Orexin-A Exerts Equivocal Role in Atherosclerosis Process Depending on the Duration of Exposure: In Vitro Study

 , ,
, , 
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Cell Culture and Treatment
2.2. MTS Cell Proliferation Assay
2.3. qRT-PCR
2.4. SDS-PAGE and Western-Blot Analysis
2.5. Gelatin Zymography
2.6. Enzyme Linked Immunosorbent Assay (ELISA)
2.7. Statistical Analysis
3. Results
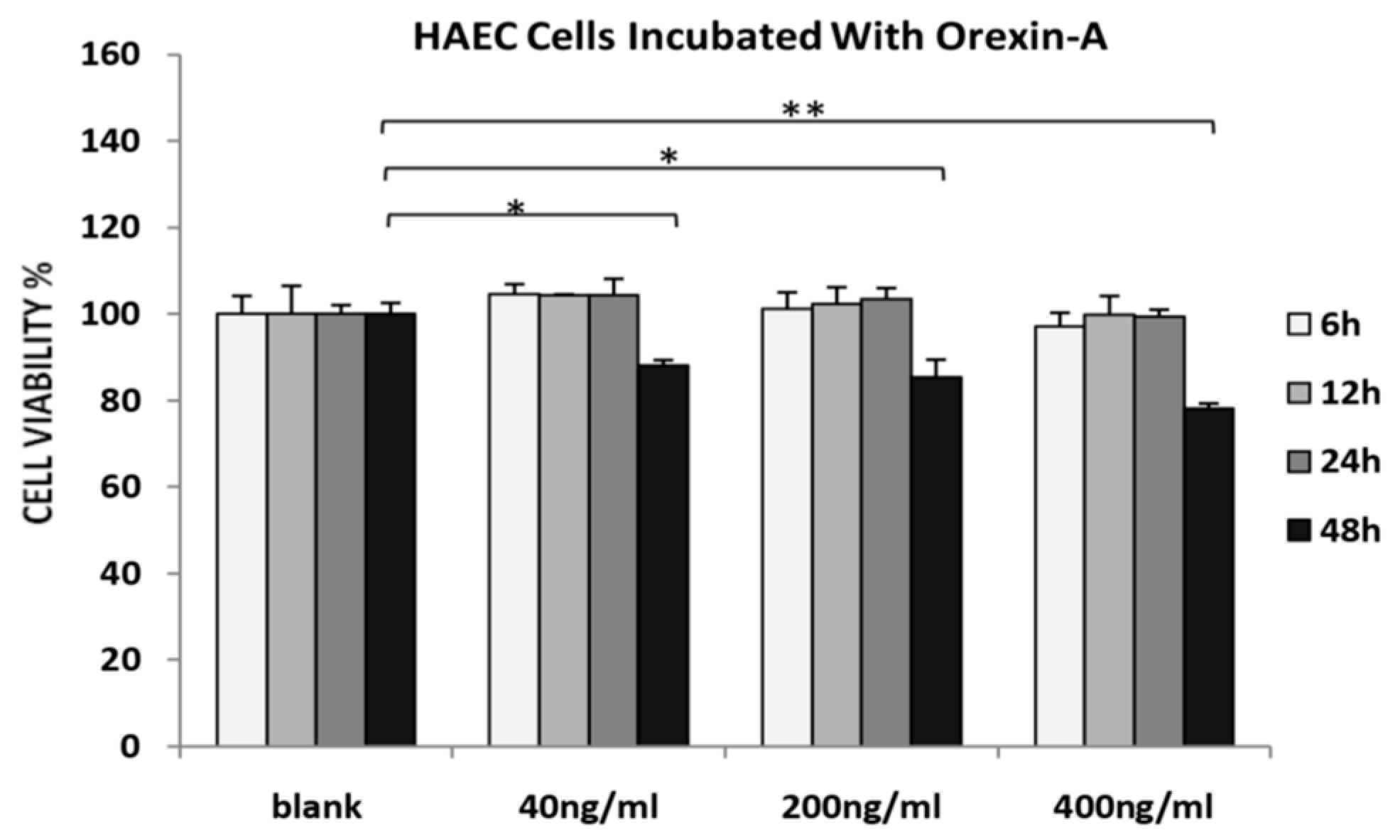
3.1. Orexin-A Did Not Affect Cell Viability/ Proliferation of HAECs after 24 Hours of Incubation
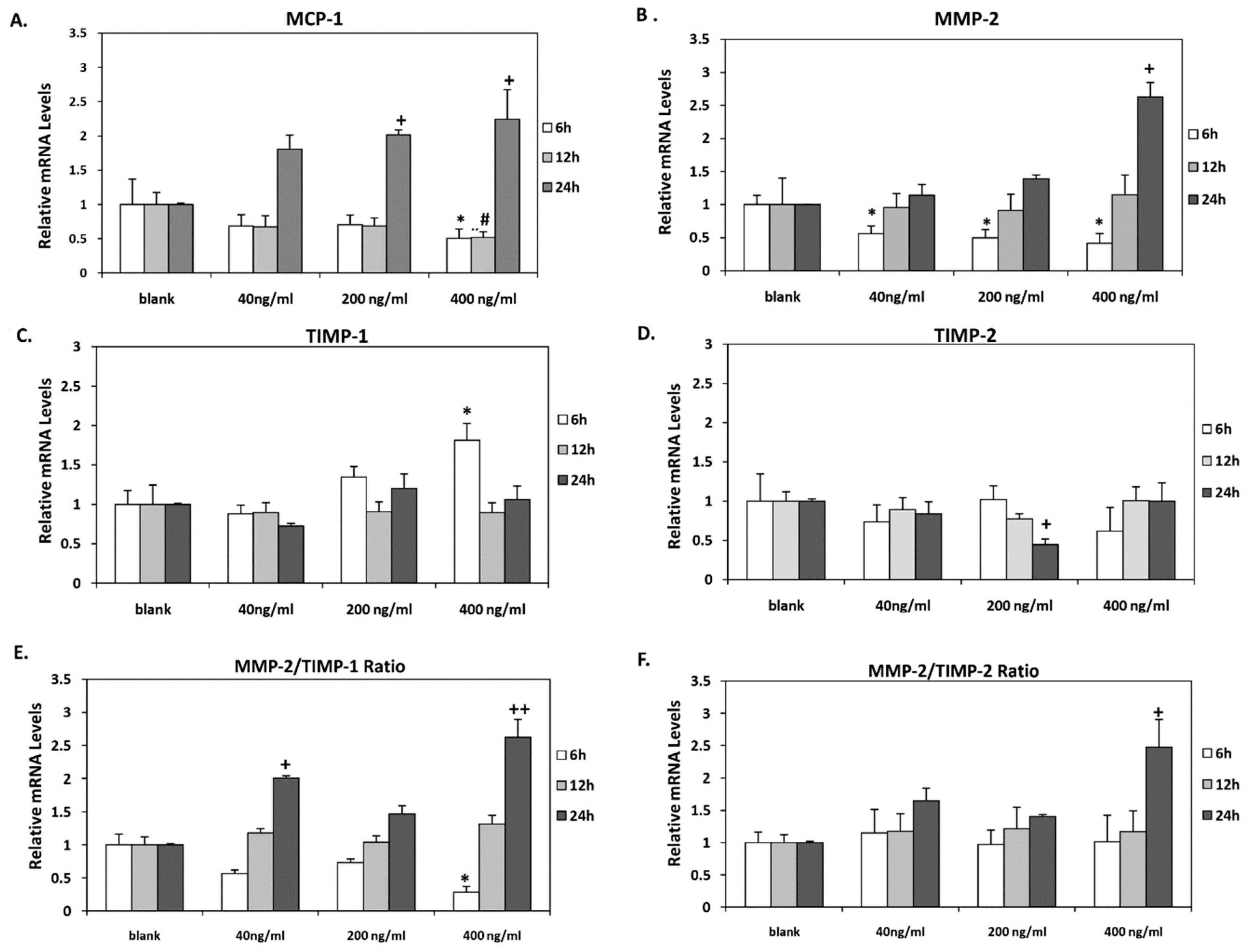
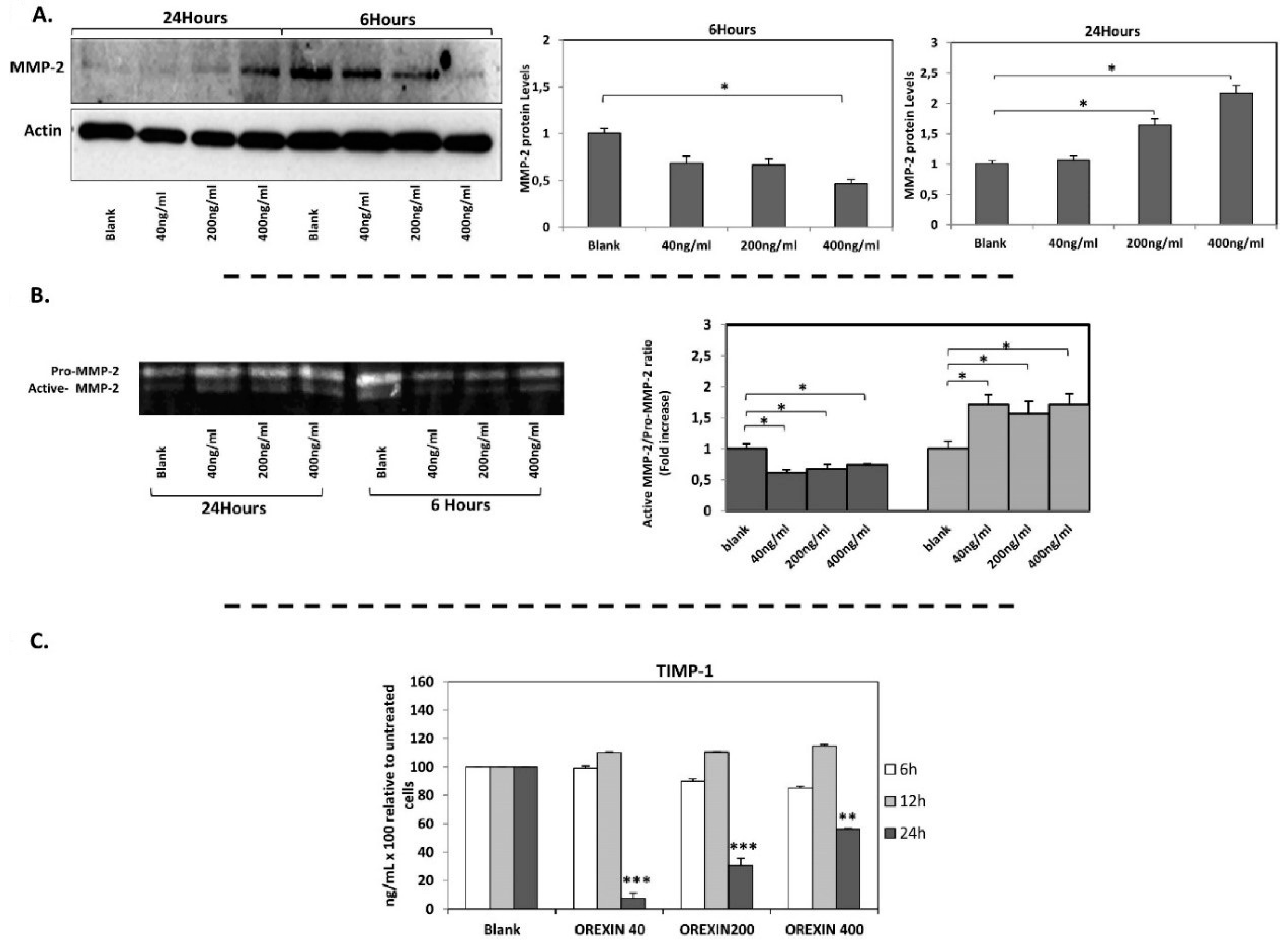
3.2. Orexin-A Exerted A Dual Role in MCP-1, MMP-2, TIMP-1 Expression and in MMP-2 Activity
3.3. Orexin-A Differently Regulated MMPs/TIMPs Ratio Depending on the Duration of Exposure
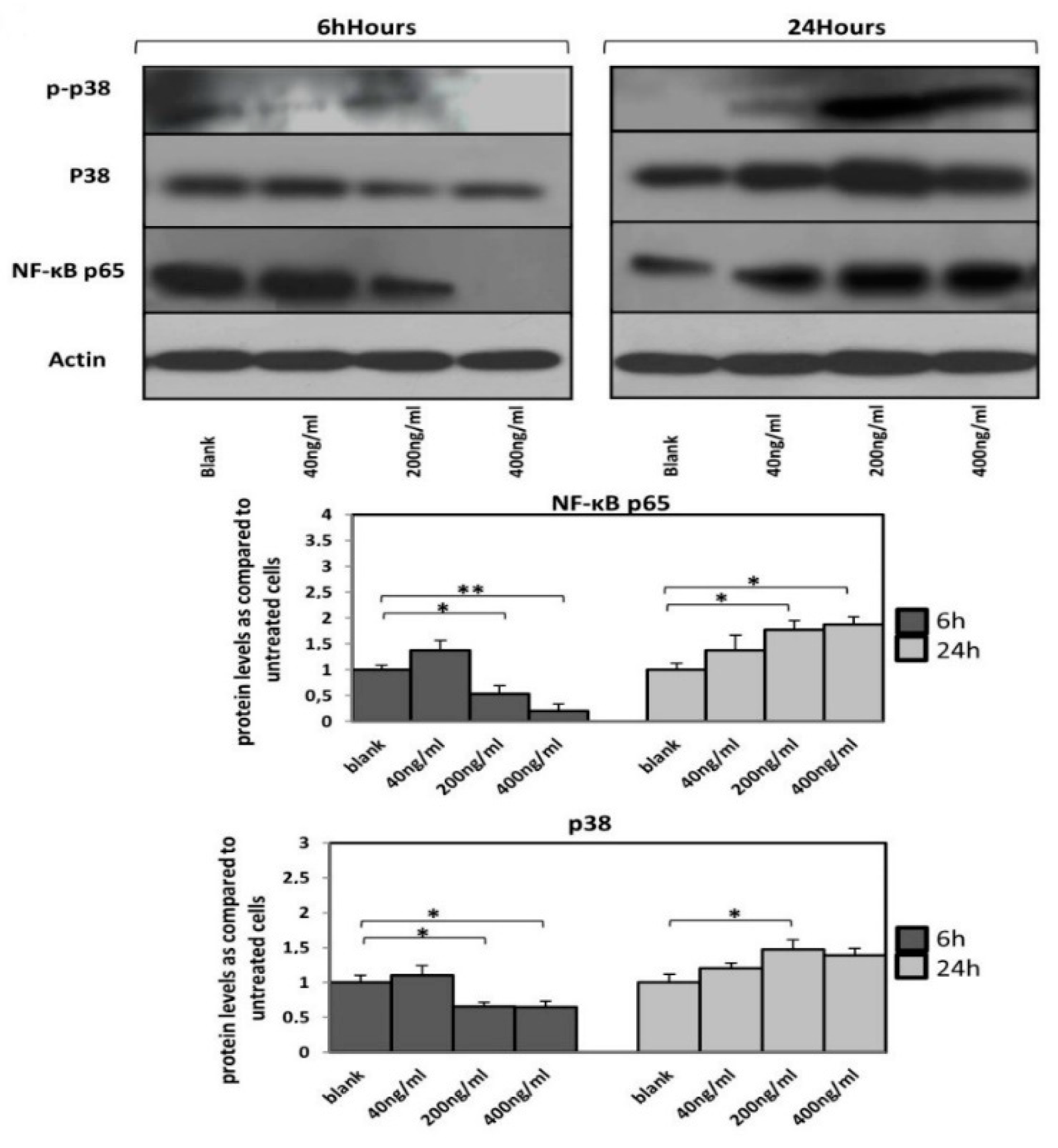
3.4. Orexin-A Can Either Activate or Deactivate p38 and NF-κB p65 Pathway Depending on the Duration of Exposure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HAECs | human aortic endothelial cells |
| HUVECs | human umbilical vein endothelial cells |
| OX1R | Orexin receptor type 1 |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| MMP-2 | matrix metalloproteinase-2 |
| MMP-9 | matrix metalloproteinase-9 |
| MCP-1 | monocyte chemoattractant protein 1 |
| TIMP-1 | tissue inhibitor of metalloproteinases-1 |
