Plasma Levels of the Bioactive Sphingolipid Metabolite S1P in Adult Cystic Fibrosis Patients: Potential Target for Immunonutrition?

 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Collection and Storage of Blood Samples
2.3. S1P Measurements
2.3.1. Measurement of Total Plasma S1P and SPH Levels
2.3.2. Measurement of Unbound S1P Plasma Titers
2.4. Statistical Evaluation
3. Results
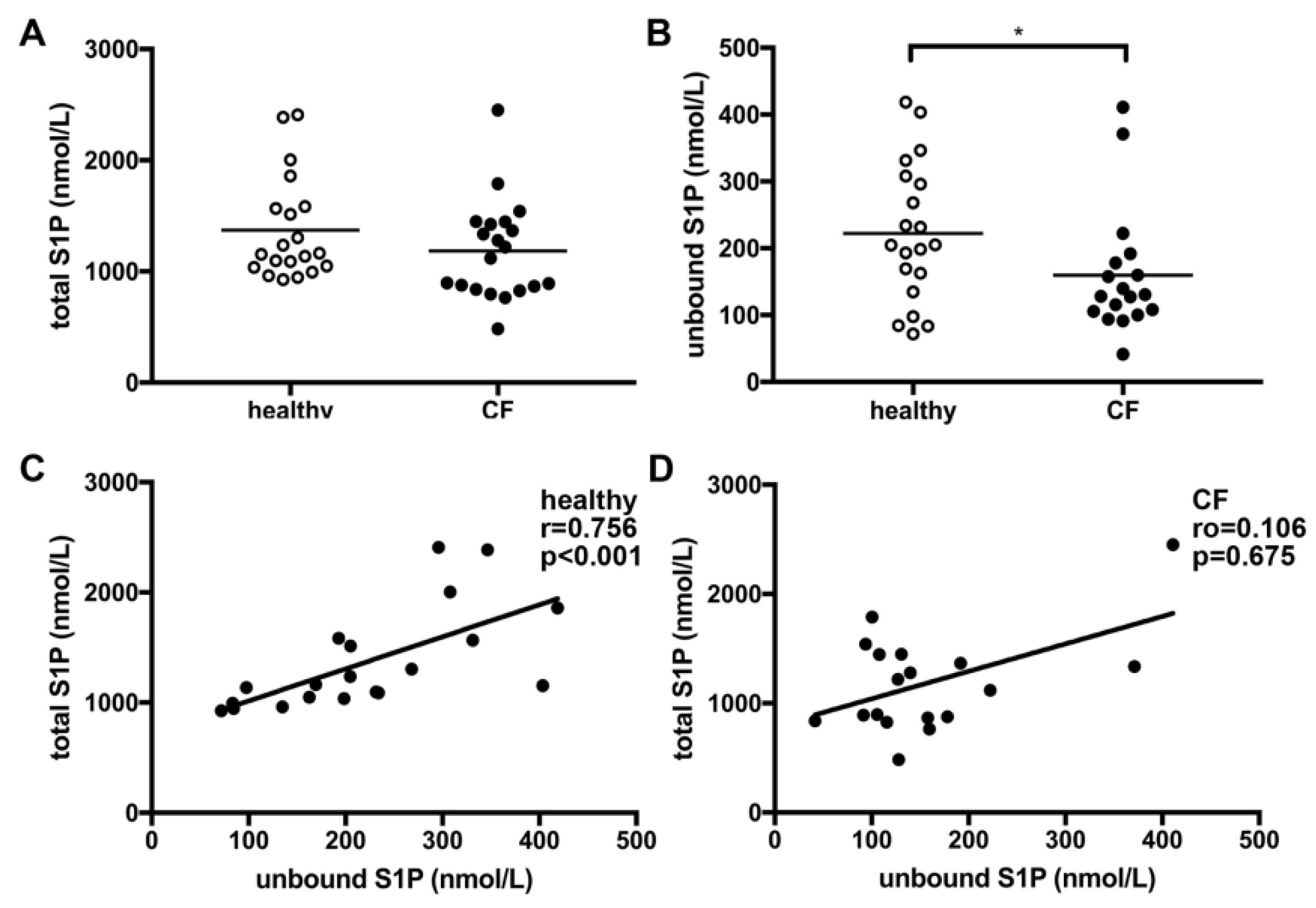
3.1. Unbound Plasma S1P Levels Were Lower in CF Patients Compared to Healthy Controls
3.2. Unbound but Not Total S1P Plasma Levels Significantly Correlate with Hemoglobin and TG Levels in CF Patients
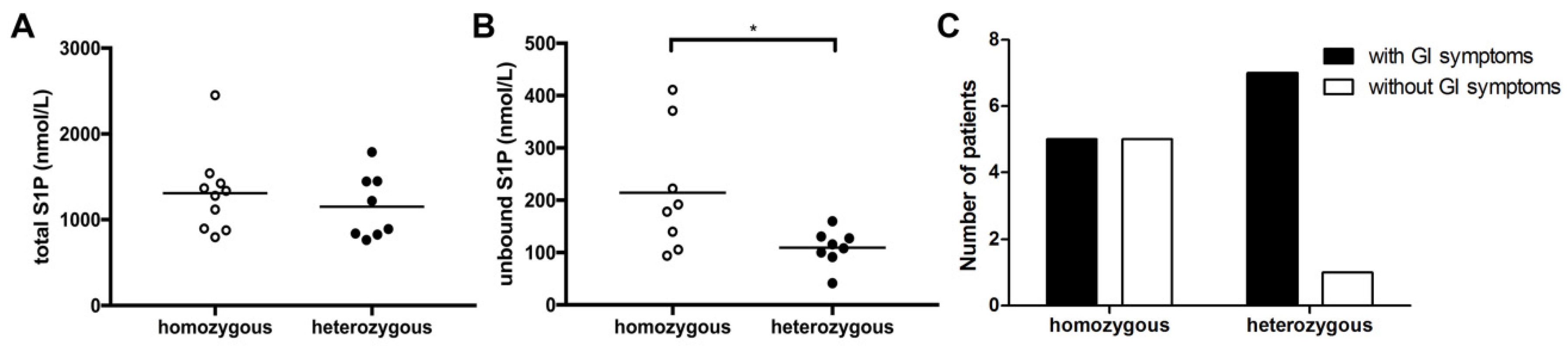
3.3. CF Genotype Is Associated with Differences in Unbound S1P Levels and GI Symptoms
3.4. Correlation of Lung Function, Lung Pathogen Colonization and Immunosuppressive Treatment with S1P Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Teichgräber, V.; Ulrich, M.; Endlich, N.; Riethmuller, J.; Wilker, B.; De Oliveira-Munding, C.C.; van Heeckeren, A.M.; Barr, M.L.; von Kurthy, G.; Schmid, K.W.; et al. Ceramide accumulation mediates inflammation, cell death and infection susceptibility in cystic fibrosis. Nat. Med. 2008, 14, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Ziobro, R.; Henry, B.; Edwards, M.J.; Lentsch, A.B.; Gulbins, E. Ceramide mediates lung fibrosis in cystic fibrosis. Biochem. Biophys. Res. Commun. 2013, 434, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, D.N.; Welsh, M.J. Structure and function of the CFTR chloride channel. Physiol. Rev. 1999, 79, S23–S45. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.A.; Henry, B.; Ziobro, R.; Tummler, B.; Gulbins, E.; Grassme, H. Role of CD95 in pulmonary inflammation and infection in cystic fibrosis. J. Mol. Med. (Berl.) 2012, 90, 1011–1023. [Google Scholar] [CrossRef] [PubMed]
- Nährlich, L.; Mainz, J.G.; Adams, C.; Engel, C.; Herrmann, G.; Icheva, V.; Lauer, J.; Deppisch, C.; Wirth, A.; Unger, K.; et al. Therapy of CF-patients with amitriptyline and placebo—A randomised, double-blind, placebo-controlled phase IIb multicenter, cohort-study. Cell. Physiol. Biochem. 2013, 31, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Riethmüller, J.; Anthonysamy, J.; Serra, E.; Schwab, M.; Döring, G.; Gulbins, E. Therapeutic efficacy and safety of amitriptyline in patients with cystic fibrosis. Cell. Physiol. Biochem. 2009, 24, 65–72. [Google Scholar] [CrossRef]
