Diminished Vitamin D Receptor Protein Levels in Crohn’s Disease Fibroblasts: Effects of Vitamin D

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Primary Fibroblasts and Epithelial Cells Isolation
2.3. Fibroblasts Wound Healing Assay
2.4. Immunohistochemical Studies
2.5. Mice
2.6. Induction of Intestinal Fibrosis by Heterotopic Transplant of Colonic Tissue and Vitamin D Treatment
2.7. Sirius Red Staining
2.8. RNA Extraction and Real-Time Quantitative PCR (RT-qPCR)
2.9. Protein Extraction and Western Blot Analysis
2.10. Statistical Analysis
3. Results
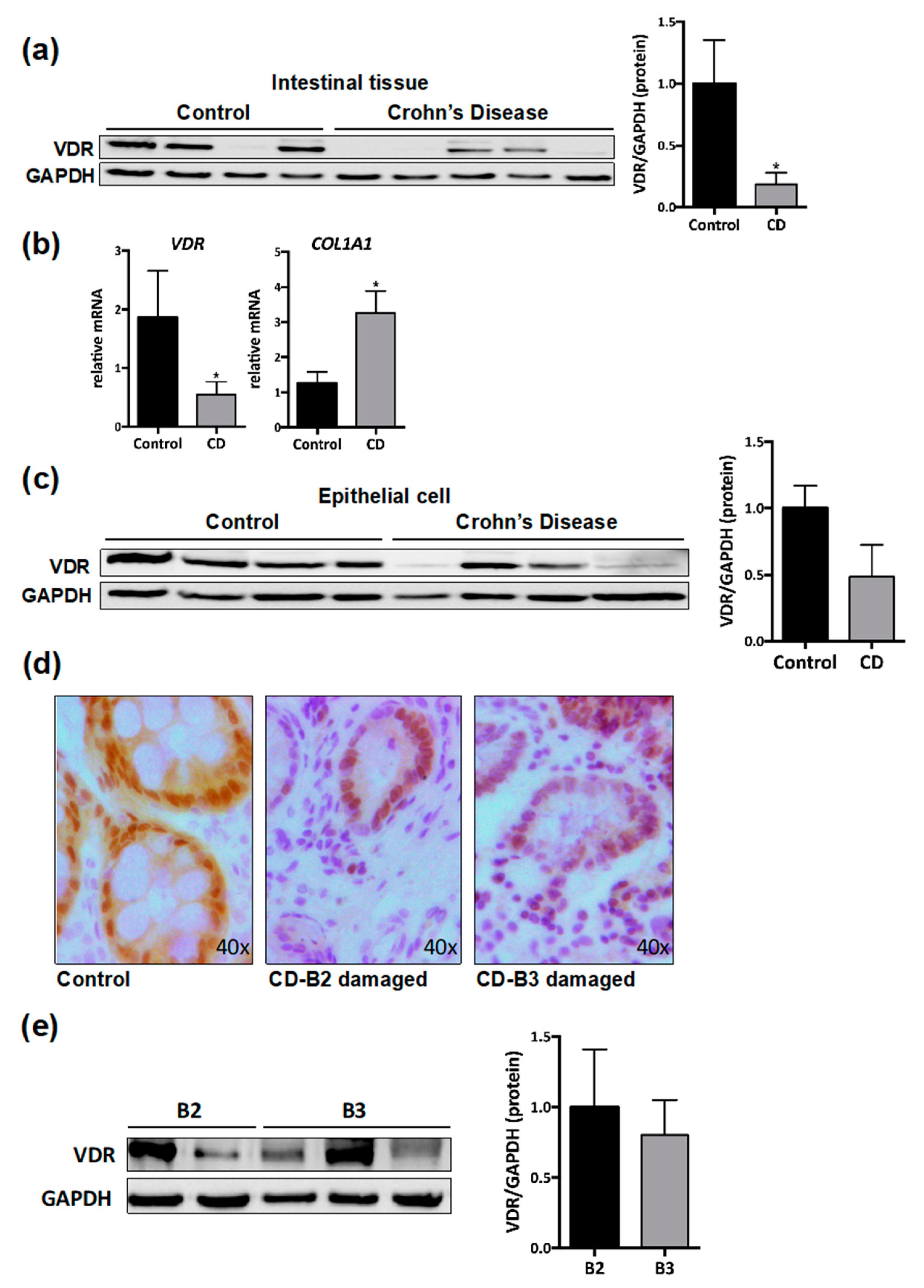
3.1. VDR Expression Is Diminished in Intestinal Resections of CD Patients
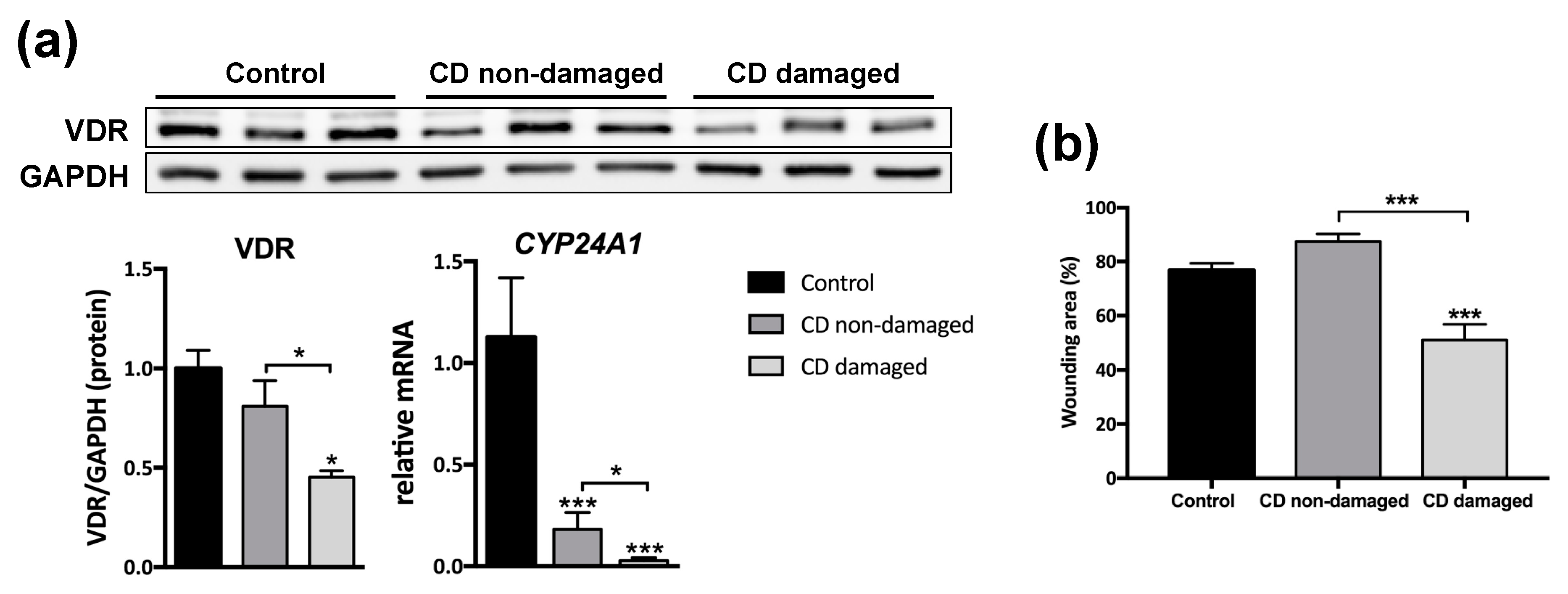
3.2. Reduced VDR Protein Levels Are Associated with Increased Migration in Fibroblasts from CD Patients
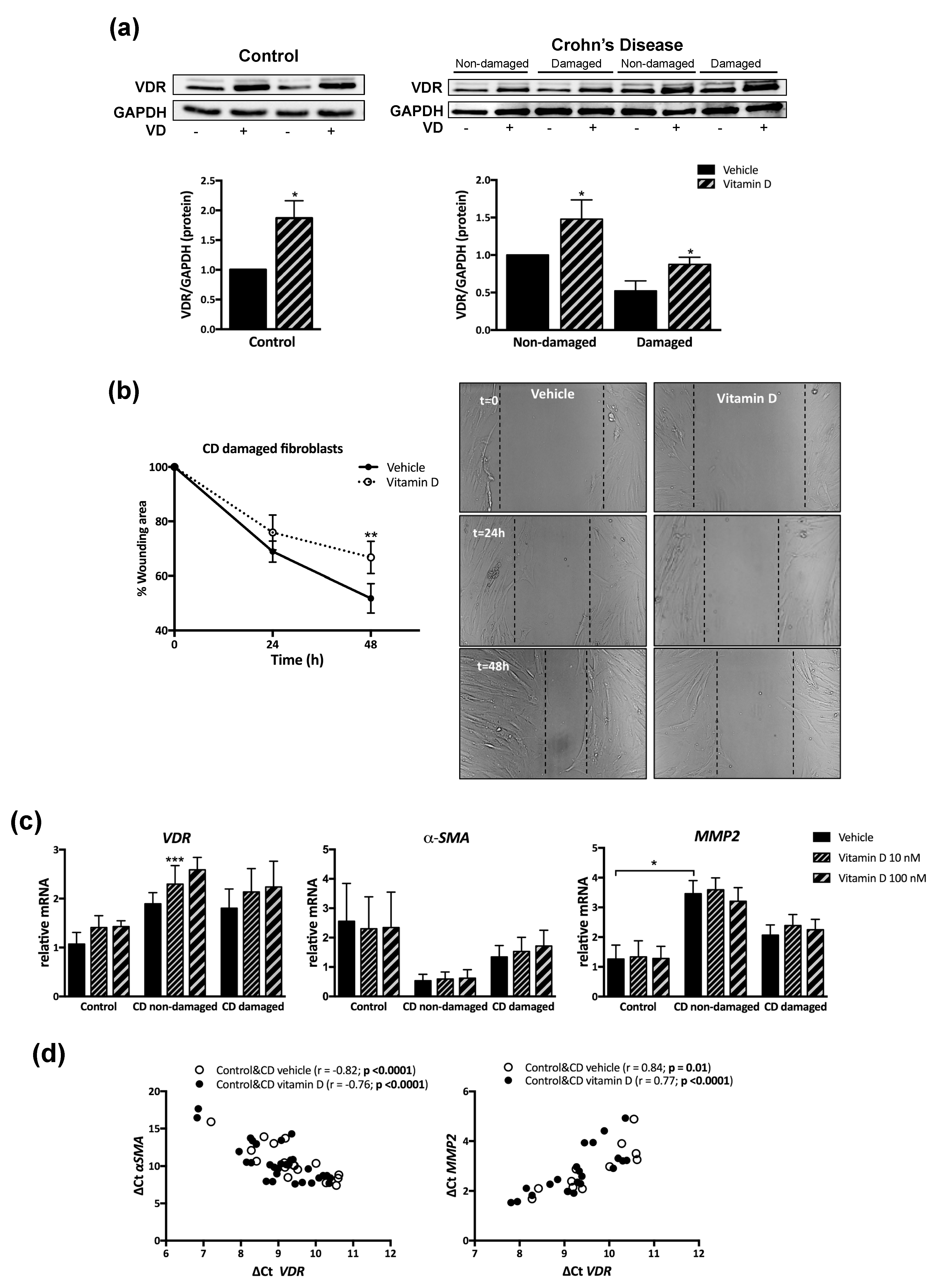
3.3. VD Increased VDR Protein Levels and Prevented the Accelerated Migration in Fibroblasts from CD Patients
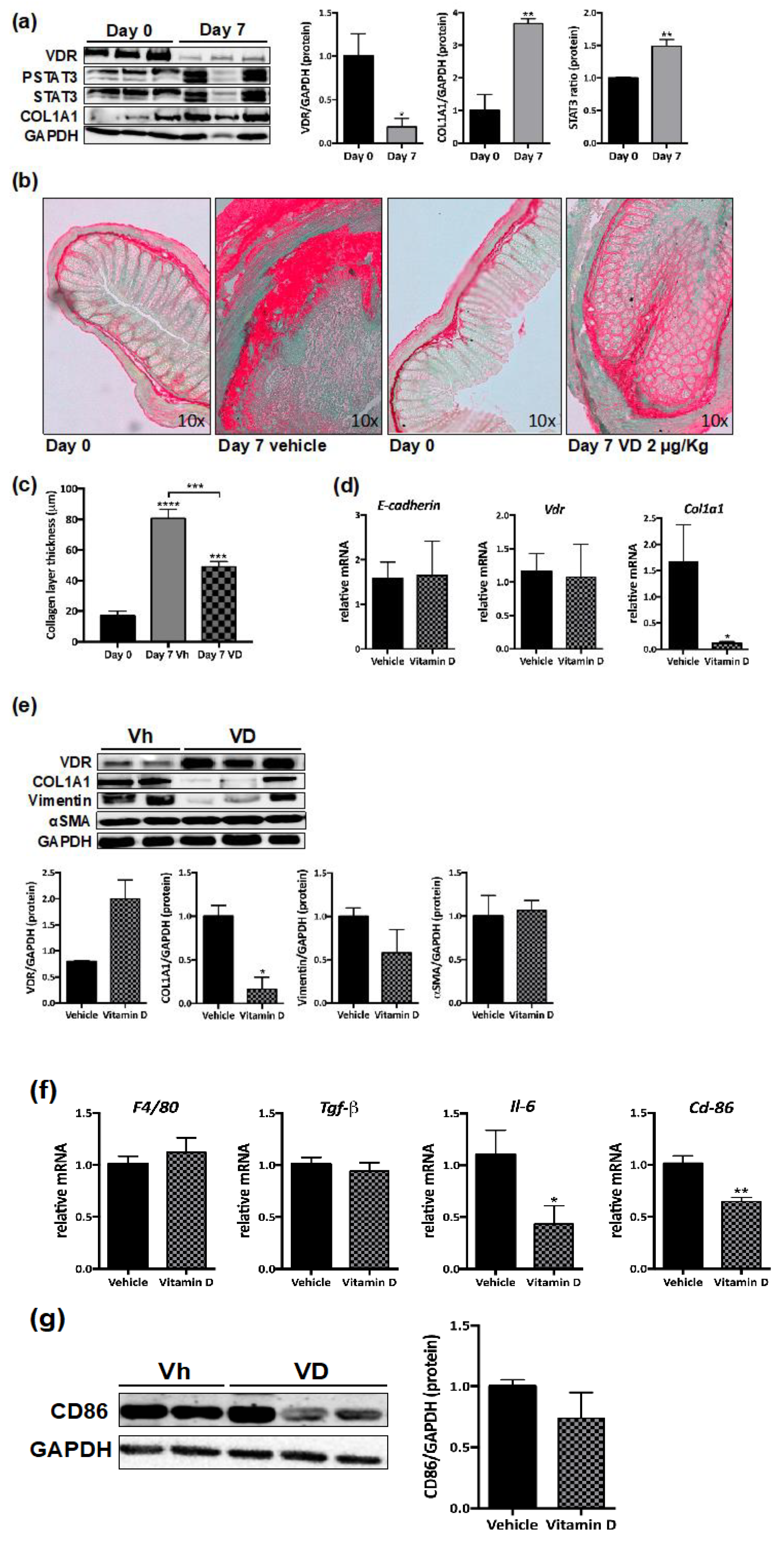
3.4. Reduced VDR Expression in Murine Intestinal Fibrosis
3.5. VD Reduces Murine Intestinal Fibrosis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ramos, G.P.; Papadakis, K.A. Mechanisms of Disease: Inflammatory Bowel Diseases. Mayo Clin. Proc. 2019, 94, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Danese, S.; Bonovas, S.; Lopez, A.; Fiorino, G.; Sandborn, W.J.; Rubin, D.T.; Kamm, M.A.; Colombel, J.F.; Sands, B.E.; Vermeire, S.; et al. Identification of Endpoints for Development of Antifibrosis Drugs for Treatment of Crohn’s Disease. Gastroenterology 2018, 155, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Cannito, S.; Novo, E.; Parola, M. Therapeutic pro-fibrogenic signaling pathways in fibroblasts. Adv. Drug Deliv. Rev. 2017, 121, 57–84. [Google Scholar] [CrossRef] [PubMed]
