Cellular Function of Annexin A1 Protein Mimetic Peptide Ac2-26 in Human Skin Keratinocytes HaCaT and Fibroblast Detroit 551 Cells

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Reagents
2.2. Cell Viability Assay
2.3. Chemokine and Cytokine Analysis
2.4. Western Blot Analysis
2.5. RNA Isolation and Quantitative Real-Time PCR
2.6. Statistical Analysis
3. Results
3.1. Effects of Ac2-26 on HaCaT Cells Cytotoxicity
3.2. Effects of Ac2-26 on the Production of TNF-α/IFN-γ-Stimulated Inflammatory Chemokines and Cytokines in HaCaT Cells
3.3. Inhibitory Effect of Ac2-26 on Chemokines mRNA Expression in TNF-α/IFN-γ-Stimulated HaCaT Cells
3.4. Effects of Ac2-26 on MAPK Phosphorylation in TNF-α/IFN-γ-Treated HaCaT Cells
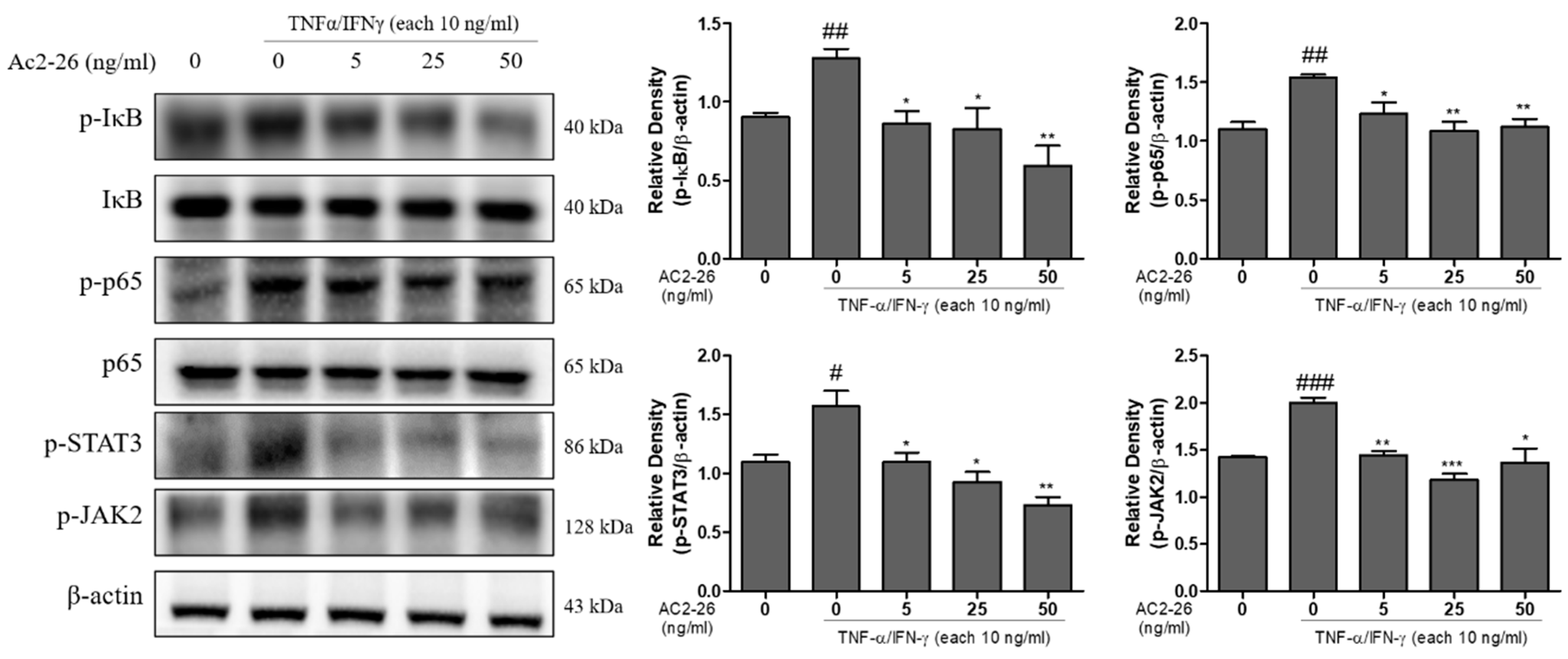
3.5. Effects of Ac2-26 on NF-κB Signaling and JAK 2/STAT 3 Phosphorylation in TNF-α/IFN-γ-Treated HaCaT Cells
3.6. Expression of Collagen on Ac2-26 Treated Human Fibroblast Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yazdi, A.S.; Rocken, M.; Ghoreschi, K. Cutaneous immunology: Basics and new concepts. Semin. Immunopathol. 2016, 38, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Welsch, K.; Holstein, J.; Laurence, A.; Ghoreschi, K. Targeting JAK/STAT signalling in inflammatory skin diseases with small molecule inhibitors. Eur. J. Immunol. 2017, 47, 1096–1107. [Google Scholar] [CrossRef] [PubMed]
- Salvioli, S.; Monti, D.; Lanzarini, C.; Conte, M.; Pirazzini, C.; Bacalini, M.G.; Garagnani, P.; Giuliani, C.; Fontanesi, E.; Ostan, R.; et al. Immune system, cell senescence, aging and longevity—Inflamm-aging reappraised. Curr. Pharm. Des. 2013, 19, 1675–1679. [Google Scholar] [PubMed]
- Subedi, L.; Lee, T.H.; Wahedi, H.M.; Baek, S.H.; Kim, S.Y. Resveratrol-Enriched Rice Attenuates UVB-ROS-Induced Skin Aging via Downregulation of Inflammatory Cascades. Oxid. Med. Cell. Longev. 2017, 2017, 8379539. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, J.; Cameron, J. Assessment of skin integrity in the elderly 1. Br. J. Community Nurs. 2008, 13, S26–S32. [Google Scholar] [CrossRef]
- Miodovnik, M.; Koren, R.; Ziv, E.; Ravid, A. The inflammatory response of keratinocytes and its modulation by vitamin D: The role of MAPK signaling pathways. J. Cell. Physiol. 2012, 227, 2175–2183. [Google Scholar] [CrossRef]
- Park, J.-H.; Kim, M.-S.; Jeong, G.-S.; Yoon, J. Xanthii fructus extract inhibits TNF-α/IFN-γ-induced Th2-chemokines production via blockade of NF-κB, STAT1 and p38-MAPK activation in human epidermal keratinocytes. J. Ethnopharmacol. 2015, 171, 85–93. [Google Scholar] [CrossRef]
- Kwon, D.J.; Bae, Y.S.; Ju, S.M.; Goh, A.R.; Youn, G.S.; Choi, S.Y.; Park, J. Casuarinin suppresses TARC/CCL17 and MDC/CCL22 production via blockade of NF-kappaB and STAT1 activation in HaCaT cells. Biochem. Biophys. Res. Commun. 2012, 417, 1254–1259. [Google Scholar] [CrossRef]