| TIMP-2 | tissue inhibitor of metalloproteinases-2 |
| CCL-2 | chemokine (C-C motif) ligand 2 |
| HO_1 | heme oxygenase-1 |
References
- Conti, P.; Shaik-Dasthagirisaeb, Y. Atherosclerosis: A chronic inflammatory disease mediated by mast cells. Cent. J. Immunol. 2015, 40, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Rognoni, A.; Cavallino, C.; Veia, A.; Bacchini, S.; Rosso, R.; Facchini, G.; Secco, G.; Lupi, A.; Nardi, F.; Rametta, F.; et al. Pathophysiology of Atherosclerotic Plaque Development. Cardiovasc. Hematol. Agents Med. Chem. 2015, 13, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Sridevi, D.; Yan, X.D.; Ishwarlal, J. C-Reactive Protein Increases Plasminogen Activator Inhibitor-1 Expression and Activity in Human Aortic Endothelial Cells. Circulation 2003, 107, 398–404. [Google Scholar]
- Lin, J.; Kakkar, V.; Lu, X. Impact of MCP-1 in atherosclerosis. Curr. Pharm. Des. 2014, 20, 4580–4588. [Google Scholar] [CrossRef] [PubMed]
- Masciantonio, M.G. The Balance Between Metalloproteinases and TIMPs: Critical Regulator of Microvascular Endothelial Cell Function in Health and Disease. Prog. Mol. Biol. Transl. Sci. 2017, 147, 101–131. [Google Scholar]
- Kodadek, T.; Cai, D. Chemistry and biology of orexin signaling. Mol. Biosyst. 2010, 6, 1366–1375. [Google Scholar] [CrossRef]