- Herrera, J.; Henke, C.A.; Bitterman, P.B. Extracellular matrix as a driver of progressive fibrosis. J. Clin. Investig. 2018, 128, 45–53. [Google Scholar] [CrossRef]
- Szabò, H.; Fiorino, G.; Spinelli, A.; Rovida, S.; Repici, A.; Malesci, A.C.; Danese, S. Review Article: Anti-Fibrotic Agents for the Treatment of Crohn’s Disease—Lessons Learnt from Other Diseases. Aliment. Pharmacol. Ther. 2010, 31, 189–201. [Google Scholar] [CrossRef]
- D’Haens, G.; Rieder, F.; Feagan, B.G.; Higgins, P.D.R.; Panes, J.; Maaser, C.; Rogler, G.; Löwenberg, M.; van der Voort, R.; Pinzani, M.; et al. Challenges in the Pathophysiology, Diagnosis and Management of Intestinal Fibrosis in Inflammatory Bowel Disease. Gastroenterology 2019. [Google Scholar] [CrossRef]
- Powell, D.W.; Mifflin, R.C.; Valentich, J.D.; Crowe, S.E.; Saada, J.I.; West, A.B. Myofibroblasts. II. Intestinal Subepithelial Myofibroblasts. Am. J. Physiol. 1999, 277, C183–C201. [Google Scholar] [CrossRef]
- Bettenworth, D.; Rieder, F. Pathogenesis of Intestinal Fibrosis in Inflammatory Bowel Disease and Perspectives for Therapeutic Implication. Dig. Dis. 2017, 35, 25–31. [Google Scholar] [CrossRef]
- Harries, A.D.; Brown, R.; Heatley, R.V.; Williams, L.A.; Woodhead, S.; Rhodes, J. Vitamin D Status in Crohn’s Disease: Association with Nutrition and Disease Activity. Gut 1985, 26, 1197–1203. [Google Scholar] [CrossRef]
- White, J.H. Vitamin D Deficiency and the Pathogenesis of Crohn’s Disease. J. Steroid Biochem. Mol. Biol. 2018, 175, 23–28. [Google Scholar] [CrossRef]
