agaricus blazei suppresses tumor growth and angiogenesis in vivo. Oncol. Rep. 2009, 21, 145–152. [Google Scholar]
- Niu, Y.C.; Liu, J.C.; Zhao, X.M.; Cao, J. A low molecular weight polysaccharide isolated from Agaricus blazei Murill (LMPAB) exhibits its anti-metastatic effect by down-regulating metalloproteinase-9 and up-regulating Nm23-H1. Am. J. Chin. Med. 2009, 37, 909–921. [Google Scholar] [CrossRef]
- Yu, C.H.; Kan, S.F.; Shu, C.H.; Lu, T.J.; Sun-Hwang, L.; Wang, P.S. Inhibitory mechanisms of Agaricus blazei Murill on the growth of prostate cancer in vitro and in vivo. J. Nutr. Biochem. 2009, 20, 753–764. [Google Scholar] [CrossRef]
- Murakawa, K.; Fukunaga, K.; Tanouchi, M.; Hosokawa, M.; Hossain, Z.; Takahashi, K. Therapy of Myeloma In Vivo Using Marine Phospholipid in Combination with Agaricus blazei Murill as an Immune Respond Activator. J. Oleo. Sci. 2007, 56, 179–188. [Google Scholar] [CrossRef][Green Version]
- Prabhu, V.V.; Siddikuzzaman Grace, V.M.; Guruvayoorappan, C. Targeting tumor metastasis by regulating Nm23 gene expression. Asian Pac. J. Cancer Prev. 2012, 13, 3539–3548. [Google Scholar] [CrossRef]
- Tangen, J.M.; Holien, T.; Mirlashari, M.R.; Misund, K.; Hetland, G. Cytotoxic Effect on Human Myeloma Cells and Leukemic Cells by the Agaricus blazei Murill Based Mushroom Extract, Andosan™. Biomed. Res. Int. 2017, 2017, 2059825. [Google Scholar] [CrossRef] [PubMed]
- Sovrani, V.; da Rosa, J.; Drewinski, M.P.; Colodi, F.G.; Tominaga, T.T.; Santa, H.S.D.; Rebeca, R. In Vitro and In Vivo Antitumoral Activity of Exobiopolymers from the Royal Sun Culinary-Medicinal Mushroom Agaricus brasiliensis (Agaricomycetes). Int. J. Med. Mushrooms 2017, 19, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Hui, T.; Guo, L.; Wang, J.; Ito, H.; Simura, K. Clinical observation on treatment of acute nonlymphocytic leukemia with Agaricus blazei Murill. J. Lanzhou Med. Coll. 1994, 20, 169–171. [Google Scholar]
- Ahn, W.-S.; Kim, D.-J.; Chae, G.-T.; Lee, J.-M.; Bae, S.-M.; Sin, J.-I.; Kim, Y.-W.; Namkoong, S.-E.; Lee, I.P. Natural killer cell activity and quality of life were improved by consumption of a mushroom extract, Agaricus blazei Murill Kyowa, in gynecological cancer patients undergoing chemotherapy. Int. J. Gynecol. Cancer 2004, 14, 589–594. [Google Scholar] [CrossRef]
- Tangen, J.M.; Tierens, A.; Caers, J.; Binsfeld, M.; Olstad, O.K.; Trøseid, A.M.; Wang, J.; Tjønnfjord, G.E.; Hetland, G. Immunomodulatory effects of the Agaricus blazei Murrill-based mushroom extract AndoSan in patients with multiple myeloma undergoing high dose chemotherapy and autologous stem cell transplantation: A randomized, double blinded clinical study. Biomed. Res. Int. 2015, 2015, 718539. [Google Scholar] [CrossRef]
- Yoshimura, K.; Kamoto, T.; Ogawa, O.; Matsui, S.; Tsuchiya, N.; Tada, H.; Murata, K.; Yoshimura, K.; Habuchi, T.; Fukushima, M. Medical mushrooms used for biochemical failure after radical treatment for prostate cancer: An open-label study. Int. J. Urol. 2010, 17, 548–554. [Google Scholar] [CrossRef]
- Grinde, B.; Hetland, G.; Johnson, E. Effects on gene expression and viral load of a medicinal extract from Agaricus blazei in patients with chronic hepatitis C infection. Int. Immunopharmacol. 2006, 6, 1311–1314. [Google Scholar] [CrossRef]
- Suzuki, I.; Itani, T.; Ohno, N.; Oikawa, S.; Sato, K.; Miyamzaki, T.; Yadomae, T. Effect of a polysaccharide fraction from Grifola frondosa on immune response in mice. J. Pharm. Dyn. 1985, 8, 217–226. [Google Scholar] [CrossRef]
- Takeyama, T.; Suzuki, I.; Ohno, N.; Oikawa, S.; Sato, K.; Ohsawa, M.; Yadomae, T. Host-mediated antitumor effect of grifolan NMF-5N, a polysaccharide obtained from Grifola frondosa. J. Pharmacobiodyn. 1987, 10, 644–651. [Google Scholar] [CrossRef]
