Diet Quality, Food Groups and Nutrients Associated with the Gut Microbiota in a Nonwestern Population

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population and Design
2.2. Ethics
2.3. Dietary Data
2.4. Gut Microbiota Data
2.5. Statistical Analysis
2.6. Data Availability
3. Results
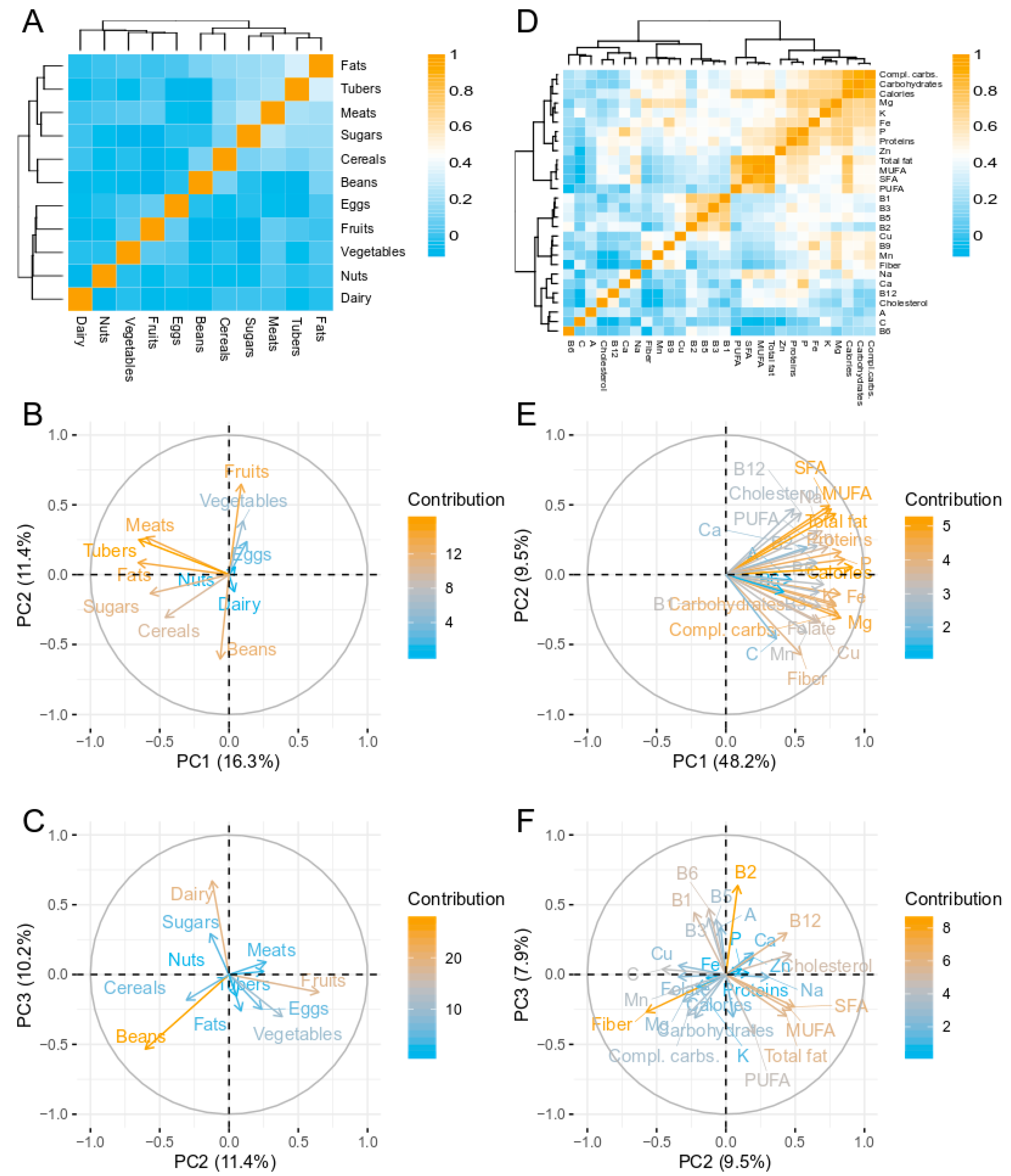
3.1. Dietary Analysis
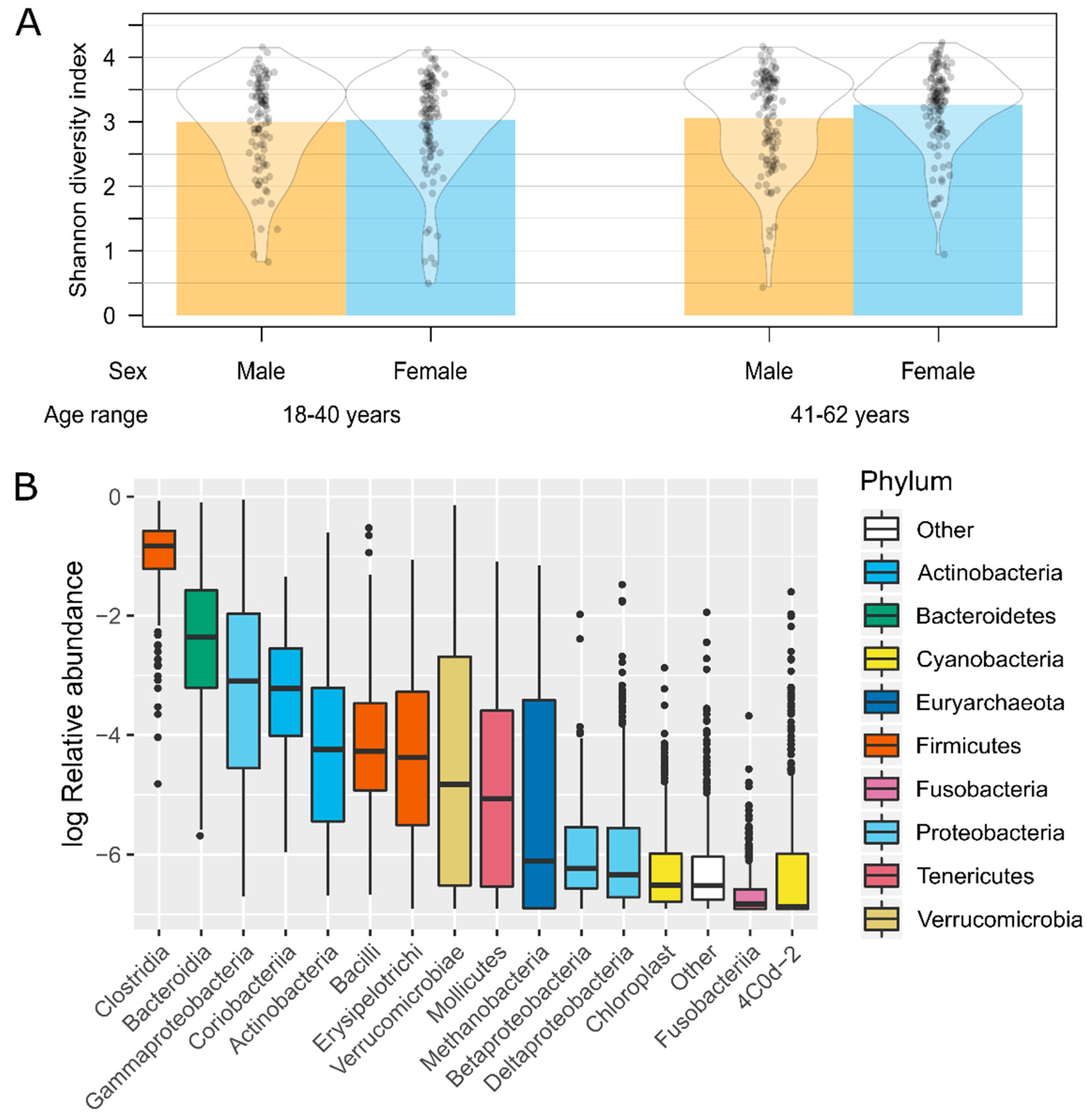
3.2. Gut Microbiota Analysis
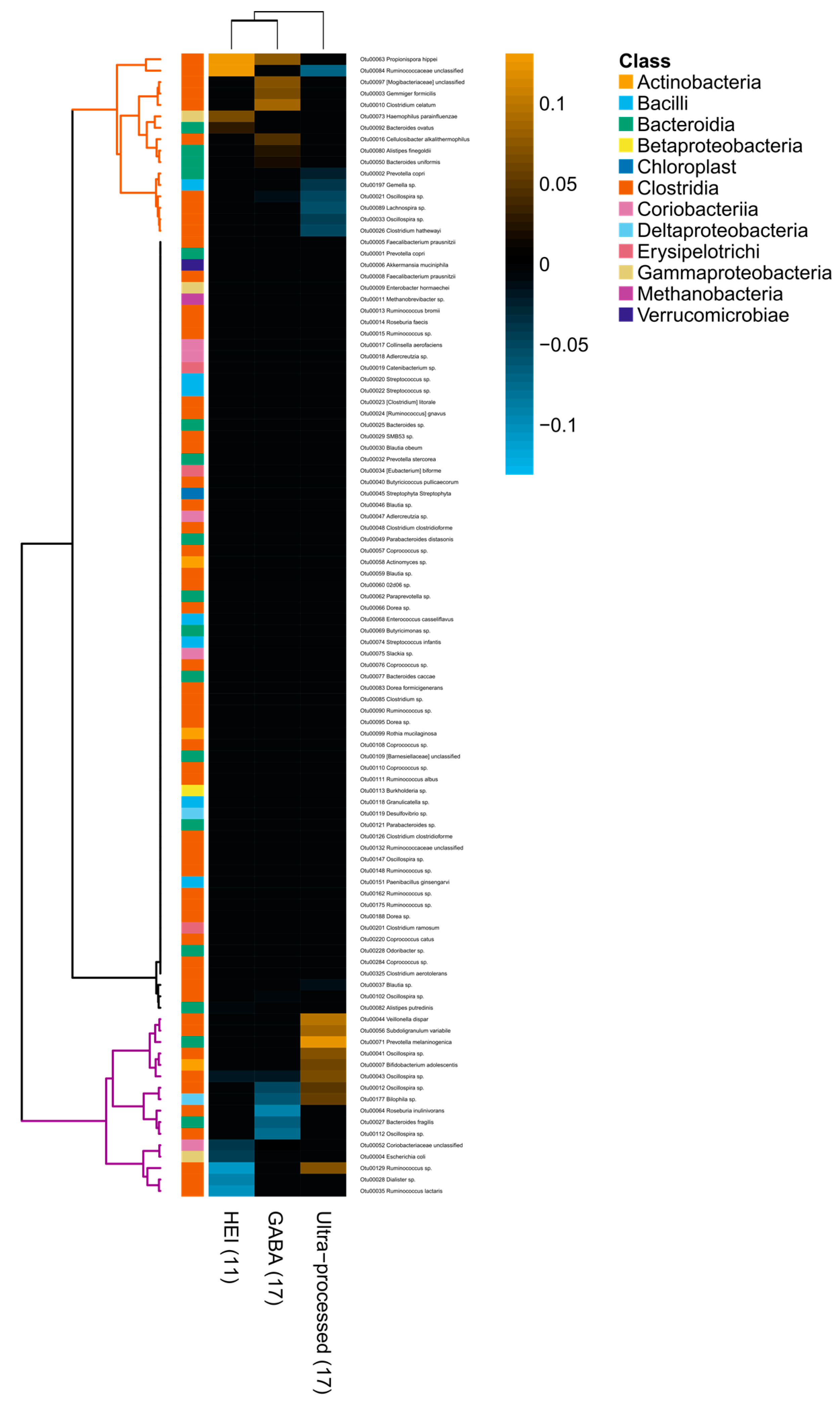
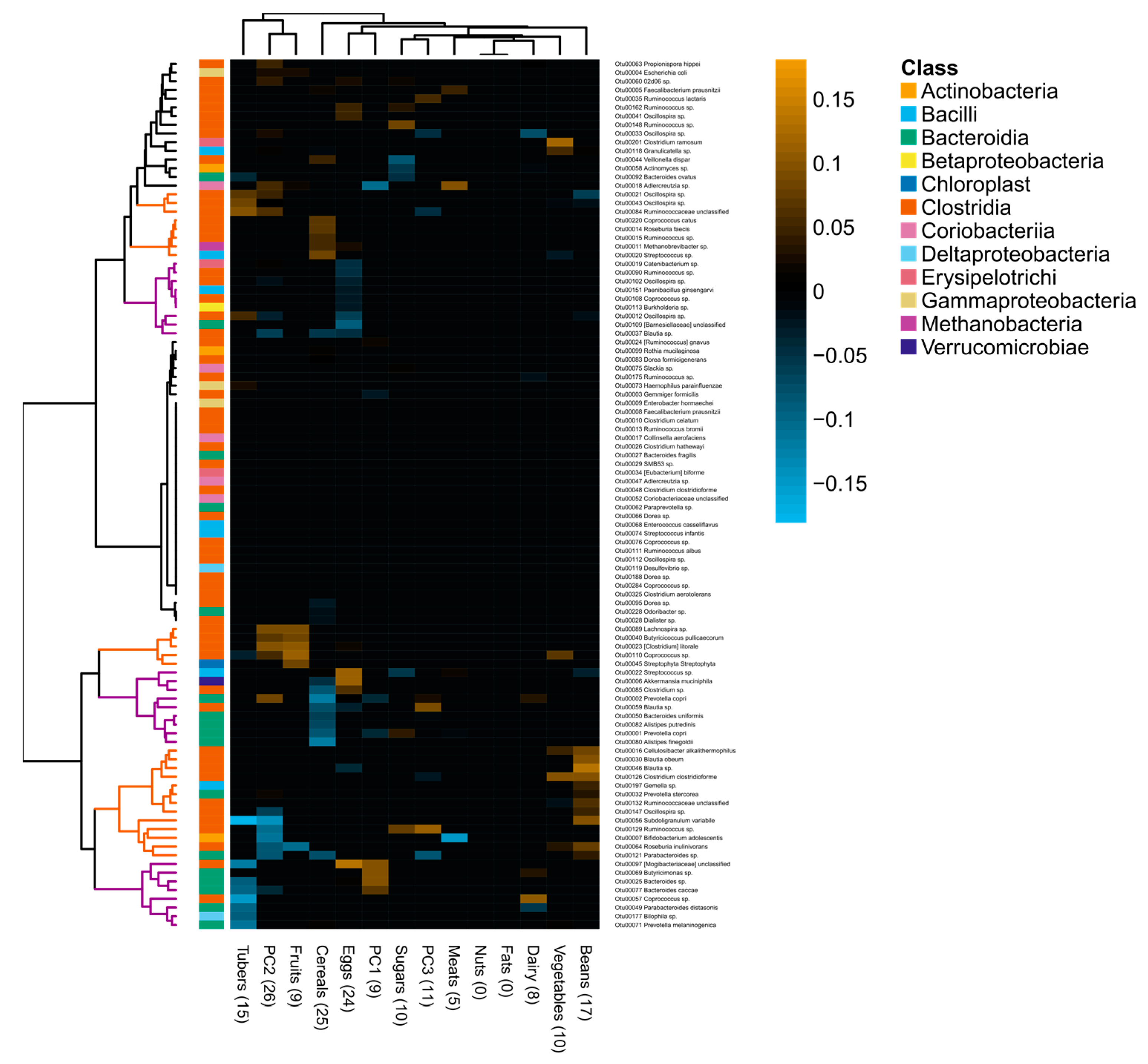
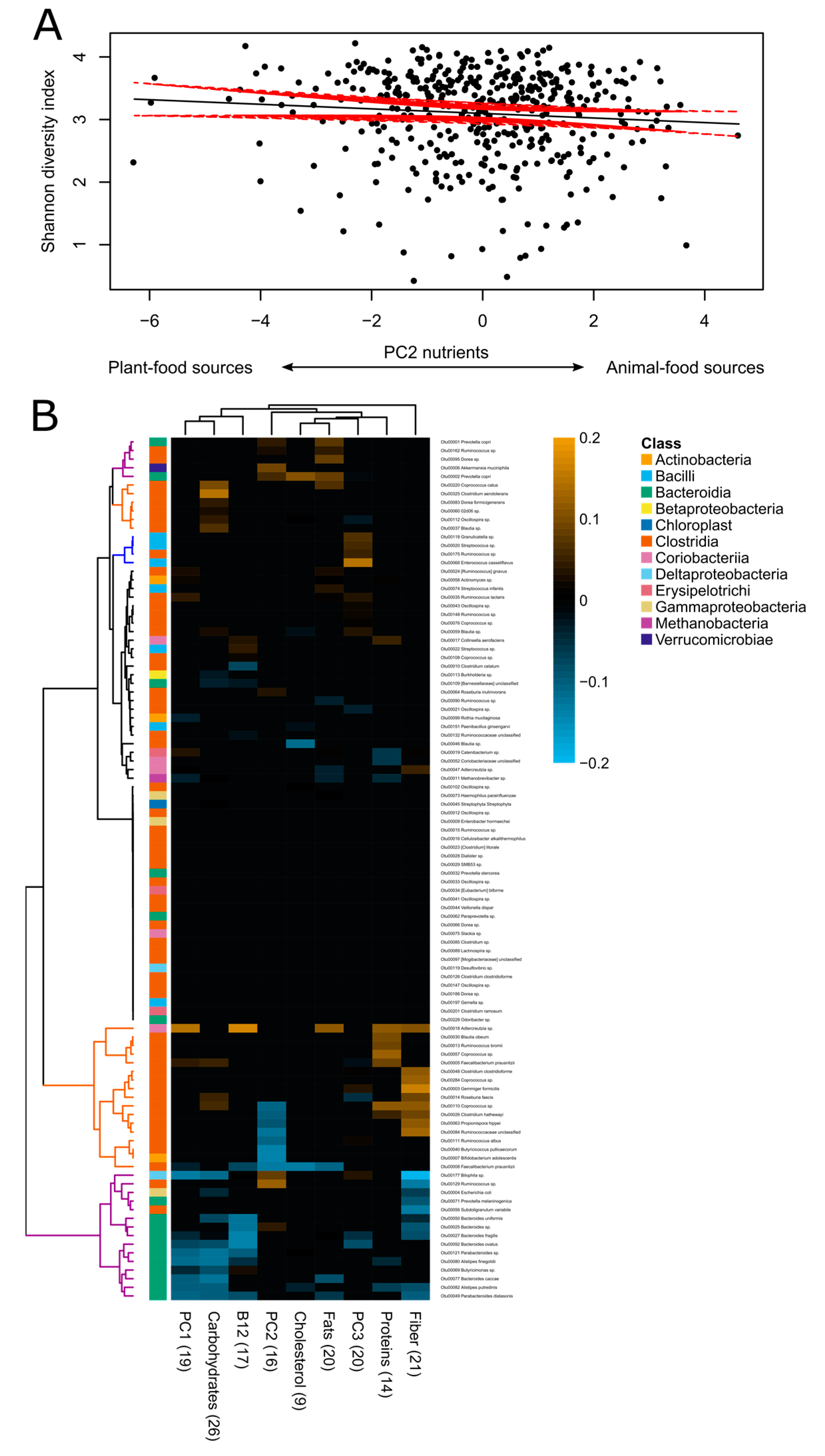
3.3. Associations between Diet and Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef]
- Huttenhower, C.; Gevers, D.; Knight, R.; Abubucker, S.; Badger, J.H.; Chinwalla, A.T.; Creasy, H.H.; Earl, A.M.; Fitzgerald, M.G.; Fulton, R.S.; et al. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef]
- Mancabelli, L.; Milani, C.; Lugli, G.A.; Turroni, F.; Ferrario, C.; van Sinderen, D.; Ventura, M. Meta-analysis of the human gut microbiome from urbanized and pre-agricultural populations. Environ. Microbiol. 2017, 19, 1379–1390. [Google Scholar] [CrossRef]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef]
- De la Cuesta-Zuluaga, J.; Kelley, S.T.; Chen, Y.; Escobar, J.S.; Mueller, N.T.; Ley, R.E.; McDonald, D.; Huang, S.; Swafford, A.D.; Knight, R.; et al. Age- and sex-dependent patterns of gut microbial diversity in human adults. Msystems 2019, 4, 1–12. [Google Scholar] [CrossRef]
- Kau, A.L.; Ahern, P.P.; Griffin, N.W.; Goodman, A.L.; Gordon, J.I. Human nutrition, the gut microbiome and the immune system. Nature 2011, 474, 327–336. [Google Scholar] [CrossRef]
- Albenberg, L.G.; Wu, G.D. Diet and the intestinal microbiome: Associations, functions, and implications for health and disease. Gastroenterology 2014, 146, 1564–1572. [Google Scholar] [CrossRef]
- Annalisa, N.; Alessio, T.; Claudette, T.D.; Erald, V.; Antonino, D.L.; Nicola, D.D. Gut microbioma population: An indicator really sensible to any change in age, diet, metabolic syndrome, and life-style. Mediat. Inflamm. 2014, 2014, 1–11. [Google Scholar] [CrossRef]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef]
- Muegge, B.D.; Kuczynski, J.; Knights, D.; Clemente, J.C.; González, A.; Fontana, L.; Henrissat, B.; Knight, R.; Gordon, J.I. Diet drives convergence in gut microbiome functions across mammalian phylogeny and within humans. Science 2011, 332, 970–974. [Google Scholar] [CrossRef]
- Gomez, A.; Petrzelkova, K.J.; Burns, M.B.; Yeoman, C.J.; Amato, K.R.; Vlckova, K.; Modry, D.; Todd, A.; Jost Robinson, C.A.; Remis, M.J.; et al. Gut microbiome of coexisting BaAka pygmies and bantu reflects gradients of traditional subsistence patterns. Cell Rep. 2016, 14, 2142–2153. [Google Scholar] [CrossRef]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef]