- Rivera, J.; Proia, R.L.; Olivera, A. The alliance of sphingosine-1-phosphate and its receptors in immunity. Nat. Rev. Immunol. 2008, 8, 753–763. [Google Scholar] [CrossRef]
- Hait, N.C.; Oskeritzian, C.A.; Paugh, S.W.; Milstien, S.; Spiegel, S. Sphingosine kinases, sphingosine 1-phosphate, apoptosis and diseases. Biochim. Biophys. Acta 2006, 1758, 2016–2026. [Google Scholar] [CrossRef]
- Maceyka, M.; Milstien, S.; Spiegel, S. Sphingosine-1-phosphate: The Swiss army knife of sphingolipid signaling. J. Lipid Res. 2009, 50, S272–S276. [Google Scholar] [CrossRef]
- Oskeritzian, C.A.; Milstien, S.; Spiegel, S. Sphingosine-1-phosphate in allergic responses, asthma and anaphylaxis. Pharmacol. Ther. 2007, 115, 390–399. [Google Scholar] [CrossRef]
- Karuppuchamy, T.; Behrens, E.H.; Gonzalez-Cabrera, P.; Sarkisyan, G.; Gima, L.; Boyer, J.D.; Bamias, G.; Jedlicka, P.; Veny, M.; Clark, D.; et al. Sphingosine-1-phosphate receptor-1 (S1P1) is expressed by lymphocytes, dendritic cells, and endothelium and modulated during inflammatory bowel disease. Mucosal Immunol. 2017, 10, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Maceyka, M.; Spiegel, S. Sphingolipid metabolites in inflammatory disease. Nature 2014, 510, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Sandborn, W.J.; Feagan, B.G.; Wolf, D.C.; D’Haens, G.; Vermeire, S.; Hanauer, S.B.; Ghosh, S.; Smith, H.; Cravets, M.; Frohna, P.A.; et al. Ozanimod Induction and Maintenance Treatment for Ulcerative Colitis. N. Engl. J. Med. 2016, 374, 1754–1762. [Google Scholar] [CrossRef] [PubMed]
- Malik, F.A.; Meissner, A.; Semenkov, I.; Molinski, S.; Pasyk, S.; Ahmadi, S.; Bui, H.H.; Bear, C.E.; Lidington, D.; Bolz, S.S. Sphingosine-1-Phosphate Is a Novel Regulator of Cystic Fibrosis Transmembrane Conductance Regulator (CFTR) Activity. PLoS ONE 2015, 10, e0130313. [Google Scholar] [CrossRef] [PubMed]
- Tabeling, C.; Yu, H.; Wang, L.; Ranke, H.; Goldenberg, N.M.; Zabini, D.; Noe, E.; Krauszman, A.; Gutbier, B.; Yin, J.; et al. CFTR and sphingolipids mediate hypoxic pulmonary vasoconstriction. Proc. Natl. Acad. Sci. USA 2015, 112, E1614–E1623. [Google Scholar] [CrossRef] [PubMed]