- Kodama, N.; Komuta, K.; Nanba, H. Effect of Maitake (Grifola frondosa) D-Fraction on the activation of NK cells in cancer patients. J. Med. Food 2003, 6, 371–377. [Google Scholar] [CrossRef]
- Masuda, Y.; Matsumoto, A.; Toida, T.; Oikawa, T.; Ito, K.; Nanba, H. Characterization and antitumor effect of a novel polysaccharide from Grifola frondosa. J. Agric. Food Chem. 2009, 57, 10143–10149. [Google Scholar] [CrossRef]
- Masuda, Y.; Nakayama, Y.; Tanaka, A.; Naito, K.; Konishi, M. Antitumor activity of orally administered maitake α-glucan by stimulating antitumor immune response in murine tumor. PLoS ONE 2017, 12, e0173621. [Google Scholar] [CrossRef]
- Masuda, Y.; Inoue, M.; Miyata, A.; Mizuno, S.; Nanba, H. Maitake beta-glucan enhances therapeutic effect and reduces myelosupression and nephrotoxicity of cisplatin in mice. Int. Immunopharmacol. 2009, 9, 620–626. [Google Scholar] [CrossRef]
- Mao, G.H.; Zhang, Z.H.; Fei, F.; Ding, Y.Y.; Zhang, W.J.; Chen, H.; Ali, S.S.; Zhao, T.; Feng, W.W.; Wu, X.Y.; et al. Effect of Grifola frondosa polysaccharide on anti-tumor activity in combination with 5-Fu in Heps-bearing mice. Int. J. Biol. Macromol. 2019, 121, 930–935. [Google Scholar] [CrossRef]
- Mao, G.; Li, Q.; Deng, C.; Wang, Y.; Ding, Y.; Zhang, W.; Chen, Y.; Zhao, T.; Wei, F.; Yang, L.; et al. The synergism and attenuation effect of Selenium (Se)-enriched Grifola frondosa (Se)-polysaccharide on 5-Fluorouracil (5-Fu) in Heps-bearing mice. Int. J. Biol. Macromol. 2018, 107 Pt B, 2211–2216. [Google Scholar] [CrossRef]
- Vetchinkina, E.; Shirokov, A.; Bucharskaya, A.; Navolokin, N.; Prilepskii, A.; Burov, A.; Maslyakova, G.; Nikitina, V.E. Antitumor Activity of Extracts from Medicinal Basidiomycetes Mushrooms. Int. J. Med. Mushrooms 2016, 18, 955–964. [Google Scholar] [CrossRef]
- Kodama, N.; Mizuno, S.; Nanba, H.; Saito, N. Potential antitumor activity of a low-molecular-weight protein fraction from Grifola frondosa through enhancement of cytokine production. J. Med. Food 2010, 13, 20–30. [Google Scholar] [CrossRef]
- Kim, S.P.; Kang, M.Y.; Choi, Y.H.; Kim, J.H.; Nam, S.H.; Friedman, M. Mechanism of Hericium erinaceus (Yamabushitake) mushroom-induced apoptosis of U937 human monocytic leukemia cells. Food Funct. 2011, 2, 348–356. [Google Scholar] [CrossRef]
- Kim, S.P.; Nam, S.H.; Friedman, M. Hericium erinaceus (Lion’s Mane) mushroom extracts inhibit metastasis of cancer cells to the lung in CT-26 colon cancer-transplanted mice. J. Agric. Food Chem. 2013, 61, 4898–4904. [Google Scholar] [CrossRef]
- Li, G.; Yu, K.; Li, F.; Xu, K.; Li, J.; He, S.; Cao, S.; Tan, G.-S. Anticancer potential of Hericium erinaceus extracts against human gastrointestinal cancers. J. Ethnopharmacol. 2014, 153, 521–530. [Google Scholar] [CrossRef]
- Nakamura, A.; Zhu, Q.; Yokoyama, Y.; Kitamura, N.; Uchida, S.; Kumadaki, K.; Tsubota, K.; Watanabe, M. Agaricus brasiliensis KA21 May Prevent Diet-Induced Nash Through Its Antioxidant, Anti-Inflammatory, and Anti-Fibrotic Activities in the Liver. Foods 2019, 8, 546. [Google Scholar] [CrossRef]
- Val, C.H.; Brant, F.; Miranda, A.S.; Rodrigues, F.G.; Oliveira, B.C.L.; Dos Santos, E.A.; De Assis, D.R.R.; Esper, L.; Silva, B.C.; Rachid, M.A.; et al. Effect of mushroom Agaricus blazei on immune response and development of experimental cerebral malaria. Malar. J. 2015, 14, 311. [Google Scholar] [CrossRef]
- Croccia, C.; Lopes, A.J.; Pinto, L.F.R.; Sabaa-Srur, A.U.O.; Vaz, L.C.A.; Trotte, M.N.D.S.; Tessarollo, B.; Silva, A.C.; De Matos, H.J.; Nunes, R.A. Royal Sun Medicinal Mushroom Agaricus Brasiliensis (Higher Basidiomycetes) and the Attenuation of Pulmonary Inflammation Induced by 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK). Int. J. Med. Mushrooms 2013, 15, 345–355. [Google Scholar] [CrossRef]