- Heinonen, M.V.; Purhonen, A.K.; Makela, K.A.; Herzig, K.H. Functions of orexins in peripheral tissues. Acta Physiol. (Oxf.) 2008, 192, 471–485. [Google Scholar] [CrossRef]
- Duffy, C.M.; Yuan, C.; Wisdorf, L.E.; Billington, C.J.; Kotz, C.M.; Nixon, J.P.; Butterick, T.A. Role of orexin A signaling in dietary palmitic acid-activated microglial cells. Neurosci. Lett. 2015, 606, 140–144. [Google Scholar] [CrossRef]
- Messal, N.; Fernandez, N.; Dayot, S.; Gratio, V.; Nicole, P.; Prochasson, C.; Chantret, I.; LeGuilloux, G.; Jarry, A.; Couvelard, A.; et al. Ectopic expression of OX1R in ulcerative colitis mediates anti-inflammatory effect of orexin-A. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3618–3628. [Google Scholar] [CrossRef]
- Fatemi, I.; Shamsizadeh, A.; Ayoobi, F.; Taghipour, Z.; Sanati, M.H.; Roohbakhsh, A.; Motevalian, M. Role of orexin-A in experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2016, 291, 101–109. [Google Scholar] [CrossRef]
- Kim, M.-K.; Park, H.-J.; Kim, S.-R.; Choi, Y.K.; Bae, S.-K.; Bae, M.-K. Involvement of Heme Oxygenase-1 in Orexin-A-induced Angiogenesis in Vascular Endothelial Cells. Korean J. Physiol. Pharm. Off. J. Korean Physiol. Soc. Korean Soc. Pharmacol. 2015, 19, 327–334. [Google Scholar] [CrossRef]
- Tomasik, P.J.; Sztefko, K. The effect of enteral and parenteral feeding on secretion of orexigenic peptides in infants. BMC Gastroenterol. 2009, 9, 92. [Google Scholar] [CrossRef] [PubMed]
