Leukocyte Activation and Antioxidative Defense Are Interrelated and Moderately Modified by n-3 Polyunsaturated Fatty Acid-Enriched Eggs Consumption—Double-Blind Controlled Randomized Clinical Study

 ,
,  ,
,  ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants and Study Design
2.2. Functional Eggs Production and Fatty Acids Content
2.3. Anthropometric, Body Composition, Body Fluid Status, and Arterial Blood Pressure Measurement
2.4. Assessment of Forearm Skin Microvascular Reactivity
2.5. Venous Blood Biochemical Analysis
2.6. Profiling of Plasma Fatty Acid Concentrations
2.7. Measurement of Thiobarbituric Acid Reactive Substances (TBARS) and Ferric-Reducing Ability of Plasma (FRAP)
2.8. Measurement of Glutathione Peroxidase 1 (GPx1) and Superoxide Dismutase (SOD) Serum Activity
2.9. Measurement of Plasma Lipid Hydroperoxides (LOOH) Concentration
2.10. Flow Cytometry
2.11. Statistical Analysis
3. Results
3.1. Patients Characteristics, Hemodynamic and Biochemical Parameters
3.2. Anthropometric Measures, Body Fluid Status, and Body Composition
3.3. Plasma Fatty Acid Concentrations
3.4. Acetylcholine-Induced Dilation, Sodium Nitroprusside-Induced Dilation, and Post-Occlusive Reactive Hyperaemia in Forearm Skin Microcirculation
3.5. Markers of Oxidative Stress and Antioxidative Defence
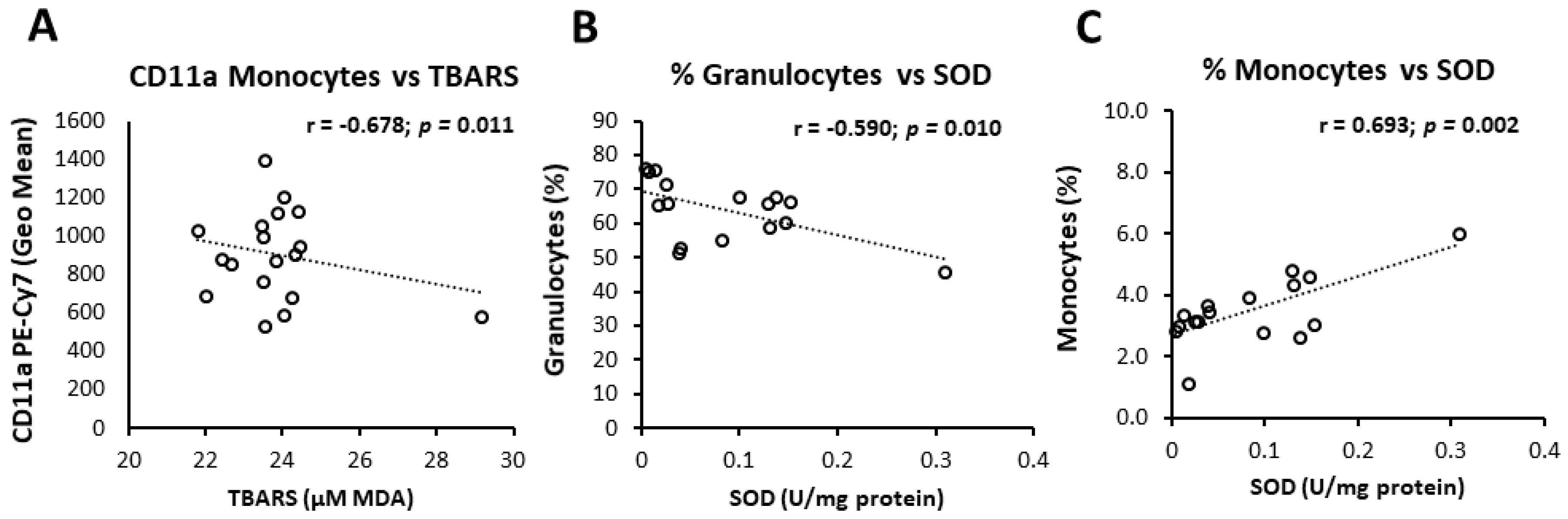
3.6. Interrelation of Vascular Measurements, Markers of Oxidative Balance, and Leukocyte Activation
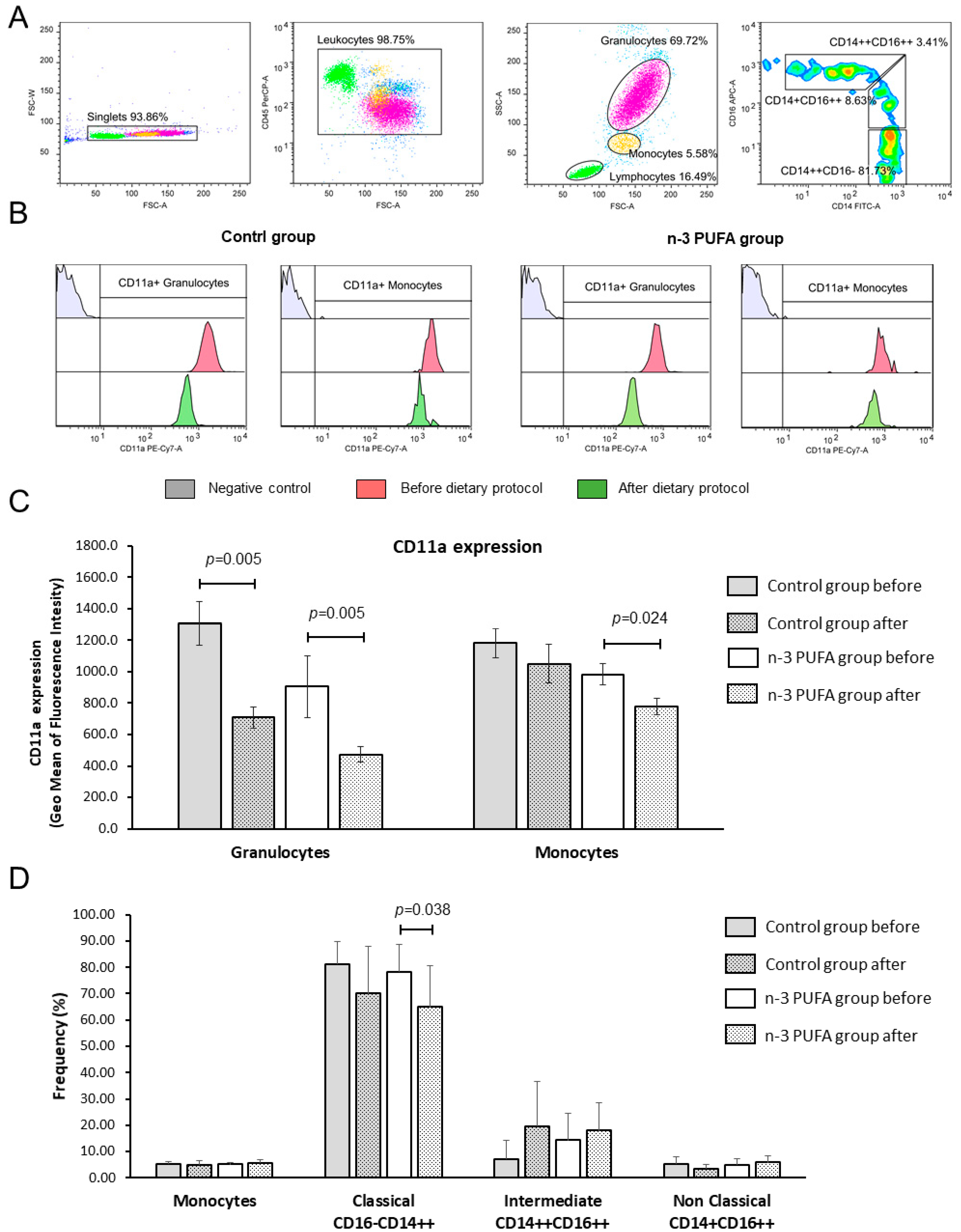
3.7. Peripheral Blood Leukocyte Subpopulations and CD11a (LFA-1) Integrin Expression
4. Discussion
4.1. n-3 PUFA Enriched Hen Eggs Effects on Microvascular Functional Responses
4.2. n-3 PUFA-Enriched Hen Eggs, Blood Pressure Level, and Serum Lipid Profile
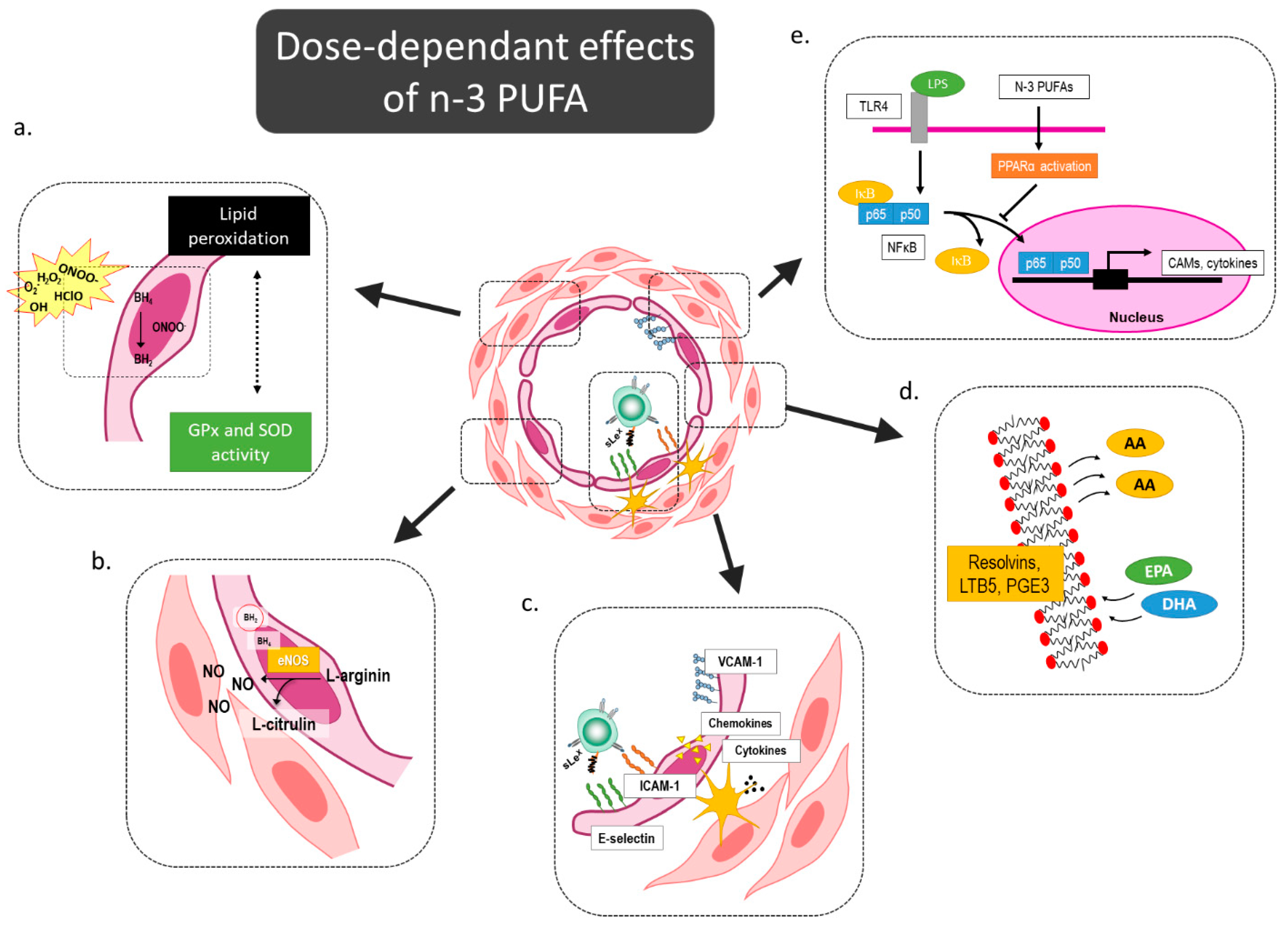
4.3. Modulation of Oxidative Stress and Antioxidative Capacity by n-3 PUFA-Enriched Diet
4.4. Supplementation with Intermediate Doses of n-3 PUFAs Deactivates Circulating Leukocytes at Comparable Levels to Regular Eggs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zehr, K.R.; Walker, M.K. Omega-3 polyunsaturated fatty acids improve endothelial function in humans at risk for atherosclerosis: A review. Prostaglandins Other. Lipid Mediat. 2018, 134, 131–140. [Google Scholar] [CrossRef]