- Li, I.C.; Lee, L.Y.; Chen, Y.J.; Chou, M.Y.; Wang, M.F.; Chen, W.P.; Chen, Y.P.; Chen, C.C. Erinacine A-enriched Hericium erinaceus mycelia promotes longevity in Drosophila melanogaster and aged mice. PLoS ONE 2019, 14, e0217226. [Google Scholar] [CrossRef]
- Ren, Y.; Geng, Y.; Du, Y.; Wang, L.; Lu, Z.-M.; Xu, H.-Y.; Xu, G.-H.; Shi, J.-S.; Xu, Z.-H. Polysaccharide of Hericium Erinaceus Attenuates Colitis in C57BL/6 Mice via Regulation of Oxidative Stress, Inflammation-Related Signaling Pathways and Modulating the Composition of the Gut Microbiota. J. Nutr. Biochem. 2018, 57, 67–76. [Google Scholar] [CrossRef]
- Diling, C.; Xin, Y.; Chaoqun, Z.; Jian, Y.; Xiaocui, T.; Jun, C.; Ou, S.; Yizhen, X. Extracts from Hericium erinaceus relieve inflammatory bowel disease by regulating immunity and gut microbiota. Oncotarget 2017, 8, 85838–85857. [Google Scholar] [CrossRef]
- Lee, K.-F.; Chen, J.-H.; Teng, C.-C.; Shen, C.-H.; Hsieh, M.-C.; Lu, C.-C.; Lee, K.-C.; Lee, L.-Y.; Chen, W.-P.; Chen, C.-C.; et al. Protective effects of Hericium erinaceus mycelium and its isolated erinacine A against ischemia-injury-induced neuronal cell death via the inhibition of iNOS/p38 MAPK and nitrotyrosine. Int. J. Mol. Sci. 2014, 15, 15073–15089. [Google Scholar] [CrossRef]
- Li, X.; Zeng, F.; Huang, Y.; Liu, B. The Positive Effects of Grifola frondosa Heteropolysaccharide on NAFLD and Regulation of the Gut Microbiota. Int. J. Mol. Sci. 2019, 20, 5302. [Google Scholar] [CrossRef]
- Han, C.; Cui, B. Pharmacological and pharmacokinetic studies with agaricoglycerides, extracted from Grifola frondosa, in animal models of pain and inflammation. Inflammation 2012, 35, 1269–1275. [Google Scholar] [CrossRef]
- Lee, J.S.; Park, S.-Y.; Thapa, D.; Choi, M.K.; Chung, I.-M.; Park, Y.-J.; Yong, C.S.; Choi, H.G.; Kim, J.-A. Grifola frondosa water extract alleviates intestinal inflammation by suppressing TNF-alpha production and its signaling. Exp. Mol. Med. 2010, 42, 143–154. [Google Scholar] [CrossRef]
- Therkelsen, S.P.; Hetland, G.; Lyberg, T.; Lygren, I.; Johnson, E. Cytokine Levels after Consumption of a Medicinal Agaricus blazei Murill-Based Mushroom Extract, AndoSan™, in Patients with Crohn’s Disease and Ulcerative Colitis in a Randomized Single-Blinded Placebo-Controlled Study. Scand. J. Immunol. 2016, 84, 323–331. [Google Scholar] [CrossRef]
- Therkelsen, S.P.; Hetland, G.; Lyberg, T.; Lygren, I.; Johnson, E. Effect of a Medicinal Agaricus blazei Murill-Based Mushroom Extract, AndoSan™, on Symptoms, Fatigue and Quality of Life in Patients with Ulcerative Colitis in a Randomized Single-Blinded Placebo Controlled Study. PLoS ONE 2016, 11, e0150191. [Google Scholar] [CrossRef]
- Therkelsen, S.P.; Hetland, G.; Lyberg, T.; Lygren, I.; Johnson, E. Effect of the Medicinal Agaricus blazei Murill-Based Mushroom Extract, AndoSanTM, on Symptoms, Fatigue and Quality of Life in Patients with Crohn’s Disease in a Randomized Single-Blinded Placebo Controlled Study. PLoS ONE 2016, 11, e0159288. [Google Scholar] [CrossRef]
- Førland, D.T.; Johnson, E.; Saetre, L.; Lyberg, T.; Lygren, I.; Hetland, G. Effect of an extract based on the medicinal mushroom Agaricus blazei Murill on expression of cytokines and calprotectin in patients with ulcerative colitis and Crohn’s disease. Scand. J. Immunol. 2011, 73, 66–75. [Google Scholar] [CrossRef]
- Johnson, E.; Førland, D.T.; Hetland, G.; Sætre, L.; Olstad, O.K.; Lyberg, T. Effect of AndoSan™ on expression of adhesion molecules and production of reactive oxygen species in human monocytes and granulocytes in vivo. Scand. J. Gastroenterol. 2012, 47, 984–992. [Google Scholar] [CrossRef]