- Almeneessier, A.S.; Alzoghaibi, M.; BaHammam, A.A.; Ibrahim, M.G.; Olaish, A.H.; Nashwan, S.Z.; BaHammam, A.S. The effects of diurnal intermittent fasting on the wake-promoting neurotransmitter orexin-A. Ann. Thorac. Med. 2018, 13, 48–54. [Google Scholar] [PubMed]
- Yousefi, B.; Faghfoori, Z.; Samadi, N.; Karami, H.; Ahmadi, Y.; Badalzadeh, R.; Shafiei-Irannejad, V.; Majidinia, M.; Ghavimi, H.; Jabbarpour, M. The effects of Ramadan fasting on endothelial function in patients with cardiovascular diseases. Eur. J. Clin. Nutr. 2014, 68, 835–839. [Google Scholar] [CrossRef] [PubMed]
- Nasiri-Ansari, N.; Dimitriadis, G.K.; Agrogiannis, G.; Perrea, D.; Kostakis, L.D.; Kaltsas, G.; Papavassiliou, A.G.; Randeva, H.S.; Kassi, E. Canagliflozin attenuates the progression of atherosclerosis and inflammation process in APOE knockout mice. Cardiovasc. Diabetol. 2018, 17, 106. [Google Scholar] [CrossRef]
- Kassi, E.; Nasiri-Ansari, N.; Spilioti, E.; Kalotychou, V.; Apostolou, P.E.; Moutsatsou, P.; Papavassiliou, A.G. Vitamin D interferes with glucocorticoid responsiveness in human peripheral blood mononuclear target cells. Cell. Mol. Life Sci. 2016, 73, 4341–4354. [Google Scholar] [CrossRef]
- Zhang, H.; Liang, B.; Li, T.; Zhou, Y.; Shang, D.; Du, Z. Orexin A Suppresses Oxidized LDL Induced Endothelial Cell Inflammation via MAPK p38 and NF-kappaB Signaling Pathway. IUBMB Life 2018, 70, 961–968. [Google Scholar] [CrossRef]