- Schnorr, S.L.; Candela, M.; Rampelli, S.; Centanni, M.; Consolandi, C.; Basaglia, G.; Turroni, S.; Biagi, E.; Peano, C.; Severgnini, M.; et al. Gut microbiome of the Hadza hunter-gatherers. Nat. Commun. 2014, 5, 1–12. [Google Scholar] [CrossRef]
- Clemente, J.C.; Pehrsson, E.C.; Blaser, M.J.; Sandhu, K.; Gao, Z.; Wang, B.; Magris, M.; Hidalgo, G.; Contreras, M.; Noya-Alarcón, Ó.; et al. The microbiome of uncontacted Amerindians. Sci. Adv. 2015, 1, e1500183. [Google Scholar] [CrossRef]
- Obregon-Tito, A.J.; Tito, R.Y.; Metcalf, J.; Sankaranarayanan, K.; Clemente, J.C.; Ursell, L.K.; Zech Xu, Z.; Van Treuren, W.; Knight, R.; Gaffney, P.M.; et al. Subsistence strategies in traditional societies distinguish gut microbiomes. Nat. Commun. 2015, 6, 1–9. [Google Scholar] [CrossRef]
- Sonnenburg, E.D.; Smits, S.A.; Tikhonov, M.; Higginbottom, S.K.; Wingreen, N.S.; Sonnenburg, J.L. Diet-induced extinctions in the gut microbiota compound over generations. Nature 2016, 529, 212–215. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- O’Keefe, S.J.D.; Li, J.V.; Lahti, L.; Ou, J.; Carbonero, F.; Mohammed, K.; Posma, J.M.; Kinross, J.; Wahl, E.; Ruder, E.; et al. Fat, fibre and cancer risk in African Americans and rural Africans. Nat. Commun. 2015, 6, 1–14. [Google Scholar] [CrossRef]
- Liu, Y.; Ajami, N.J.; El-Serag, H.B.; Hair, C.; Graham, D.Y.; White, D.L.; Chen, L.; Wang, Z.; Plew, S.; Kramer, J.; et al. Dietary quality and the colonic mucosa-associated gut microbiome in humans. Am. J. Clin. Nutr. 2019, 110, 701–712. [Google Scholar] [CrossRef]
- Meslier, V.; Laiola, M.; Munch, H.; De Filippis, F.; Roume, H.; Quinquis, B.; Giacco, R.; Mennella, I.; Ferracane, R.; Pons, N.; et al. Gut microbiota Mediterranean diet intervention in overweight and obese subjects lowers plasma cholesterol and causes changes in the gut microbiome and metabolome independently of energy intake. Gut 2020, 69, 1258–1268. [Google Scholar] [CrossRef]
- Claesson, M.J.; Cusack, S.; O’Sullivan, O.; Greene-Diniz, R.; De Weerd, H.; Flannery, E.; Marchesi, J.R.; Falush, D.; Dinan, T.; Fitzgerald, G.; et al. Composition, variability, and temporal stability of the intestinal microbiota of the elderly. Proc. Natl. Acad. Sci. USA 2011, 108, 4586–4591. [Google Scholar] [CrossRef]
- Profamilia; Instituto Nacional de Salud; Universidad de Antioquia; Organización Panamericana de la Salud; Instituto Colombiano de Bienestar Familiar (ICBF). Encuesta Nacional De La Situación Nutricional En Colombia (ENSIN) 2005; Instituto Colombiano de Bienestar Familiar: Bogotá D.C., Colombia, 2006. [Google Scholar]
