Betaine Supplementation May Improve Heat Tolerance: Potential Mechanisms in Humans


Abstract
1. Introduction and Methodology
2. Betaine
2.1. Methyl Donation
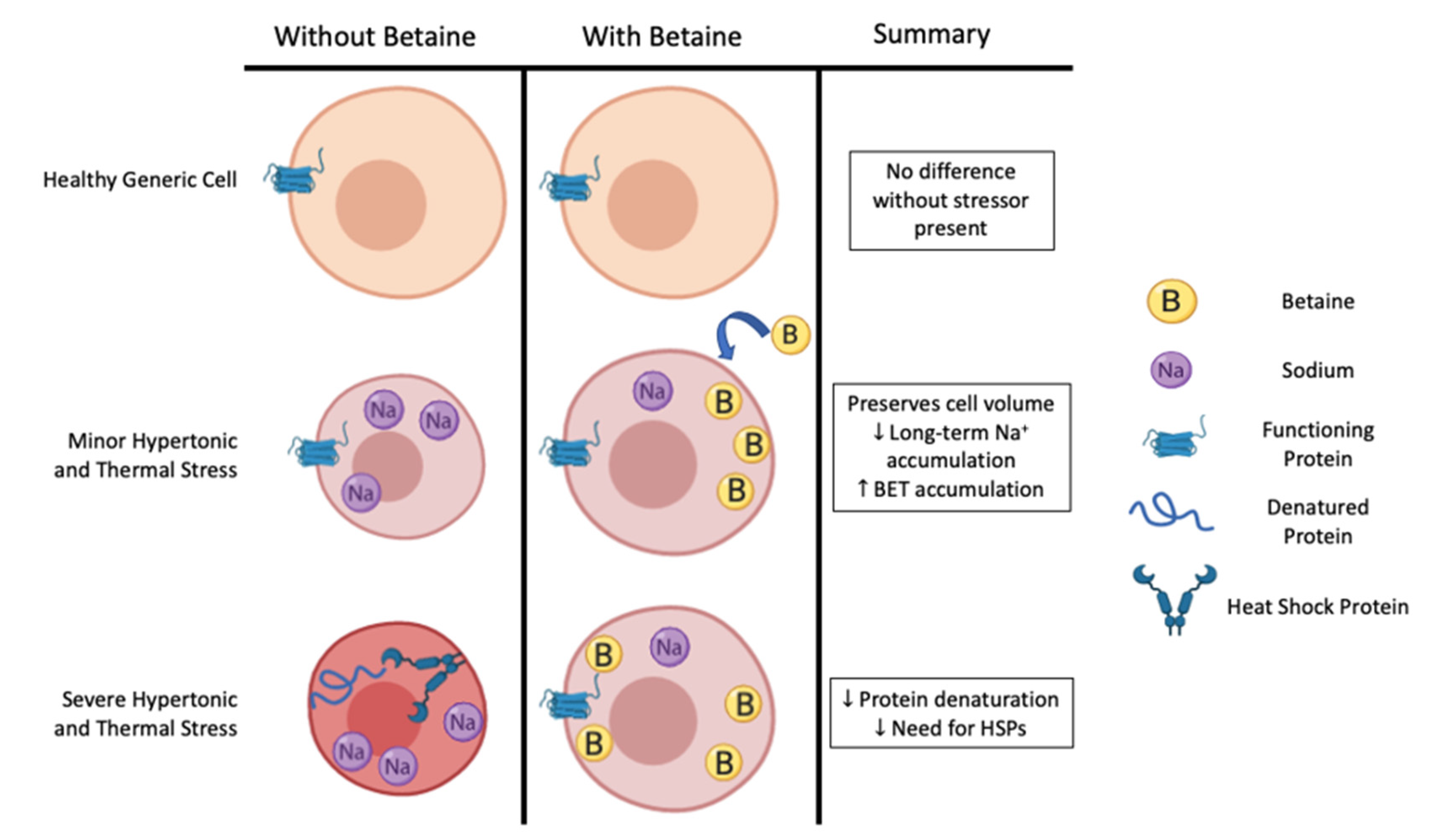
2.2. Osmolyte
3. Heat Stress and Heat Shock Proteins
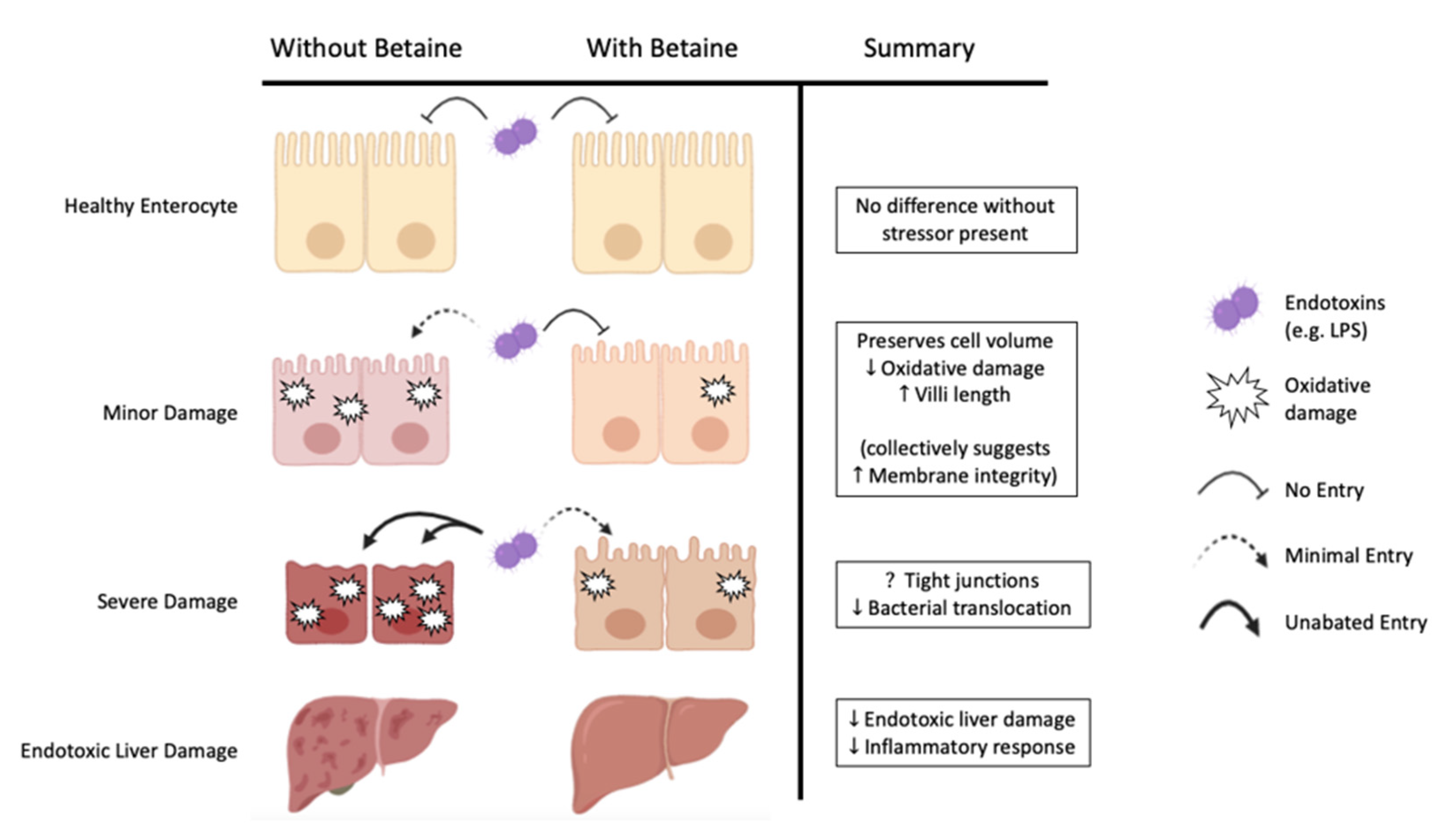
4. Gut and Immune Health
5. Animal Models of Heat Stress
6. Human Models of Heat Stress
7. Conclusions
8. Future Research
Author Contributions
Funding
Conflicts of Interest
References
- Kark, J.A.; Burr, P.Q.; Wenger, C.B.; Gastaldo, E.; Gardner, J.W. Exertional heat illness in marine corps recruit training. Aviat. Space Environ. Med. 1996, 67, 354–360. [Google Scholar] [PubMed]
- Wallace, R.F.; Kriebel, D.; Punnett, L.; Wegman, D.H.; Wenger, C.B.; Gardner, J.W.; Gonzalez, R.R. The effects of continuous hot weather training on risk of exertional heat illness. Med. Sci. Sports Exerc. 2005, 37, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Berko, J.; Ingram, D.D.; Saha, S.; Parker, J.D. Deaths attributed to heat, cold, and other weather events in the United States, 2006–2010. Natl. Health Stat. Rep. 2014, 2014, 1–15. [Google Scholar]
- Argaud, L.; Ferry, T.; Le, Q.H.; Marfisi, A.; Ciorba, D.; Achache, P.; Ducluzeau, R.; Robert, D. Short- and long-term outcomes of heatstroke following the 2003 heat wave in Lyon, France. Arch. Intern. Med. 2007, 167, 2177–2183. [Google Scholar] [CrossRef] [PubMed]
- Azhar, G.S.; Mavalankar, D.; Nori-Sarma, A.; Rajiva, A.; Dutta, P.; Jaiswal, A.; Sheffield, P.; Knowlton, K.; Hess, J.J. Heat-related mortality in India: Excess all-cause mortality associated with the 2010 Ahmedabad heat wave. PLoS ONE 2014, 9, e91831. [Google Scholar] [CrossRef]
- Hoffmann, B.; Hertel, S.; Boes, T.; Weiland, D.; Jöckel, K.H. Increased cause-specific mortality associated with 2003 heat wave in Essen, Germany. J. Toxicol. Environ. Health Part A Curr. Issues 2008, 71, 759–765. [Google Scholar] [CrossRef]
- Garth, A.K.; Burke, L.M. What do athletes drink during competitive sporting activities? Sports Med. 2013, 43, 539–564. [Google Scholar] [CrossRef]
- McCartney, D.; Desbrow, B.; Irwin, C. The Effect of Fluid Intake Following Dehydration on Subsequent Athletic and Cognitive Performance: A Systematic Review and Meta-analysis. Sports Med.-Open 2017, 3, 13. [Google Scholar] [CrossRef]
- Kenefick, R.W. Drinking Strategies: Planned Drinking Versus Drinking to Thirst. Sports Med. 2018, 48, 31–37. [Google Scholar] [CrossRef]
- McDermott, B.P.; Anderson, S.A.; Armstrong, L.E.; Casa, D.J.; Cheuvront, S.N.; Cooper, L.; Larry Kenney, W.; O’Connor, F.G.; Roberts, W.O. National athletic trainers’ association position statement: Fluid replacement for the physically active. J. Athl. Train. 2017, 52, 877–895. [Google Scholar] [CrossRef]
- Casa, D.J.; DeMartini, J.K.; Bergeron, M.F.; Csillan, D.; Eichner, E.R.; Lopez, R.M.; Ferrara, M.S.; Miller, K.C.; O’Connor, F.; Sawka, M.N.; et al. National athletic trainers’ association position statement: Exertional heat illnesses. J. Athl. Train. 2015, 50, 986–1000. [Google Scholar] [CrossRef] [PubMed]