- Li, X.X.; Liu, Y.; Luo, J.; Huang, Z.D.; Zhang, C.; Fu, Y. Vitamin D Deficiency Associated with Crohn’s Disease and Ulcerative Colitis: A Meta-Analysis of 55 Observational Studies. J. Transl. Med. 2019, 17, 323. [Google Scholar] [CrossRef]
- Ananthakrishnan, A.N.; Cagan, A.; Gainer, V.S.; Cai, T.; Cheng, S.C.; Savova, G.; Chen, P.; Szolovits, P.; Xia, Z.; De Jager, P.L.; et al. Normalization of Plasma 25-Hydroxy Vitamin D is Associated with Reduced Risk of Surgery in Crohn’s Disease. Inflamm. Bowel Dis. 2013, 19, 1921–1927. [Google Scholar] [CrossRef]
- Hawthorne, A.B. Editorial: Clinical Benefits of Vitamin D Therapy in Inflammatory Bowel Disease. Aliment. Pharmacol. Ther. 2017, 45, 1365–1366. [Google Scholar] [CrossRef]
- Garg, M.; Rosella, O.; Rosella, G.; Wu, Y.; Lubel, J.S.; Gibson, P.R. Evaluation of a 12-Week Targeted Vitamin D Supplementation Regimen in Patients with Active Inflammatory Bowel Disease. Clin. Nutr. 2018, 37, 1375–1382. [Google Scholar] [CrossRef]
- Murdaca, G.; Tonacci, A.; Negrini, S.; Greco, M.; Borro, M.; Puppo, F.; Gangemi, S. Emerging Role of Vitamin D in Autoimmune Diseases: An Update on Evidence and Therapeutic Implications. Autoimmun. Rev. 2019, 18, 102350. [Google Scholar] [CrossRef]
- Pludowski, P.; Holick, M.F.; Grant, W.B.; Konstantynowicz, J.; Mascarenhas, M.R.; Haq, A.; Povoroznyuk, V.; Balatska, N.; Barbosa, A.P.; Karonova, T.; et al. Vitamin D supplementation guidelines. J. Steroid Biochem. Mol. Biol. 2018, 175, 125–135. [Google Scholar] [CrossRef]
- Nielsen, O.H.; Irgens Hansen, T.; Gubatan, J.M.; Jensen, K.B.; Rejnmark, L. Managing vitamin D deficiency in inflammatory bowel disease. Frontline Gastroenterol. 2019, 10, 394–400. [Google Scholar] [CrossRef]