- Manjarrés, L.M.; de Universidad, A.; Manjarrés, S. Programa de Evaluación de Ingesta Dietética EVINDI v4; Universidad de Antioquia: Medellín, Colombia, 2008. [Google Scholar]
- Instituto Colombiano de Bienestar Familiar (ICBF). Tabla de Composición de Alimentos Colombianos. Reimpresión Año 2000; División de Recursos Materiales y Físicos ICBF: Bogotá D.C., Colombia, 2000. [Google Scholar]
- Quintero, D.; Escobar, L. Tabla de Composición de Alimentos; Centro de atención Nutricional: Medellín, Colombia, 1996. [Google Scholar]
- Krebs-Smith, S.M.; Pannucci, T.R.E.; Subar, A.F.; Kirkpatrick, S.I.; Lerman, J.L.; Tooze, J.A.; Wilson, M.M.; Reedy, J. Update of the healthy eating index: HEI-2015. J. Acad. Nutr. Diet. 2018, 118, 1591–1602. [Google Scholar] [CrossRef] [PubMed]
- Instituto Colombiano de Bienestar Familiar (ICBF). Documento técnico. Guías Alimentarias Basadas en Alimentos Para la Población Colombiana Mayor de 2 Años; ICBF: Bogotá D.C., Colombia, 2016. [Google Scholar]
- Monteiro, C.A.; Cannon, G.; Moubarac, J.C.; Levy, R.B.; Louzada, M.L.C.; Jaime, P.C. The UN decade of nutrition, the NOVA food classification and the trouble with ultra-processing. Public Health Nutr. 2018, 21, 5–17. [Google Scholar] [CrossRef]
- Iowa. Iowa State University Dietary Assessment Research Series Report 8; Iowa, Department of Statistics, PC-SIDE: Ames, IA, USA, 1996. [Google Scholar]
- De la Cuesta-Zuluaga, J.; Mueller, N.T.; Corrales-Agudelo, V.; Velásquez-Mejía, E.P.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Metformin is associated with higher relative abundance of mucin-degrading Akkermansia muciniphila and several short-chain fatty acid-producing microbiota in the gut. Diabetes Care 2017, 40, 54–62. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the Miseq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef]
- Evans, J.; Sheneman, L.; Foster, J. Relaxed neighbor joining: A fast distance-based phylogenetic tree construction method. J. Mol. Evol. 2006, 62, 785–792. [Google Scholar] [CrossRef]
- Kindt, R.; Coe, R. Tree Diversity Analysis. A Manual and Software for Common Statistical Methods for Ecological and Biodiversity Studies; World Agroforestry Centre (ICRAF): Nairobi, Kenya, 2005; ISBN 92-9059-179-X. [Google Scholar]
- Chen, J.; Bittinger, K.; Charlson, E.S.; Hoffmann, C.; Lewis, J.; Wu, G.D.; Collman, R.G.; Bushman, F.D.; Li, H. Associating microbiome composition with environmental covariates using generalized UniFrac distances. Bioinformatics 2012, 28, 2106–2113. [Google Scholar] [CrossRef]