- Thies, F.; Garry, J.M.; Yaqoob, P.; Rerkasem, K.; Williams, J.; Shearman, C.P.; Gallagher, P.J.; Calder, P.C.; Grimble, R.F. Association of n-3 polyunsaturated fatty acids with stability of atherosclerotic plaques: A randomised controlled trial. Lancet 2003, 361, 477–485. [Google Scholar] [CrossRef]
- Stupin, M.; Kibel, A.; Stupin, A.; Selthofer-Relatić, K.; Matić, A.; Mihalj, M.; Mihaljević, Z.; Jukic, I.; Drenjančević, I. The Physiological Effect of n-3 Polyunsaturated Fatty Acids (n-3 PUFAs) Intake and Exercise on Hemorheology, Microvascular Function, and Physical Performance in Health and Cardiovascular Diseases; Is There an Interaction of Exercise and Dietary n-3 PUFA Intake? Front. Physiol. 2019, 10, 1129. [Google Scholar] [CrossRef]
- Versari, D.; Daghini, E.; Virdis, A.; Ghiadoni, L.; Taddei, S. Endothelial dysfunction as a target for prevention of cardiovascular disease. Diabetes Care 2009, 32 (Suppl. S2), S314–S321. [Google Scholar] [CrossRef]
- Goodfellow, J.; Bellamy, M.F.; Ramsey, M.W.; Jones, C.J.; Lewis, M.J. Dietary supplementation with marine omega-3 fatty acids improve systemic large artery endothelial function in subjects with hypercholesterolemia. J. Am. Coll. Cardiol. 2000, 35, 265–270. [Google Scholar] [CrossRef]
- Siasos, G.; Tousoulis, D.; Oikonomou, E.; Zaromitidou, M.; Verveniotis, A.; Plastiras, A.; Kioufis, S.; Maniatis, K.; Miliou, A.; Siasou, Z.; et al. Effects of Ω-3 fatty acids on endothelial function, arterial wall properties, inflammatory and fibrinolytic status in smokers: A cross over study. Int. J. Cardiol. 2013, 166, 340–346. [Google Scholar] [CrossRef]
- Stupin, A.; Mihalj, M.; Kolobarić, N.; Šušnjara, P.; Kolar, L.; Mihaljević, Z.; Matić, A.; Stupin, M.; Jukić, I.; Kralik, Z.; et al. Anti-Inflammatory Potential of n-3 Polyunsaturated Fatty Acids Enriched Hen Eggs Consumption in Improving Microvascular Endothelial Function of Healthy Individuals—Clinical Trial. Int. J. Mol. Sci. 2020, 21, 4149. [Google Scholar] [CrossRef]
- Radzikowska, U.; Rinaldi, A.O.; Çelebi Sözener, Z.; Karaguzel, D.; Wojcik, M.; Cypryk, K.; Akdis, M.; Akdis, C.A.; Sokolowska, M. The Influence of Dietary Fatty Acids on Immune Responses. Nutrients 2019, 11, 2990. [Google Scholar] [CrossRef]
- Gutiérrez, S.; Svahn, S.L.; Johansson, M.E. Effects of Omega-3 Fatty Acids on Immune Cells. Int. J. Mol. Sci. 2019, 20, 5028. [Google Scholar] [CrossRef]