- Pike, J.W.; Meyer, M.B.; Lee, S.M.; Onal, M.; Benkusky, N.A. The Vitamin D Receptor: Contemporary Genomic Approaches Reveal New Basic and Translational Insights. J. Clin. Investig. 2017, 127, 1146–1154. [Google Scholar] [CrossRef]
- Li, X.Y.; Boudjelal, M.; Xiao, J.H.; Peng, Z.H.; Asuru, A.; Kang, S.; Fisher, G.J.; Voorhees, J.J. 1,25-Dihydroxyvitamin D3 Increases Nuclear Vitamin D3 Receptors by Blocking Ubiquitin/Proteasome-Mediated Degradation in Human Skin. Mol. Endocrinol. 1999, 13, 1686–1694. [Google Scholar] [CrossRef]
- Ortiz-Masia, D.; Cosin-Roger, J.; Calatayud, S.; Hernandez, C.; Alos, R.; Hinojosa, J.; Apostolova, N.; Alvarez, A.; Barrachina, M.D. Hypoxic Macrophages Impair Autophagy in Epithelial Cells through Wnt1: Relevance in IBD. Mucosal Immunol. 2014, 7, 929–938. [Google Scholar] [CrossRef]
- Macias-Ceja, D.C.; Ortiz-Masia, D.; Salvador, P.; Gisbert-Ferrandiz, L.; Hernandez, C.; Hausmann, M.; Rogler, G.; Esplugues, J.V.; Hinojosa, J.; Alos, R.; et al. Succinate Receptor Mediates Intestinal Inflammation and Fibrosis. Mucosal Immunol. 2019, 12, 178–187. [Google Scholar] [CrossRef]
- Ortiz-Masia, D.; Hernandez, C.; Quintana, E.; Velazquez, M.; Cebrian, S.; Riano, A.; Calatayud, S.; Esplugues, J.V.; Barrachina, M.D. iNOS-Derived Nitric Oxide Mediates the Increase in TFF2 Expression Associated with Gastric Damage: Role of HIF-1. FASEB J. 2010, 24, 136–145. [Google Scholar] [CrossRef]
- Cosin-Roger, J.; Ortiz-Masia, D.; Calatayud, S.; Hernandez, C.; Alvarez, A.; Hinojosa, J.; Esplugues, J.V.; Barrachina, M.D. M2 Macrophages Activate WNT Signaling Pathway in Epithelial Cells: Relevance in Ulcerative Colitis. PLoS ONE 2013, 8, e78128. [Google Scholar] [CrossRef]