- De la Cuesta-Zuluaga, J.; Corrales-Agudelo, V.; Velásquez-Mejía, E.P.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Gut microbiota is associated with obesity and cardiometabolic disease in a population in the midst of Westernization. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- De la Cuesta-Zuluaga, J.; Corrales-Agudelo, V.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Body size phenotypes comprehensively assess cardiometabolic risk and refine the association between obesity and gut microbiota. Int. J. Obes. 2018, 42, 424–432. [Google Scholar] [CrossRef]
- Zackular, J.P.; Baxter, N.T.; Chen, G.Y.; Schloss, P.D. Manipulation of the gut microbiota reveals role in colon tumorigenesis. Msphere 2015, 1. [Google Scholar] [CrossRef] [PubMed]
- Kovalskys, I.; Fisberg, M.; Gómez, G.; Pareja, R.G.; Yépez García, M.C.; Cortés Sanabria, L.Y.; Herrera-Cuenca, M.; Rigotti, A.; Guajardo, V.; Zalcman Zimberg, I.; et al. Energy intake and food sources of eight Latin American countries: Results from the Latin American Study of Nutrition and Health (ELANS). Public Health Nutr. 2018, 21, 2535–2547. [Google Scholar] [CrossRef]
- Ruiz Pardo, Y.A. Caracterización de la Calidad Nutricional a Partir del Índice de Alimentación Saludable (HEI). Población de 18 a 59 Años 11 Meses; Estudio Latinoamericano de Nutrición y Salud Colombia ELANS-C, Pontificia Universidad Javeriana: Bogotá, Colombia, 2019. [Google Scholar]
- Agudelo Cañas, S.; Hernandez, T.B.C. Recomendaciones de Ingesta de Energía y Nutrientes (RIEN) Para la Población Colombiana. Documento Técnico; Subdirección de Salud Nutricional Alimentos y Bebidas, Ministerio de Salud y Protección Social: Bogotá, Colombia, 2016. [Google Scholar]
- Claesson, M.J.; Jeffery, I.B.; Conde, S.; Power, S.E.; O’Connor, E.M.; Cusack, S.; Harris, H.M.B.; Coakley, M.; Lakshminarayanan, B.; O’Sullivan, O.; et al. Gut microbiota composition correlates with diet and health in the elderly. Nature 2012, 488, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Cotillard, A.; Kennedy, S.P.; Kong, L.C.; Prifti, E.; Pons, N.; Le Chatelier, E.; Almeida, M.; Quinquis, B.; Levenez, F.; Galleron, N.; et al. Dietary intervention impact on gut microbial gene richness. Nature 2013, 500, 585–588. [Google Scholar] [CrossRef]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humarán, L.G.; Gratadoux, J.J.; Blugeon, S.; Bridonneau, C.; Furet, J.P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef] [PubMed]
- Roediger, W.E.W. Utilization of nutrients by isolated epithelial cells of the rat colon. Gastroenterology 1982, 83, 424–429. [Google Scholar] [CrossRef]
- Vital, M.; Karch, A.; Pieper, D.H. Colonic butyrate-producing communities in humans: An overview using omics data. Msystems 2017, 2. [Google Scholar] [CrossRef]
- Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless, A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk between microbiota-derived short-chain fatty acids and intestinal epithelial HIF augments tissue barrier function. Cell Host Microbe 2015, 17, 662–671. [Google Scholar] [CrossRef]
- Vieira, A.T.; MacIa, L.; Galvão, I.; Martins, F.S.; Canesso, M.C.C.; Amaral, F.A.; Garcia, C.C.; Maslowski, K.M.; De Leon, E.; Shim, D.; et al. A role for gut microbiota and the metabolite-sensing receptor GPR43 in a murine model of gout. Arthritis Rheumatol. 2015, 67, 1646–1656. [Google Scholar] [CrossRef] [PubMed]
- Devkota, S.; Wang, Y.; Musch, M.W.; Leone, V.; Fehlner-Peach, H.; Nadimpalli, A.; Antonopoulos, D.A.; Jabri, B.; Chang, E.B. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10-/- mice. Nature 2012, 487, 104–108. [Google Scholar] [CrossRef]
- Natividad, J.M.; Lamas, B.; Pham, H.P.; Michel, M.L.; Rainteau, D.; Bridonneau, C.; Da Costa, G.; Van Hylckama Vlieg, J.; Sovran, B.; Chamignon, C.; et al. Bilophila wadsworthia aggravates high fat diet induced metabolic dysfunctions in mice. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Litvak, Y.; Byndloss, M.X.; Tsolis, R.M.; Bäumler, A.J. Dysbiotic Proteobacteria expansion: A microbial signature of epithelial dysfunction. Curr. Opin. Microbiol. 2017, 39, 1–6. [Google Scholar] [CrossRef]
- Rivas, L.; Mellor, G.E.; Gobius, K.; Fegan, N. Introduction to pathogenic Escherichia coli. In Detection and Typing Strategies for Pathogenic Escherichia Coli; Springer: Berlin, Germany, 2015; pp. 1–38. [Google Scholar]
- Brennan, C.A.; Garrett, W.S. Gut microbiota, inflammation, and colorectal cancer. Annu. Rev. Microbiol. 2016, 70, 395–411. [Google Scholar] [CrossRef]
- Zhang, G.; Weintraub, A. Prevalence of enterotoxigenic Bacteroides fragilis in adult patients with diarrhea and healthy controls. Clin. Infect. Dis. 1999, 29, 590–594. [Google Scholar] [CrossRef]
- Brook, I. Anaerobic Infections. Diagnosis and Management, 1st ed.; CRC Press: Boca Raton, FL, USA, 2007; ISBN 978-0849382574. [Google Scholar]
- Instituto Colombiano de Bienestar Familiar (ICBF); Ministerio de Salud; Instituto Nacional de Salud; Universidad Nacional de Colombia; Fundación Santa Fe de Bogotá. Encuesta Nacional de la Situación Nutricional de Colombia (ENSIN) 2015; Instituto Colombiano de Bienestar Familiar: Bogotá D.C., Colombia, 2015. [Google Scholar]
- Food and Agriculture Organization (FAO). Panorama de la Seguridad Alimentaria y Nutricional en América Latina y el Caribe 2017; Food and Agriculture Organization: Santiago, Chile, 2017. [Google Scholar]