- Ito, F.; Sono, Y.; Ito, T. Measurement and Clinical Significance of Lipid Peroxidation as a Biomarker of Oxidative Stress: Oxidative Stress in Diabetes, Atherosclerosis, and Chronic Inflammation. Antioxidants 2019, 8, 72. [Google Scholar] [CrossRef]
- Shiomi, N.; Waisundara, V. Superfood and Functional Food: The Development of Superfoods and Their Roles as Medicine; BoD—Books on Demand: Norderstedt, Germany, 2017. [Google Scholar]
- Kumar, N.; Gupta, G.; Anilkumar, K.; Fatima, N.; Karnati, R.; Reddy, G.V.; Giri, P.V.; Reddanna, P. 15-Lipoxygenase metabolites of α-linolenic acid, [13-(S)-HPOTrE and 13-(S)-HOTrE], mediate anti-inflammatory effects by inactivating NLRP3 inflammasome. Sci. Rep. 2016, 6, 31649. [Google Scholar] [CrossRef] [PubMed]
- Okumura, T.; Fujioka, Y.; Morimoto, S.; Tsuboi, S.; Masai, M.; Tsujino, T.; Ohyanagi, M.; Iwasaki, T. Eicosapentaenoic acid improves endothelial function in hypertriglyceridemic subjects despite increased lipid oxidizability. Am. J. Med. Sci. 2002, 324, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, K.; Shimabukuro, M.; Higa, N.; Asahi, T.; Ohba, K.; Arasaki, O.; Higa, M.; Oshiro, Y.; Yoshida, H.; Higa, T.; et al. Eicosapentaenoic Acid supplementation changes Fatty Acid composition and corrects endothelial dysfunction in hyperlipidemic patients. Cardiol. Res. Pract. 2012, 2012, 754181. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Watts, G.F.; Burke, V.; Hilme, E.; Puddey, I.B.; Beilin, L.J. Differential effects of eicosapentaenoic acid and docosahexaenoic acid on vascular reactivity of the forearm microcirculation in hyperlipidemic, overweight men. Circulation 2000, 102, 1264–1269. [Google Scholar] [CrossRef]
- McVeigh, G.E.; Brennan, G.M.; Johnston, G.D.; McDermott, B.J.; McGrath, L.T.; Henry, W.R.; Andrews, J.W.; Hayes, J.R. Dietary fish oil augments nitric oxide production or release in patients with type 2 (non-insulin-dependent) diabetes mellitus. Diabetologia 1993, 36, 33–38. [Google Scholar] [CrossRef]
- Anon. Csapó: Chemical Composition of Milk from Red Deer—Google Znalac. Available online: https://scholar.google.com/scholar_lookup?title=Chemical+composition+of+milk+from+red+deer+roe+and+fallow+deer+kept+in+captivit&author=Csap%C3%B3,+J.&author=Sug%C3%A1r,+L.&author=Horn,+A.&author=Kiss,+C.&publication_year=1987&journal=Acta+Agron+Hung.&volume=36&pages=359%E2%80%93372 (accessed on 24 July 2020).