- Johnson, E.; Førland, D.T.; Saetre, L.; Bernardshaw, S.V.; Lyberg, T.; Hetland, G. Effect of an extract based on the medicinal mushroom Agaricus blazei murill on release of cytokines, chemokines and leukocyte growth factors in human blood ex vivo and in vivo. Scand. J. Immunol. 2009, 69, 242–250. [Google Scholar] [CrossRef]
- Bernardshaw, S.; Hetland, G.; Ellertsen, L.K.; Tryggestad, A.M.; Johnson, E. An extract of the medicinal mushroom Agaricus blazei Murill differentially stimulates production of pro-inflammatory cytokines in human monocytes and human vein endothelial cells in vitro. Inflammation 2005, 29, 147–153. [Google Scholar] [CrossRef]
- Oh, T.W.; Kim, Y.A.; Jang, W.J.; Byeon, J.I.; Ryu, C.H.; Kim, J.O.; Ha, Y.L. Semipurified fractions from the submerged-culture broth of Agaricus blazei Murill reduce blood glucose levels in streptozotocin-induced diabetic rats. J. Agric. Food Chem. 2010, 58, 4113–4119. [Google Scholar] [CrossRef]
- Padilha, M.M.; Avila, A.A.; Sousa, P.J.; Cardoso, L.G.; Perazzo, F.F.; Carvalho, J.C. Anti-inflammatory activity of aqueous and alkaline extracts from mushrooms (Agaricus blazei Murill). J. Med. Food 2009, 12, 359–364. [Google Scholar] [CrossRef]
- Izawa, S.; Inoue, Y. A screening system for antioxidants using thioredoxin-deficient yeast: Discovery of thermostable antioxidant activity from Agaricus blazei Murill. Appl. Microbiol. Biotechnol. 2004, 64, 537–542. [Google Scholar] [CrossRef]
- Choi, Y.H.; Yan, G.H.; Chai, O.H.; Choi, Y.H.; Zhang, X.; Lim, J.M.; Kim, J.H.; Lee, M.S.; Han, E.H.; Kim, H.T.; et al. Inhibitory effects of Agaricus blazei on mast cell-mediated anaphylaxis-like reactions. Biol. Pharm. Bull. 2006, 29, 1366–1371. [Google Scholar] [CrossRef]
- Takimoto, H.; Kato, H.; Kaneko, M.; Kumazawa, Y. Amelioration of skewed Th1/Th2 balance in tumor-bearing and asthma-induced mice by oral administration of Agaricus blazei extracts. Immunopharmacol. Immunotoxicol. 2008, 30, 747–760. [Google Scholar] [CrossRef]
- Bouike, G.; Nishitani, Y.; Shiomi, H.; Yoshida, M.; Azuma, T.; Hashimoto, T.; Kanazawa, K.; Mizuno, M. Oral Treatment with Extract of Agaricus blazei Murill Enhanced Th1 Response through Intestinal Epithelial Cells and Suppressed OVA-Sensitized Allergy in Mice. Evid. Based Complement Altern. Med. 2011, 2011, 1–11. [Google Scholar] [CrossRef]
- Mahmood, F.; Hetland, G.; Nentwich, I.; Mirlashari, M.R.; Ghiasvand, R.; Nissen-Meyer, L.S.H. Agaricus blazei-Based Mushroom Extract Supplementation to Birch Allergic Blood Donors: A Randomized Clinical Trial. Nutrients 2019, 11, 2339. [Google Scholar] [CrossRef]
- Kawai, J.; Mori, K.; Hirasawa, N. Grifola frondosa extract and ergosterol reduce allergic reactions in an allergy mouse model by suppressing the degranulation of mast cells. Biosci. Biotechnol. Biochem. 2019, 83, 2280–2287. [Google Scholar] [CrossRef]
- Park, H.S.; Hwang, Y.H.; Kim, M.K.; Hong, G.E.; Lee, H.J.; Nagappan, A.; Yumnam, S.; Kim, E.H.; Heo, J.D.; Lee, S.J.; et al. Functional Polysaccharides from Grifola Frondosa Aqueous Extract Inhibit Atopic Dermatitis-Like Skin Lesions in NC/Nga Mice. Biosci. Biotechnol. Biochem. 2015, 79, 147–154. [Google Scholar] [CrossRef]
- Mukai, H.; Watanabe, T.; Ando, M.; Katsumata, N. An alternative medicine, Agaricus blazei, may have induced severe hepatic dysfunction in cancer patients. Jpn. J. Clin. Oncol. 2006, 36, 808–810. [Google Scholar] [CrossRef]
- Suehiro, M.; Katoh, N.; Kishimoto, S. Cheilitis due to Agaricus blazei Murill mushroom extract. Contact Dermat. 2007, 56, 293–294. [Google Scholar] [CrossRef]
- Engdal, S.; Nilsen, O.G. In vitro inhibition of CYP3A4 by herbal remedies frequently used by cancer patients. Phytother. Res. 2009, 23, 906–912. [Google Scholar] [CrossRef]