- Takabe, K.; Spiegel, S. Export of sphingosine-1-phosphate and cancer progression. J. Lipid Res. 2014, 55, 1839–1846. [Google Scholar] [CrossRef] [PubMed]
- Boujaoude, L.C.; Bradshaw-Wilder, C.; Mao, C.; Cohn, J.; Ogretmen, B.; Hannun, Y.A.; Obeid, L.M. Cystic fibrosis transmembrane regulator regulates uptake of sphingoid base phosphates and lysophosphatidic acid: Modulation of cellular activity of sphingosine 1-phosphate. J. Biol. Chem. 2001, 276, 35258–35264. [Google Scholar] [CrossRef]
- Xu, Y.; Krause, A.; Limberis, M.; Worgall, T.S.; Worgall, S. Low sphingosine-1-phosphate impairs lung dendritic cells in cystic fibrosis. Am. J. Respir. Cell Mol. Biol. 2013, 48, 250–257. [Google Scholar] [CrossRef]
- Yang, Y.; Uhlig, S. The role of sphingolipids in respiratory disease. Ther. Adv. Respir. Dis. 2011, 5, 325–344. [Google Scholar] [CrossRef]
- Norris, G.H.; Blesso, C.N. Dietary and Endogenous Sphingolipid Metabolism in Chronic Inflammation. Nutrients 2017, 9, 1180. [Google Scholar] [CrossRef]
- Hloch, O.; Charvat, J.; Fila, L.; Jan, H. Immunonutrition in patients with cyctic fibrosis leads to drop of serum amyloid A and increase of oxidative stress. J. Clin. Biochem. Nutr. 2017, 60, 176–179. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ooi, C.Y.; Durie, P.R. Cystic fibrosis from the gastroenterologist’s perspective. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 175–185. [Google Scholar] [CrossRef] [PubMed]
- World Medical Association Declaration of Helsinki: Ethical principles for medical research involving human subjects. JAMA 2013, 310, 2191–2194. [CrossRef] [PubMed]
- Hänel, P.; Andreani, P.; Graler, M.H. Erythrocytes store and release sphingosine 1-phosphate in blood. FASEB J. 2007, 21, 1202–1209. [Google Scholar] [CrossRef]
- Japtok, L.; Schaper, K.; Baumer, W.; Radeke, H.H.; Jeong, S.K.; Kleuser, B. Sphingosine 1-phosphate modulates antigen capture by murine Langerhans cells via the S1P2 receptor subtype. PLoS ONE 2012, 7, e49427. [Google Scholar] [CrossRef]