- Craig, S.A. Betaine in human nutrition. Am. J. Clin. Nutr. 2004, 80, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H.; Mar, M.H.; Howe, J.C.; Holden, J.M. Concentrations of choline-containing compounds and betaine in common foods. J. Nutr. 2003, 133, 1302–1307. [Google Scholar] [CrossRef] [PubMed]
- Lever, M.; Sizeland, P.C.B.; Bason, L.M.; Hayman, C.M.; Chambers, S.T. Glycine betaine and proline betaine in human blood and urine. BBA-Gen. Subj. 1994, 1200, 259–264. [Google Scholar] [CrossRef]
- Schwahn, B.C.; Hafner, D.; Hohlfeld, T.; Balkenhol, N.; Laryea, M.D.; Wendel, U. Pharmacokinetics of oral betaine in healthy subjects and patients with homocystinuria. Br. J. Clin. Pharmacol. 2003, 55, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Alfieri, R.R.; Cavazzoni, A.; Petronini, P.G.; Bonelli, M.A.; Caccamo, A.E.; Borghetti, A.F.; Wheeler, K.P. Compatible osmolytes modulate the response of porcine endothelial cells to hypertonicity and protect them from apoptosis. J. Physiol. 2002, 540, 499–508. [Google Scholar] [CrossRef]
- Ueland, P.M. Choline and betaine in health and disease. J. Inherit. Metab. Dis. 2011, 34, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Anioła, M.; Dega-Szafran, Z.; Katrusiak, A.; Szafran, M. NH⋯O and OH⋯O interactions of glycine derivatives with squaric acid. New J. Chem. 2014, 38, 3556–3568. [Google Scholar] [CrossRef]
- Finkelstein, J.D.; Harris, B.J.; Kyle, W.E. Methionine metabolism in mammals: Kinetic study of betaine-homocysteine methyltransferase. Arch. Biochem. Biophys. 1972, 153, 320–324. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Blusztajn, J.K. Choline and human nutrition. Annu. Rev. Nutr. 1994, 14, 269–296. [Google Scholar] [CrossRef]
- Borsook, H.; Borsook, M.E. The biochemical basis of betaine-glycocyamine therapy. Ann. West. Med. Surg. 1951, 5, 825–829. [Google Scholar] [PubMed]
- Da Silva, R.P.; Nissim, I.; Brosnan, M.E.; Brosnan, J.T. Creatine synthesis: Hepatic metabolism of guanidinoacetate and creatine in the rat In Vitro and In Vivo. Am. J. Physiol.-Endocrinol. Metab. 2009, 296, E256–E261. [Google Scholar] [CrossRef] [PubMed]
- Pekkinen, J.; Olli, K.; Huotari, A.; Tiihonen, K.; Keski-Rahkonen, P.; Lehtonen, M.; Auriola, S.; Kolehmainen, M.; Mykkänen, H.; Poutanen, K.; et al. Betaine supplementation causes increase in carnitine metabolites in the muscle and liver of mice fed a high-fat diet as studied by nontargeted LC-MS metabolomics approach. Mol. Nutr. Food Res. 2013, 57, 1959–1968. [Google Scholar] [CrossRef]
- Carnicelli, D.; Arfilli, V.; Onofrillo, C.; Alfieri, R.R.; Petronini, P.G.; Montanaro, L.; Brigotti, M. Cap-independent protein synthesis is enhanced by betaine under hypertonic conditions. Biochem. Biophys. Res. Commun. 2017, 483, 936–940. [Google Scholar] [CrossRef] [PubMed]
- Best, C.H.; Ridout, J.H.; Lucas, C.C. Alleviation of dietary cirrhosis by betaine and other lipotrophic agents. Nutr. Rev. 1969, 27, 269–271. [Google Scholar]
- Schwab, U.; Törrönen, A.; Toppinen, L.; Alfthan, G.; Saarinen, M.; Aro, A.; Uusitupa, M. Betaine supplementation decreases plasma homocysteine concentrations but does not affect body weight, body composition, or resting energy expenditure in human subjects. Am. J. Clin. Nutr. 2002, 76, 961–967. [Google Scholar] [CrossRef]
