1. Introduction
Flavonoids are a diverse group of compounds which are ubiquitously distributed among plants, especially in fruits and vegetables. Being a regular and essential part of the human diet, their toxicity is considered as negligible. In addition, their biological activities, benefits to health, as well as pharmacological properties have been an important focus of several studies. Among the vast group of flavonoids, some of the most well-known compounds are the polyphenols under the subgroup flavonol, which include quercetin and its derivatives [
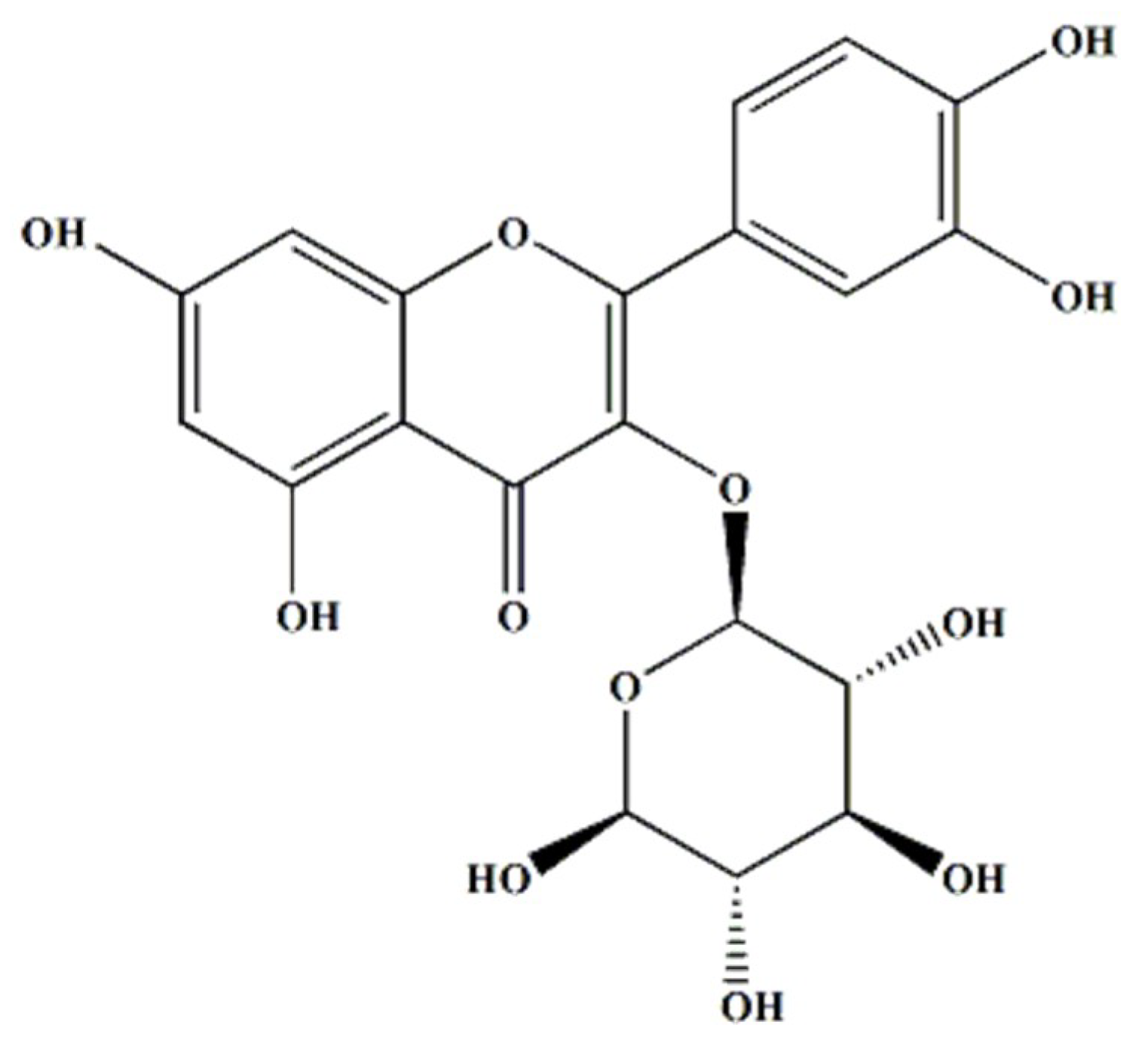
1]. Isoquercitrin (IQ) or quercetin-3-O-
β-D-glucopyranoside is a major glycosidic derivative of the naturally and abundantly occurring flavonoid quercetin, alongside rutin. Similar to other flavonols, IQ is often found in fruits, vegetables, cereals, juices and beverages, and other plant-based foods [
2]. A considerable amount of IQ has been found and isolated from Prunus mume [
3], also known as Japanese apricot, a traditional food and medicinal plant which originated from Southern part of China and was further cultivated in East Asian countries including Taiwan, Japan, and Korea [
4]. Different extracts of Prunus mume fruit, including ethyl acetate extract [
5] and methanol extract [
6], contain IQ, to which the biological activities of these extracts can be largely attributed.