- Yatomi, Y.; Igarashi, Y.; Yang, L.; Hisano, N.; Qi, R.; Asazuma, N.; Satoh, K.; Ozaki, Y.; Kume, S. Sphingosine 1-phosphate, a bioactive sphingolipid abundantly stored in platelets, is a normal constituent of human plasma and serum. J. Biochem. 1997, 121, 969–973. [Google Scholar] [CrossRef]
- Veltman, M.; Stolarczyk, M.; Radzioch, D.; Wojewodka, G.; De Sanctis, J.B.; Dik, W.A.; Dzyubachyk, O.; Oravecz, T.; de Kleer, I.; Scholte, B.J. Correction of lung inflammation in a F508del CFTR murine cystic fibrosis model by the sphingosine-1-phosphate lyase inhibitor LX2931. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 311, L1000–L1014. [Google Scholar] [CrossRef]
- Meissner, A.; Yang, J.; Kroetsch, J.T.; Sauve, M.; Dax, H.; Momen, A.; Noyan-Ashraf, M.H.; Heximer, S.; Husain, M.; Lidington, D.; et al. Tumor necrosis factor-alpha-mediated downregulation of the cystic fibrosis transmembrane conductance regulator drives pathological sphingosine-1-phosphate signaling in a mouse model of heart failure. Circulation 2012, 125, 2739–2750. [Google Scholar] [CrossRef]
- Jolly, P.S.; Bektas, M.; Olivera, A.; Gonzalez-Espinosa, C.; Proia, R.L.; Rivera, J.; Milstien, S.; Spiegel, S. Transactivation of sphingosine-1-phosphate receptors by FcepsilonRI triggering is required for normal mast cell degranulation and chemotaxis. J. Exp. Med. 2004, 199, 959–970. [Google Scholar] [CrossRef]
- Spiegel, S.; Milstien, S. Sphingosine-1-phosphate: An enigmatic signalling lipid. Nat. Rev. Mol. Cell Biol. 2003, 4, 397–407. [Google Scholar] [CrossRef]
- Maceyka, M.; Harikumar, K.B.; Milstien, S.; Spiegel, S. Sphingosine-1-phosphate signaling and its role in disease. Trends Cell Biol. 2012, 22, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Bamias, G.; Rivera-Nieves, J. Targeting S1P Receptors, A New Mechanism of Action for Inflammatory Bowel Disease Therapy. Gastroenterology 2016, 151, 1025–1027. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Liang, J.; Nagahashi, M.; Avni, D.; Yamada, A.; Maceyka, M.; Wolen, A.R.; Kordula, T.; Milstien, S.; Takabe, K.; et al. Sphingosine-1-phosphate phosphatase 2 promotes disruption of mucosal integrity, and contributes to ulcerative colitis in mice and humans. FASEB J. 2016, 30, 2945–2958. [Google Scholar] [CrossRef] [PubMed]