- Abdelmalek, M.F.; Sanderson, S.O.; Angulo, P.; Soldevila-Pico, C.; Liu, C.; Peter, J.; Keach, J.; Cave, M.; Chen, T.; McClain, C.J.; et al. Betaine for nonalcoholic fatty liver disease: Results of a randomized placebo-controlled trial. Hepatology 2009, 50, 1818–1826. [Google Scholar] [CrossRef]
- Lee, E.C.; Maresh, C.M.; Kraemer, W.J.; Yamamoto, L.M.; Hatfield, D.L.; Bailey, B.L.; Armstrong, L.E.; Volek, J.S.; McDermott, B.P.; Craig, S.A. Ergogenic effects of betaine supplementation on strength and power performance. J. Int. Soc. Sports Nutr. 2010, 7, 27. [Google Scholar] [CrossRef]
- Hoffman, J.R.; Ratamess, N.A.; Kang, J.; Gonzalez, A.M.; Beller, N.A.; Craig, S.A.S. Effect of 15 days of betaine ingestion on concentric and eccentric force outputs during isokinetic exercise. J. Strength Cond. Res. 2011, 25, 2235–2241. [Google Scholar] [CrossRef]
- Pryor, J.L.; Craig, S.A.; Swensen, T. Effect of betaine supplementation on cycling sprint performance. J. Int. Soc. Sports Nutr. 2012, 9, 12. [Google Scholar]
- Cholewa, J.M.; Guimarães-Ferreira, L.; Zanchi, N.E. Effects of betaine on performance and body composition: A review of recent findings and potential mechanisms. Amino Acids 2014, 46, 1785–1793. [Google Scholar] [CrossRef] [PubMed]
- Ismaeel, A. Effects of Betaine Supplementation on Muscle Strength and Power: A Systematic Review. J. Strength Cond. Res. 2017, 31, 2338–2346. [Google Scholar] [CrossRef] [PubMed]
- Burg, M.B. Molecular basis of osmotic regulation. Am. J. Physiol.-Ren. Fluid Electrolyte Physiol. 1995, 268, F983–F996. [Google Scholar] [CrossRef] [PubMed]
- Sheikh-Hamad, D.; Garcia-Perez, A.; Ferraris, J.D.; Peters, E.M.; Burg, M.B. Induction of Gene Expression by Heat Shock Versus Osmotic Stress. Am. J. Physiol.-Ren. Fluid Electrolyte Physiol. 1994, 267, F28–F34. [Google Scholar] [CrossRef]
- Garcia-Perez, A.; Burg, M.B. Renal medullary organic osmolytes. Physiol. Rev. 1991, 71, 1081–1115. [Google Scholar] [CrossRef]
- Kenefick, R.W.; Cheuvront, S.N.; Sawka, M.N. Thermoregulatory function during the marathon. Sports Med. 2007, 37, 312–315. [Google Scholar] [CrossRef]
- Rowell, L.B. Human cardiovascular adjustments to exercise and thermal stress. Physiol. Rev. 1974, 54, 75–159. [Google Scholar] [CrossRef]
- Taylor, N.A. Eccrine sweat glands. Adaptations to Physical Training and Heat Acclimation. Sports Med. 1986, 3, 387–397. [Google Scholar] [CrossRef]
- Baskakov, I.; Bolen, D.W. Forcing thermodynamically unfolded proteins to fold. J. Biol. Chem. 1998, 273, 4831–4834. [Google Scholar] [CrossRef]
- Yancey, P.H.; Clark, M.E.; Hand, S.C.; Bowlus, R.D.; Somero, G.N. Living with water stress: Evolution of osmolyte systems. Science 1982, 217, 1214–1222. [Google Scholar] [CrossRef]
- Street, T.O.; Krukenberg, K.A.; Rosgen, J.; Bolen, D.W.; Agard, D.A. Osmolyte-Induced Conformational Changes in the Hsp90 Molecular Chaperone. Protein Sci. 2010, 19, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Zininga, T.; Ramatsui, L.; Shonhai, A. Heat shock proteins as immunomodulants. Molecules 2018, 23, 2846. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fernández, M.R.; Valpuesta, J.M. Hsp70 chaperone: A master player in protein homeostasis. F1000Research 2018, 7, 1497. [Google Scholar]