- Shapiro, L.E.; Shear, N.H. Drug interactions: Proteins, pumps, and P-450s. J. Am. Acad. Dermatol. 2002, 47, 467–484. [Google Scholar] [CrossRef] [PubMed]
- Sumiya, T.; Ikeda, Y.; Broadmeadow, A.; May, K.; Pritchard, L.; Horne, C.; Burlinson, B. Himematsutake (Iwade Strain 101) extract (ABM-FD): Genetic toxicology and a 3-month dietary toxicity study in rats. Food Chem. Toxicol. 2008, 46, 1949–1959. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.P.; Kang, B.H.; Roh, J.K.; Kim, J.R. Lack of carcinogenicity of lyophilized Agaricus blazei Murill in a F344 rat two year bioassay. Food Chem. Toxicol. 2008, 46, 87–95. [Google Scholar] [CrossRef]
- Tanaka, H.; Tsunematsu, K.; Nakamura, N.; Suzuki, K.; Tanaka, N.; Takeya, I.; Saikai, T.; Abe, S. Successful treatment of hypersensitivity pneumonitis caused by Grifola frondosa (Maitake) mushroom using a HFA-BDP extra-fine aerosol. Intern. Med. 2004, 43, 737–740. [Google Scholar] [CrossRef][Green Version]
- Deng, G.; Lin, H.; Seidman, A.; Fornier, M.; D’Andrea, G.; Wesa, K.; Yeung, K.; Cunningham-Rundles, S.; Vickers, A.; Cassileth, B. A phase I/II trial of a polysaccharide extract from Grifola frondosa (Maitake mushroom) in breast cancer patients: Immunological effects. J. Cancer Res. Clin. Oncol. 2009, 135, 1215–1221. [Google Scholar] [CrossRef]
- Chan, G.C.; Chan, W.K.; Sze, D.M. The effects of beta-glucan on human immune and cancer cells. J. Hematol. Oncol. 2009, 2, 25. [Google Scholar] [CrossRef]
- Lehne, G.; Haneberg, B.; Gaustad, P.; Johansen, P.W.; Preus, H.; Abrahamsen, T.G. Oral administration of a new soluble branched beta-1,3-D-glucan is well tolerated and can lead to increased salivary concentrations of immunoglobulin A in healthy volunteers. Clin. Exp. Immunol. 2006, 143, 65–69. [Google Scholar] [CrossRef]
- Samuelsen, A.B.; Schrezenmeir, J.; Knutsen, S.H. Effects of orally administered yeast-derived beta-glucans: A review. Mol. Nutr. Food Res. 2014, 58, 183–193. [Google Scholar] [CrossRef]
- Berven, L.; Karppinen, P.; Hetland, G.; Samuelsen, A.B. The polar high molecular weight fraction of the Agaricus blazei Murill extract, AndoSan™, reduces the activity of the tumor-associated protease, legumain, in RAW 264.7 cells. J. Med. Food 2015, 18, 429–438. [Google Scholar] [CrossRef]
- Hetland, G.; Johnson, E.; Lyberg, T.; Bernardshaw, S.; Tryggestad, A.M.; Grinde, B. Effects of the medicinal mushroom Agaricus blazei Murill on immunity, infection and cancer. Scand. J. Immunol. 2008, 68, 363–370. [Google Scholar] [CrossRef]
- Batbayar, S.; Lee, D.H.; Kim, H.W. Immunomodulation of Fungal β-Glucan in Host Defense Signaling by Dectin-1. Biomol. Ther. 2012, 20, 433–445. [Google Scholar] [CrossRef] [PubMed]


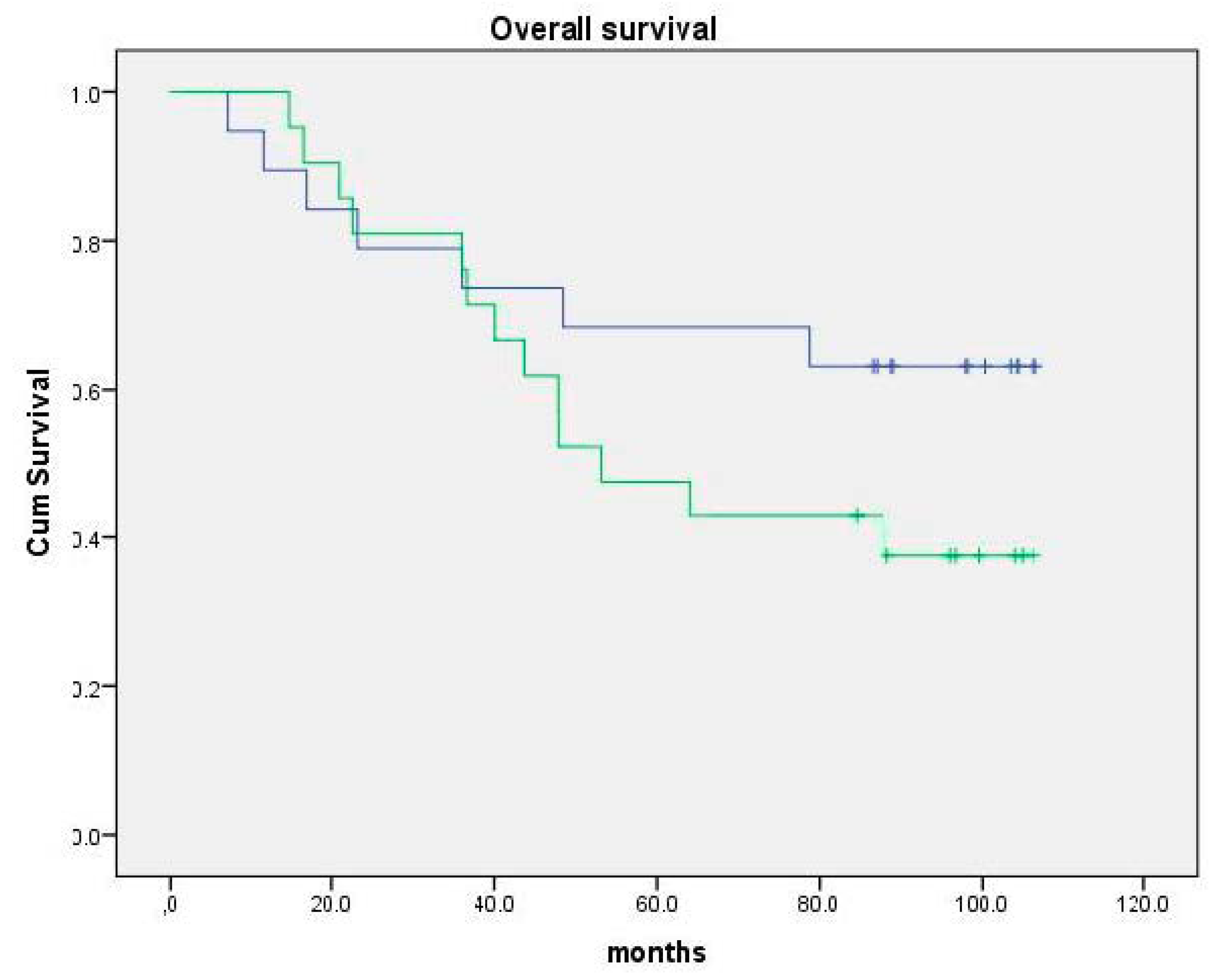{kind=link}
| Product Admin. | Study In, Of | Effects | Mechanism | Author, Year [Ref] |
|---|---|---|---|---|
| Solid-state fermented mycelia, p.o. | Mice, Sarcoma | Growth inhibition | Immunomodulation ↑ T cell and ↓ B cell | Rubel et al., 2018 [40] |
| Ergosterol deriv. (Agarol), i.p. | SCID Mice, Lung Adenoarcinoma | Inhibition | Apoptosis | Shimizu et al., 2016 [7] |
| Water mycelia extract incl. HE, GF, p.o. (Andosan) | Min Mice, spon. Adenocarcinoma | Inhibition | Immunomodulation, Apoptosis | Hetland et al., 2016 [16] |
| Water mycelia extract, p.o. | Mice, Sarcoma | Antitumor | Immunoprotection | Bertéli et al., 2016 [39] |
| Water extract + Chitosan, p.o. | SCID Mice, Hepatoma | Inhibition | Angiogenetic effect | Yeh et al., 2015 [41] |
| Water extract, p.o. | Mice, Murine Leukemia | ↓ Liver & spleen size ↑ IL-1β,IL-6, IFNγ, ↓ IL-4 | ↑ T% B cells, ↓ MΦ Immunomodulation | Lin et al., 2012 [42] |