- Christoffersen, C.; Obinata, H.; Kumaraswamy, S.B.; Galvani, S.; Ahnstrom, J.; Sevvana, M.; Egerer-Sieber, C.; Muller, Y.A.; Hla, T.; Nielsen, L.B.; et al. Endothelium-protective sphingosine-1-phosphate provided by HDL-associated apolipoprotein M. Proc. Natl. Acad. Sci. USA 2011, 108, 9613–9618. [Google Scholar] [CrossRef] [PubMed]
- Ksiazek, M.; Chacinska, M.; Chabowski, A.; Baranowski, M. Sources, metabolism, and regulation of circulating sphingosine-1-phosphate. J. Lipid Res. 2015, 56, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, W.J.; Lindgren, F.T.; Whalen, J.B.; Abraham, S. Serum lipoprotein concentrations in cystic fibrosis. Science 1978, 199, 783–786. [Google Scholar] [CrossRef]
- Worgall, T.S. Lipid metabolism in cystic fibrosis. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 105–109. [Google Scholar] [CrossRef]
- Worgall, T.S. Lipoprotein patterns in adult cystic fibrosis: A cause for concern or marker for survival? Respirology 2010, 15, 731–732. [Google Scholar] [CrossRef]
- Levy, E.; Lepage, G.; Bendayan, M.; Ronco, N.; Thibault, L.; Galeano, N.; Smith, L.; Roy, C.C. Relationship of decreased hepatic lipase activity and lipoprotein abnormalities to essential fatty acid deficiency in cystic fibrosis patients. J. Lipid Res. 1989, 30, 1197–1209. [Google Scholar]
- Rhodes, B.; Nash, E.F.; Tullis, E.; Pencharz, P.B.; Brotherwood, M.; Dupuis, A.; Stephenson, A. Prevalence of dyslipidemia in adults with cystic fibrosis. J. Cyst. Fibros. 2010, 9, 24–28. [Google Scholar] [CrossRef]
- Becker, K.A.; Henry, B.; Ziobro, R.; Riethmuller, J.; Gulbins, E. Lipids in cystic fibrosis. Expert Rev. Respir. Med. 2011, 5, 527–535. [Google Scholar] [CrossRef]
- Kowalski, M.P.; Pier, G.B. Localization of cystic fibrosis transmembrane conductance regulator to lipid rafts of epithelial cells is required for Pseudomonas aeruginosa-induced cellular activation. J. Immunol. 2004, 172, 418–425. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| CF Patients | Healthy Controls | |
|---|---|---|
| Total population (N (%)) | 20 | 20 |
| Male (N (%)) | 13 (65%) | 10 (50%) |
| Age (y) at inclusion (mean ± SD) | 38.5 ± 8.8 | 31.2 ± 5.7 |
| BMI (kg/m2) (mean ± SD) | 20.7 ± 2.5 | n.d. |
| LT (N (%)) | 20 (100%) | n.a. |
| Age (y) at LT (mean ± SD) | 31.9 ± 6.5 | n.a. |