- Lee, H.Y.; Lee, S.Y.; Kim, S.D.; Shim, J.W.; Kim, H.J.; Jung, Y.S.; Kwon, J.Y.; Baek, S.-H.; Chung, J.; Bae, Y.-S. Sphingosylphosphorylcholine stimulates CCL2 production from human umbilical vein endothelial cells. J. Immunol. 2011, 186, 4347–4353. [Google Scholar] [CrossRef]
- Nijboer, C.H.; Heijnen, C.J.; Groenendaal, F.; May, M.J.; van Bel, F.; Kavelaars, A. A dual role of the NF-kappaB pathway in neonatal hypoxic-ischemic brain damage. Stroke 2008, 39, 2578–2586. [Google Scholar] [CrossRef]
- Werle, M.; Schmal, U.; Hanna, K.; Kreuzer, J. MCP-1 induces activation of MAP-kinases ERK, JNK and p38 MAPK in human endothelial cells. Cardiovasc. Res. 2002, 56, 284–292. [Google Scholar] [CrossRef]
- Yang, C.-Q.; Li, W.; Li, S.Q.; Li, J.; Li, Y.W.; Kong, S.X.; Liu, R.-M.; Wang, S.-M.; Lv, W.M. MCP-1 stimulates MMP-9 expression via ERK 1/2 and p38 MAPK signaling pathways in human aortic smooth muscle cells. Cell. Physiol. Biochem. 2014, 34, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Guenzi, E.; Töpolt, K.; Lubeseder-Martellato, C.; Jörg, A.; Naschberger, E.; Benelli, R.; Albini, A.; StuÈrzl, M. The guanylate binding protein-1 GTPase controls the invasive and angiogenic capability of endothelial cells through inhibition of MMP-1 expression. EMBO J. 2003, 22, 3772–3782. [Google Scholar] [CrossRef] [PubMed]