- Young, J.C.; Moarefi, I.; Ulrich Hartl, F. Hsp90: A specialized but essential protein-folding tool. J. Cell Biol. 2001, 154, 267–273. [Google Scholar] [CrossRef]
- Pires, W.; Veneroso, C.E.; Wanner, S.P.; Pacheco, D.A.S.; Vaz, G.C.; Amorim, F.T.; Tonoli, C.; Soares, D.D.; Coimbra, C.C. Association Between Exercise-Induced Hyperthermia and Intestinal Permeability: A Systematic Review. Sports Med. 2017, 47, 1389–1403. [Google Scholar] [CrossRef]
- Street, T.O.; Bolen, D.W.; Rose, G.D. A molecular mechanism for osmolyte-induced protein stability. Proc. Natl. Acad. Sci. USA 2006, 103, 13997–14002. [Google Scholar] [CrossRef]
- Seeliger, J.; Estel, K.; Erwin, N.; Winter, R. Cosolvent effects on the fibrillation reaction of human IAPP. Phys. Chem. Chem. Phys. 2013, 15, 8902–8907. [Google Scholar] [CrossRef]
- Fan, Y.Q.; Lee, J.; Oh, S.; Liu, H.J.; Li, C.; Luan, Y.S.; Yang, J.M.; Zhou, H.M.; Lü, Z.R.; Wang, Y.L. Effects of osmolytes on human brain-type creatine kinase folding in dilute solutions and crowding systems. Int. J. Biol. Macromol. 2012, 51, 845–858. [Google Scholar] [CrossRef]
- Beck, F.X.; Grünbein, R.; Lugmayr, K.; Neuhofer, W. Heat shock proteins and the cellular response to osmotic stress. Cell. Physiol. Biochem. 2000, 10, 303–306. [Google Scholar] [CrossRef]
- Alfieri, R.R.; Petronini, P.G.; Bonelli, M.A.; Desenzani, S.; Cavazzoni, A.; Borghetti, A.F.; Wheeler, K.P. Roles of compatible osmolytes and heat shock protein 70 in the induction of tolerance to stresses in porcine endothelial cells. J. Physiol. 2004, 555, 757–767. [Google Scholar] [CrossRef]
- Neuhofer, W.; Müller, E.; Grünbein, R.; Thurau, K.; Beck, F.-X. Influence of NaCl, urea, potassium and pH on HSP72 expression in MDCK cells. Pflügers Arch.-Eur. J. Physiol. 1999, 439, 195–200. [Google Scholar]
- Oliva, J.; Bardag-Gorce, F.; Tillman, B.; French, S.W. Protective effect of quercetin, EGCG, catechin and betaine against oxidative stress induced by ethanol in vitro. Exp. Mol. Pathol. 2011, 90, 295–299. [Google Scholar] [PubMed]
- Dangi, S.S.; Dangi, S.K.; Chouhan, V.S.; Verma, M.R.; Kumar, P.; Singh, G.; Sarkar, M. Modulatory effect of betaine on expression dynamics of HSPs during heat stress acclimation in goat (Capra hircus). Gene 2016, 575, 543–550. [Google Scholar] [PubMed]
- Walsh, R.C.; Koukoulas, I.; Garnham, A.; Moseley, P.L.; Hargreaves, M.; Febbraio, M.A. Exercise increases serum Hsp72 in humans. Cell Stress Chaperones 2001, 6, 386–393. [Google Scholar] [PubMed]
- Bruzdźiak, P.; Panuszko, A.; Jourdan, M.; Stangret, J. Protein thermal stabilization in aqueous solutions of osmolytes. Acta Biochim. Pol. 2016, 63, 65–70. [Google Scholar] [PubMed]
- Stewart, I.; Schluter, P.J.; Shaw, G.R. Cyanobacterial lipopolysaccharides and human health—A review. Environ. Health A Glob. Access Sci. Source 2006, 5, 7. [Google Scholar]
- Zuhl, M.; Schneider, S.; Lanphere, K.; Conn, C.; Dokladny, K.; Moseley, P. Exercise regulation of intestinal tight junction proteins. Br. J. Sports Med. 2012, 48, 980–986. [Google Scholar]
- Lim, C.L.; Mackinnon, L.T. The roles of exercise-induced immune system disturbances in the pathology of heat stroke: The dual pathway model of heat stroke. Sports Med. 2006, 36, 39–64. [Google Scholar]