- Barić, L.; Drenjančević, I.; Mihalj, M.; Matić, A.; Stupin, M.; Kolar, L.; Mihaljević, Z.; Mrakovčić-Šutić, I.; Šerić, V.; Stupin, A. Enhanced Antioxidative Defense by Vitamins C and E Consumption Prevents 7-Day High-Salt Diet-Induced Microvascular Endothelial Function Impairment in Young Healthy Individuals. J. Clin. Med. 2020, 9, 843. [Google Scholar] [CrossRef]
- Stupin, A.; Stupin, M.; Baric, L.; Matic, A.; Kolar, L.; Drenjancevic, I. Sex-related differences in forearm skin microvascular reactivity of young healthy subjects. Clin. Hemorheol. Microcirc. 2019, 72, 339–351. [Google Scholar] [CrossRef]
- Stupin, M.; Stupin, A.; Rasic, L.; Cosic, A.; Kolar, L.; Seric, V.; Lenasi, H.; Izakovic, K.; Drenjancevic, I. Acute exhaustive rowing exercise reduces skin microvascular dilator function in young adult rowing athletes. Eur. J. Appl. Physiol. 2018, 118, 461–474. [Google Scholar] [CrossRef]
- Bujak, M.; Bujak, I.T.; Sobočanec, S.; Mihalj, M.; Novak, S.; Ćosić, A.; Levak, M.T.; Kopačin, V.; Mihaljević, B.; Balog, T.; et al. Trefoil Factor 3 Deficiency Affects Liver Lipid Metabolism. Cell. Physiol. Biochem. 2018, 47, 827–841. [Google Scholar] [CrossRef]
- Cosic, A.; Jukic, I.; Stupin, A.; Mihalj, M.; Mihaljevic, Z.; Novak, S.; Vukovic, R.; Drenjancevic, I. Attenuated flow-induced dilatation of middle cerebral arteries is related to increased vascular oxidative stress in rats on a short-term high salt diet. J. Physiol. (Lond.) 2016, 594, 4917–4931. [Google Scholar] [CrossRef]
- Mihaljević, Z.; Matić, A.; Stupin, A.; Rašić, L.; Jukić, I.; Drenjančević, I. Acute Hyperbaric Oxygenation, Contrary to Intermittent Hyperbaric Oxygenation, Adversely Affects Vasorelaxation in Healthy Sprague-Dawley Rats due to Increased Oxidative Stress. Oxid. Med. Cell. Longev. 2018, 2018, 7406027. [Google Scholar] [CrossRef] [PubMed]
- Vuković, R.; Blažetić, S.; Oršolić, I.; Heffer, M.; Vari, S.G.; Gajdoš, M.; Krivošíková, Z.; Kramárová, P.; Kebis, A.; Has-Schön, E. Impact of ovariectomy, high fat diet, and lifestyle modifications on oxidative/antioxidative status in the rat liver. Croat. Med. J. 2014, 55, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Novak, S.; Drenjancevic, I.; Vukovic, R.; Kellermayer, Z.; Cosic, A.; Tolusic Levak, M.; Balogh, P.; Culo, F.; Mihalj, M. Anti-Inflammatory Effects of Hyperbaric Oxygenation during DSS-Induced Colitis in BALB/c Mice Include Changes in Gene Expression of HIF-1α, Proinflammatory Cytokines, and Antioxidative Enzymes. Mediat. Inflamm. 2016, 2016, 7141430. [Google Scholar] [CrossRef] [PubMed]
- Gasparovic, A.C.; Jaganjac, M.; Mihaljevic, B.; Sunjic, S.B.; Zarkovic, N. Assays for the measurement of lipid peroxidation. Methods Mol. Biol. 2013, 965, 283–296. [Google Scholar] [CrossRef]
- Mihaljević, B.; Katusin-Razem, B.; Razem, D. The reevaluation of the ferric thiocyanate assay for lipid hydroperoxides with special considerations of the mechanistic aspects of the response. Free Radic. Biol. Med. 1996, 21, 53–63. [Google Scholar] [CrossRef]
- Matic, A.; Jukic, I.; Stupin, A.; Baric, L.; Mihaljevic, Z.; Unfirer, S.; Tartaro Bujak, I.; Mihaljevic, B.; Lombard, J.H.; Drenjancevic, I. High salt intake shifts the mechanisms of flow-induced dilation in the middle cerebral arteries of Sprague-Dawley rats. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H718–H730. [Google Scholar] [CrossRef]
- Stupin, A.; Rasic, L.; Matic, A.; Stupin, M.; Kralik, Z.; Kralik, G.; Grcevic, M.; Drenjancevic, I. Omega-3 polyunsaturated fatty acids-enriched hen eggs consumption enhances microvascular reactivity in young healthy individuals. Appl. Physiol. Nutr. Metab. 2018, 43, 988–995. [Google Scholar] [CrossRef]
- Khan, F.; Elherik, K.; Bolton-Smith, C.; Barr, R.; Hill, A.; Murrie, I.; Belch, J.J. The effects of dietary fatty acid supplementation on endothelial function and vascular tone in healthy subjects. Cardiovasc. Res. 2003, 59, 955–962. [Google Scholar] [CrossRef]
- Appel, L.J.; Miller, E.R.; Seidler, A.J.; Whelton, P.K. Does supplementation of diet with ‘fish oil’ reduce blood pressure? A meta-analysis of controlled clinical trials. Arch. Int. Med. 1993, 153, 1429–1438. [Google Scholar] [CrossRef]
- Knapp, H.R.; FitzGerald, G.A. The antihypertensive effects of fish oil. A controlled study of polyunsaturated fatty acid supplements in essential hypertension. N. Engl. J. Med. 1989, 320, 1037–1043. [Google Scholar] [CrossRef]
- Bønaa, K.H.; Bjerve, K.S.; Straume, B.; Gram, I.T.; Thelle, D. Effect of eicosapentaenoic and docosahexaenoic acids on blood pressure in hypertension. A population-based intervention trial from the Tromsø study. N. Engl. J. Med. 1990, 322, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Kestin, M.; Clifton, P.; Belling, G.B.; Nestel, P.J. n-3 fatty acids of marine origin lower systolic blood pressure and triglycerides but raise LDL cholesterol compared with n-3 and n-6 fatty acids from plants. Am. J. Clin. Nutr. 1990, 51, 1028–1034. [Google Scholar] [CrossRef] [PubMed]
- Grieger, J.A.; Miller, M.D.; Cobiac, L. Investigation of the effects of a high fish diet on inflammatory cytokines, blood pressure, and lipids in healthy older Australians. Food Nutr. Res. 2014, 58, 20369. [Google Scholar] [CrossRef]