Previous studies about the biological activities, specifically the chemopreventive potential, have been intensively conducted about the flavonols quercitin and rutin. Conversely, only few studies have published data about the equally important and biologically active compound, isoquercitrin. In addition, IQ is more soluble in aqueous phase and has greater bioavailability as compared with the aglycone, quercetin, making it highly suitable in many food and pharmacological applications [
2]. The biological activities of IQ were vastly influenced by its antioxidant and anti-inflammatory activities. In vitro studies conducted on IQ were mostly focused on its ROS scavenging activity, anti-angiogenesis, inhibition of melanogenesis, and anti-carcinogenic properties [
2]. In a study performed by Li et al. (2016), it was discovered that IQ exhibited greater activity in terms of the Fe
2+-binding, ET-based ferric ion reducing antioxidant power, and multi-pathways-based superoxide anion-scavenging as compared with the well-studied quercetin [
7]. Meanwhile, the findings of Matsubara et al. (2004) showed that IQ exhibits high anti-angiogenesis effect, which is also an important manifestation of anti-cancer potential [
8]. Studies about the anti-cancer properties, particularly breast, colon, liver, prostate, fibrosarcoma, pancreatic, and cerebral cancers, of isoquercitrin and its modified forms have been conducted in the past years [
9] but no recent reports were published about the mechanisms behind the effect of IQ against the melanoma cells.
Melanoma, which is brought about by malignant changes in the pigment-producing cells, is considered as one of the most aggressive types of cancer [
10,
11]. The percentage of incidence of malignant melanoma has been rising recently at a relatively fast rate, accounting for approximately 1.6% new cancer cases on an annual basis [
12]. This scenario is considered unusual considering the demographics of the occurrence of the disease such as the age and the region. The socio-economic burden of melanoma is construed to be unbalanced, and interestingly, the highest incidence was found among the younger population [
10]. The most significant risk factors were environmental aspects, particularly prolonged exposure to UV radiation, as well as the individual genetics [
10,
13]. Although advances in the treatment of melanoma, even the metastatic and malignant forms, have been made available in the past years, the efficacy of these therapies are still being explored and identification of promising combination of treatments is still under study [
14,
15].
Apoptosis is a physiological phenomenon involving the programmed cell death and is an important process for the control of cell population and systematic removal of damaged cells with impaired DNA [
16,
17]. Deregulation in the normal path of apoptosis is one of the main causative factors in many diseases including cancer development and progression. In melanoma, most anti-cancer therapy and treatment approaches commonly targeted serine/threonine protein kinase inhibitors, particularly those involved in the Ras/Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR signaling pathways [
11].
The present study aimed to investigate the effect of a major flavonol found in Prunus mume, isoquercitrin, in the viability of melanoma and normal skin cells, and to elucidate the mechanism, particularly the apoptotic cell death-related signaling pathways, behind the anti-cancer effect of IQ.
2. Materials and Methods
2.1. Chemicals and Reagents
Isoquercitrin or quercetin-3-O-
β-D-glucopyranoside (
Figure 1) was obtained from Sigma-Aldrich (St. Louis, MO, USA). Dulbecco’s modified eagle medium (DMEM), RPMI 1640 medium, fetal bovine serum (FBS), trypsin-EDTA, penicillin, and antibiotic-antimycotic were purchased from GIBCO BRL Co. (Gaithersburg, MD, USA). Sulforhodamin B (SRB), RNase, and Hoechst 33258 bis-benzamide were purchased from Sigma-Aldrich Co. Ltd. (St. Louis, Missouri USA). Anti-Bid (sc-514622), anti-Bax (sc-7480), anti-Bcl-2 (sc-7382), anti-procaspase-3, anti-procaspase-8, anti-procaspase-9, anti- Endonuclease G (Endo G) (sc-365359), anti-AIF (sc-13116), anti-poly (ADPribose) polymerase-1 (PARP-1) (sc-56197), anti-Akt (sc-5298), anti-p-Akt (sc-7985-R), anti-mTOR (sc-8319), anti-p-mTOR (sc-101738), anti-PI3K (sc-423), and anti-β-actin (sc-47778) antibodies were procured from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The ECL kit was purchased from Amersham Life Science (Amersham, UK).
2.2. Cell Culture
SK-MEL-2 (malignant melanoma cells derived from metastasis on human thigh skin), SK-MEL-28 (malignant melanoma cells derived from human skin), B16 (melanoma cells derived from mouse skin tissue), and HaCaT (normal keratinocytes derived from adult human skin) were purchased from American Type Culture Collection (ATCC, Rockville, ND, USA). The cells were cultured in RPMI (SK-MEL-2) and DMEM (SK-MEL-28, B16, and HaCaT) supplemented with 10% fetal bovine serum (FBS), penicillin and streptomycin (Gibco, ThermoScientific Co., Waltham, MA, USA). The cultures were incubated and maintained at humidified atmosphere condition with 5% CO2 at 37 °C.