- Meier, R.; Lutz, C.; Cosin-Roger, J.; Fagagnini, S.; Bollmann, G.; Hunerwadel, A.; Mamie, C.; Lang, S.; Tchouboukov, A.; Weber, F.E.; et al. Decreased Fibrogenesis After Treatment with Pirfenidone in a Newly Developed Mouse Model of Intestinal Fibrosis. Inflamm. Bowel Dis. 2016, 22, 569–582. [Google Scholar] [CrossRef]
- Liu, W.; Chen, Y.; Golan, M.A.; Annunziata, M.L.; Du, J.; Dougherty, U.; Kong, J.; Musch, M.; Huang, Y.; Pekow, J.; et al. Intestinal Epithelial Vitamin D Receptor Signaling Inhibits Experimental Colitis. J. Clin. Investig. 2013, 123, 3983–3996. [Google Scholar] [CrossRef]
- Abreu-Delgado, Y.; Isidro, R.A.; Torres, E.A.; Gonzalez, A.; Cruz, M.L.; Isidro, A.A.; Gonzalez-Keelan, C.I.; Medero, P.; Appleyard, C.B. Serum Vitamin D and Colonic Vitamin D Receptor in Inflammatory Bowel Disease. World J. Gastroenterol. 2016, 22, 3581–3591. [Google Scholar] [CrossRef]
- Rafiq, S.; Ieppesen, P.B. Body Mass Index, Vitamin D, and Type 2 Diabetes: A Systematic Review and Meta-Analysis. Nutrients 2018, 10, 1182. [Google Scholar] [CrossRef]
- Holmes, E.A.; Rodney Harri, R.M.; Lucas, R.M. Low Sun Exposure and Vitamin D Deficiency as Risk Factors for Inflammatory Bowel Disease, With a Focus on Childhood Onset. Photochem. Photobiol. 2019, 95, 105–118. [Google Scholar] [CrossRef]
- Zenata, O.; Vrzal, R. Fine Tuning of Vitamin D Receptor (VDR) Activity by Post-Transcriptional and Post-Translational Modifications. Oncotarget 2017, 8, 35390–35402. [Google Scholar] [CrossRef]
- Mohri, T.; Nakajima, M.; Takagi, S.; Komagata, S.; Yokoi, T. MicroRNA Regulates Human Vitamin D Receptor. Int. J. Cancer 2009, 125, 1328–1333. [Google Scholar] [CrossRef]