- Singhal, A.; Wells, J.; Cole, T.J.; Fewtrell, M.; Lucas, A. Programming of lean body mass: A link between birth weight, obesity, and cardiovascular disease? Am. J. Clin. Nutr. 2003, 77, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, J.G.; Forsen, T.J.; Kajantie, E.; Osmond, C.; Barker, D.J.P. Childhood growth and hypertension in later life. Hypertension 2007, 49, 1415–1421. [Google Scholar] [CrossRef] [PubMed]
- Harder, T.; Bergmann, R.; Kallischnigg, G.; Plagemann, A. Duration of breastfeeding and risk of overweight: A meta-analysis. Am. J. Epidemiol. 2005, 162, 397–403. [Google Scholar] [CrossRef]
- Singhal, A.; Cole, T.J.; Fewtrell, M.; Lucas, A. Breastmilk feeding and lipoprotein profile in adolescents born preterm: Follow-up of a prospective randomised study. Lancet 2004, 363, 1571–1578. [Google Scholar] [CrossRef]
- Singhal, A.; Cole, T.J.; Lucas, A. Early nutrition in preterm infants and later blood pressure: Two cohorts after randomised trials. Lancet 2001, 357, 413–419. [Google Scholar] [CrossRef]
- Martin, R.M.; Ness, A.R.; Gunnell, D.; Emmett, P.; Smith, G.D. Does breast-feeding in infancy lower blood pressure in childhood? The Avon Longitudinal Study of Parents and Children (ALSPAC). Circulation 2004, 109, 1259–1266. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nasiri Ansari, N.; Spentza, F.; Dimitriadis, G.K.; Daskalopoulou, A.; Karapanagioti, A.; Siasos, G.; Lianidou, E.; Papavassiliou, A.G.; Kassi, E.; Randeva, H.S. Orexin-A Exerts Equivocal Role in Atherosclerosis Process Depending on the Duration of Exposure: In Vitro Study. Nutrients 2020, 12, 53. https://doi.org/10.3390/nu12010053
Nasiri Ansari N, Spentza F, Dimitriadis GK, Daskalopoulou A, Karapanagioti A, Siasos G, Lianidou E, Papavassiliou AG, Kassi E, Randeva HS. Orexin-A Exerts Equivocal Role in Atherosclerosis Process Depending on the Duration of Exposure: In Vitro Study. Nutrients. 2020; 12(1):53. https://doi.org/10.3390/nu12010053
Chicago/Turabian StyleNasiri Ansari, Narjes, Flora Spentza, Georgios K. Dimitriadis, Aphrodite Daskalopoulou, Angeliki Karapanagioti, Gerasimos Siasos, Evi Lianidou, Athanasios G. Papavassiliou, Eva Kassi, and Harpal S. Randeva. 2020. "Orexin-A Exerts Equivocal Role in Atherosclerosis Process Depending on the Duration of Exposure: In Vitro Study" Nutrients 12, no. 1: 53. https://doi.org/10.3390/nu12010053
APA StyleNasiri Ansari, N., Spentza, F., Dimitriadis, G. K., Daskalopoulou, A., Karapanagioti, A., Siasos, G., Lianidou, E., Papavassiliou, A. G., Kassi, E., & Randeva, H. S. (2020). Orexin-A Exerts Equivocal Role in Atherosclerosis Process Depending on the Duration of Exposure: In Vitro Study. Nutrients, 12(1), 53. https://doi.org/10.3390/nu12010053