- Hlais, S.; El-Bistami, D.; El Rahi, B.; Mattar, M.A.; Obeid, O.A. Combined fish oil and high oleic sunflower oil supplements neutralize their individual effects on the lipid profile of healthy men. Lipids 2013, 48, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Root, M.; Collier, S.R.; Zwetsloot, K.A.; West, K.L.; McGinn, M.C. A randomized trial of fish oil omega-3 fatty acids on arterial health, inflammation, and metabolic syndrome in a young healthy population. Nutr. J. 2013, 12, 40. [Google Scholar] [CrossRef]
- Oh, S.Y.; Ryue, J.; Hsieh, C.H.; Bell, D.E. Eggs enriched in omega-3 fatty acids and alterations in lipid concentrations in plasma and lipoproteins and in blood pressure. Am. J. Clin. Nutr. 1991, 54, 689–695. [Google Scholar] [CrossRef]
- Bovet, P.; Faeh, D.; Madeleine, G.; Viswanathan, B.; Paccaud, F. Decrease in blood triglycerides associated with the consumption of eggs of hens fed with food supplemented with fish oil. Nutr. Metab. Cardiovasc. Dis. 2007, 17, 280–287. [Google Scholar] [CrossRef]
- Burns-Whitmore, B.; Haddad, E.; Sabaté, J.; Rajaram, S. Effects of supplementing n-3 fatty acid enriched eggs and walnuts on cardiovascular disease risk markers in healthy free-living lacto-ovo-vegetarians: A randomized, crossover, free-living intervention study. Nutr. J. 2014, 13, 29. [Google Scholar] [CrossRef]
- Gutteridge, J.M. Lipid peroxidation and antioxidants as biomarkers of tissue damage. Clin. Chem. 1995, 41, 1819–1828. [Google Scholar] [CrossRef]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Anon. Manafa: Assessment of Superoxide Dismutase Activity—Google Znalac. Available online: https://scholar.google.com/scholar_lookup?journal=The+Journal+of+Medical+Research&title=Assessment+of+superoxide+dismutase+activity+and+total+antioxidant+capacity+in+adult+male+cigarette+smokers+in+Nnewi+metropolis,+Nigeria&author=P.+O.+Manafa&author=C.+C.+Okafor&author=C.+O.+Okeke&volume=3&issue=1&publication_year=2017&pages=23-26&doi=10.31254/jmr.2017.3109& (accessed on 23 July 2020).
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Mao, C.; Yuan, J.-Q.; Lv, Y.-B.; Gao, X.; Yin, Z.-X.; Kraus, V.B.; Luo, J.-S.; Chei, C.L.; Matchar, D.B.; Zeng, Y.; et al. Associations between superoxide dismutase, malondialdehyde and all-cause mortality in older adults: A community-based cohort study. BMC Geriatr. 2019, 19, 104. [Google Scholar] [CrossRef] [PubMed]
- Witztum, J.L. The oxidation hypothesis of atherosclerosis. Lancet 1994, 344, 793–795. [Google Scholar] [CrossRef]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Marcos, M.A.; Blázquez-Medela, A.M.; Gamella-Pozuelo, L.; Recio-Rodriguez, J.I.; García-Ortiz, L.; Martínez-Salgado, C. Serum Superoxide Dismutase Is Associated with Vascular Structure and Function in Hypertensive and Diabetic Patients. Oxid. Med. Cell. Longev. 2016, 2016, 9124676. [Google Scholar] [CrossRef]
- Wassmann, S.; Wassmann, K.; Nickenig, G. Modulation of oxidant and antioxidant enzyme expression and function in vascular cells. Hypertension 2004, 44, 381–386. [Google Scholar] [CrossRef]
- Anon. (15) (PDF) Evaluation of Antioxidant Deficit and Lipid Profile in Type—2 Diabetes Mellitus Patients. ResearchGate. Available online: https://www.researchgate.net/publication/286487072_Evaluation_of_antioxidant_deficit_and_lipid_profile_in_type_-_2_diabetes_mellitus_patients (accessed on 23 July 2020).
- Niwa, Y. Lipid peroxides and superoxide dismutase (SOD) induction in skin inflammatory diseases, and treatment with SOD preparations. Dermatologica 1989, 179 (Suppl. S1), 101–106. [Google Scholar] [CrossRef]
- Lee, R.; Margaritis, M.; Channon, K.M.; Antoniades, C. Evaluating oxidative stress in human cardiovascular disease: Methodological aspects and considerations. Curr. Med. Chem. 2012, 19, 2504–2520. [Google Scholar] [CrossRef]
- Yamaji, Y.; Nakazato, Y.; Oshima, N.; Hayashi, M.; Saruta, T. Oxidative stress induced by iron released from transferrin in low pH peritoneal dialysis solution. Nephrol. Dial. Transplant. 2004, 19, 2592–2597. [Google Scholar] [CrossRef]
- Recalcati, S.; Locati, M.; Marini, A.; Santambrogio, P.; Zaninotto, F.; De Pizzol, M.; Zammataro, L.; Girelli, D.; Cairo, G. Differential regulation of iron homeostasis during human macrophage polarized activation. Eur. J. Immunol. 2010, 40, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Bowen, K.J.; Harris, W.S.; Kris-Etherton, P.M. Omega-3 Fatty Acids and Cardiovascular Disease: Are There Benefits? Curr. Treat. Options Cardiovasc. Med. 2016, 18, 69. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.J.; Yusof, M.H.; Yaqoob, P.; Miles, E.A.; Calder, P.C. Omega-3 fatty acids and leukocyte-endothelium adhesion: Novel anti-atherosclerotic actions. Mol. Asp. Med. 2018, 64, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Granger, D.N.; Senchenkova, E. Leukocyte–Endothelial Cell Adhesion. Morgan & Claypool Life Sciences. 2010. Available online: https://www.ncbi.nlm.nih.gov/books/NBK53380/ (accessed on 23 July 2020).