- Brock-Utne, J.G.; Gaffin, S.L.; Wells, M.T.; Gathiram, P.; Sohar, E.; James, M.F.; Morrell, D.F.; Norman, R.J. Endotoxaemia in exhausted runners after a long-distance race. S. Afr. Med. J. 1988, 73, 533–536. [Google Scholar]
- Chao, T.C.; Sinniah, R.; Pakiam, J.E. Acute heat stroke deaths. Pathology 1981, 13, 145–156. [Google Scholar]
- Shapiro, Y.; Seidman, D.S. Field and clinical observations of exertional heat stroke patients. Med. Sci. Sports Exerc. 1990, 22, 6–14. [Google Scholar] [CrossRef]
- Clausen, J.P. Effect of physical training on cardiovascular adjustments to exercise in man. Physiol. Rev. 1977, 57, 779–815. [Google Scholar] [CrossRef] [PubMed]
- Granger, D.N.; Kvietys, P.R. The Splanchnic Circulation: Intrinsic Regulation. Annu. Rev. Physiol. 1981, 43, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Sakurada, S.; Hales, J.R.S. A Role for Gastrointestinal Endotoxins in Enhancement of Heat Tolerance by Physical Fitness. J. Appl. Physiol. 1998, 84, 207–214. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Van Wijck, K.; Lenaerts, K.; van Loon, L.J.C.; Peters, W.H.M.; Buurman, W.A.; Dejong, C.H.C. Exercise-Induced splanchnic hypoperfusion results in gut dysfunction in healthy men. PLoS ONE 2011, 6, e22366. [Google Scholar] [CrossRef]
- Moseley, P.L.; Gisolfi, C.V. New Frontiers in Thermoregulation and Exercise. Sports Med. 1993, 16, 163–167. [Google Scholar] [CrossRef]
- Hall, D.M.; Baumgardner, K.R.; Oberley, T.D.; Gisolfi, C.V. Splanchnic tissues undergo hypoxic stress during whole body hyperthermia. Am. J. Physiol.-Gastrointest. Liver Physiol. 1999, 276, G1195. [Google Scholar] [CrossRef]
- Walsh, N.P.; Gleeson, M.; Pyne, D.B.; Nieman, D.C.; Dhabhar, S.; Shephard, R.J.; Oliver, S.J.; Bermon, S.; Kajeniene, A. Position Statement. Part two: Maintaining immune health. Exerc. Immunol. Rev. 2011, 17, 64–103. [Google Scholar]
- Hall, D.M.; Buettner, G.R.; Oberley, L.W.; Xu, L.; Matthes, R.D.; Gisolfi, C.V. Mechanisms of Circulatory and Intestinal Barrier Dysfunction during Whole Body Hyperthermia. Am. J. Physiol.-Hear. Circ. Physiol. 2001, 280, H509–H521. [Google Scholar] [CrossRef]
- Lambert, G.P. Stress-Induced Gastrointestinal Barrier Dysfunction and its Inflammatory Effects. J. Anim. Sci. 2009, 87, E101–E108. [Google Scholar] [CrossRef]
- Selkirk, G.A.; McLellan, T.M.; Wright, H.E.; Rhind, S.G. Mild endotoxemia, NF-κB translocation, and cytokine increase during exertional heat stress in trained and untrained individuals. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2008, 295. [Google Scholar] [CrossRef] [PubMed]
- Vargas, N.; Marino, F. Heat stress, gastrointestinal permeability and interleukin-6 signaling—Implications for exercise performance and fatigue. Temperature 2016, 3, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Alhenaky, A.; Abdelqader, A.; Abuajamieh, M.; Al-Fataftah, A.R. The effect of heat stress on intestinal integrity and Salmonella invasion in broiler birds. J. Therm. Biol. 2017, 70, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Hamesch, K.; Borkham-Kamphorst, E.; Strnad, P.; Weiskirchen, R. Lipopolysaccharide-induced inflammatory liver injury in mice. Lab. Anim. 2015, 49, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Kim, Y.C. Attenuation of bacterial lipopolysaccharide-induced hepatotoxicity by betaine or taurine in rats. Food Chem. Toxicol. 2002, 40, 545–549. [Google Scholar] [CrossRef]