- Food and Agriculture Organization (FAO). Panorama de la Seguridad Alimentaria y Nutricional en América Latina y el Caribe 2019: Hacia Entornos Alimentarios Más Saludables Que Hagan Frente a Todas Las Formas de Malnutrición; Food and Agriculture Organization: Santiago, Chile, 2019. [Google Scholar]
- Kovalskys, I.; Rigotti, A.; Koletzko, B.; Fisberg, M.; Gómez, G.; Herrera-Cuenca, M.; Cortés Sanabria, L.Y.; Yépez García, M.C.; Pareja, R.G.; Zimberg, I.Z.; et al. Latin American consumption of major food groups: Results from the ELANS study. PLoS ONE 2019, 14, e0225101. [Google Scholar] [CrossRef]
- Sonnenburg, E.D.; Sonnenburg, J.L. The ancestral and industrialized gut microbiota and implications for human health. Nat. Rev. Microbiol. 2019, 17, 383–390. [Google Scholar] [CrossRef]
- Duvallet, C.; Gibbons, S.M.; Gurry, T.; Irizarry, R.A.; Alm, E.J. Meta-analysis of gut microbiome studies identifies disease-specific and shared responses. Nat. Commun. 2017, 8, 1–10. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Global Status Report on Noncommunicable Diseases 2014; WHO Press: Geneva, Switzerland, 2014; ISBN 978-92-4-156485-4. [Google Scholar]
- Guzmán-Castañeda, S.J.; Ortega-Vega, E.L.; de la Cuesta-Zuluaga, J.; Velásquez-Mejía, E.P.; Rojas, W.; Bedoya, G.; Escobar, J.S. Gut microbiota composition explains more variance in the host cardiometabolic risk than genetic ancestry. Gut Microbes 2019, 11, 191–204. [Google Scholar] [CrossRef]
- Escobar, J.S.; Klotz, B.; Valdes, B.E.; Agudelo, G.M. The gut microbiota of Colombians differs from that of Americans, Europeans and Asians. BMC Microbiol. 2014, 14, 1–14. [Google Scholar] [CrossRef]
- Carpenter, C.L. Dietary Assessment. In Nutritional Oncology, 2nd ed.; Academic Press: Boston, MA, USA, 2006; ISBN 9780120883936. [Google Scholar]
- Johnson, A.J.; Vangay, P.; Al-Ghalith, G.A.; Hillmann, B.M.; Ward, T.L.; Shields-Cutler, R.R.; Kim, A.D.; Shmagel, A.K.; Syed, A.N.; Walter, J.; et al. Daily sampling reveals personalized diet-microbiome associations in humans. Cell Host Microbe 2019, 25, 789–802.e5. [Google Scholar] [CrossRef]
- Zeevi, D.; Korem, T.; Zmora, N.; Israeli, D.; Rothschild, D.; Weinberger, A.; Ben-Yacov, O.; Lador, D.; Avnit-Sagi, T.; Lotan-Pompan, M.; et al. Personalized nutrition by prediction of glycemic responses. Cell 2015, 163, 1079–1094. [Google Scholar] [CrossRef]
- Korem, T.; Zeevi, D.; Zmora, N.; Weissbrod, O.; Bar, N.; Lotan-Pompan, M.; Avnit-Sagi, T.; Kosower, N.; Malka, G.; Rein, M.; et al. Bread affects clinical parameters and induces gut microbiome-associated personal glycemic responses. Cell Metab. 2017, 25. [Google Scholar] [CrossRef]