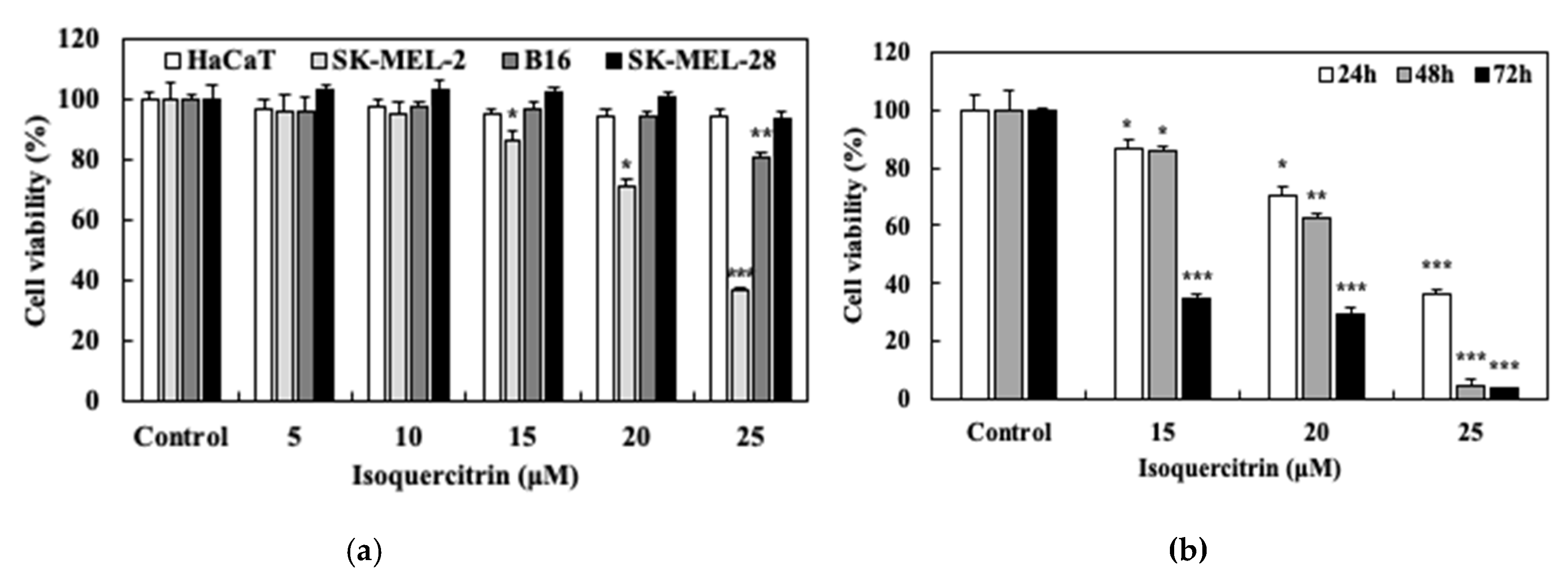
2.3. Cell Proliferation Analysis via Sulforhodamin B (SRB) Assay
The anti-proliferative activity of isoquercitrin against both the normal skin and melanoma cell lines was evaluated using the sulforhodamin B assay. The normal skin cell HaCaT and the human melanoma cells, namely SK-MEL-28, SK-MEL-2, and murine melanoma B16, were seeded at a density of 3 × 104 cells/well in 48-well cell culture plates and treated with various concentrations of isoquercitrin. After incubation for 24, 48, and 72 h, the medium was removed and 10% trichloro-acetic acid was added. After 1 h of incubation at 4 °C, the wells were washed and the cells were stained with 0.4% (w/v) SRB at room temperature for 1 h and then washed using 1% acetic acid. Bound SRB was solubilized with 10 mM Tris and the absorbance was measured at 540 nm using a microplate reader (Molecular Devices, Inc., California, USA).
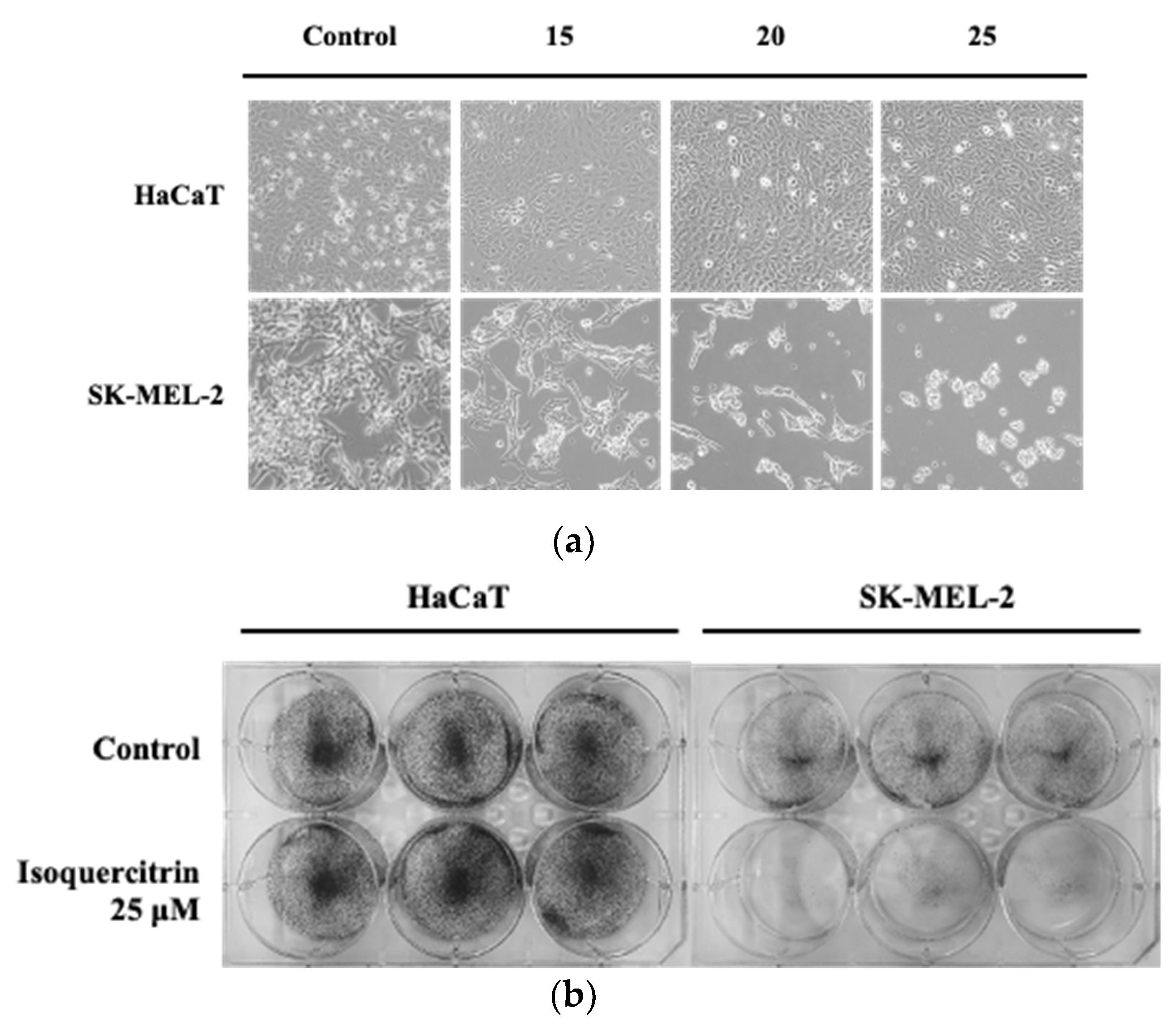
2.4. Clonogenic Survival Assay
The ability of HaCaT keratinocytes and SK-MEL-2 melanoma cells to form colonies after long-term treatment with isoquercitrin was evaluated. Briefly, 2 × 104 cells were seeded on a 6-well culture plate. After 24 h, the cells were treated with 25 μM of IQ for another 24 h and incubated at 37 °C. Fresh medium was replaced every after 24–48 h until the 14 days of incubation. Colony formation was evaluated and photographed after fixing the cells with 100% methanol and staining with 0.5% crystal violet.