- Jirik, F.R.; Podor, T.J.; Hirano, T.; Kishimoto, T.; Loskutoff, D.J.; Carson, D.A.; Lotz, M. Bacterial lipopolysaccharide and inflammatory mediators augment IL-6 secretion by human endothelial cells. J. Immunol. 1989, 142, 144–147. [Google Scholar]
- Snipe, R.M.J.; Costa, R.J.S. Does the temperature of water ingested during exertional-heat stress influence gastrointestinal injury, symptoms, and systemic inflammatory profile? J. Sci. Med. Sport 2018, 21, 771–776. [Google Scholar] [CrossRef]
- Costa, R.J.S.; Camões-Costa, V.; Snipe, R.M.J.; Dixon, D.; Russo, I.; Huschtscha, Z. Impact of Exercise-Induced Hypohydration on Gastrointestinal Integrity, Function, Symptoms, and Systemic Endotoxin and Inflammatory Profile. J. Appl. Physiol. 2019, 126, 1281–1291. [Google Scholar] [CrossRef]
- Olesen, J.; Biensø, R.S.; Meinertz, S.; Van Hauen, L.; Rasmussen, S.M.; Gliemann, L.; Plomgaard, P.; Pilegaard, H. Impact of training status on LPS-induced acute inflammation in humans. J. Appl. Physiol. 2015, 118, 818–829. [Google Scholar] [CrossRef]
- Ganesan, B.; Anandan, R.; Lakshmanan, P.T. Studies on the protective effects of betaine against oxidative damage during experimentally induced restraint Stress in Wistar albino rats. Cell Stress Chaperones 2011, 16, 641–652. [Google Scholar] [CrossRef]
- Wang, H.; Li, S.; Fang, S.; Yang, X.; Feng, J. Betaine improves intestinal functions by enhancing digestive enzymes, ameliorating intestinal morphology, and enriching intestinal microbiota in high-salt stressed rats. Nutrients 2018, 10, 907. [Google Scholar]
- Orawan, C.; Aengwanich, W. Blood cell characteristics, hematological values and average daily gained weight of Thai indigenous, Thai indigenous crossbred and broiler chickens. Pakistan J. Biol. Sci. 2007, 10, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Hao, Y.; Li, J.; Bao, W.; Li, G.; Gao, Y.; Gu, X. Chronic heat stress induces immune response, oxidative stress response, and apoptosis of finishing pig liver: A proteomic approach. Int. J. Mol. Sci. 2016, 17, 393. [Google Scholar] [CrossRef] [PubMed]
- Khattak, F.M.; Acamovic, T.; Sparks, N.; Pasha, T.N.; Joiya, M.H.; Hayat, Z.; Ali, Z. Comparative efficacy of different supplements used to reduce heat stress in broilers. Pak. J. Zool. 2012, 44, 31–41. [Google Scholar]
- Costa, R.J.S.; Crockford, M.J.; Moore, J.P.; Walsh, N.P. Heat acclimation responses of an ultra-endurance running group preparing for hot desert-based competition. Eur. J. Sport Sci. 2014, 14, 131–141. [Google Scholar]
- Snipe, R.M.J.; Khoo, A.; Kitic, C.M.; Gibson, P.R.; Costa, R.J.S. Carbohydrate and protein intake during exertional heat stress ameliorates intestinal epithelial injury and small intestine permeability. Appl. Physiol. Nutr. Metab. 2017, 42, 1283–1292. [Google Scholar] [CrossRef]