- Li, F.; Zhang, A.; Shi, Y.; Ma, Y.; Du, Y. 1alpha,25-Dihydroxyvitamin D3 Prevents the Differentiation of Human Lung Fibroblasts Via microRNA-27b Targeting the Vitamin D Receptor. Int. J. Mol. Med. 2015, 36, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Li, Y.; Shao, T.; Zhao, Z.; Wang, Y.; Wu, A.; Chen, H.; Li, S.; Jiang, C.; Xu, J.; et al. Integrating Analysis Reveals microRNA-Mediated Pathway Crosstalk among Crohn’s Disease, Ulcerative Colitis and Colorectal Cancer. Mol. Biosyst. 2014, 10, 2317–2328. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Qian, J.; Yu, Z. Budesonide Up-Regulates Vitamin D Receptor Expression in Human Bronchial Fibroblasts and Enhances the Inhibitory Effect of Calcitriol on Airway Remodeling. Allergol. Immunopathol. (Madr) 2019, 47, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Panizo, S.; Carrillo-Lopez, N.; Naves-Diaz, M.; Solache-Berrocal, G.; Martinez-Arias, L.; Rodrigues-Diez, R.R.; Fernandez-Vazquez, A.; Martinez-Salgado, C.; Ruiz-Ortega, M.; Dusso, A.; et al. Regulation of miR-29b and miR-30c by Vitamin D Receptor Activators Contributes to Attenuate Uraemia-Induced Cardiac Fibrosis. Nephrol. Dial. Transplant. 2017, 32, 1831–1840. [Google Scholar] [CrossRef]
- Tao, Q.; Wang, B.; Zheng, Y.; Jiang, X.; Pan, Z.; Ren, J. Vitamin D Prevents the Intestinal Fibrosis Via Induction of Vitamin D Receptor and Inhibition of Transforming Growth Factor-Beta1/Smad3 Pathway. Dig. Dis. Sci. 2015, 60, 868–875. [Google Scholar] [CrossRef]
- Tian, Y.; Lv, G.; Yang, Y.; Zhang, Y.; Yu, R.; Zhu, J.; Xiao, L.; Zhu, J. Effects of Vitamin D on Renal Fibrosis in Diabetic Nephropathy Model Rats. Int. J. Clin. Exp. Pathol. 2014, 7, 3028–3037. [Google Scholar]
- Tzilas, V.; Bouros, E.; Barbayianni, I.; Karampitsakos, T.; Kourtidou, S.; Ntassiou, M.; Ninou, I.; Aidinis, V.; Bouros, D.; Tzouvelekis, A. Vitamin D Prevents Experimental Lung Fibrosis and Predicts Survival in Patients with Idiopathic Pulmonary Fibrosis. Pulm. Pharmacol. Ther. 2019, 55, 17–24. [Google Scholar] [CrossRef]