| Water extract, p.o. | SCID Mice, colon cancer, Hepatoma Melanoma | Inhibition, Inhibition, ↑ Life span | Dose-dependent Immunomodulation | Wu et al., 2011 [43,44] |
| β-glucan-rich extract, i.t. | Mice, Ehrlich, tumor | Inhibition, ↑ IFNγ, T cc, MΦ, ↓ IL-10 | Immune cell tumor migration Cytokine switch, Apoptosis | Pinto et al., 2009 [45] |
| Crude fruiting body, p.o. | Rats, colon ca. | No effect on colon carcinogenesis | - | Ziliotto et al., 2009 [46] |
| LMW polysacc., i.t., i.p. | Mice, Sarcoma Melanoma | Inhibition ↓ Lung metastasis | Inhib. Angiogenesis Metalloproteinase modulation | Niu et al., 2009 [47,48] |
| Broth fraction, p.o. | SCID mice, Prostate ca. | Inhibition | Apoptosis, Antiangiogenesis | Yu et al., 2009 [49] |
| Water extract + marine phospholipid, p.o. | Nude Mice, Myeloma | Inhibition | Immunomodulation ↑ uptake of extract | Murakawa et al., 2007 [50] |
| Ergosterol, p.o. | Mice, Sarcoma | Inhibition | ↓ Neo-vascularization | Takaku et al., 2001 [36] |
| Water extract, p.o. | Mice, CP-induced clastogenicity | Anticarcinogenic | Antimutagenic | Delmanto et al., 2001 [37] |
| Mycelia polysaccprotein complex, i.p., p.o. | Mice, Sarcoma | Inhibition | Immunological | Ito et al., 1997 [35] |
| Product Admin. | Study In, Of | Effects | Mechanism | Author, Year [Ref] |
|---|---|---|---|---|
| Water mycelia extract incl. HE, GF, p.o. (Andosan) | Placebo-ctr RCT Myeloma pat. (n = 40) | Immunomodula-ting | Immunomodulation | Tangen et al., 2015 [56] |
| Water extract, p.o. (Senseiro) | Prostate ca. patients (n = 32) | Longer PSA doubling time, no testosterone correlation | - | Yoshimura et al., 2010 [57] |
| Water mycelia extract incl. HE, GF, p.o. (Andosan) | Patient with chron. HCV infection (n = 5) | ↑ Expression of “antitumor” genes | Immunomodulation | Grinde et al., 2006 [58] |
| Water extract, p.o. | Gynecol. ca. patients (n = 100) | ↑ QoL, ↑ NK cc activ. | Immunomodulation | Ahn et al., 2004 [55] |
| Water extract, p.o. | Ac. non-lymphoblastic leukemic patients (n = 10) | Inhibition | - | Hui et al., 1994 [54] |
| Product, Admin. | Study In, Of | Effects | Mechanism | Author, Year [Ref] |
|---|---|---|---|---|
| GF β-glucan, p.o. | Mice, Heps tumor (hepatoma) | ↑ Inhibitory effect of 5-Fu against Heps-tumor | Synergism and immune regulation | Mao et al., 2019 [65] |
| Selenium-enriched GF polysaccharide, p.o. | Mice, Heps tumor (hepatoma) | Inhibition of Heps-tumor | Improved immune function | Mao et al., 2018 [66] |
| GF β-glucan, p.o. | Mice, Colon tumor, Melanoma | Inhibition, Systemic antitumor response ↑ Survival | MΦ activ. in Peyer’s patches and ↑ IFNγ | Masuda et al., 2017 [63] |
| GF mycelia extract, p.o. | Rats, Kidney ca. | Inhibition | Immunomodulation and tumor necrosis | Vetchinkina et al., 2016 [67] |
| GF LMW protein fraction, i.p. | Mice, Colon ca. | Inhibition | ↑ IL-1, TNFα, IL-10, IL-12, IFNγ, activ. NK and DC cc, MΦ | Kodama et al., 2010 [68] |
| GF polysacc., i.p. | Mice, Colon ca. | Inhibition | Induced cell mediated immunity ↑ Th1 cytokines | Masuda et al., 2009 [62] |
| GF polysacc., i.p. | Mice, Colon ca. | ↑ Antitumor and-metastatic effect on cisplatin, ↓myelo- and nephrotoxicity | Synergistic effect of cisplatin cytotoxicity and GF immunomodulation | Masuda et al., 2009 [64] |
| GF D fraction, p.o. | Diff. Cancer pat. (n = 10) | Hindered metastat. prog. ↓expression of tumor markers | Immunomodulation ↑NK cell activ. and ↑Th 1/↓Th2 | Kodama et al., 2003 [61] |
| GF β-glucan, i.p. | Mice, Colon ca. | Inhibition | Induced cellular mediated immunity and Th1 dominance | Kodama et al., 2002 [25] |
| GF β-glucan, i.p. | Mice, Sarcoma, Carcinoma | Inhibition | Host-mediated mechanism involving MΦ and cytotoxic T cc | Takeyama et al., 1987 [60] |