- Sahebi Ala, F.; Hassanabadi, A.; Golian, A. Effects of dietary supplemental methionine source and betaine replacement on the growth performance and activity of mitochondrial respiratory chain enzymes in normal and heat-stressed broiler chickens. J. Anim. Physiol. Anim. Nutr. (Berl.) 2019, 103, 87–99. [Google Scholar] [CrossRef]
- Zhang, L.; An, W.J.; Lian, H.; Zhou, G.B.; Han, Z.Y.; Ying, S.J. Effects of dietary betaine supplementation subjected to heat stress on milk performances and physiology indices in dairy cow. Genet. Mol. Res. 2014, 13, 7577–7586. [Google Scholar]
- Ratriyanto, A.; Mosenthin, R. Osmoregulatory function of betaine in alleviating heat stress in poultry. J. Anim. Physiol. Anim. Nutr. (Berl.) 2018, 102, 1634–1650. [Google Scholar]
- Zulkifli, I.; Mysahra, S.A.; Jin, L.Z. Dietary Supplementation of Betaine (Betafin®) and Response to High Temperature Stress in Male Broiler Chickens. Asian-Australasian J. Anim. Sci. 2004, 17, 244–249. [Google Scholar] [CrossRef]
- DiGiacomo, K.; Simpson, S.; Leury, B.J.; Dunshea, F.R. Dietary betaine impacts the physiological responses to moderate heat conditions in a dose dependent manner in sheep. Animals 2016, 6, 51. [Google Scholar] [CrossRef] [PubMed]
- Attia, Y.A.; Hassan, R.A.; Qota, E.M.A. Recovery from adverse effects of heat stress on slow-growing chicks in the tropics 1: Effect of ascorbic acid and different levels of betaine. Trop. Anim. Health Prod. 2009, 41, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Jiao, H.; Song, Z.; Zhao, J.; Wang, X.; Lin, H. Heat stress impairs mitochondria functions and induces oxidative injury in broiler chickens. J. Anim. Sci. 2015, 93, 2144–2153. [Google Scholar] [CrossRef] [PubMed]
- Garrett, Q.; Khandekar, N.; Shih, S.; Flanagan, J.L.; Simmons, P.; Vehige, J.; Willcox, M.D.P. Betaine stabilizes cell volume and protects against apoptosis in human corneal epithelial cells under hyperosmotic stress. Exp. Eye Res. 2013, 108, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.T.; Fujita, M.; Yamamoto, S.; Iwasaki, K.; Ikawa, R.; Oyama, H.; Horikawa, H. Effects of Glucose in Drinking Water on the Changes in Whole Blood Viscosity and Plasma Osmolality of Broiler Chickens during High Temperature Exposure. Poult. Sci. 1998, 77, 644–647. [Google Scholar] [CrossRef] [PubMed]
- Park, S.O.; Kim, W.K. Effects of betaine on biological functions in meat-type ducks exposed to heat stress. Poult. Sci. 2017, 96, 1212–1218. [Google Scholar] [CrossRef]
- Borges, S.A.; Fischer Da Silva, A.V.; Majorka, A.; Hooge, D.M.; Cummings, K.R. Physiological responses of broiler chickens to heat stress and dietary electrolyte balance (sodium plus potassium minus chloride, milliequivalents per kilogram). Poult. Sci. 2004, 83, 1551–1558. [Google Scholar] [CrossRef]
- Armstrong, L.E.; Casa, D.J.; Roti, M.W.; Lee, E.C.; Craig, S.A.S.; Sutherland, J.W.; Fiala, K.A.; Maresh, C.M. Influence of betaine consumption on strenuous running and sprinting in a hot environment. J. Strength Cond. Res. 2008, 22, 851–860. [Google Scholar] [CrossRef]
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Ringuet, M.; Furness, J.B.; Dunshea, F.R. Betaine and antioxidants improve growth performance, breast muscle development and ameliorate thermoregulatory responses to cyclic heat exposure in broiler chickens. Animals 2018, 8, 162. [Google Scholar] [CrossRef]