- Holvoet, T.; Devriese, S.; Castermans, K.; Boland, S.; Leysen, D.; Vandewynckel, Y.P.; Devisscher, L.; Van den Bossche, L.; Van Welden, S.; Dullaers, M.; et al. Treatment of Intestinal Fibrosis in Experimental Inflammatory Bowel Disease by the Pleiotropic Actions of a Local Rho Kinase Inhibitor. Gastroenterology 2017, 153, 1054–1067. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | CD | |
|---|---|---|
| Number of patients | 10 | 12 |
| Age | ||
| 17–40 years | 3 | 5 |
| >40 years | 7 | 7 |
| Sex | ||
| Female | 4 | 7 |
| Male | 6 | 5 |
| Behavior | ||
| B2 | 6 | |
| B3 | 6 |
| Gene | Sense (5′-3′) | Antisense (5′-3′) |
|---|---|---|
| VDR | TGGAGACTTTGACCGGAACG | AAGGGGCAGGTGAATAGTGC |
| CYP24A1 | ACCAGGGGAAGTGATGAAGC | TCATCCTCCCAAACGTGCTC |
| COL1A1 | GGAGCAGACGGGAGTTTCTC | CCGTTCTGTACGCAGGTGAT |
| ACTA2 (α-SMA) | GACCTTTGGCTTGGCTTGTC | AGCTGCTTCACAGGATTCCC |
| MMP2 | CATTCCCTGCAAAGAACACA | GTATTTGATGGCATCGCTCA |
| ACTB (β-actin) | GGACTTCGAGCAAGAGATGG | AGCACTGTGTTGGCGTACAG |
| Gene | Sense (5′-3′) | Antisense (5′-3′) |
|---|---|---|
| Vdr | ACAAGACCTACGACCCCACCT | AGCCGATGACCTTTTGGATGCT |
| E-cadherin | ACCCAAGCACGTATCAGGG | ACTGCTGGTCAGGATCGTTG |
| Col1a1 | CAGGCTGGTGTGATGGGATT | AAACCTCTCTCGCCTCTTGC |
| Adgre1 (F4/80) | CTTCCCAGAATCCAGTCTTTCC | TGACTCACCTTGTGGTCCTAA |
| Tgfb | GCGGACTACTATGCTAAAGAGG | TCAAAAGACAGCCACTCAGG |
| Il-6 | GAGTCCTTCAGAGAGATACAGAAAC | TGGTCTTGGTCCTTAGCCAC |
| Cd86 | GCACGGACTTGAACAACCAG | CCTTTGTAAATGGGCACGGC |
| Actb (β-actin) | GCCAACCGTGAAAAGATGACC | GAGGCATACAGGGACAGCAC |
| Antibody | Supplier | Dilution |
|---|---|---|
| VDR | 12550, Cell Signaling | 1:1000 |