- Yoo, J.; Park, K.; Yoo, Y.; Kim, J.; Yang, H.; Shin, Y. Effects of Egg Shell Membrane Hydrolysates on Anti-Inflammatory, Anti-Wrinkle, Anti-Microbial Activity and Moisture-Protection. Korean J. Food Sci. Anim. Resour. 2014, 34, 26–32. [Google Scholar] [CrossRef]
- Liu, P.; Lee, M.K.; Choi, J.W.; Choi, Y.H.; Nam, T.J. Crude protein from spirulina increases the viability of CCD-986sk cells via the EGFR/MAPK signaling pathway. Int. J. Mol. Med. 2019, 43, 771–778. [Google Scholar] [CrossRef]
- Na, J.; Bak, D.H.; Im, S.I.; Choi, H.; Hwang, J.H.; Kong, S.Y.; No, Y.A.; Lee, Y.; Kim, B.J. Anti-apoptotic effects of glycosaminoglycans via inhibition of ERK/AP-1 signaling in TNF-α-stimulated human dermal fibroblasts. Int. J. Mol. Med. 2018, 41, 3090–3098. [Google Scholar] [CrossRef]
- Herman, M.P.; Sukhova, G.K.; Libby, P.; Gerdes, N.; Tang, N.; Horton, D.B.; Kilbride, M.; Breitbart, R.E.; Chun, M.; Schönbeck, U. Expression of neutrophil collagenase (matrix metalloproteinase-8) in human atheroma: A novel collagenolytic pathway suggested by transcriptional profiling. Circulation 2001, 104, 1899–1904. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Jung, H.J.; Choi, J.S.; Nam, T.J. Anti-wrinkle effects of a tuna heart H2O fraction on Hs27 human fibroblasts. Int. J. Mol. Med. 2016, 37, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, R.A.; Mimura, K.K.; Araujo, L.P.; Greco, K.V.; Oliani, S.M. The essential role of annexin A1 mimetic peptide in the skin allograft survival. J. Tissue Eng. Regen. Med. 2016, 10, E44–E53. [Google Scholar] [CrossRef]
- Rhee, H.J.; Kim, G.Y.; Huh, J.W.; Kim, S.W.; Na, D.S. Annexin I is a stress protein induced by heat, oxidative stress and a sulfhydryl-reactive agent. Eur. J. Biochem. 2000, 267, 3220–3225. [Google Scholar] [CrossRef]
- Gil, C.D.; La, M.; Perretti, M.; Oliani, S.M. Interaction of human neutrophils with endothelial cells regulates the expression of endogenous proteins annexin 1, galectin-1 and galectin-3. Cell Biol. Int. 2006, 30, 338–344. [Google Scholar] [CrossRef]
- Yang, Y.H.; Aeberli, D.; Dacumos, A.; Xue, J.R.; Morand, E.F. Annexin-1 regulates macrophage IL-6 and TNF via glucocorticoid-induced leucine zipper. J. Immunol. 2009, 183, 1435–1445. [Google Scholar] [CrossRef] [PubMed]
- Girol, A.P.; Mimura, K.K.; Drewes, C.C.; Bolonheis, S.M.; Solito, E.; Farsky, S.H.; Gil, C.D.; Oliani, S.M. Anti-inflammatory mechanisms of the annexin A1 protein and its mimetic peptide Ac2-26 in models of ocular inflammation in vivo and in vitro. J. Immunol. 2013, 190, 5689–5701. [Google Scholar] [CrossRef]
- Min, Y.D.; Choi, C.H.; Bark, H.; Son, H.Y.; Park, H.H.; Lee, S.; Park, J.W.; Park, E.K.; Shin, H.I.; Kim, S.H. Quercetin inhibits expression of inflammatory cytokines through attenuation of NF-kappaB and p38 MAPK in HMC-1 human mast cell line. Inflamm. Res. 2007, 56, 210–215. [Google Scholar] [CrossRef]