- Mayer, K.; Merfels, M.; Muhly-Reinholz, M.; Gokorsch, S.; Rosseau, S.; Lohmeyer, J.; Schwarzer, N.; Krull, M.; Suttorp, N.; Grimminger, F.; et al. Omega-3 fatty acids suppress monocyte adhesion to human endothelial cells: Role of endothelial PAF generation. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H811–H818. [Google Scholar] [CrossRef]
- Nimalaratne, C.; Wu, J. Hen Egg as an Antioxidant Food Commodity: A Review. Nutrients 2015, 7, 8274–8293. [Google Scholar] [CrossRef] [PubMed]
- Tagashira, A.; Nishi, K.; Sugahara, T. Lysozyme from hen egg white ameliorates lipopolysaccharide-induced systemic inflammation in mice. Cytotechnology 2019, 71, 497–506. [Google Scholar] [CrossRef]
- Sethi, S.; Ziouzenkova, O.; Ni, H.; Wagner, D.D.; Plutzky, J.; Mayadas, T.N. Oxidized omega-3 fatty acids in fish oil inhibit leukocyte-endothelial interactions through activation of PPAR alpha. Blood 2002, 100, 1340–1346. [Google Scholar] [CrossRef]
- Chang, H.Y.; Lee, H.-N.; Kim, W.; Surh, Y.-J. Docosahexaenoic acid induces M2 macrophage polarization through peroxisome proliferator-activated receptor γ activation. Life Sci. 2015, 120, 39–47. [Google Scholar] [CrossRef]
- Brown, A.L.; Zhu, X.; Rong, S.; Shewale, S.; Seo, J.; Boudyguina, E.; Gebre, A.K.; Alexander-Miller, M.A.; Parks, J.S. Omega-3 fatty acids ameliorate atherosclerosis by favorably altering monocyte subsets and limiting monocyte recruitment to aortic lesions. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2122–2130. [Google Scholar] [CrossRef]
- Dai Perrard, X.-Y.; Lian, Z.; Bobotas, G.; Dicklin, M.R.; Maki, K.C.; Wu, H. Effects of n-3 fatty acid treatment on monocyte phenotypes in humans with hypertriglyceridemia. J. Clin. Lipidol. 2017, 11, 1361–1371. [Google Scholar] [CrossRef]
- Arnardottir, H.H.; Freysdottir, J.; Hardardottir, I. Dietary fish oil decreases the proportion of classical monocytes in blood in healthy mice but increases their proportion upon induction of inflammation. J. Nutr. 2012, 142, 803–808. [Google Scholar] [CrossRef]
- Niethammer, P.; Grabher, C.; Look, A.T.; Mitchison, T.J. A tissue-scale gradient of hydrogen peroxide mediates rapid wound detection in zebrafish. Nature 2009, 459, 996–999. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Y.; Gu, Y.; Lewis, D.F.; Wang, Y. Heme oxygenase-1 mediates up-regulation of adhesion molecule expression induced by peroxynitrite in endothelial cells. J. Soc. Gynecol. Investig. 2004, 11, 465–471. [Google Scholar] [CrossRef]
- Granger, D.N.; Senchenkova, E. Inflammation and the Microcirculation; Morgan & Claypool Life Sciences: San Rafael, CA, USA, 2010. Available online: http://www.ncbi.nlm.nih.gov/books/NBK53373/ (accessed on 23 July 2020).
- Ryu, J.-W.; Hong, K.H.; Maeng, J.H.; Kim, J.-B.; Ko, J.; Park, J.Y.; Lee, K.-U.; Hong, M.K.; Park, S.W.; Kim, Y.H.; et al. Overexpression of uncoupling protein 2 in THP1 monocytes inhibits beta2 integrin-mediated firm adhesion and transendothelial migration. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 864–870. [Google Scholar] [CrossRef]
- Friedrich, K.; Sommer, M.; Strobel, S.; Thrum, S.; Blüher, M.; Wagner, U.; Rossol, M. Perturbation of the Monocyte Compartment in Human Obesity. Front. Immunol. 2019, 10, 1874. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Feeding Mixture (n = 3) (g/100 g Total Fatty Acids) | Eggs (n = 10) (mg/100 g Egg 1) | |||
|---|---|---|---|---|
| Fatty acid | For production of regular eggs | For production of n-3 PUFA-enriched eggs | Regular eggs | n-3 PUFA-enriched eggs |
| ∑SFA | 21 ± 0.8 | 17 ± 0.3 * | 3107 ± 104 | 2192 ± 71 * |
| ∑MUFA | 27 ± 1.4 * | 37 ± 1.2 | 4091 ± 180 | 2980 ± 174 * |