| COL1A1 | 84336S, Cell Signaling | 1:1000 |
| STAT3 | ab68153, Abcam | 1:1000 |
| Phospho STAT3 | ab76315, Abcam | 1:1000 |
| Alpha SMA | PA5-16697, ThermoFisher | 1:1000 |
| Vimentin | ab92547, Abcam | 1:1000 |
| CD86 | ab53004, Abcam | 1:1000 |
| GAPDH | G9545, Sigma-Aldrich | 1:10000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gisbert-Ferrándiz, L.; Cosín-Roger, J.; Hernández, C.; Macias-Ceja, D.C.; Ortiz-Masiá, D.; Salvador, P.; Esplugues, J.V.; Hinojosa, J.; Navarro, F.; Calatayud, S.; et al. Diminished Vitamin D Receptor Protein Levels in Crohn’s Disease Fibroblasts: Effects of Vitamin D. Nutrients 2020, 12, 973. https://doi.org/10.3390/nu12040973
Gisbert-Ferrándiz L, Cosín-Roger J, Hernández C, Macias-Ceja DC, Ortiz-Masiá D, Salvador P, Esplugues JV, Hinojosa J, Navarro F, Calatayud S, et al. Diminished Vitamin D Receptor Protein Levels in Crohn’s Disease Fibroblasts: Effects of Vitamin D. Nutrients. 2020; 12(4):973. https://doi.org/10.3390/nu12040973
Chicago/Turabian StyleGisbert-Ferrándiz, Laura, Jesús Cosín-Roger, Carlos Hernández, Dulce C. Macias-Ceja, Dolores Ortiz-Masiá, Pedro Salvador, Juan V. Esplugues, Joaquín Hinojosa, Francisco Navarro, Sara Calatayud, and et al. 2020. "Diminished Vitamin D Receptor Protein Levels in Crohn’s Disease Fibroblasts: Effects of Vitamin D" Nutrients 12, no. 4: 973. https://doi.org/10.3390/nu12040973
APA StyleGisbert-Ferrándiz, L., Cosín-Roger, J., Hernández, C., Macias-Ceja, D. C., Ortiz-Masiá, D., Salvador, P., Esplugues, J. V., Hinojosa, J., Navarro, F., Calatayud, S., & Barrachina, M. D. (2020). Diminished Vitamin D Receptor Protein Levels in Crohn’s Disease Fibroblasts: Effects of Vitamin D. Nutrients, 12(4), 973. https://doi.org/10.3390/nu12040973