- Pan, X.; Cao, X.; Li, N.; Xu, Y.; Wu, Q.; Bai, J.; Yin, Z.; Luo, L.; Lan, L. Forsythin inhibits lipopolysaccharide-induced inflammation by suppressing JAK-STAT and p38 MAPK signalings and ROS production. Inflamm. Res. 2014, 63, 597–608. [Google Scholar] [CrossRef]
- Lee, H.J.; Im, A.R.; Kim, S.M.; Kang, H.S.; Lee, J.D.; Chae, S. The flavonoid hesperidin exerts anti-photoaging effect by downregulating matrix metalloproteinase (MMP)-9 expression via mitogen activated protein kinase (MAPK)-dependent signaling pathways. BMC Complement. Altern. Med. 2018, 18, 39. [Google Scholar] [CrossRef]
- Kim, S.R.; Jung, Y.R.; An, H.J.; Kim, D.H.; Jang, E.J.; Choi, Y.J.; Moon, K.M.; Park, M.H.; Park, C.H.; Chung, K.W.; et al. Anti-wrinkle and anti-inflammatory effects of active garlic components and the inhibition of MMPs via NF-kappaB signaling. PLoS ONE 2013, 8, e73877. [Google Scholar] [CrossRef]
- Gröne, A. Keratinocytes and cytokines. Vet. Immunol. Immunopathol. 2002, 88, 1–12. [Google Scholar] [CrossRef]
- Sebastiani, S.; Albanesi, C.; De, P.O.; Puddu, P.; Cavani, A.; Girolomoni, G. The role of chemokines in allergic contact dermatitis. Arch. Dermatol. Res. 2002, 293, 552–559. [Google Scholar] [CrossRef]
- Song, H.Y.; Lee, J.A.; Ju, S.M.; Yoo, K.Y.; Won, M.H.; Kwon, H.J.; Eum, W.S.; Jang, S.H.; Choi, S.Y.; Park, J. Topical transduction of superoxide dismutase mediated by HIV-1 Tat protein transduction domain ameliorates 12-O-tetradecanoylphorbol-13-acetate (TPA)-induced inflammation in mice. Biochem. Pharmacol. 2008, 75, 1348–1357. [Google Scholar] [CrossRef]
- Albanesi, C.; Pastore, S. Pathobiology of chronic inflammatory skin diseases: Interplay between keratinocytes and immune cells as a target for anti-inflammatory drugs. Curr. Drug Metab. 2010, 11, 210–227. [Google Scholar] [CrossRef] [PubMed]
- Seo, W.Y.; Youn, G.S.; Choi, S.Y.; Park, J. Butein, a tetrahydroxychalcone, suppresses pro-inflammatory responses in HaCaT keratinocytes. BMB Rep. 2015, 48, 495–500. [Google Scholar] [CrossRef]
- Yang, J.H.; Yoo, J.M.; Cho, W.K.; Ma, J.Y. Anti-inflammatory effects of Sanguisorbae Radix water extract on the suppression of mast cell degranulation and STAT-1/Jak-2 activation in BMMCs and HaCaT keratinocytes. BMC Complement. Altern. Med. 2016, 16, 347. [Google Scholar] [CrossRef]
- Yang, J.H.; Yoo, J.M.; Lee, E.; Lee, B.; Cho, W.K.; Park, K.I.; Yeul Ma, J. Anti-inflammatory effects of Perillae Herba ethanolic extract against TNF-alpha/IFN-gamma-stimulated human keratinocyte HaCaT cells. J. Ethnopharmacol. 2018, 211, 217–223. [Google Scholar] [CrossRef]
- Sung, Y.Y.; Kim, Y.S.; Kim, H.K. Illicium verum extract inhibits TNF-alpha- and IFN-gamma-induced expression of chemokines and cytokines in human keratinocytes. J. Ethnopharmacol. 2012, 144, 182–189. [Google Scholar] [CrossRef]