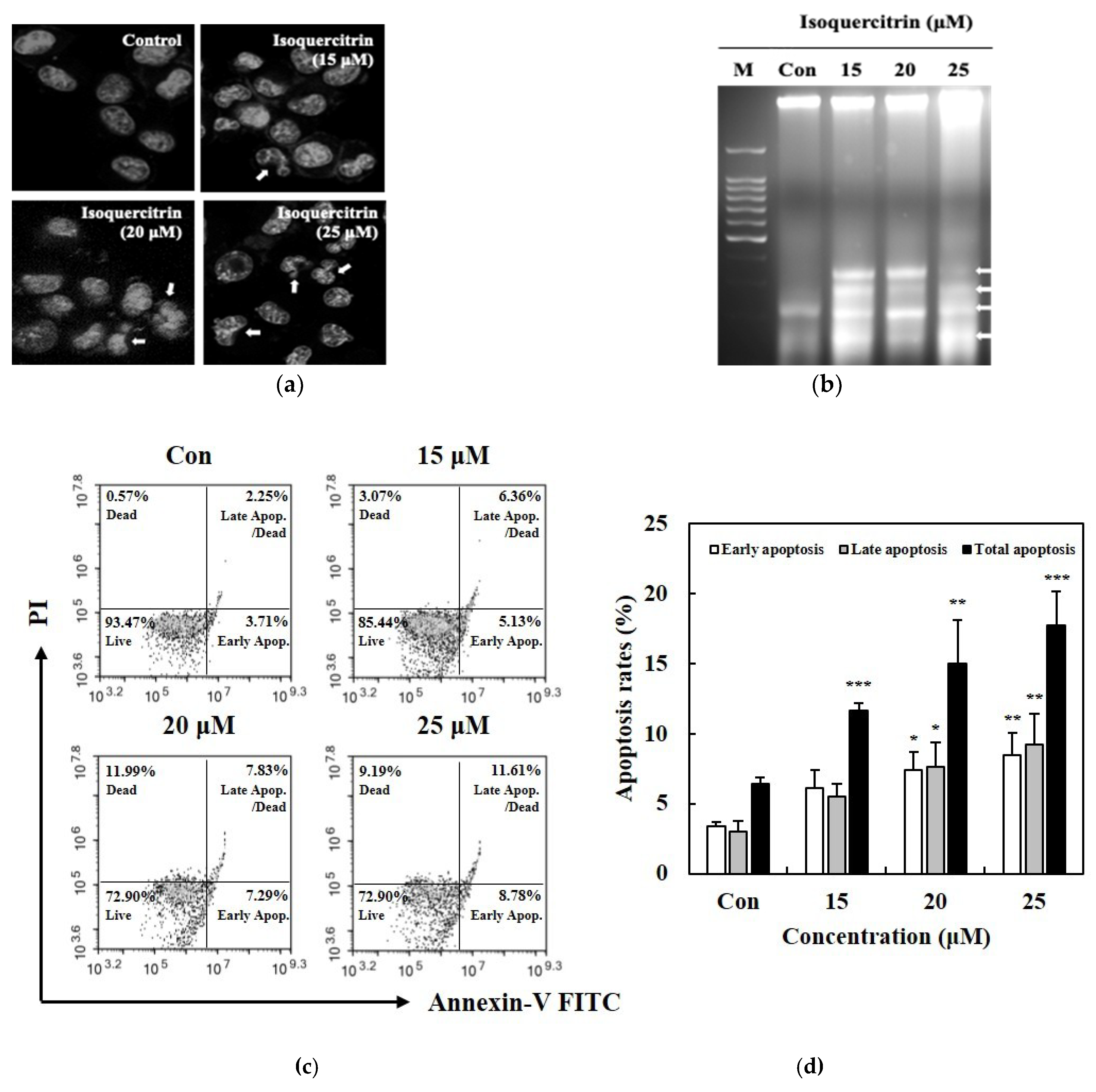
2.5. Evaluation of Morphological Apoptosis
The morphological changes in the SK-MEL-2 cells undergoing apoptosis were analyzed using the Hoechst staining assay and detected by fluorescent microscopy. The cells were first seeded in 6-well plates at a density of 1 × 106 cells per well. Then, the cells were subjected to treatment with 15, 20, and 25 μM IQ for 24 h. For the staining assay, the cells were harvested, washed with PBS and stained with 200 μL of Hoechst 33258 bis-benzamide (5 μg/mL) for 10 min at room temperature. Then, cell suspension was placed on the glass slide and examined using a fluorescence microscope (Olympus Optical, Co., Ltd., Tokyo, Japan).