| Time (m) since LT (median (min–max)) | 82.5 (5–251) | n.a. |
| Pancreas insufficiency (N (%)) | 20 (100%) | n.a. |
| Diabetes mellitus (N (%)) | 19 (95%) | n.a. |
| NODAT (N (%)) | 5 (25%) | n.a. |
| CFRD (N (%)) | 14 (75%) | n.a. |
| Immunosuppressive therapy | 20 (100%) | n.a. |
| CNI (N (%)) | 19 (95%) | n.a. |
| Steroids (N (%)) | 18 (90%) | n.a. |
| Mycofenolate mofetil (N (%)) | 6 (30%) | n.a. |
| mTOR inhibitor (N (%)) | 2 (10%) | n.a. |
| CFTR genotype, determined in (N (%)) | 20 (100%) | n.d. |
| ∆F508-homozygous (N (%)) | 10 (50%) | |
| ∆F508-heterozygous (N (%)) | 8 (40%) | |
| other than ∆F508 (N (%)) | 2 (10%) |
| ∆F508-Homozygous (n = 10) | ∆F508-Heterozygous (n = 8) | Other than ∆F508 (n = 2) | p-Value | |
|---|---|---|---|---|
| Male (N (%)) | 8 (80%) | 4 (50%) | 1 (50%) | 0.372 |
| Age (y) at inclusion (mean ± SD) | 36.9 ± 8.8 | 41.6 ± 9.2 | 34.0 5.7 | 0.286 |
| BMI (kg/m2) (mean ± SD) | 21.5 ± 2.4 | 19.5 ± 2.6 | 21.2 0.3 | 0.134 |
| Age (y) at LT (mean ± SD) | 30.5 ± 6.9 | 34.1 ± 6.6 | 29.5 0.7 | 0.278 |
| Time (m) since LT (mean ± SD) | 65 (7–251) | 95.5 (5–170) | 102.5 (96–109) | 0.706 |
| Diabetes mellitus (N (%)) | 9 (90%) | 8 (100%) | 2 (100%) | 0.608 |
| NODAT (N (%)) | 3 (30%) | 1 (13%) | 1 (50%) | |
| CFRD (N (%)) | 6 (60%) | 7 (87%) | 1 (50%) | |
| GI Symptoms (N (%)) | 4 (40%) | 6 (75%) | 2 (100%) | 0.153 |
| Pancreatin Dose (kU/d) (mean ± SD) | 418.5 ± 119.0 | 411.3 ± 214.8 | 142.5 ± 24.7 | 0.929 |
| Lung function test parameters | ||||
| FEV1 (L) (mean ± SD) | 2.9 ± 1.0 | 2.5 ± 1.2 | 1.9 ± 0.3 | 0.483 |
| FEV1 (% of predicted) (mean ± SD) | 74.3 ± 25.8 | 67.3 ± 25.3 | 61.7 ± 2.7 | 0.571 |
| FEV1/VC (mean ± SD) | 82.0 ± 17.3 | 76.3 ± 16.8 | 79.1 ± 24.5 | 0.496 |
| FEV1/VC (% of predicted) (mean ± SD) | 97.4 ± 20.5 | 91.4 ± 18.7 | 91.5 ± 26.9 | 0.531 |
| MEF50 (L) | 3.9 ± 2.2 | 3.0 ± 2.0 | 2.8 ± 2.4 | 0.381 |
| MEF50 (% of predicted) (mean ± SD) | 82.6 ± 45.0 | 64.4 ± 39.2 | 66.4 ± 55.6 | 0.396 |
| TLC (L) (mean ± SD) | 5.6 ± 0.9 | 5.2 ± 1.2 | 4.4 ± 0.6 | 0.389 |
| TLC (% of predicted) (mean ± SD) | 82.5 ± 26.8 | 88.6 ± 16.2 | 91.0 ± 16.9 | 0.582 |
| Laboratory parameters | ||||
| RBC count (G/L) (mean ± SD) | 4.7 ± 0.5 | 4.1 ± 0.5 | 3.5 ± 0.4 | 0.029 |