- Choi, J.H.; Jin, S.W.; Park, B.H.; Kim, H.G.; Khanal, T.; Han, H.J.; Hwang, Y.P.; Choi, J.M.; Chung, Y.C.; Hwang, S.K.; et al. Cultivated ginseng inhibits 2,4-dinitrochlorobenzene-induced atopic dermatitis-like skin lesions in NC/Nga mice and TNF-alpha/IFN-gamma-induced TARC activation in HaCaT cells. Food Chem. Toxicol. 2013, 56, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Hu, J.H.; Zhu, Q.G.; Li, F.Q.; Wang, J.; Sun, H.J. Effect of matrine on the expression of substance P receptor and inflammatory cytokines production in human skin keratinocytes and fibroblasts. Int. Immunopharmacol. 2007, 7, 816–823. [Google Scholar] [CrossRef] [PubMed]
- Oikarinen, A. The aging of skin: Chronoaging versus photoaging. Photodermatol. Photoimmunol. Photomed. 1990, 7, 3–4. [Google Scholar] [PubMed]
- Imokawa, G. Mechanism of UVB-induced wrinkling of the skin: Paracrine cytokine linkage between keratinocytes and fibroblasts leading to the stimulation of elastase. J. Investig. Dermatol. Symp. Proc. 2009, 14, 36–43. [Google Scholar] [CrossRef]
- Makrantonaki, E.; Zouboulis, C.C. The skin as a mirror of the aging process in the human organism—State of the art and results of the aging research in the German National Genome Research Network 2 (NGFN-2). Exp. Gerontol. 2007, 42, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Fagot, D.; Asselineau, D.; Bernerd, F. Direct role of human dermal fibroblasts and indirect participation of epidermal keratinocytes in MMP-1 production after UV-B irradiation. Arch. Dermatol. Res. 2002, 293, 576–583. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.M.; Ha, S.E.; Vetrivel, P.; Kim, H.H.; Bhosale, P.B.; Park, J.E.; Heo, J.D.; Kim, Y.S.; Kim, G.S. Cellular Function of Annexin A1 Protein Mimetic Peptide Ac2-26 in Human Skin Keratinocytes HaCaT and Fibroblast Detroit 551 Cells. Nutrients 2020, 12, 3261. https://doi.org/10.3390/nu12113261
Kim SM, Ha SE, Vetrivel P, Kim HH, Bhosale PB, Park JE, Heo JD, Kim YS, Kim GS. Cellular Function of Annexin A1 Protein Mimetic Peptide Ac2-26 in Human Skin Keratinocytes HaCaT and Fibroblast Detroit 551 Cells. Nutrients. 2020; 12(11):3261. https://doi.org/10.3390/nu12113261
Chicago/Turabian StyleKim, Seong Min, Sang Eun Ha, Preethi Vetrivel, Hun Hwan Kim, Pritam Bhagwan Bhosale, Jung Eun Park, Jeong Doo Heo, Young Sil Kim, and Gon Sup Kim. 2020. "Cellular Function of Annexin A1 Protein Mimetic Peptide Ac2-26 in Human Skin Keratinocytes HaCaT and Fibroblast Detroit 551 Cells" Nutrients 12, no. 11: 3261. https://doi.org/10.3390/nu12113261
APA StyleKim, S. M., Ha, S. E., Vetrivel, P., Kim, H. H., Bhosale, P. B., Park, J. E., Heo, J. D., Kim, Y. S., & Kim, G. S. (2020). Cellular Function of Annexin A1 Protein Mimetic Peptide Ac2-26 in Human Skin Keratinocytes HaCaT and Fibroblast Detroit 551 Cells. Nutrients, 12(11), 3261. https://doi.org/10.3390/nu12113261