2.6. DNA Fragmentation Analysis
SK-MEL-2 cells were seeded on a 100-mm dish at a density of 2 × 106 and cultured in RPMI medium for 24 h. Then, the cells were treated with the indicated concentrations of IQ for 24 h. After treatment, the cell suspensions were centrifuged and the cells were lysed using the lysis buffer consisting of 10 mM Tris-HCl, 10 mM EDTA, 0.5% Triton X-100, 20% SDS, and 10 mg/mL proteinase K. Then, extraction was performed with phenol:chloroform:isoamyl alcohol (25:24:1) solution and DNA was precipitated with cold absolute ethanol. The resulting pellets were incubated in Tris-HCl EDTA buffer and RNAse (2 mg/mL) for 1 h at 37 °C. Electrophoresis separation was performed on 2% agarose gel containing ethidium bromide. The resulting DNA bands were visualized and examined using the UV Trans illuminator imaging system (Entela Co. Ltd., Entzheim, France).
2.7. Apoptotic Cell Death Analysis Using Annexin V/PI Staining Assay
Analysis of the apoptotic cell death was performed using the Muse® Annexin V/PI and dead cell assay kit according to the manufacturer’s protocol (Merck KGaA, Darmstadt, Germany). SK-MEL-2 cells were seeded into 24-well plates at 1 × 105 cells/well density and incubated for 24 h. The indicated concentrations of IQ were treated into the cells for another 24 h. Then, the cells were trypsinized, washed with PBS, and added with the staining reagent. Suspensions of the stained cells were transferred into microcentrifuge tubes and incubated for 20 min at room temperature. Flow cytometry, obtaining data from 5000 events (gated cells) per sample, was performed using Muse cell analyzer (Merck KGaA, Darmstadt, Germany). The percentages of cells were calculated from the mean fluorescence intensity in each of the four quadrants.
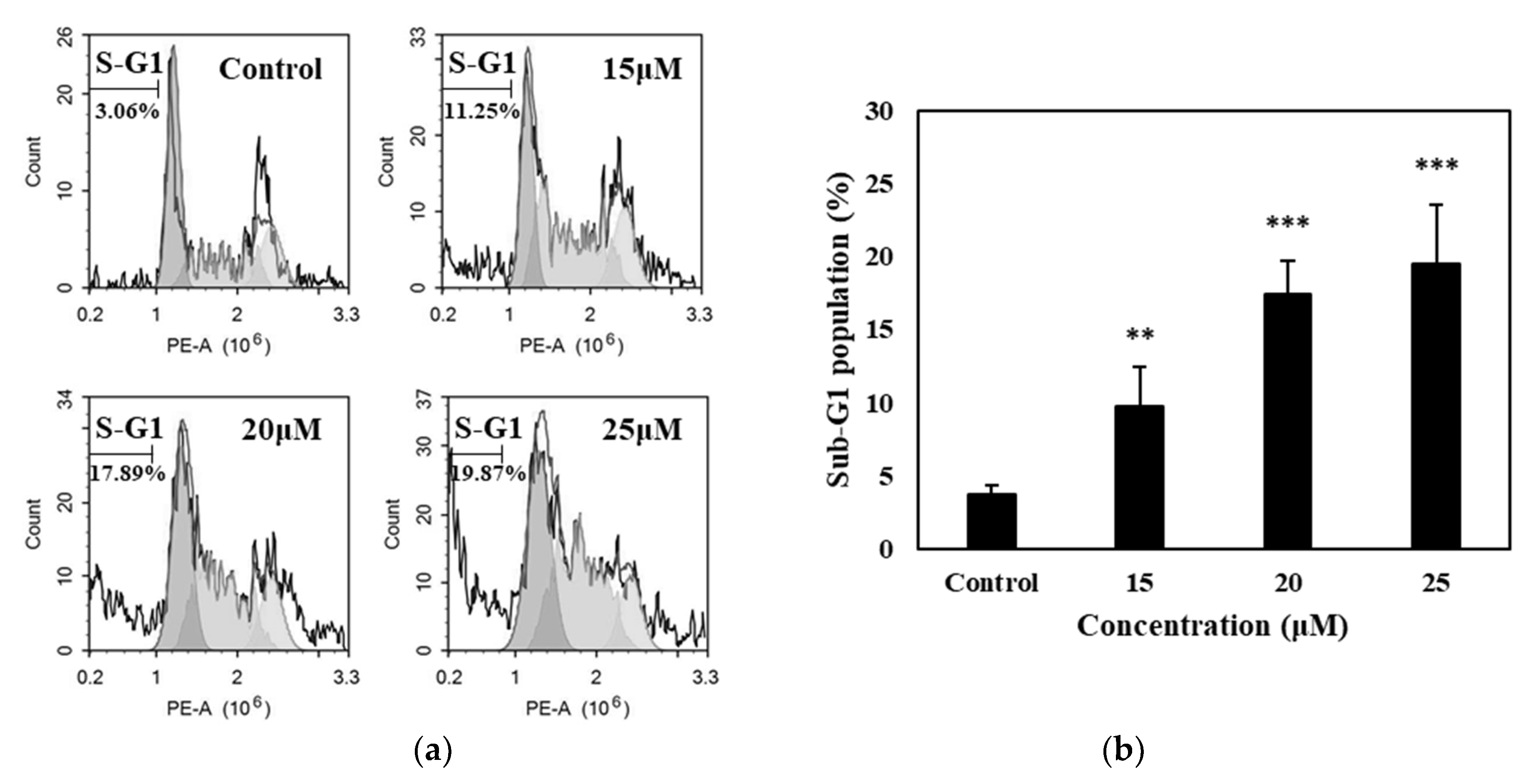
2.8. Sub-G1 Population Analysis
Flow cytometry was performed to quantify the sub-G1 population in the cell cycle phase. SK-MEL-2 melanoma cells were seeded at a density of 1 × 106 cells/well in 6-well plates and cultured in RPMI for 24 h. The cells were then collected and fixed in ice-cold 70% ethanol solution and stored at 4 °C overnight. The cells were then washed and incubated in a medium containing 1 μL of 1 mg/mL RNAse, 20 μL of 1 mg/mL propidium iodide and 500 mL PBS at 37 °C for 30 min. Flow cytometry (BD FACS, CantoTM II Flow cytometry, BD Biosciences, San Jose, USA) was used to analyze the sub-G1 DNA content.