| Hemoglobin (mg/l) (mean ± SD) | 12.9 ± 1.7 | 11.6 ± 1.3 | 10.8 ± 0.5 | 0.085 |
| Platelets count (G/L) (mean ± SD) | 286 ± 149 | 262 ± 110 | 282 ± 33.2 | 0.704 |
| WBC count (G/L) (mean ± SD) | 7.3 ± 2.3 | 8.1 ± 3.7 | 8.0 ± 0.6 | 0.550 |
| CRP (mg/dl) (mean ± SD) | 0.99 ± 1.7 | 0.38 ± 0.25 | 0.46 ± 0.08 | 0.829 |
| Albumin (mg/dl) | 41.6 ± 3.1 | 41.4 ± 3.0 | 36.9 ± 2.3 | 0.884 |
| Cholesterol (mg/dl) (mean ± SD) | 151.8 ± 27.9 | 149.0 ± 91.7 | 179.0 ± 33.9 | 0.928 |
| HDL (mg/dl) (mean ± SD) | 46.1 ± 13.3 | 81.2 ± 47.6 | 72.0 ± 26.9 | 0.112 |
| LDL (mg/dl) (mean ± SD) | 76.7 ± 20.2 | 92.0 ± 17.7 | 76.0 ± 15.0 | 0.180 |
| TG (mg/dl) (mean ± SD) | 124.3 ± 78.5 | 141.8 ± 66.3 | 155.0 ± 110.3 | 0.351 |
| HbA1c (%) (mean ± SD) | 6.7 ± 1.4 | 6.4 ± 0.5 | 5.6 ± 0.6 | 0.601 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Halilbasic, E.; Fuerst, E.; Heiden, D.; Japtok, L.; Diesner, S.C.; Trauner, M.; Kulu, A.; Jaksch, P.; Hoetzenecker, K.; Kleuser, B.; et al. Plasma Levels of the Bioactive Sphingolipid Metabolite S1P in Adult Cystic Fibrosis Patients: Potential Target for Immunonutrition? Nutrients 2020, 12, 765. https://doi.org/10.3390/nu12030765
Halilbasic E, Fuerst E, Heiden D, Japtok L, Diesner SC, Trauner M, Kulu A, Jaksch P, Hoetzenecker K, Kleuser B, et al. Plasma Levels of the Bioactive Sphingolipid Metabolite S1P in Adult Cystic Fibrosis Patients: Potential Target for Immunonutrition? Nutrients. 2020; 12(3):765. https://doi.org/10.3390/nu12030765
Chicago/Turabian StyleHalilbasic, Emina, Elisabeth Fuerst, Denise Heiden, Lukasz Japtok, Susanne C. Diesner, Michael Trauner, Askin Kulu, Peter Jaksch, Konrad Hoetzenecker, Burkhard Kleuser, and et al. 2020. "Plasma Levels of the Bioactive Sphingolipid Metabolite S1P in Adult Cystic Fibrosis Patients: Potential Target for Immunonutrition?" Nutrients 12, no. 3: 765. https://doi.org/10.3390/nu12030765
APA StyleHalilbasic, E., Fuerst, E., Heiden, D., Japtok, L., Diesner, S. C., Trauner, M., Kulu, A., Jaksch, P., Hoetzenecker, K., Kleuser, B., Kazemi-Shirazi, L., & Untersmayr, E. (2020). Plasma Levels of the Bioactive Sphingolipid Metabolite S1P in Adult Cystic Fibrosis Patients: Potential Target for Immunonutrition? Nutrients, 12(3), 765. https://doi.org/10.3390/nu12030765