- Bashiardes, S.; Godneva, A.; Elinav, E.; Segal, E. Towards utilization of the human genome and microbiome for personalized nutrition. Curr. Opin. Biotechnol. 2018, 51, 57–63. [Google Scholar] [CrossRef]
- Zmora, N.; Suez, J.; Elinav, E. You are what you eat: Diet, health and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 35–56. [Google Scholar] [CrossRef]
- Spector, M.P. Metabolism, central (intermediary). Encycl. Microbiol. 2009, 728–752. [Google Scholar] [CrossRef]
- Faust, K.; Sathirapongsasuti, J.F.; Izard, J.; Segata, N.; Gevers, D.; Raes, J.; Huttenhower, C. Microbial co-occurrence relationships in the human microbiome. PLoS Comput. Biol. 2012, 8, e1002606. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.; Gasbarrini, A.; Mele, M. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Males | Females | ||||
|---|---|---|---|---|---|
| 18–40 Years | 41–62 Years | 18–40 Years | 41–62 Years | ||
| (n = 98) | (n = 114) | (n = 109) | (n = 120) | p-Value 1 | |
| Diet quality | |||||
| HEI 2 | 37.3 (7.44) | 40.4 (8.80) | 39.1 (9.64) | 41.7 (9.18) | /*** |
| GABA 3 | 22.3 (10.0) | 27.6 (10.6) | 24.1 (10.7) | 28.6 (10.0) | NS/*** |
| Ultraprocessed foods (%) 4 | 34.7 (16.2) | 30.9 (15.8) | 39.0 (18.6) | 34.7 (15.6) | */** |
| Food groups | |||||
| Dairy (g) | 172 (196) | 183 (196) | 186 (240) | 201 (168) | NS/NS |
| Meats (g) | 170 (122) | 136 (94.3) | 113 (81.7) | 86.5 (60.8) | ***/*** |
| Eggs (g) | 40.8 (59.9) | 39.0 (43.7) | 38.2 (45.0) | 32.9 (47.1) | NS/NS |
| Beans (g) | 78.4 (185) | 38.0 (94.4) | 29.1 (63.1) | 29.9 (69.3) | */. |
| Nuts (g) | 1.53 (7.85) | 2.91 (12.4) | 2.37 (9.45) | 3.51 (26.8) | NS/NS |
| Fruits (g) | 200 (238) | 232 (243) | 171 (197) | 221 (257) | NS/. |
| Vegetables (g) | 72.9 (80.8) | 97.6 (133) | 74.2 (75.3) | 105 (106) | NS/** |
| Cereals (g) | 350 (164) | 333 (201) | 230 (152) | 203 (137) | ***/NS |
| Tubers (g) | 220 (208) | 174 (180) | 136 (147) | 90.7 (113) | ***/** |
| Fats (g) | 33.6 (29.7) | 25.6 (29.9) | 22.3 (26.7) | 14.2 (18.2) | ***/** |
| Sugars (g) | 339 (348) | 213 (239) | 178 (203) | 141 (202) | ***/*** |
| Nutrients | |||||
| Calories (kcal) | 2240 (389) | 2030 (487) | 1830 (347) | 1670 (316) | ***/*** |
| Macronutrients | |||||
| Carbohydrates (g) | 305 (53.4) | 286 (76.5) | 248 (44.9) | 232 (50.3) | ***/** |
| Proteins (g) | 81.5 (12.4) | 76.6 (11.5) | 70.4 (9.65) | 67.5 (10.5) | ***/*** |
| Fats (g) | 72.0 (15.6) | 62.9 (16.1) | 62.0 (14.0) | 55.0 (12.3) | ***/*** |
| SFA (g) 5 | 28.9 (7.35) | 24.6 (7.32) | 24.6 (7.11) | 21.7 (5.52) | ***/*** |
| MUFA (g) 6 | 24.4 (4.70) | 21.8 (4.95) | 21.5 (4.69) | 19.3 (4.14) | ***/*** |