2.9. Western Blot Analysis
SK-MEL-2 melanoma cells were seeded at a density of 5 × 106 cells in a 100-mm cell culture plate and cultured for 24 h in RPMI medium. Then, the cells were treated with 25 μM of isoquercitrin for 24 h. After the treatment time, the cell suspensions were centrifuged and the resulting pellets were lysed in a medium containing the lysis buffer (50 mM Tris-HCl, 150 mM NaCl, 1 mM EDTA, 50 nM NaF, 30 mM Na4P2O7, 1 mM PMSF, and 2 μg/mL aprotinin) for 30 min on ice. The cell lysates were obtained and the total protein content in the supernatant was measured using a Bicinchoninic acid(BCA) protein kit (Pierce, Rockford, IL, USA). The extracted protein was then subjected to separation by SDS-PAGE. Protein detection was performed using the ECL kit (Santa Cruz, CA, USA).
2.10. Statistical Analysis
All data were performed in triplicate and the data obtained were subjected to one-way analysis of variance followed by Dunnett’s test or unpaired Student’s t-test. Results are expressed as a percentage. Data values are expressed as mean ± SD of triplicate determinations. Significant difference was established using Dunnett’s test or unpaired Student’s t-test at * p < 0.05, ** p < 0.01, and *** p < 0.001. Statistical analysis was conducted using the Prism software (GraphPad, La Jolla, CA, USA).
4. Discussion
Prunus mume, Japanese apricot, has been found to contain the flavonol compounds among the fruits in the Prunus genus. Furthermore, it contains an appreciable amount of 4.9 mg of the specific flavonol isoquercitrin per 100 g dry weight of fruit [
3]. Isoquercitrin (IQ), a glycosylated derivative of the flavonoid quercetin, has gained attention as a focus for research in the past years because of its relatively higher specific biological activities, namely, its antioxidant property in terms of free-radical scavenging activity [
7], its anti-angiogenic activity [
8], as well as its anti-cancer property [
9]. Plant extracts containing IQ as well as the sole compound isolated from different plant extracts revealed its great potential for treatment of various types of cancers such as pancreatic, colon, liver, breast and glioblastoma cancers. However, the mechanism behind the anti-tumor effect of IQ, especially in malignant melanoma, still remains ambiguous. In this study, we demonstrated that IQ promotes the mitochondrial apoptotic cell death of melanoma cells, particularly the SK-MEL-2, through the modulation of the PI3K/AKT/mTOR signaling pathway, without exhibiting cytotoxicity against normal skin cells.
Studies about the chemopreventive effect of IQ showed that this compound exhibits a wide spectrum of anti-proliferative activity against different types of cancer [
9]. IQ displayed strong anti-proliferative activity against pancreatic cancer both in vitro and in vivo through induction of apoptosis and cell cycle arrest in pancreatic cancer cells [
18]. Isoquercitrin isolated from
Hyptis fasciculata also inhibited the proliferation of glioblastoma cell in a dose- and time-dependent manner [
19]. Similarly, IQ reduced the growth and viability of three selected colon cancer cell lines, but did not affect the non-tumoral cell line tested [
20]. In our experiments on the anti-proliferative activities of IQ, its treatment displayed potency against skin cancer cells, particularly the SK-MEL-2, by preventing the growth as well as the long-term survival of the tested melanoma cell without affecting the viability of the normal HaCaT keratinocytes. These findings were also in accordance with the results of previous studies on the effect of IQ on melanoma cells. De Freitas et al. (2016) showed that the ethanolic extract of
Morus nigra L. leaves, containing IQ as one of the main flavonoids, exhibited higher cytotoxicity against murine carcinoma cell line [
21]. Likewise, in the study performed by Ohguchi et al. (2010), IQ isolated from the peels of Japanese persimmon displayed an ability to inhibit melanogenesis in murine melanoma B16 [
22].
Apoptosis has played a crucial role in the development and progression of cancer; hence, it has always become an important target for cancer treatments or therapies. In the present study, we have established in the results that the treatment with IQ can lead to apoptosis in SK-MEL-2 melanoma cells as manifested in the morphological changes, DNA fragmentation, and increase in the population of cells undergoing stages of apoptosis. Accumulation of the cells in the sub-G1 phase of the cell cycle was also evident, where DNA flocculation or partial relaxation of DNA occurs, and the cells lost some of their DNA on late stage of apoptosis.