| ∑n-6 PUFA | 49 ± 1.8 | 33 ± 1.5 * | 1775 ± 270 | 1397 ± 153 |
| LA | 42 ± 2.5 | 33 ± 1.2 * | 1551 ± 258 | 1302 ± 146 |
| AA | n.d. | 0.04 ± 0.01 | 197 ± 15 | 79 ± 10 * |
| ∑N-3 PUFA | 3.47 ± 0.69 * | 14 ± 1.88 | 75 ± 4 * | 403 ± 49 |
| ALA | 3.33 ± 0.70 * | 11.9 ± 1.97 | 23 ± 2.9 * | 198 ± 55 |
| EPA | n.d. | 0.63 ± 0.06 | 12 ± 2.0 * | 22 ± 3.2 |
| DHA | 0.14 ± 0.02 * | 0.93 ± 0.05 | 39 ± 3.5 * | 180 ± 14 |
| ∑n-6/n-3 PUFA | 14.07 | 2.44 * | 23.79 | 3.46 * |
| Parameter | Control Group | n-3 PUFA Group | ||
|---|---|---|---|---|
| Before | After | Before | After | |
| N (W/M) | 9 (2/7) | 11 (7/4) | ||
| Age (years) | 21 ± 2 | 21 ± 2 | ||
| SBP (mmHg) | 124 ± 12 | 122 ± 10 | 117 ± 10 | 118 ± 8 |
| DBP (mmHg) | 78 ± 9 | 75 ± 10 | 75 ± 9 | 77 ± 7 |
| MAP (mmHg) | 93 ± 9 | 91 ± 9 | 89 ± 9 | 90 ± 6 |
| HR (beats per minute) | 75 ± 16 | 68 ± 11 | 72 ± 15 | 71 ± 10 |
| erythrocytes (×1012/L) | 4.9 ± 0.4 | 5.0 ± 0.5 | 4.8 ± 0.5 | 4.8 ± 0.5 |
| hemoglobin (g/L) | 144 ± 12 | 148 ± 14 * | 139 ± 14 | 139 ± 13 |
| hematocrit (%) | 42.3 ± 3.8 | 43.9 ± 4.0 * | 41.7 ± 3.8 | 42.2 ± 3.9 |
| leukocytes (×109/L) | 7.3 ± 1.1 | 7.2 ± 1.6 | 7.5 ± 1.6 | 8.2 ± 1.3 |
| thrombocytes (×109/L) | 231 ± 36 | 258 ± 39 | 266 ± 69 | 289 ± 75 * |
| urea (mol/L) | 5.7 ± 1.4 | 5.5 ± 1.4 | 5.4 ± 1.5 | 5.4 ± 1.4 |
| creatinine (µmol/L) | 83 ± 15 | 80 ± 16 | 79 ± 13 | 79 ± 14 |
| urates (µmol/L) | 343 ± 27 | 332 ± 41 | 298 ± 76 | 291 ± 69 |
| sodium (mol/L) | 138 ± 2 | 137 ± 2 | 140 ± 2 † | 139 ± 2 |
| potassium (mol/L) | 4.1 ± 0.2 | 4.1 ± 0.3 | 4.3 ± 0.2 | 4.4 ± 0.5 |
| calcium (mol/L) | 2.4 ± 0.1 | 2.5 ± 0.1 | 2.4 ± 0.1 | 2.5 ± 0.1 |
| iron (µmol/L) | 16 ± 5 | 17 ± 6 | 18 ± 8 | 15 ± 9 |
| transferrin (g/L) | 2.5 ± 0.3 | 2.7 ± 0.3 * | 2.7 ± 0.2 | 3.0 ± 0.4 * |
| glucose (mol/L) | 5.2 ± 0.6 | 4.9 ± 0.7 | 4.9 ± 0.4 | 4.4 ± 0.4 * |
| hsCRP (mg/L) | 1.7 ± 2.8 | 0.6 ± 0.4 | 1.6 ± 2.3 | 1.2 ± 1.0 |
| cholesterol (mol/L) | 4.0 ± 0.7 | 4.2 ± 0.3 | 4.5 ± 0.8 | 4.6 ± 0.8 |
| triglycerides (mol/L) | 1.1 ± 0.3 † | 1.1 ± 0.6 | 0.7 ± 0.3 | 0.9 ± 0.5 |
| HDL cholesterol (mol/L) | 1.4 ± 0.2 | 1.4 ± 0.2 | 1.7 ± 0.4 † | 1.7 ± 0.4 |
| LDL cholesterol (mol/L) | 2.1 ± 0.5 | 2.3 ± 0.2 | 2.3 ± 0.4 | 2.4 ± 0.5 |
| apoA (g/L) | 1.6 ± 0.2 | 1.6 ± 0.2 | 1.8 ± 0.3 | 1.7 ± 0.3 |
| apoB (g/L) | 0.6 ± 0.1 | 0.7 ± 0.1 | 0.7 ± 0.1 | 0.7 ± 0.1 |
| Parameter | Control Group | n-3 PUFAs Group | ||
|---|---|---|---|---|
| Before | After | Before | After | |
| BMI (kg/m2) | 24.4 ± 2.3 | 24.3 ± 2.3 | 25.2 ± 3.7 | 25.0 ± 4.2 |
| WHR | 0.80 ± 0.02 | 0.80 ± 0.02 | 0.79 ± 0.04 | 0.77 ± 0.04 |
| Fat Free Mass (%) | 83.6 ± 7.7 * | 83.0 ± 7.9 | 75.1 ± 7.6 | 75.3 ± 8.8 |
| Fat (%) | 16.4 ± 7.5 | 17.0 ± 7.9 | 24.9 ± 7.6 * | 24.7 ± 8.8 |
| Total Body Water (%) | 62.9 ± 8.7 | 61.6 ± 7.5 | 56.0 ± 7.0 | 55.8 ± 7.6 |
| Extracellular Water (%) | 43.8 ± 2.8 | 43.0 ± 1.4 | 42.7 ± 1.5 | 42.8 ± 1.8 |
| Intracellular Water (%) | 56.2 ± 2.8 | 57.0 ± 1.4 | 57.3 ± 1.5 | 57.2 ± 1.8 |
| Plasma Fluid (L) | 4.61 ± 0.98 | 4.44 ± 1.07 | 3.79 ± 0.98 | 3.79 ± 1.22 |
| Body Density (kg/L) | 1.061 ± 0.015 * | 1.060 ± 0.020 | 1.043 ± 0.017 | 1.043 ± 0.020 |
| Parameter | Control Group (n = 9) | Omega-3 Group (n = 11) | ||
|---|---|---|---|---|
| Before | After | Before | After | |
| %SFA (%FAME) | 36.7 ± 2.2 | 33.9 ± 10.2 | 36.9 ± 1.8 | 29.3 ± 11.7 |
| %MUFA (%FAME) | 23.2 ± 5.1 | 24.7 ± 10.0 | 19.4 ± 2.1 | 28.1 ± 14.9 |
| %PUFA (%FAME) | 40.6 ± 5.5 | 41.3 ± 5.5 | 43.6 ± 2.8 | 42.6 ± 4.8 |
| SFA (%FAME) | ||||
| C14:0 Myristic acid | 0.82 ± 0.74 | 1.31 ± 0.76 | 1.02 ± 0.34 | 1.30 ± 0.57 |
| C16:0 Palmitic acid | 25.8 ± 2.9 | 25.5 ± 3.3 | 26.6 ± 1.6 | 27.0 ± 2.5 |