| GF polysacc. fraction, i.p., i.t. | Mice, Sarcoma | Repression | ↑ Weight of spleen cc and number | Suzuki et al., 1985 [59] |
| Product, Admin. | Study In, Of | Effects | Mechanism | Author, Year [Ref] |
|---|---|---|---|---|
| AbM dry feed, p.o. | Mice, non-alcoholic steato-hepatitis | Prevention | Prevention of oxidative stress | Nakamura et al., 2019 [72] |
| AbM water extract fractions | Mice, cerebral malaria | Improved consequence of cerebral malaria | ↓ TNFα, IL-6, IL-1β Antimalarial activity | Val et al., 2015 [73] |
| AbM extract, p.o. | Rats, Pulmonary inflammation | ↓ Lung damage induced by carcinogen | Attenuation of pulmonary inflammation & gross consolidation | Croccia et al., 2013 [74] |
| Erinacine A-enriched HE mycelia, p.o. | Mice, Life-prolonging activity | Increased longevity in aged mice | Induction of endogenous antioxidant enzymes | Li et al., 2019 [75] |
| HE Polysaccharide, p.o. | Mice, Colitis | Attenuation of colitis, reversing of gut dysbiosis | Downregulation of oxidative stress and inflamm.-related signaling pathways, Maintaining intestinal barrier | Ren et al., 2018 [76] |
| HE alcohol extract & polysacc., p.o. | Rats, IBD | Improved damages in colonic mucosa of induced IBD | ↓ MPO activ., NFKB, TNFα, ↑T cc activ. Beneficial gut bacteria growth and improved host immunity | Diling et al., 2017 [77] |
| HE mycelium alcohol extract & erinacine A, p.o. | Rats, brain ischemia | Protection against brain ischemia injury induced neuronal cell death | Inhibition of iNOS/P3 MAPK, ↓ IL-1β, IL-6, TNFα, ↑ nerve growth properties | Lee et al., 2014 [78] |
| GF polysacc., p.o. | Rats, non-alcoholic steato-hepatitis | Protection | Beneficial regulation of microbiota | Li et al., 2019 [79] |
| Fermented GF extract, p.o. | Rats ET-induced uveitis | Anti-inflammatory | ↓ IL-1β, TNFα, NFΚB activ., iNOS express. | Han et al.., 2012 [80] |
| GF water extract, p.o. | Rats, IBD | ↓ Colon ulceration | Amelioration by ↓MPO, TNFα colon express. and NFΚB signaling | Lee et al., 2010 [81] |
| Product, Admin. | Study In, Of | Effects | Mechanism | Author, Year [Ref] |
|---|---|---|---|---|
| AbM mycelia water extract incl. HE, GF, p.o. (Andosan) | Placebo-ctr RCT, IBD patients; 50 UC, 50 CD | Improved symptoms & QoL espec. in UC | ↓ Proinflammatory effect | Therkelsen et al., 2016 [82,83,84] |
| AbM mycelia water extract incl. HE, GF, p.o. (Andosan) | Pilot study, IBD patients; 10 UC, 11 CD | Anti-inflammatory | ↓ Proinflammatory cytokines, ↓ fecal calprotectin | Førland et al., 2011 [85] |
| AbM mycelia water extract incl. HE, GF, p.o. (Andosan) | Healthy Volunteers (n = 8) | Antioxidant effect | ↓iROS prod. and Adhesion molec. express. in MΦ and granuloc. | Johnson et al., 2012 [86] |
| AbM mycelia water extract incl. HE, GF, p.o. (Andosan) | Healthy Volunteers (n = 10) | Predominantly anti-inflammatory effect | ↓Proinflammatory cytokines | Johnson et al., 2009 [87] |
| Product, Admin. | Study In, Of | Effects | Mechanism | Author, Year, [Ref.] |
|---|---|---|---|---|
| AbM mycelia water extract incl. HE, GF, p.o. (Andosan) | Placebo-ctr RCT in blood donors, Pollen allergy & asthma (n = 60) | ↓ General symptoms, and medication | ↓ Spec. IgE, reduced basophil sensitivity | Mahmood et al., 2019 [95] |
| Water AbM extract, p.o. | Mice, Allergy | ↓ OVA sensitization | ↓ Spec. IgE, improv. Th1/Th2 balance via MΦ activ. by epithelial cc, diff. promotion of naïve T cc to Th1 cc | Bouike et al., 2011 [94] |
| AbM mycelia water extract incl. HE, GF, p.o. (Andosan) | Mice, Allergy | ↓ OVA sensitization | ↓ Spec. IgE, IgG1 and improved Th1/Th2 balance | Ellertsen & Hetland 2009 [15] |
| AbM water extract, p.o. | Mice, Asthma | ↓ Spec. IgE, IgG1 and bronchial eosinophils | Amelioration of skewed Th1/Th2 balance | Takimoto et al., 2008 [93] |