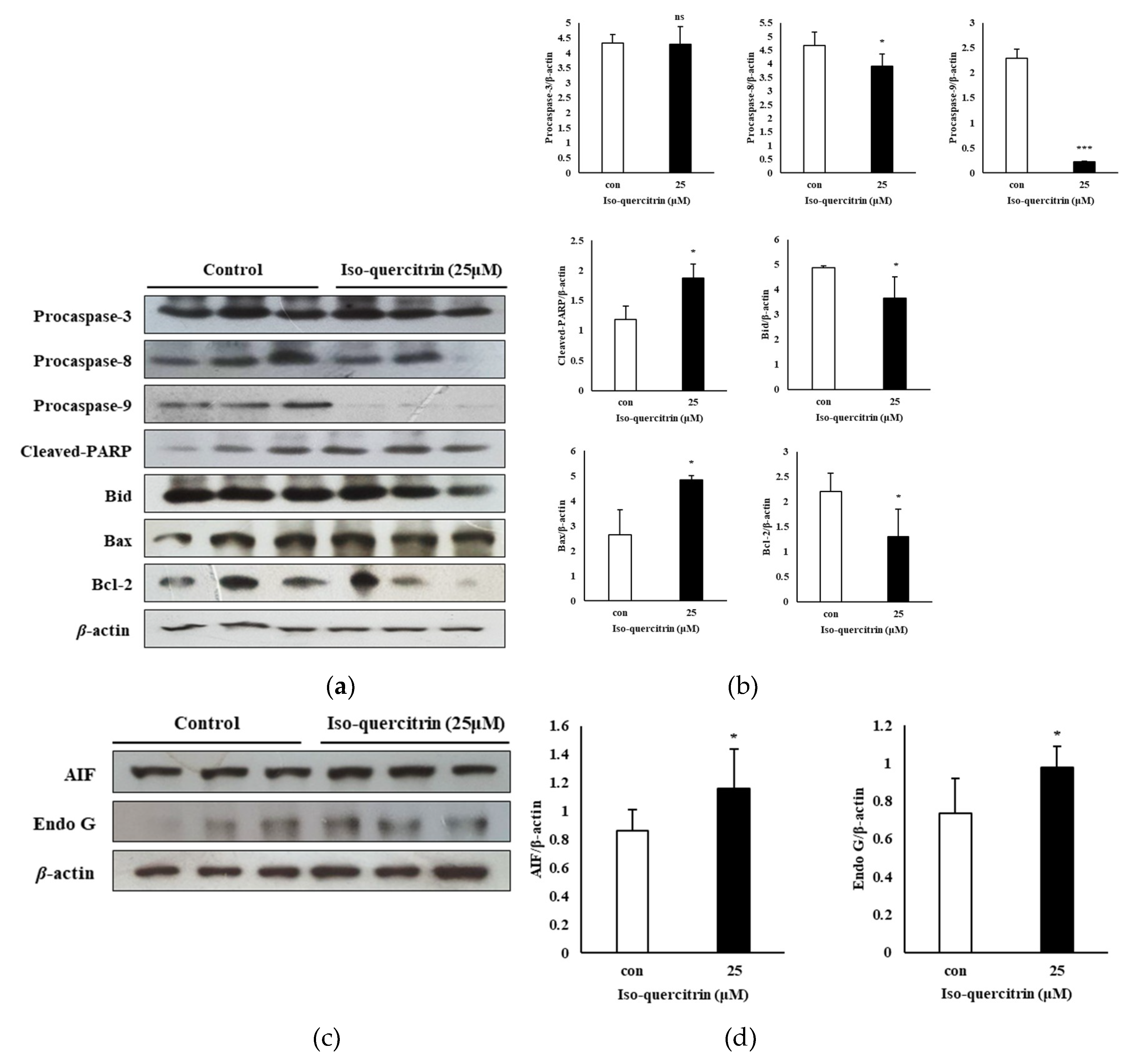
Caspases play a fundamental role in apoptosis as these molecules serve as the initiator and executioner of the cell death process [
23]. Isoquercitrin is one of the flavonoids which can target caspases [
1]. In the current study, although a significant effect in the executioner caspase-3 was not evident in the results, treatment with IQ resulted in the downregulation of the expression of procaspase-8 and-9. This action leads to self-processing and activation of initiator caspases, which promotes the commencement of the apoptosis pathway [
23,
24]. Activation of related caspases also leads to the cleavage of the cellular protein poly(ADP-ribose) polymerase-1 (PARP-1), which further results in the destruction of apoptotic cells [
25]. PARP is considered to be a pro-death signaling molecule as it serves as a nuclear and mitochondrial signal to release apoptosis inducing factor (AIF) from the mitochondria in PARP-1 dependent cell death [
26]. The results of the present study showed the ability of IQ to increase the levels of cleaved PARP in SK-MEL-2 cells. Moreover, treatment with IQ brough about significant upregulation of AIF and Endo G. These signaling molecules are apoptosis promoting proteins which are released during the mitochondrial-associated pathway and can act independently of the caspase activation [
27,
28].