| C18:0 Stearic acid | 10.1 ± 2.0 | 10.2 ± 3.8 | 9.5 ± 1.1 | 9.3 ± 1.2 |
| MUFA and PUFA (%FAME) | ||||
| C16:1 (n-7) Palmitoleic acid | 1.21 ± 0.76 | 1.49 ± 0.55 | 1.24 ± 0.28 | 1.40 ± 0.65 |
| C18:1 (n-7) Vaccenic acid | 1.24 ± 0.72 | 1.67 ± 0.31 | 1.29 ± 0.18 | 1.38 ± 0.24 |
| C18:1 (n-9) Oleic acid | 20.7 ± 4.6 †* | 18.4 ± 2.4 | 16.6 ± 2.1 | 16.9 ± 2.8 |
| C18:2 (n-6) Linoleic acid | 30.3 ± 4.9 | 30.4 ± 4.3 | 34.0 ± 2.9 | 32.8 ± 4.1 |
| C20:3 (n-6) Dihomo-gamma-linolenic acid | 1.46 ± 0.73 | 1.51 ± 0.73 | 1.74 ± 0.91 | 1.69 ± 0.73 |
| C20:4 (n-6) Arachidonic acid | 7.0 ± 2.4 | 7.7 ± 1.6 | 6.8 ± 1.5 | 6.8 ± 1.6 |
| C18:3 (n-3) α-Linolenic acid | 1.04 ± 1.86 * | 0.61 ± 1.1.10 | 0 ± 0 | 0.08 ± 0.18 |
| C20:5 (n-3) Eicosapentaenoic acid | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| C22:6 (n-3) Docosahexaenoic acid | 0.85 ± 0.86 | 1.14 ± 0.26 | 0.91 ± 0.48 | 1.09 ± 0.66 |
| n-6/n-3 PUFA | 20.5 | 22.6 | 46.7 | 35.3 |
| Parameter | Control Group | n-3 PUFAs Group | ||
|---|---|---|---|---|
| Before | After | Before | After | |
| TBARS (µM MDA) | 23.5 ± 0.6 | 24.1 ± 2.1 | 26.7 ± 2.6 * | 26.1 ± 2.8 * |
| FRAP (mM Trolox) | 0.47 ± 0.06 | 0.48 ± 0.05 | 0.46 ± 0.07 | 0.42 ± 0.08 † |
| GPx (U/mg protein) | 3.83 × 10−3 ± 0.7 × 10−3 | 3.45 × 10−3 ± 0.5 × 10−3 | 3.59 × 10−3 ± 0.4 × 10−3 | 4.04 × 10−3 ± 0.9 × 10−3 |
| SOD (U/mg protein) | 0.11 ± 0.05 | 0.10 ± 0.02 | 0.06 ± 0.06 | 0.10 ± 0.10 † |
| LOOH (µM) | 1.46 ± 0.66 | 1.35 ± 0.51 | 1.30 ± 0.65 | 1.10 ± 0.47 |
| Control Group | n-3 PUFAs Group | |||
|---|---|---|---|---|
| Body Mass Index (kg/m2) | ||||
| r = | p = | r = | p = | |
| Rate of peripheral blood monocytes (%) | −0.684 | 0.010 * | −0.468 | 0.039 * |
| FRAP (mM Trolox) | 0.092 | 0.717 | 0.534 | 0.015 * |
| TBARS (µM MDA) | 0.113 | 0.654 | 0.462 | 0.039 * |
| SOD activity (U/mg protein) | −0.052 | 0.839 | −0.548 | 0.028 * |
| GPx activity (U/mg protein) | −0.066 | 0.793 | −0.376 | 0.146 |
| LOOH (µM) | −0.252 | 0.313 | −0.098 | 0.665 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mihalj, M.; Stupin, A.; Kolobarić, N.; Tartaro Bujak, I.; Matić, A.; Kralik, Z.; Jukić, I.; Stupin, M.; Drenjančević, I. Leukocyte Activation and Antioxidative Defense Are Interrelated and Moderately Modified by n-3 Polyunsaturated Fatty Acid-Enriched Eggs Consumption—Double-Blind Controlled Randomized Clinical Study. Nutrients 2020, 12, 3122. https://doi.org/10.3390/nu12103122
Mihalj M, Stupin A, Kolobarić N, Tartaro Bujak I, Matić A, Kralik Z, Jukić I, Stupin M, Drenjančević I. Leukocyte Activation and Antioxidative Defense Are Interrelated and Moderately Modified by n-3 Polyunsaturated Fatty Acid-Enriched Eggs Consumption—Double-Blind Controlled Randomized Clinical Study. Nutrients. 2020; 12(10):3122. https://doi.org/10.3390/nu12103122
Chicago/Turabian StyleMihalj, Martina, Ana Stupin, Nikolina Kolobarić, Ivana Tartaro Bujak, Anita Matić, Zlata Kralik, Ivana Jukić, Marko Stupin, and Ines Drenjančević. 2020. "Leukocyte Activation and Antioxidative Defense Are Interrelated and Moderately Modified by n-3 Polyunsaturated Fatty Acid-Enriched Eggs Consumption—Double-Blind Controlled Randomized Clinical Study" Nutrients 12, no. 10: 3122. https://doi.org/10.3390/nu12103122
APA StyleMihalj, M., Stupin, A., Kolobarić, N., Tartaro Bujak, I., Matić, A., Kralik, Z., Jukić, I., Stupin, M., & Drenjančević, I. (2020). Leukocyte Activation and Antioxidative Defense Are Interrelated and Moderately Modified by n-3 Polyunsaturated Fatty Acid-Enriched Eggs Consumption—Double-Blind Controlled Randomized Clinical Study. Nutrients, 12(10), 3122. https://doi.org/10.3390/nu12103122