- Egbuniwe, I.C.; Ayo, J.O.; Ocheja, O.B. Betaine and ascorbic acid modulate indoor behavior and some performance indicators of broiler chickens in response to hot-dry season. J. Therm. Biol. 2018, 76, 38–44. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Author | Population | Supplementation | Thermal Stress | Significant Findings (Compared to Identified CON) |
|---|---|---|---|---|
| Zulkifli et al., 2004 | Chickens (N = 150) | Ad libitum intake, water supplemented with 0 (CON) or 50 g·kg−1 BET | Cyclical heat stress (36 °C, 75% RH) for 4 h·day−1, 6 days | BET core temperature immediately post-heat stress |
| Attia et al., 2009 | Chickens (N = 300) | Ad libitum intake, feed supplemented with 0 (CON) or 1.0 g·kg−1 BET | Cyclical heat stress (38 °C, 49% RH) for 6 h·day−1, 3 days·week−1, 9 weeks | BET core temperature during and after heat stress BET Hgb during and after heat stress BET blood pH during, but not after heat stress |
| Khattak et al., 2012 | Chickens (N = 250) | Ad libitum intake, feed supplemented with 0 (CON) or 1.2 g·kg−1 BET | Natural daily cyclical heat stress (30–41 °C, 40–93% RH), 35 days | BET total leukocyte count BET mortality 10-fold |
| Dangi et al., 2015 | Goats (N = 18) | Intramuscular injections of saline (CON) or saline + 0.2 g·kg−1 BET immediately prior to heat stress | Cyclical heat stress (42 °C, 36% RH) for 6 h·day−1, 16 days | BET HSP60, HSP70, and HSP90 |
| DiGiacomo et al., 2016 | Sheep (N = 36) | Ad libitum intake, feed supplemented with 0 (CON), 2, or 4 g BET daily in morning feed | Cyclical heat stress (43 °C, 49% RH) for 8 h·day−1, 21 days | BET (2 g) + Heat core and skin temperature |
| Sahebi Ala et al., 2017 | Chickens (N = 1200) | BET as a replacement for 30% methionine needs according to methyl groups | Cyclical heat stress (32 °C, 40% RH) for 6 h·day−1, 31 days | BET + Heat mitochondrial Complex-1 function, whereas Heat alone mitochondrial Complex-1 function |
| Park and Kim, 2017 | Ducks (N = 360) | Ad libitum intake, water supplemented with 0 (CON), 700, 1000, or 1300 ppm BET | Cyclical heat stress (33–43 °C, 70% RH) for 8 h·day−1, 20 days | All doses of BET RBC count, Hct, and Hgb PO2 and PCO2 blood pH Blood electrolyte concentrations (Na+, K+, Cl−) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Willingham, B.D.; Ragland, T.J.; Ormsbee, M.J. Betaine Supplementation May Improve Heat Tolerance: Potential Mechanisms in Humans. Nutrients 2020, 12, 2939. https://doi.org/10.3390/nu12102939
Willingham BD, Ragland TJ, Ormsbee MJ. Betaine Supplementation May Improve Heat Tolerance: Potential Mechanisms in Humans. Nutrients. 2020; 12(10):2939. https://doi.org/10.3390/nu12102939
Chicago/Turabian StyleWillingham, Brandon D., Tristan J. Ragland, and Michael J. Ormsbee. 2020. "Betaine Supplementation May Improve Heat Tolerance: Potential Mechanisms in Humans" Nutrients 12, no. 10: 2939. https://doi.org/10.3390/nu12102939
APA StyleWillingham, B. D., Ragland, T. J., & Ormsbee, M. J. (2020). Betaine Supplementation May Improve Heat Tolerance: Potential Mechanisms in Humans. Nutrients, 12(10), 2939. https://doi.org/10.3390/nu12102939