The Bcl-2 family of proteins are key regulators of the intrinsic pathway of apoptosis or the mitochondrial-associated pathway in cancer because the anti-apoptotic factors belonging to this family of proteins are often overexpressed in various types of cancer and are also crucial in the ability of tumor cells to evade apoptosis [
29]. These proteins also primarily play a significant role in the regulation of the release of signaling molecules from the mitochondria which act as caspase activators [
30]. This family of proteins is comprised both of pro-apoptotic and anti-apoptotic effectors, serving as an apoptotic cell death “switch” [
17,
29]. Consequently, the Bcl-2 family is an important target for therapeutic anti-cancer agents mainly by suppressing the activity of the anti-apoptotic proteins [
23]. In the findings presented in this study, IQ treatment significantly upregulated the expression of the pro-apoptotic protein Bax while downregulating Bcl-2 expression. Taken together, the results showed that IQ brought about mitochondrial dysfunction and prompted apoptotic cell death in SK-MEL-2 melanoma cells by regulating signaling proteins associated with the mitochondrial apoptosis.
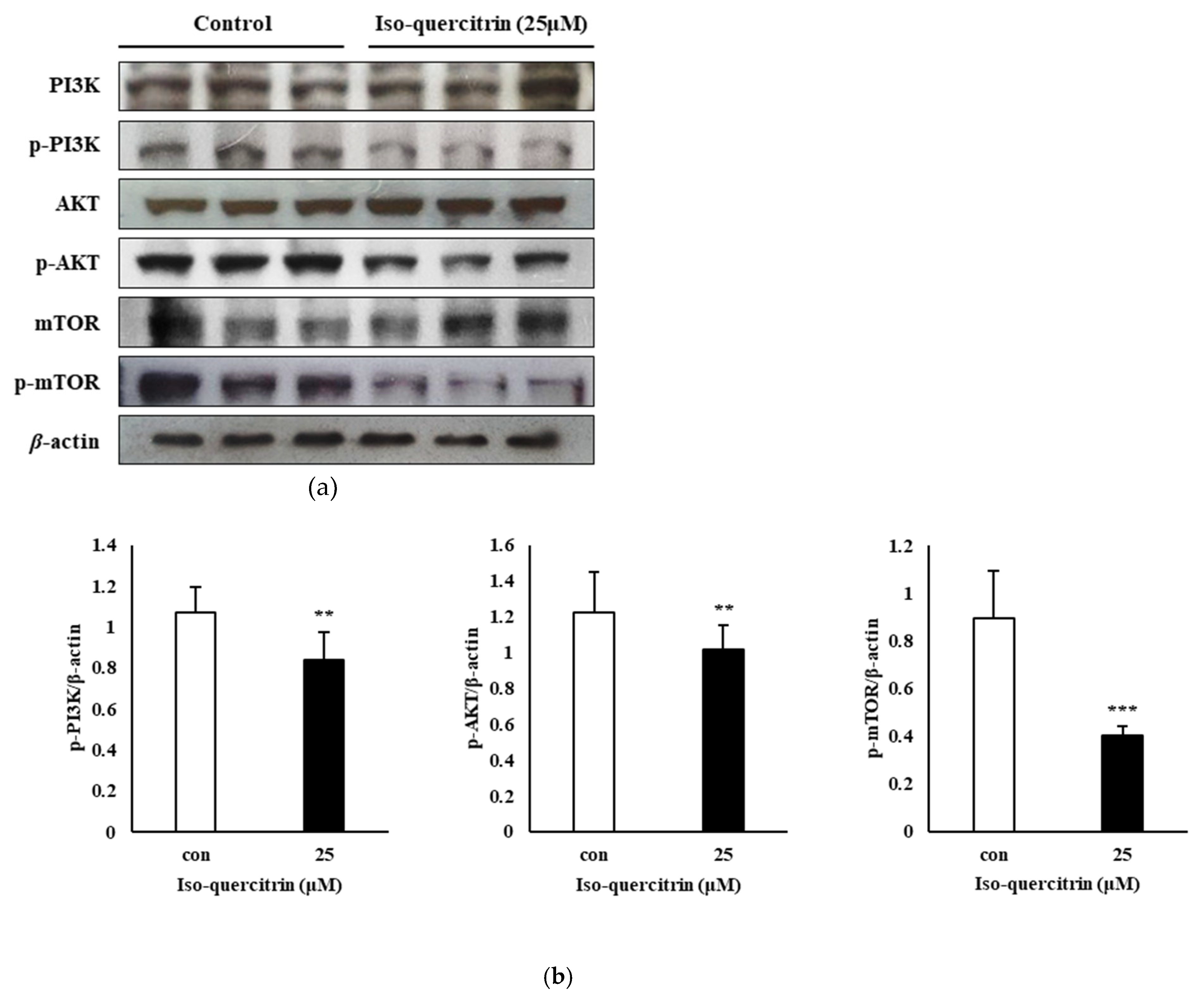
In the present study, we have also shown the involvement of the PI3K/AKT/mTOR signaling pathway in the mechanism behind the ability of IQ to inhibit the proliferation and survival of SK-MEL-2 melanoma cells. The PI3K/AKT pathway has a significant function in the cell survival mechanism and in promoting mitochondrial-associated apoptosis through its interaction with the Bcl-2 family protein, Bax. In the research conducted by Tsuruta, Masuyama, and Gotoh (2002), the PI3K/AKT pathway can block the Bax protein, leading to cell survival [
31]. Hence, suppression of the PI3K/AKT pathway plays a role in the regulation of the mitochondrial pathway leading to apoptosis. The results of this study showed that IQ exhibits an anti-proliferative property against SK-MEL-2 human skin cancer cells by downregulating the expression of phosphorylated PI3K, AKT and mTOR signaling proteins.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