| PUFA (g) 7 | 14.7 (4.74) | 12.4 (4.68) | 11.9 (4.00) | 9.84 (3.44) | ***/*** |
| Cholesterol (mg) | 354 (37.9) | 346 (34.7) | 336 (32.8) | 329 (35.0) | ***/* |
| Fiber (g) | 19.4 (5.09) | 18.6 (4.87) | 16.3 (4.46) | 16.7 (5.26) | ***/NS |
| Micro-nutrients | |||||
| Ca (mg) | 664 (269) | 632 (235) | 581 (230) | 623 (213) | ./NS |
| p (mg) | 1150 (247) | 1070 (235) | 961 (213) | 931 (182) | ***/* |
| Total Fe (mg) | 14.5 (2.22) | 13.9 (2.19) | 12.8 (1.82) | 12.6 (1.87) | ***/NS |
| Na (mg) | 1420 (389) | 1270 (384) | 1280 (367) | 1160 (292) | ***/*** |
| K (mg) | 3410 (767) | 3300 (870) | 2820 (781) | 2750 (738) | ***/NS |
| Mg (mg) | 269 (54.2) | 257 (62.1) | 221 (46.1) | 214 (46.9) | ***/. |
| Zn (mg) | 10.7 (0.608) | 10.6 (0.596) | 10.2 (0.694) | 10.1 (0.659) | ***/* |
| Cu (mg) | 2.16 (1.16) | 1.87 (1.13) | 1.41 (0.667) | 1.36 (0.583) | ***/. |
| Mn (mg) | 3.36 (0.489) | 3.26 (0.629) | 2.99 (0.498) | 2.93 (0.492) | ***/NS |
| Vitamin A (RE) | 836 (213) | 834 (177) | 756 (148) | 798 (148) | ***/NS |
| B1 (mg) | 1.38 (0.673) | 1.17 (0.318) | 1.06 (0.296) | 1.03 (0.274) | ***/** |
| B2 (mg) | 1.97 (0.842) | 1.80 (0.444) | 1.66 (0.440) | 1.62 (0.453) | ***/. |
| B3 (mg) | 19.7 (7.43) | 17.5 (5.16) | 15.2 (3.71) | 14.1 (3.22) | ***/*** |
| B5 (mg) | 5.79 (2.13) | 5.37 (1.41) | 4.53 (0.867) | 4.51 (1.05) | ***/NS |
| B6 (mg) | 1.56 (0.798) | 1.56 (0.641) | 1.51 (0.675) | 1.40 (0.532) | ./NS |
| B9 (folate) (µg) | 378 (78.1) | 359 (73.6) | 332 (58.1) | 328 (64.0) | ***/. |
| B12 (mg) | 7.28 (0.376) | 7.21 (0.375) | 7.11 (0.350) | 7.02 (0.373) | ***/* |
| Vitamin C (mg) | 171 (68.5) | 183 (62.0) | 150 (58.3) | 162 (59.9) | **/* |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Vega, Á.S.; Corrales-Agudelo, V.; Reyes, A.; Escobar, J.S. Diet Quality, Food Groups and Nutrients Associated with the Gut Microbiota in a Nonwestern Population. Nutrients 2020, 12, 2938. https://doi.org/10.3390/nu12102938
García-Vega ÁS, Corrales-Agudelo V, Reyes A, Escobar JS. Diet Quality, Food Groups and Nutrients Associated with the Gut Microbiota in a Nonwestern Population. Nutrients. 2020; 12(10):2938. https://doi.org/10.3390/nu12102938
Chicago/Turabian StyleGarcía-Vega, Ángela S., Vanessa Corrales-Agudelo, Alejandro Reyes, and Juan S. Escobar. 2020. "Diet Quality, Food Groups and Nutrients Associated with the Gut Microbiota in a Nonwestern Population" Nutrients 12, no. 10: 2938. https://doi.org/10.3390/nu12102938
APA StyleGarcía-Vega, Á. S., Corrales-Agudelo, V., Reyes, A., & Escobar, J. S. (2020). Diet Quality, Food Groups and Nutrients Associated with the Gut Microbiota in a Nonwestern Population. Nutrients, 12(10), 2938. https://doi.org/10.3390/nu12102938