| AbM water extract, p.o. | Mice, Anaphylaxis | Inhib. of induced anaphylactic reaction and ear swelling | Treatment of mast cell mediated anaphylactic reaction | Choi et al., 2006 [92] |
| GF alcohol extract and ergosterol, p.o. | Mice, Allergic inflammation | Inhib. mast cc degranulation, alleviated anaphylactic cutaneous response | ↓ Type 1 allergic reaction by suppression of mast cc degranulation | Kawai et al., 2019 [96] |
| GF polysacch, p.o. | Mice, AD | Inhib. AD-like skin lesion | ↓ IgE, mast cc infiltr., cytokine express. controlling Th1/Th2 | Park et al., 2015 [97] |
| Product Admin. | Study In, Of | Effects | Mechanism | Author, Year [Ref] |
|---|---|---|---|---|
| AbM powder, p.o. | Rats, Toxicity and oncogenicity | ↓ Mortality in 2-yrs toxicity study | Possible anti-mutagenic and antioxidant effect, No carcinogenicity | Lee et al., 2008 [103] |
| AbM water extract, p.o. | Rats, Subchronic toxicity | Low subchronic toxicity at very high doses | Neg. genotoxicity test, possible clastogenic activity but no direct effect on DNA | Sumiya et al., 2008 [102] |
| AbM water extract, p.o. | Human case (n = 1), Contact dermatitis | Allergic chron. cheilitis, pos. delayed reaction after patch testing | Daily AbM intake for 6 months | Suehiro et al., 2007 [99] |
| AbM extract, p.o. | Cancer patients (n = 3), Liver function | Severe hepatic dysfunction | - | Mukai et al., 2006 [98] |
| GF polysacc., p.o. | Phase I/II safety study in 34 breast ca. pat. | No dose-limiting toxicity | - | Deng et al., 2009 [105] |
| GF spore, inhalation | Human case (n = 1), pneumonitis | Occupational hypersensitivity pneumonitis | Work for 3 mo. in mushroom farm | Tanaka et al., 2004 [104] |
| Erinacine A-enriched HE mycelia, p.o. | Mice, Life-prolonging activity | Increased longevity in aged mice | Induction of endogenous antioxidant enzymes | Li et al., 2019 [75] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hetland, G.; Tangen, J.-M.; Mahmood, F.; Mirlashari, M.R.; Nissen-Meyer, L.S.H.; Nentwich, I.; Therkelsen, S.P.; Tjønnfjord, G.E.; Johnson, E. Antitumor, Anti-inflammatory and Antiallergic Effects of Agaricus blazei Mushroom Extract and the Related Medicinal Basidiomycetes Mushrooms, Hericium erinaceus and Grifola frondosa: A Review of Preclinical and Clinical Studies. Nutrients 2020, 12, 1339. https://doi.org/10.3390/nu12051339
Hetland G, Tangen J-M, Mahmood F, Mirlashari MR, Nissen-Meyer LSH, Nentwich I, Therkelsen SP, Tjønnfjord GE, Johnson E. Antitumor, Anti-inflammatory and Antiallergic Effects of Agaricus blazei Mushroom Extract and the Related Medicinal Basidiomycetes Mushrooms, Hericium erinaceus and Grifola frondosa: A Review of Preclinical and Clinical Studies. Nutrients. 2020; 12(5):1339. https://doi.org/10.3390/nu12051339
Chicago/Turabian StyleHetland, Geir, Jon-Magnus Tangen, Faiza Mahmood, Mohammad Reza Mirlashari, Lise Sofie Haug Nissen-Meyer, Ivo Nentwich, Stig Palm Therkelsen, Geir Erland Tjønnfjord, and Egil Johnson. 2020. "Antitumor, Anti-inflammatory and Antiallergic Effects of Agaricus blazei Mushroom Extract and the Related Medicinal Basidiomycetes Mushrooms, Hericium erinaceus and Grifola frondosa: A Review of Preclinical and Clinical Studies" Nutrients 12, no. 5: 1339. https://doi.org/10.3390/nu12051339
APA StyleHetland, G., Tangen, J.-M., Mahmood, F., Mirlashari, M. R., Nissen-Meyer, L. S. H., Nentwich, I., Therkelsen, S. P., Tjønnfjord, G. E., & Johnson, E. (2020). Antitumor, Anti-inflammatory and Antiallergic Effects of Agaricus blazei Mushroom Extract and the Related Medicinal Basidiomycetes Mushrooms, Hericium erinaceus and Grifola frondosa: A Review of Preclinical and Clinical Studies. Nutrients, 12(5), 1339. https://doi.org/10.3390/nu12051339